
A subscription to JoVE is required to view this content. Sign in or start your free trial.
Method Article
Dissection ו-הרכבה שטוחה של שלד Threespine קוצן branchial
In This Article
Summary
The branchial skeleton, including gill rakers, pharyngeal teeth, and branchial bones, serves as the primary site of food processing in most fish. Here we describe a protocol to dissect and flat-mount this internal skeleton in threespine sticklebacks. This method is also applicable to a variety of other fish species.
Abstract
The posterior pharyngeal segments of the vertebrate head give rise to the branchial skeleton, the primary site of food processing in fish. The morphology of the fish branchial skeleton is matched to a species' diet. Threespine stickleback fish (Gasterosteus aculeatus) have emerged as a model system to study the genetic and developmental basis of evolved differences in a variety of traits. Marine populations of sticklebacks have repeatedly colonized countless new freshwater lakes and creeks. Adaptation to the new diet in these freshwater environments likely underlies a series of craniofacial changes that have evolved repeatedly in independently derived freshwater populations. These include three major patterning changes to the branchial skeleton: reductions in the number and length of gill raker bones, increases in pharyngeal tooth number, and increased branchial bone lengths. Here we describe a detailed protocol to dissect and flat-mount the internal branchial skeleton in threespine stickleback fish. Dissection of the entire three-dimensional branchial skeleton and mounting it flat into a largely two-dimensional prep allows for the easy visualization and quantification of branchial skeleton morphology. This dissection method is inexpensive, fast, relatively easy, and applicable to a wide variety of fish species. In sticklebacks, this efficient method allows the quantification of skeletal morphology in genetic crosses to map genomic regions controlling craniofacial patterning.
Introduction
סכום מדהים של מגוון קיים בשלד הראש בין חוליות, במיוחד בקרב דגים. במקרים רבים המגוון הזה הופך לפשוט אסטרטגיות האכלה שונות 1 - 4, והוא יכול לכלול שינויים משמעותיים הוא דפוסי craniofacial חיצוניים ופנימיים. שלד branchial ממוקם באופן פנימי בגרון של דג והמקיף ביותר של חלל buccal. שלד branchial מורכב 5 מקטעים הומולוגיים סדר, הקדמיים וארבעת מהם לתמוך הזימים. יחד חמישה קטעים אלה כממשק בין דגים ופירות שלהם 5. וריאצית שפע של תכונות כולל rakers זימים, שיני בלוע, ועצמות branchial לתרום ליקוט יעיל על סוגים שונים של מזון.
הקוצנים עברו קרינה אדפטיבית לאחר צורות אושיאניק האבות יישבו אגמי מים מתוקים ונחלים ברחבי חצי הכדור הצפוני. המעבר בתזונהמ זואופלנקטון קטן באוקיינוס כדי טרף גדול במים מתוקים הביא וריאציה trophic דרמטית תכונות כמה craniofacial 6. בעוד מחקרים רבים התמקדו על הבדלי craniofacial חיצוניים הקוצנים 7 - 13, שינויי craniofacial חשובים להתפתח שוב ושוב בשלד branchial הפנימי. היכולת ליצור כלאיים פורים בין אוכלוסיות קוצן ברורות מורפולוגית מספקת הזדמנות מצוינת כדי למפות את הבסיס הגנטי של שינויים התפתחו עד שלד branchial.
תכונה אחת trophic של משמעות אקולוגית היא הדפוסים של rakers זימים, עצמות עורי תקופתיות המרפדות את פניהם הקדמיים וגם אחוריים של עצמות branchial ומשמשים לסנן פריטים טרפים. דגים כי בדרך כלל ניזונים טרף קטנים נוטים להיות יותר ויותר בצפיפות הזימים במרווחים rakers לעומת דגים הניזונים טרף גדול יותר 14,15. וריאציה rakers הזימים דווחה הן within ובין מיני 14-19, והיבטים של דפוסי מגרפות זימים לתרום נישות trophic וכושר 16. עשרות שנים של מחקר תיעדו בהרחבה מספר זיז הזים וריאציה בהרחבה הקוצנים threespine 17 - 21; עם זאת, מחקרים אלה מתמקדים בדרך כלל בשורה הראשונה של rakers זימים. מחקר שנערך לאחרונה הוכיח מודולריות בבקרה הגנטית של מספר מגרפות זימים פני שלד branchial 22,23 ועל פני שורה אחת מגרפות זימים ריווח 23 ואורך 24 המדגיש את החשיבות של לימוד יותר משורה אחת או מגרפות זימים אחת להבין את בסיס גנטי ההתפתחותי של הפחתת מגרפות זימים.
אחת מתכונותיהן trophic שניות של שניהם המשמעות האקולוגית ביו הן הדפוסים של שיניים בלועות. שיני דגים יכולות להיות ממוקמות בשני הלסת האוראלית בשלד branchial, המכונות שיני בלוע. שיני Oral משמשות בעיקר עמ 'rey ללכוד בשיניים בלוע משמשים מניפולציה לעיסה וטרף 25 - 27. שתי הסדרות יוצרים באמצעות מנגנונים התפתחותיים משותף ונחשבים מבחינה התפתחותית הומולוגיים 28. מודולריות מעניין מתרחש תוך שלחלק מינים, כגון דג זברה, שיני בלוע פה ועל גב חוסר 29 בעוד מינים אחרים ceratobranchials מספר שיניים, pharyngobranchials, ולפעמים שיני basihyal ו hypobranchials 30. בשנת הקוצנים, שיניים בלוע נמצאים ventrally על ceratobranchial ו dorsally החמישי על הקדמי וגם האחורי pharyngobranchials 31. קינמטיקה על האכלת קוצן להראות הלסת האוראלית משמשת בעיקר עבור לכידת טרף והקלת האכלת יניקת 9 עוזב לעיסה ללסת הבלוע. בשנת ציקלידים, מורפולוגיה הלסת בלוע התחתון משתנה באופן דרמטי 32,33 ו הוכח להיות אדפטיבית בקורלציה עם נישה trophic 34. רַבאוכלוסיות קוצן מים מתוקים ple התפתחו מגדילה דרמטית במספר שן בלוע גחון 23,35,36. מחקר שנערך לאחרונה הוכיח כי הבסיס הגנטי ההתפתחותי של רווח השן התפתח זה הנו ייחודי, בעיקר בשתי אוכלוסיות נגזרות עצמאי של הקוצנים מים מתוקים 36. בניגוד שיניים יונקים, דגים להתחדש שיניהם ברציפות לאורך החיים הבוגרים 37. שתי אוכלוסיות מים מתוקות שיניים גבוהות שתוארו לעיל אלה התפתחו בקצב החלפת שן מואץ, ובכך יוצרים מערכת החולייתנים נדירה לחקר הבסיס הגנטי של התחדשות 36.
אחת מתכונותיהן trophic שלישי התפתח שוב ושוב הקוצנים מים מתוקים ארוכה עצמות epibranchial ו ceratobranchial, את homologs קשת המגזרי branchial של הלסת העליונה והתחתונה, בהתאמה 38. עצמות branchial ארוכות להעניק חלל buccal גדול וסבירות הם אדפטיבית על שאפשרו פריטי טרף גדולים להיות גonsumed. יתר על כן, בדגים אחרים, עצמות epibranchial חשובות לדיכאון של צלחות שן בלוע הגבה 25. כמו זיז זים ושיני בלוע, עצמות branchial הן פנימיות ולכן, קשה לדמיין או לכמת בקלות.
כאן אנו מציגים פרוטוקול מפורט לנתח שטוח הר שלד branchial, המאפשרים הדמיה קלה וכימות של מגוון תכונות craniofacial חשובים. בעוד פרוטוקול זה מתאר לנתיחת קוצן, באותה שיטה זו פועלת על מגוון רחב של דגים אחרים.
Access restricted. Please log in or start a trial to view this content.
Protocol
כל העבודה דגים אושרה על ידי ועדת טיפול בבעלי חיים מוסדיים השתמש מאוניברסיטת קליפורניה-ברקלי (מספר פרוטוקול R330). המתת חסד בוצעה באמצעות טבילה 0.025% Tricaine-S שנאגרו עם 0.1% נתרן ביקרבונט 39. כל השלבים מבוצעים בטמפרטורת החדר.
1. הכנה
הערה: בצע את פעולות 1.1-1.5 צינורות חרוטים או צלוחיות נצנץ שיכול לאטום בחוזקה להיות מונח אופקי. דגים לא צריכים להיות מזועזע כל הזמן, אבל מנסה לערבב הפתרון לעתים קרובות ככל האפשר על ידי היפוך בעדינות או טלטול מתלה של צינורות או צלוחיות לחשוף את כל הצדדים של הדג לפתרון מכתים ולאפשר כתם לחדור לרקמה שווה. אל תניחו קבוצה גדולה של דגים על שייקר פלטפורמה, כמו המשקל הכבד של הנוזל ישבור שייקר.
- תקן או דג טרי מורדם או דגים מאוחסן אתנול עם 10% Neutral שנאגר פורמלין (NBF) הלילה. לחלופין, להשתמש 4% paraformאלדהיד בתמיסה 1x PBS במקום 10% NBF.
הערה: אם להפיק ממנה DNA, קליפ חלק קטן של סנפירי זנב או חזה לפני הקיבוע ולאחסן באתנול. - השלך לתקן כראוי במנדף כימי ולהחליף עם מי ברז (כלומר ~ pH 7.0) עבור 2 שעות. הימנע משימוש במי דה מיונן כפי שקרה לעתים קרובות יכול להיות חומצי יכול decalcify עצם.
- סר מים ודגי כתם עם 0.008% Alizarin האדום S ב 1% KOH במים למשך 24 שעות. עבור דגים פחות מ -20 מ"מ אורך סטנדרטי, השתמש 0.004% Alizarin האדום ס (ערוך 100x (פתרון מניות 0.8%) של Alizarin האדום S אז מה שיכול להיות מדולל).
- הסר כתם (הצבת מיכל פסולת המתאים במנדף) ודגים המקום במי ברז למשך מספר שעות. שינוי מים לפי הצורך עד לשטוף במים הוא בעיקר ברור.
- סור דגי מים ומקום לתוך 50% גליצרול, 0.25% KOH לסליקה עדינה לנתיחה שלאחר מכן.
הערה: פרוטוקול מכתים זה שונה מן השיטות שתוארו קודם לכן 40,41.
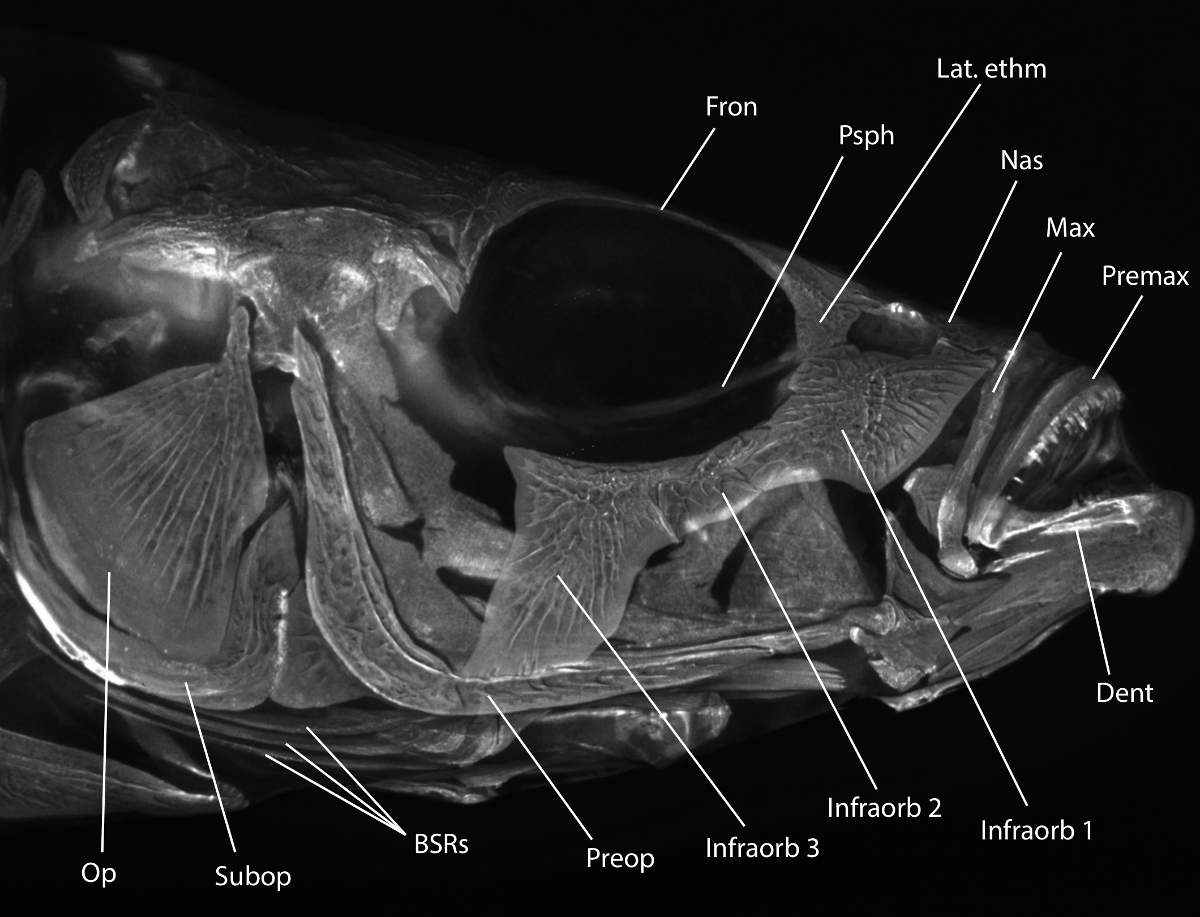
הערה: ראה איור 1 עבור סקירה של מורפולוגיה שלד ראש רלוונטי.

איור 1:. קוצן ראש מורפולוגיה שלד Alizarin האדום מוכתם ראש קוצן threespine צלם עם קרינה תחת סט מסנן rhodamine B. מורפולוגיה שימושיים מתויג: אופ = opercle, Subop = subopercle, BSRs = branchiostegal קרני, Preop = preopercle, Infraorb 1-3 = 1-3 infraorbital (circumorbitals המכונה גם או suborbitals), דנט = dentary, Premax = premaxilla, מקס = maxilla נאס, = אף, לאט. ethm = לרוחב כִּברָתִי, Psph = parasphenoid, Fron = העצם הפרונטלית. לתיאור מפורט יותר של השלד ראש קוצן, לראות אנקר (1974) 31. אנא לחץ כאן כדי להציג גרסה גדולה יותר שלנתון זה.
{kind=link}
- הנח את הדג השטוח (איור 2 א) וכנס המלקחיים של השען # 5 חד לתוך הצד של העין בזווית של 45 מעלות ~ לנקב את הקרום המכסה את העין.
- מקלף את הקרום הרחק מן העין, דומה לקליפת מכסת יוגורט (איור 2 ב).
- הכנס מלקחיים פתוחים מאחורי העין, לאחוז את עצב הראייה מאחורי העין, ולהסיר את העין (איור 2 ג). אין לנקב את העין כפי שהוא ידלוף מלנין. אם ניקב, מלנין ניתן נשטף במהלך השלבים מאוחר יותר.
- חזור על צד שני.
- החל האחורי, למקום להב מספריים לנתח קטן אחד תחת כנף opercle, להב מספריים גרור dorsally מעל opercle, ואז לחתוך רקמות רכות ועד ארובת העין (איור 2 ד). חותכים הגבו עד עצם opercle.
- חותך את העצם הפרונטלית (גב אל ארובת העין) (איור 2E).
- חותכים את העצם parasphenoid קו האמצע סביב tהוא המרכז של ארובת העין (איור 2F).
- לחתוך opercle חוזר על הצד שכנגד.
- הכנס מלקחיים תחת opercle ולאט לאט לקלף את הפנים מהגוף, זמירה כל רקמות רכות עדיין מחוברות (2G איור - H). שים לב שלא לשבש את השורה הראשונה של rakers זימים.
- עם מלקחיים, לנתק את ceratohyals משני הצדדים מן basihyal קו האמצע תוך קילוף והסרה של שלד craniofacial קדמית (הלסת כולה כולל dentary, premaxilla, ו maxilla; השלד hyoid כולו כולל opercle עורי החיצוני, preopercle, subopercle, ו קרן branchiostegal ואת הגב שבבסיס ואלמנטי endochondral גחון; והחלק הקדמי של הגולגולת כולל האף, כִּברָתִי לרוחב, ועצמות infraorbital, ראה איורים 1 ו 2I).
- קוצי אגן יכולים להיות מקופלים החוצה מהגוף, ויכולים לשמש כידית עבור מלקחיים לאחוז of כאשר ההווה. שדרות ננעלו במקומו. כדי לבטל את הנעילה, משוך בעדינות את עמוד שדרה עם מלקחיים ישירות מגוף דגים, אז בעדינות לכופף בדיעבד ללחוץ עמוד שדרה שטוחה נגד דגים.
- הכניסו נסגר מלקחיים אחוריים גחון לשלד branchial (ממש מתחת צינור מעיים) ו מלקחיים גרורים קדמי, מתגרים לגזרים את השרירים ורצועות הנותרים המקובעים לשלד branchial (האיור 2J - K).
- באמצעות קצות מלקחיים סגורים, לגרד ממנו את שרירי הצמדת השלד הגבה branchial אל braincase הגחון בתוך אחורי לכיוון קדמי (האיור 2L).
- חזור על 2.9 ו 2.10 על הצד שכנגד.
- אחוז בבסיס צינור המעיים ולמשוך anteriorly להסיר את הצינור שלד ובמעיים branchial (איור 2M - N).
- הפרד את צינור המעיים על ידי יצירת האחורי חתך בניצב סוף ceratobranchial החמישי (איור 2 טו ).
- לאחר הסרת כל שברי עצמות שנותרו מן braincase בצד הגבי של שלד branchial, הכנס מספריים לתוך סל branchial לעשות חתך הגבה (חיתוך קדמי אל אחורי) בין הסטים הבילטרליים של צלחות שן הגבה (איור 3 א - ד). ודא לחתוך מרוכז כדי למנוע נזק לוחות השן הגבי.
- לעשות שני חתכים לרוחב רדודים לומן המעי גומי בקצה האחורי של השלד branchial (הקצה הקדמי של צינור המעיים) כדי לסייע עם פתיחת שלד branchial (איור 3E).
- הנח דגים וכל פיסות הרקמה לתוך צנצנת והניח שלד branchial לתוך צינור microcentrifuge עם גליצרול 50%, 0.25% KOH להמשיך סליקה עדינה, או גליצרול 100% אם לא סליקה נוספת נדרשת. צנצנות וצינורות תווית עם מזהה ייחודי, כך שניתן יהיה לעקוב. סכום סליקה הנדרש הוא בעיקר פונקציה של הגודל של הדגים, דגים בוגרים גדולים (מעל 40 מ 'מ 'אורך סטנדרטי) בדרך כלל לחייב סליקה נוספת.

איור 2:. לנתיחת שלד branchial קוצן Alizarin האדום מוכתם דגי קוצן threespine מוכנים לנתיחה. עין depigmented מ סליקה נרחבת. חצים כחולים מציינים את כיוון התנועה. (א) להציג לרוחב של ראש קוצן, הקדמי הוא בצד ימין. (ב) הסרת קרום המכסה את העין. (ג) הסרת העין. (ד) הגבה לחתוך מעל opercle. (E) לחתוך עצם פרונטאלית. (F) Parasphenoid לחתוך. (G - אני) הסרת שלד הפנים. (J) הסרת חיבורי רקמות רכות שלד branchial גחון. (K - L) הסרת חיבורי שלד הגבה branchial. ( M - N) הסרת שלד branchial. (O) הפרד את צינור המעיים מן השלד branchial. עיין בשלבים 2.1 דרך 2.16 לפרטים נוספים. סרגל קנה מידה = 5 מ"מ. אנא לחץ כאן כדי לצפות בגרסה גדולה יותר של דמות זו.
{kind=link}
3. שלד branchial מחדש מכתים (במידת הצורך)
- כדי להכתים את כהה שלד branchial או רקמות ברורות יותר, להסיר גליצרול 50%, פתרון KOH 0.25% ולשטוף עם 1% KOH פעמים (לשטוף אחד חמש דקות ואחריו לשטוף את שני 24 שעות תוך רעד אופקי על שייקר פלטפורמה).
- סר 1% KOH מחדש כתם עם 0.008% Alizarin האדום S ב KOH 1% למשך 24 שעות.
- הסר כתם ולהחליף עם KOH 1% למשך 24 שעות.
- הסר פתרון KOH ולהחליף עם 50% גליצרול, 0.25% KOH.
4. הרכבת branchial שלד
- הסר שלד branchialמ גליצרול 50%, 0.25% KOH או 100% גליצרול מקום קרוב לתחתית של תלוש כיסוי זכוכית 22 מ"מ x 60 מ"מ עם הצד הגבי פונה כלפי מעלה (איור 3F). הוסף כמה טיפות של גליצרול 50%, 0.25% KOH או 100% גליצרול על גבי שלד branchial. אם מעבר מ 50% גליצרול, 0.25% KOH ל -100% גליצרול, פתרון שינוי בצינור microcentrifuge ולנער עבור> 5 דקות לפני הרכבה לאזן רקמות.
- מרדדי שני כדורים קטנים של פלסטלינה ומניח על קצוות של להחליק את המכסה לפעול כמו מפרידים.
- רופף במקום coverslip שני על גבי בלחץ חזק מספיק כדי לשטח את שלד branchial הקדמי (האיור 3G).
- פיל לפתוח את דש הגבי עזב כולל צלחות שן מגבות, לרדד, וחלק בין coverslips (האיור 3H).
- טכניקה חזור עם דש הגבי תקין ולדחוף השלד כולו branchial מהקצה של coverslip (איור 3I).
- לשנותבאופן מקורי, להחזיק דשי הגבי של חלל פתוח עם מלקחיים ובזהירות מקום coverslip על, משטח את שלד branchial בתנועה אחת חלקה.
- לחלופין, הר השלד branchial הפוך על תלוש לכסות אחד, מפשק אותן למעלה ולמטה בכל צד הגב החוצה רוחבית כך כוח המשיכה אינו מאפשר השלד branchial לסגור לגבות. ואז לכסות עם תלוש כיסוי הזכוכית שני 22 מ"מ x 60 מ"מ ו להפוך הכנה.
הערה: טכניקות הרכבה שונות נוהגות לעבוד טוב יותר או גרוע יותר עבור כל אדם. נסה כל ולראות מה מרגיש הכי נוח.
- לחץ קל על coverslip העליון לשטח כדורי חימר מספיק כדי לשמור על שלד branchial הרכוב שטוח, אך היזהרו שלא למחוץ את הדגימה.
- במהלך תהליך ההרכבה, ceratobranchials יכול לסובב לטשטש שורה של rakers. תרופה זו על ידי הזזה מלקחיים בין coverslips מחדש המכוונת את ceratobranchials או שלד branchial כולו.
- חנות preps שטוחה במגשי שקופית roטמפרטורת אום. רכוב גליצרול 100%, ניתן לאחסן preps בין coverslips לגשר במשך עשור לפחות. מלקחיים ומספריים נקיים עם isopropanol או אתנול וטיפי כיסוי.

איור 3:. שטוח הרכבת שלד branchial המניפולציה ואת ההרכבה של שלד branchial מוצג. חצים כחולים מציינים את כיוון התנועה. (א) branchial הגבה שלד למעלה בצד. (B - D) סיבוב חתך בין לוחות שן הגבה. (E), קיצוץ רוחבי ברקמות רכות להמשיך לפתוח את הבסיס של צינור הבטן. (F) שלד branchial ממוקם בחלק התחתון של coverslip מוכן גובר. (G) coverslip השנייה מונחת על החצי הקדמי של שלד branchial (מעל צלחות שן מגבות). (H - I)הרכבה שטוחה של שלד branchial ידי דשי צלחת שן הגבה פתיחת זזה בין שני תלושים לכסות. עיין בשלבים 4.1 עד 4.6 לפרטים נוספים. סרגל קנה מידה = 5 מ"מ. אנא לחץ כאן כדי לצפות בגרסה גדולה יותר של דמות זו.
{kind=link}
Access restricted. Please log in or start a trial to view this content.
תוצאות
תוצאות פרוטוקול זה שלד branchial רכוב גזור ושטוח (איור 4) שבו מגוון של תכונות תזונתיות חשובות ניתן לכמת. מתוך תפיסת גב, כל השורות של rakers זימים, כל צלחות השן בלועה, וכמעט כל עצמות branchial ניתן מדמיינים בקלות לכמת 22 - 24,35,36,38,42. Alizarin ?...
Access restricted. Please log in or start a trial to view this content.
Discussion
The branchial skeleton is a complex set of bones in the throat of a fish that manipulates, filters, and masticates food items on their way to the esophagus. Many interesting trophic traits including the patterning of gill rakers, pharyngeal teeth, and branchial bones vary across and within species. The majority of these traits are difficult to near impossible to accurately measure with the branchial skeleton in situ (e.g., gill raker length, branchial bone length). This flat-mounting protocol places all...
Access restricted. Please log in or start a trial to view this content.
Disclosures
The authors have nothing to disclose.
Acknowledgements
This work was funded in part by NIH R01 #DE021475 to CTM and an NSF Graduate Research Fellowship to NAE. Thanks to Miles Johnson for assistance with imaging and Priscilla Erickson for critical reading of the manuscript.
Access restricted. Please log in or start a trial to view this content.
Materials
| Name | Company | Catalog Number | Comments |
| Potassium Hydroxide (KOH) | EMD | PX1480-1 | |
| Glycerol | Sigma-Aldrich | G7893-4L | |
| 10% Neutral Buffered Formalin (NBF) | Azer Scientific | NBF-4-G | |
| Alizarin Red S | EMD | AX0485-3 | |
| Microscope Cover Glasses 22 mm x 60 mm | VWR | 16004-350 | |
| 100 mm x 10 mm Glass Petri Dish | Kimble Chase | 23064-10010 | To dissect samples on |
| Sylgard 184 Silicone Elastomer Kit | Ellsworth Adhesives | 184 SIL ELAST KIT 0.5KG | Can be poured into glass or plastic Petri dishes to make dissecting plates |
| Modeling Clay | Sargent Art | 22-4000 | 1 lb cream |
| Scintillation Vials (case of 500) | Wheaton | 986586 | Borosilicate Glass with Screw Cap |
| Forceps-Dumont #5 Inox (Biologie tip) | FST | 11252-20 | Dumostars are an alternative |
| Dissecting Scissors | FST | 15003-08 | Alternate sizes are available depending on size of sample |
| Dissecting Microscope | Leica | S6E with KL300 LED | Many other models work nicely, having a flat base helps |
| Microcentrifuge Tubes 1.7 ml | Denville | C2170 | |
| Cardboard slide tray | Fisher | 12-587-10 |
References
- Cooper, W. J., Westneat, M. W. Form and function of damselfish skulls: rapid and repeated evolution into a limited number of trophic niches. BMC Evol. Biol. 9 (24), (2009).
- Albertson, R. C., Kocher, T. D. Genetic and developmental basis of cichlid trophic diversity. Heredity. 97 (3), 211-221 (2006).
- Martin, C. H., Wainwright, P. C. Trophic novelty is linked to exceptional rates of morphological diversification in two adaptive radiations of cyprinodon pupfish. Evolution. 65 (8), 2197-2212 (2011).
- Wainwright, P. C., et al. The evolution of pharyngognathy: A phylogenetic and functional appraisal of the pharyngeal jaw key innovation in labroid fishes and beyond. Syst. Biol. 61 (6), 1001-1027 (2012).
- Sibbing, F. Food capture and oral processing. Cyprinid Fishes. , 377-412 (1991).
- Bell, M., Foster, S. The Evolutionary Biology of the Threespine Stickleback. , Oxford University Press. New York. (1994).
- Kimmel, C. B., et al. Evolution and development of facial bone morphology in threespine sticklebacks. Proc. Natl. Acad. Sci. U. S. A. 102 (16), 5791-5796 (2005).
- Mcgee, M. D., Wainwright, P. C. Convergent evolution as a generator of phenotypic diversity in threespine stickleback. Evolution. 67 (4), 1204-1208 (2013).
- McGee, M. D., Schluter, D., Wainwright, P. C. Functional basis of ecological divergence in sympatric stickleback. BMC Evol. Biol. 13, 277(2013).
- McGuigan, K., Nishimura, N., Currey, M., Hurwit, D., Cresko, W. A. Quantitative genetic variation in static allometry in the threespine stickleback. Integr. Comp. Biol. 50 (6), 1067-1080 (2010).
- Caldecutt, W. J., Bell, M. A., Buckland-Nicks, J. A. Sexual dimorphism and geographic variation in dentition of threespine stickleback, Gasterosteus aculeatus. Copeia. 2001 (4), 936-944 (2001).
- Berner, D., Moser, D., Roesti, M., Buescher, H., Salzburger, W. Genetic architecture of skeletal evolution in european lake and stream stickleback. Evolution. 68 (6), 1792-1805 (2014).
- Jamniczky, H. a, Barry, T. N., Rogers, S. M. Eco-evo-devo in the study of adaptive divergence: examples from threespine stickleback (Gasterosteus aculeatus). Integr. Comp. Biol. 55 (1), 166-178 (2015).
- Magnuson, J., Heitz, J. Gill raker apparatus and food selectivity among mackerels, tunas, and dolphins. Fish. Bull. 69 (2), 361-370 (1971).
- Kahilainen, K. K., et al. The role of gill raker number variability in adaptive radiation of coregonid fish. Evol. Ecol. 25 (3), 573-588 (2011).
- Arnegard, M. E., et al. Genetics of ecological divergence during speciation. Nature. 511 (7509), 307-311 (2014).
- Gross, H. P., Anderson, J. M., Gross, H. P., Anderson, J. Geographic variation in the gillrakers and diet of European threespine sticklebacks, Gasterosteus aculeatus. Copeia. 1984 (1), 87-97 (1984).
- Hagen, D., Gilbertson, L. Geographic variation and environmental selection in Gasterosteus aculeatus L in the Pacific Northwest, America. Evolution. 26 (1), 32-51 (1972).
- McPhail, J. D. Ecology and evolution of sympatric sticklebacks (Gasterosteus): morphological and genetic evidence for a species pair in Enos Lake, British Columbia. Can. J. Zool. 62 (7), 1402-1408 (1984).
- Schluter, D., McPhail, J. D. Ecological character displacement and speciation in sticklebacks. Am. Nat. 140 (1), 85-108 (1992).
- Robinson, B. Trade offs in Habitat-specific foraging efficiency and the nascent adaptive divergence of sticklebacks in lakes. Behaviour. 137 (7), 865-888 (2000).
- Glazer, A. M., Cleves, P. A., Erickson, P. A., Lam, A. Y., Miller, C. T. Parallel developmental genetic features underlie stickleback gill raker evolution. Evodevo. 5 (1), (2014).
- Miller, C. T., Glazer, A. M., et al. Modular skeletal evolution in sticklebacks is controlled by additive and clustered quantitative trait loci. Genetics. 197 (1), 405-420 (2014).
- Glazer, A. M., Killingbeck, E. E., Mitros, T., Rokhsar, D. S., Miller, C. T. Genome assembly improvement and mapping convergently evolved skeletal traits in sticklebacks with Genotyping-by-Sequencing. G3. 5, 1463-1472 (2015).
- Wainwright, P. Functional morphology of the pharyngeal jaw apparatus. Fish Physiol. Fish Biomech. , 77-102 (2006).
- Hulsey, C. D., Fraser, G. J., Streelman, J. T. Evolution and development of complex biomechanical systems: 300 million years of fish jaws. Zebrafish. 2 (4), 243-257 (2005).
- Lauder, G. Functional design and evolution of the pharyngeal jaw apparatus in euteleostean fishes. Zool. J. Linn. Soc. 77, 1-38 (1983).
- Fraser, G. J., et al. An ancient gene network is co-opted for teeth on old and new jaws. PLoS Biol. 7 (2), e1000031(2009).
- Stock, D. Zebrafish dentition in comparative context. J. Exp. Zool. B. Mol. Dev. Evol. 308, 523-549 (2007).
- Liem, K., Greenwood, P. A functional approach to the phylogeny of the pharyngognath teleosts. Am. Zool. 21 (1), 83-101 (1981).
- Anker, G. C. Morphology and kinetics of the head of the stickleback, Gasterosteus aculeatus. Trans. Zool. Soc. London. 32 (5), 311-416 (1974).
- Meyer, A. Morphometrics and allometry in the trophically polymorphic cichlid fish, Cichlusomu citrinelfum: Alternative adaptations and ontogenetic changes in shape. J. Zool., Lond. 221, 237-260 (1990).
- Huysseune, A. Phenotypic plasticity in the lower pharyngeal jaw dentition of Astatoreochromis alluaudi (Teleostei: Cichlidae). Arch. Oral Biol. 40 (11), 1005-1014 (1995).
- Muschick, M., Indermaur, A., Salzburger, W. Convergent Evolution within an adaptive radiation of cichlid fishes. Curr. Biol. 22 (24), 2362-2368 (2012).
- Cleves, P. A., et al. Evolved tooth gain in sticklebacks is associated with a cis-regulatory allele of Bmp6. Proc. Natl. Acad. Sci. 111 (38), 13912-13917 (2014).
- Ellis, N. A., et al. Distinct developmental and genetic mechanisms underlie convergently evolved tooth gain in sticklebacks. Development. (142), 2442-2451 (2015).
- Tucker, A. S., Fraser, G. J. Evolution and developmental diversity of tooth regeneration. Semin. Cell Dev. Biol. 25-26, 71-80 (2014).
- Erickson, P. A., Glazer, A. M., Cleves, P. A., Smith, A. S., Miller, C. T. Two developmentally temporal quantitative trait loci underlie convergent evolution of increased branchial bone length in sticklebacks. Proc. R. Soc. B. 281, (2014).
- Leary, S., et al. AVMA Guidelines for the Euthanasia of Animals. , American Veterinary Medical Association. Schaumburg, IL. (2013).
- Bell, M. A. Evolutionary phenetics and genetics. Evol. Genet. Fishes. , 431-528 (1984).
- Taylor, W. R., Van Dyke, G. C. Revised procedures for staining and clearing small fishes and other vertebrates for bone and cartilage study. Cybium. 9 (2), 107-119 (1985).
- Erickson, P. A., et al. A 190 base pair, TGF-β responsive tooth and fin enhancer is required for stickleback Bmp6 expression. Dev. Biol. 401 (2), 310-323 (2015).
- Miller, C. T., et al. cis-Regulatory changes in Kit ligand expression and parallel evolution of pigmentation in sticklebacks and humans. Cell. 131 (6), 1179-1189 (2007).
- Aigler, S. R., Jandzik, D., Hatta, K., Uesugi, K., Stock, D. W. Selection and constraint underlie irreversibility of tooth loss in cypriniform fishes. Proc. Natl. Acad. Sci. U. S. A. 111 (21), 7707-7712 (2014).
- Pasco-Viel, E., et al. Evolutionary trends of the pharyngeal dentition in Cypriniformes (Actinopterygii Ostariophysi). PLoS One. 5 (6), e11293(2010).
Access restricted. Please log in or start a trial to view this content.
Reprints and Permissions
Request permission to reuse the text or figures of this JoVE article
Request PermissionExplore More Articles
This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. All rights reserved