
Method Article
טיהור של H3, H4 חלבוני היסטון, כימות של סימני היסטון Acetylated תאים ורקמות המוח
In This Article
Summary
מטרת מאמר זה היא לספק מדריך מקיף, שיטתית של טיהור יעיל של שינויים היסטוניים H3 ו- H4, כימות של שאריות היסטון acetylated.
Abstract
בכל היצורים האיקריוטים, כרומטין, התבנית פיזיולוגיים של מידע גנטי הכל, חיוני תורשה. כרומטין כפוף מערך של שינויים posttranslational מגוונות (PTMs) בעיקר מתרחשים טרמיני אמינו של חלבוני היסטון (קרי, היסטון הזנב), לווסת את הנגישות ואת למצב התפקודי של ה-DNA המשמשת כבסיס. היסטון זנבות להאריך מן הליבה של נוקלאוזום וכפופות התוספת של קבוצות אצטיל מאת היסטון acetyltransferases (כובעים), הסרת קבוצות אצטיל מאת היסטון deacetylases (HDACs) במהלך צמיחה סלולרי ובידול. דפוסי acetylation ספציפית על שאריות ליזין (K) על זנבות היסטון לקבוע הומאוסטזיס דינמי בין כרומטין transcriptionally פעיל או מודחקים transcriptionally מכלול היסטון הליבה השפעה (1) ו- (2) גיוס סינרגטי או אויבת הקשורים כרומטין חלבונים אל אתר שעתוק. מנגנון רגולטורי הבסיסית של האופי המורכב של היסטון הזנב PTMs משפיע על רוב תבניות כרומטין תהליכים ותוצאות השינויים ב ההבשלה תא ובידול בפיתוח נורמליים ופתולוגיים. מטרת הדו ח הנוכחי נועד לספק טירונים עם שיטה יעילה לטהר את חלבוני היסטון הליבה של תאים ורקמות המוח, באופן אמין לכמת סימני acetylation על שינויים היסטוניים H3 ו- H4.
Introduction
אפיגנטיקה המונח מתייחס heritable שינויים בפעילות הגן המתרחשות באופן עצמאי מפני שינויים ב- DNA רצף1,2. שעתוק גנים ודיכוי נקבעים על ידי (1) את הנגישות של ה-DNA כרומוזומלית העוטפת את octamer של חלבונים היסטון הליבה (2 עותקים כל אחד H2A, ויזת עבודה H2B, H3 ו- H4) ועל ידי (2) הזמינות של גורמי שעתוק, לגרדום חלבונים גייס יזם ספציפיים לאתרים3,4. שעתוק גנים מוסדר על ידי שינויים בתיווך האנזים DNA מקדם אתרים ספציפיים, את PTMs של היסטון זנבות5,6,7. N-טרמיני של היסטון H3 ו- H4 הם בין הרצף השמור ביותר הידוע היצורים האיקריוטים3, את השינויים posttranslational תועדו באופן נרחב כדי לשחק תפקיד מרכזי בקביעת הכרומטין, הפונקציה8,9. PTMs על זנבות היסטון (קרי, acetylation, מתילציה, זרחון ו ubiquitination) לשנות את הפוטנציאל לאינטראקציה של הזנבות, להשפיע על המדינה מבניים, קיפול של כרומטין סיבים, ועל ידי כך, לווסת את הדנ א נגישות עיבוד4,10,11,12. קבוצות אצטיל הם להוסיף, להסיר שאריות K בהיסטון זנבות על ידי מערכת של אינטראקציה היסטון epigenetic אנזימים מסוימים, כלומר כובעים ו- HDACs, בהתאמה13. לדוגמה, acetylation של היסטון H4-ליזין 12 (H4K12ac) הוכח בעבר כדי להפעיל שעתוק של גנים הקשורים זיכרון הרכישה ואיחוד14. בנוסף, מספר קווי ראיה מראים כי הפקד epigenetic בתיווך האנזים של שעתוק גנים הוא היבט חיוני של בריאה הסלולר בהתפתחותם6,15. ההתחלפות epigenetic ברגולציה של ביטוי גנים, על-ידי שינויים epigenetic של ה-DNA או מוטציה של אנזימים epigenetic עצמם, הוכח להיות dysregulated של מחלות האדם שבו שינוי בפעילות גן מסוים מהווה סימן היכר מתוך פתולוגיה (למשל, סרטן)6,16,17. לפיכך, הערכת שינויים הליבה היסטון PTMs מתגלה כיעד ערך גבוה עבור פוטנציאליים התערבויות טיפוליות. עם זאת, קביעת השפע, אינטראקציה של שותפים, והתפקידים הספציפיים של שינויים היסטוניים PTMs הוכח מאתגר18.
בדו ח הנוכחי, מתואר אסטרטגיית ממוטבת, תפוקה בינונית לטהר שינויים היסטוניים הליבה של תאים ורקמות המוח שבר יחיד, וכן פרוטוקול מלא עבור כימות של שינויים היסטוניים H3 ו- H4 PTMs. ראוי לציין, למרות כיום לאור מבוססי חומצה טיהור טכניקות ואסטרטגיות לאיתור נוגדן מבוססי היסטון נרחב שאומצו על אפיון היסטון, הם חסרים פרטים תיאורי על שלבים קריטיים של ההליך, ובכך הפרעה חילוץ מהיר ו לטבלה הניתנת לשכפול היסטון, כימות. לדוגמה, לחלץ העיבוד של התא, ביופסיות רקמות דורשת כלים שונים וטכנולוגיות עבור חילוץ מוצלח. יתר על כן, הפרוטוקול ממוטבת שהוצגו בכתב היד הנוכחי מדגים גישה מעשית תפוקה בינונית. שינויים הליבה היסטוניים מחולצים כשבר יחידה, טהור, אשר מאפשרת אמין במורד הזרם בתיווך נוגדנים PTM זיהוי ללא כל הפרעה מפני זיהומים. יתר על כן, בכתב היד הנוכחי, האתגרים לגבי זיהוי היסטון עקב משקל מולקולרי קטן שלהם יש כבר עקפו. בדרך כלל, חוסר תאימות בין טיהור, כימות ג'ל אלקטרופורזה פרוטוקולים לעכב מדענים מ קבלת התוצאות לטבלה הניתנת לשכפול ולא חד משמעי. . הנה, זרימת עבודה ממוטבת כדי לטהר שינויים היסטוניים הליבה של תאים ורקמות ולהכין אותן במורד הזרם PTM ניתוחים דרך תספיג מוצג.
בפרוטוקול הנוכחי מאפשר הטיהור של חלבוני היסטון הליבה תוך שמירה על שלהם מקורית שינויים posttranslational (קריacetylation, מתילציה, זרחון). איור 1 מציג ציר הזמן של פרוטוקול טיהור היסטון.
Protocol
כל העכברים שוכנו במתקן לחות, טמפרטורה-מבוקר, מוכר AAALAC בעלי חיים בבית הספר מילר אוניברסיטת מיאמי של הרפואה. כל הניסויים היו שאושרו על ידי אוניברסיטת מיאמי מילר בבית הספר של רפואה מוסדיים חיה טיפול ועל שימוש הוועדה (IACUC), שנערך על פי מפרט של NIH.
1. הכנת תמצית מדגם
-
תאים חסיד
- צלחת תאים במנות 10 ס מ בתקשורת תרבות המתאים (1 x 106 עונה 1 פרק 109 תאים לכל מאכל עבור שורות תאים, כגון BV2, HEK-293, ו- SH-SY5Y, אבל ~ 1 x 10 תאים15 לכל מאכל עבור תאים הראשי, כגון ראשי נוירונים בקליפת המוח). מבטיח כי התאים מופצים בצורה שווה על פני כל צלחת וגם מאפשרים לתאים לגדול במשך 48 שעות להגיע למפגש ~ 100% (37 מעלות צלזיוס, 5% CO2).
- ברגע התאים מיצית את confluency הרצויה, בעדינות תשאף התקשורת תרבות ולשטוף את התאים 2 x עם prewarmed מדיה ללא סרום ברדס תרביות רקמה.
- האחות התקשורת ללא סרום מתוך קערה ומוסיפים 1 מ"ל המאגר הקר החילוץ (0.4 M חומצה גופרתית, 1 מ מ. אשלגן כלורי, 1 מ MgCl2, 50 מ מ טריס-HCl [pH 8.0] 1 x מעכב פרוטאז קוקטייל) על כל מנה.
- להשתמש במגרדת תא פלסטיק כדי לאסוף את כל התאים למאגר החילוץ (על-ידי גירוד) ומעבירים אותם אל mL 1.5 הנקרא צינור עם פיפטה 1,000 µL. Pipet התאים למעלה ולמטה 3 x כדי להקל על המגון.
- סגור את כל צינורות, מיד לשים אותם על קרח.
-
רקמת המוח
- אם נעשה שימוש רקמות קפוא, למקם את רקמת צינור prechilled 1.5 mL, בקצרה להפשיר את זה על קרח. אם נעשה שימוש ברקמות טריים, להמשיך מיד לשלב 1.2.2.
הערה: הפרוטוקול הנוכחי מתאר הליכים באמצעות מוח העכבר קפוא ודוגמאות קליפת המוח הקדם חזיתית העכבר. - Homogenize הרקמה באמצעות מהמגן דאונס כף-יד באמצעות את הכמות המתאימה של מיצוי מאגר ואת המספר המומלץ של קווים (טבלה 1). כדי למנוע גיזום כרומטין מוגזמת, לא יעלה על מספר קווים המומלצת.
- באמצעות פיפטה µL של ערוץ אחד 1,000, להעביר homogenate את צינור prechilled 1.5 mL. סגור את כל צינורות, מיד לשים אותם על קרח.
- אם נעשה שימוש רקמות קפוא, למקם את רקמת צינור prechilled 1.5 mL, בקצרה להפשיר את זה על קרח. אם נעשה שימוש ברקמות טריים, להמשיך מיד לשלב 1.2.2.
2. הכנת תמצית היסטון גולמי
- מקום הצינורות 1.5 mL המכילה את תאים או רקמות מושעה במאגר החילוץ על פלטפורמה מסתובבת וסובב ב rpm 15 ב 4 ° C כדי לאפשר הפקת שינויים היסטוניים גסה.
הערה: הזמן החילוץ עשויים להשתנות עבור תאים שונים וסוגי רקמות, חייב להיות מותאם לכל הליך. בפרוטוקול הנוכחי מציג התוצאות המתקבלות לאחר 15 דקות, 2 h ו- 24 שעות של מיצוי (איור 2, 3 איור, איור 4ו איור 5). - Prechill את microcentrifuge עד 4 ° C. לאחר העקירה הרצויה זמן עבר, centrifuge הצינורות במהירות מקסימלית 10 דקות ב 4 º C.
- להעביר את תגובת שיקוע כולל את שינויים היסטוניים גולמי לרכבת התחתית mL 1.5 חדש, prechilled. להתעלם בגדר.
- לאחסן את תגובת שיקוע ב-80 מעלות צלזיוס (ניתן לעצור את החילוץ בשלב זה, ראה "עצור צעד" באיור1) או להמשיך לשלב הבא.
- לנטרל את שינויים היסטוניים גולמי עם נפח 1/4 של 5 x ניטרול מאגר (למשללהוסיף 250 µL של 5 x ניטרול מאגר 1 מ"ל של שינויים היסטוניים גולמי). מערבבים היטב על ידי pipetting למעלה ולמטה 6 x.
- בדוק את ה-pH של התערובת עם חומציות. משתנות בהתאם על ידי הוספת עוד מאגר ניטרול להגיע pH של 7.
- להעריך את הנוכחות של היסטון ולחלץ חלבון nonhistone היסטון גולמי כדלקמן (איור 2).
- להוסיף µL 37.5 מדגם µL 12.5 של 4 x (Laemmli) דוגמת המאגר, denature 10 דקות ב 99 º C.
- לטעון את הדגימה אל ג'ל מרחביות-דף ולהפעיל את הג'ל מאובטח ב- 100 וולט.
- מכתים את הג'ל במשך הלילה עם פתרון מוכתמים R-250 כחול מבריק Coomassie, destain במהלך שלושה ברציפות שוטף (h 1/לשטוף) עם פתרון destaining R-250 Coomassie כחול מבריק.
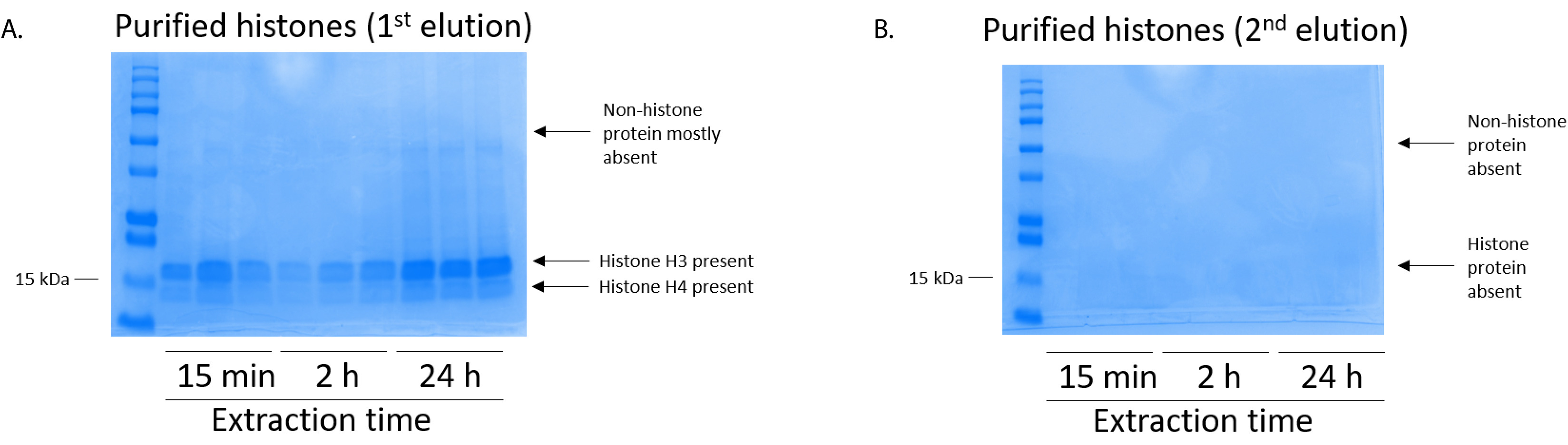
הערה: שינויים היסטוניים גולמי (איור 2) ניתן להשוות עם שינויים eluted וטיהר היסטוניים (קרי, קלט העמודה [איור 5A]).
3. טיהור של שינויים היסטוניים הליבה
-
ספין עמודה equilibration
- להוסיף 500 µL equilibration מאגר כל עמודה ספין בשימוש. אל תגעו קרום עמודה.
- צנטריפוגה ב 4 מעלות צלזיוס למשך 3 דקות ב x 800 גרם. למחוק את הזרימה דרך. חזור על 1 x.
-
היסטון טיהור
- להוסיף 500 µL של המדגם של הריבית משלב 2.6 העמודה. צנטריפוגה ב 4 מעלות צלזיוס למשך 3 דקות ב x 800 גרם. לאסוף את הזרימה דרך.
- חזור על השלב הקודם כמה פעמים לפי הצורך לטעון את המדגם כולו על העמודה. אל תמלא יותר מדי את העמודה ספין.
- לשלב את הזרימה דרך של כל שלב צנטריפוגה כדי לנתח את היעילות מחייב עמודה (איור 3).
- בצע את שלב 2.7 לנתח הזרימה עמודה של דרך.
-
שטיפת העמודה
- להוסיף 500 µL של המאגר לשטוף כל עמודה. צנטריפוגה ב 4 מעלות צלזיוס למשך 3 דקות ב x 800 גרם. לאסוף זרימה דרך בכביסה (שטיפת #1).
- חזור על צעד 3.3.1 בסך הכל שלושה שוטף. לאסוף את זרימה דרך שוטף #2 ו- #3. מאגר לא רצופות עמודה זרימה דרך שוטף.
- להערכה נוספת היסטון מחייב יעילות של העמודה, לנתח את מנקי עמודה 3 צעד בעקבות 2.7 (איור 4).
-
• תנאי היסטון
- להעביר את העמודה לתוך צינור mL 1.5 עם תוויות ברורות.
- להוסיף 50 µL של המאגר • תנאי היסטון. צנטריפוגה ב 4 מעלות צלזיוס למשך 3 דקות ב x 800 גרם. לשמור על זרימה דרך המכיל היסטון חלבונים.
- עבור • תנאי נוסף, חזור על שלב 3.4.2. אל תשלבו את הראשון והשני זרימה דרך eluates כפי שהם נבדלים היסטון כמות וטוהר.
4. משקעים של שינויים היסטוניים הליבה
- להוסיף חומצה על-כלורית (PCA) שינויים היסטוניים מטוהרים כדי ריכוז סופי של 4% PCA (למשל, להוסיף 3 µL של 70% PCA כדי 50 µL של שינויים היסטוניים מטוהרים מהשלב 3.4.2.
- צנטריפוגה 3 s כדי לאסוף את כל הנוזל שיורית מהקיר שפופרת. מערבבים על-ידי pipetting למעלה ולמטה 6 x.
- למקם את הצינורות בארון תקשורת, תקופת דגירה של 24 שעות-4 מעלות צלזיוס.
- למחרת, prechill microcentrifuge עד 4 ° C, centrifuge את הדגימות למשך 75 דקות במהירות המרבית ב 4 º C.
- עם השלמת צנטריפוגה, גלולה לבן קטן המכיל שינויים היסטוניים זירז יהיה גלוי על החלק התחתון של הצינור. לא מערבולת המדגם.
- בזהירות וארוקן את תגובת שיקוע והוסף, מבלי להפריע בגדר, 500 µL של 4% קר כקרח PCA לדגימת.
- צנטריפוגה ב 4 מעלות צלזיוס למשך 10 דקות במהירות המרבית. בזהירות וארוקן את תגובת שיקוע.
- חזור על שלב 4.7 2 x.
- מבלי להפריע בגדר, להוסיף 500 µL של אצטון קר כקרח. צנטריפוגה ב 4 מעלות צלזיוס למשך 10 דקות במהירות המרבית. בזהירות וארוקן את תגובת שיקוע.
- חזור על שלב 4.9 2 x.
- בזהירות וארוקן את תגובת שיקוע, להשאיר את צינורות uncapped, ולאפשר המדגם להתייבש על קרח במשך 30 דקות הסימון אם כל אצטון שיורית התאדו.
- . תעזוב הצינורות uncapped ולאפשר את הדגימה לייבוש בטמפרטורת החדר (RT) 5 דקות-
- Resuspend בגדר ב 30 µL של מים סטריליים. לא pipet למעלה ולמטה. קפיצי הצינור בעדינות עם האצבע.
- קאפ לכל הצינורות ולאפשר שינויים את היסטוניים צריך להשרות על קרח למשך 30-50 דקות, בהתאם לגודל גלולה. בדוק אם בגדר resuspended.
- קאפ לכל הצינורות ולאפשר בגדר עוד יותר resuspend-RT במשך 5 דקות.
הערה: פתרון זה (הראשונה והשנייה • תנאי מהשלב 3.4.3) היוו של שינויים היסטוניים מטוהרים, desalted, עשוי לשמש כמת נוספת וניתוח acetylation היסטון.
5. כימות של חלבוני היסטון Eluted
- השתמש ספקטרופוטומטרים לפי הפרוטוקול של היצרן כדי לכמת את החלבונים היסטון הכולל שהושג לאחר • סופית תנאי בשלב 4.15. למדוד את ספיגת ב- 230 ננומטר. להקליט את היחס A260/A280 מעידה על הזיהום מדגם עם חומצת גרעין.
- השתמש בנוסחה הבאה כדי לחשב את הריכוז היסטון (x):

כאן, יתר הוא הצפיפות האופטית נמדד ב A230 ננומטר. - היסטון ריכוז ~1.5 mg/mL נחשבת של התשואה הממוצעת עבור שורות תאים, בעוד היסטון ריכוז ~5.0 mg/mL נחשבת של התשואה הממוצעת עבור 30 מ"ג של רקמות.
6. תספיג ניתוח
- התאם את היסטון מטוהרים, eluted מן µg שלב 4.15-~ 10 היסטון חלבון/מדגם.
- להוסיף נפח מתאים של מים ו- 4 מאגר מדגם x Laemmli כדי להתאים את טעינת אמצעי האחסון.
- Denature את הדגימות 10 דקות ב 99 º C. מגניב אותם על קרח. צנטריפוגה 3 s כדי לאסוף את כל שאריות נוזל עיבוי מהקיר שפופרת.
- לטעון את הדוגמאות על גבי ג'ל מרחביות-דף ולהפעיל את הג'ל מאובטח ב- 100 וולט.
- כדי להמחיש היסטון הכוללת חלבון, מכתים את הג'ל במשך הלילה עם פתרון מוכתמים R-250 כחול מבריק Coomassie, destain במהלך שלושה שוטף ברציפות (1 h/לשטוף) עם פתרון destaining R-250 Coomassie כחול מבריק.
הערה: הראשון • תנאי של חלבוני היסטון מכילה שינויים באיכות גבוהה היסטוניים (איור 5A) בזמן • תנאי השני מכיל נמוך אין רמות של שינויים היסטוניים (איור 5B). - כדי לכמת PTMs היסטון, להשתמש במערכת העברת (ראה טבלה של חומרים) כדי להעביר את החלבון היסטון הג'ל מרחביות-דף (שלב 6.4) על גבי קרום PVDF.
- להרכיב את הכריך העברה, לפתוח את הקלטת מערכת העברת ולמקם הערימה ממברנה PVDF (מתויג בתור תחתון +) בתחתית הקלטת עם קרום פונה כלפי מעלה. לגלגל את הקרום בעדינות עם גלגלת חשופה כדי להסיר את כל האוויר שבין המחסנית ואת הקרום.
- להניח את הג'ל על גבי הקרום, לגלגל את הג'ל בעדינות עם גלגלת חשופה כדי להסיר את כל האוויר שבין הקרום, הג'ל, למקם את המחסנית העליונה על הג'ל. לגלגל בעדינות שוב, למקם את הכיסוי קלטת על הכריך, לחץ כלפי מטה בחוזקה, וסובב את נוב בכיוון השעון למנעול.
- הכנס את הקלטת אל החריץ מערכת העברה. במסך ' מכשיר ', בחר פרוטוקול טורבו. להשתמש בפרוטוקול 3 דקות ג'ל מיני יחיד או עבור יותר משני מיני ג'לים פרוטוקול 7 דקות.
- כתם הקרום עם כתם צבע מאכל פונסו S עבור 5 דקות והמחש את החלבונים היסטון הכולל.
- ותוא וקינ 1 x באגירה טריס תמיסת מלח (TBS) עם 0.1% 20 Tween עבור 2 h ולחסום אותם 5% חלב 1 ה Incubate עם נוגדן ראשוני ומשני (לילה 4 ° C או h 1-RT, בהתאמה) או על פי פרוטוקול בעבר ממוטבת.
הערה: ב הפרוטוקול הנוכחי, נוגדנים נגד שינויים היסטוניים acetylated H4K12 (איור 6 ואיור 7) ו H3K27 (איור 8) היו בשימוש.
תוצאות
כדי להמחיש את ההתקדמות של פרוטוקול טיהור היסטון והרכב של כל שברים שנותחה, הערכנו תמציות היסטון שונים מתאי האדם microglial BV2. כדי להדגים כימות של שינויים היסטוניים H3 ו- H4 PTMs (כלומר, acetylation), השתמשנו lysates רקמות המוח.
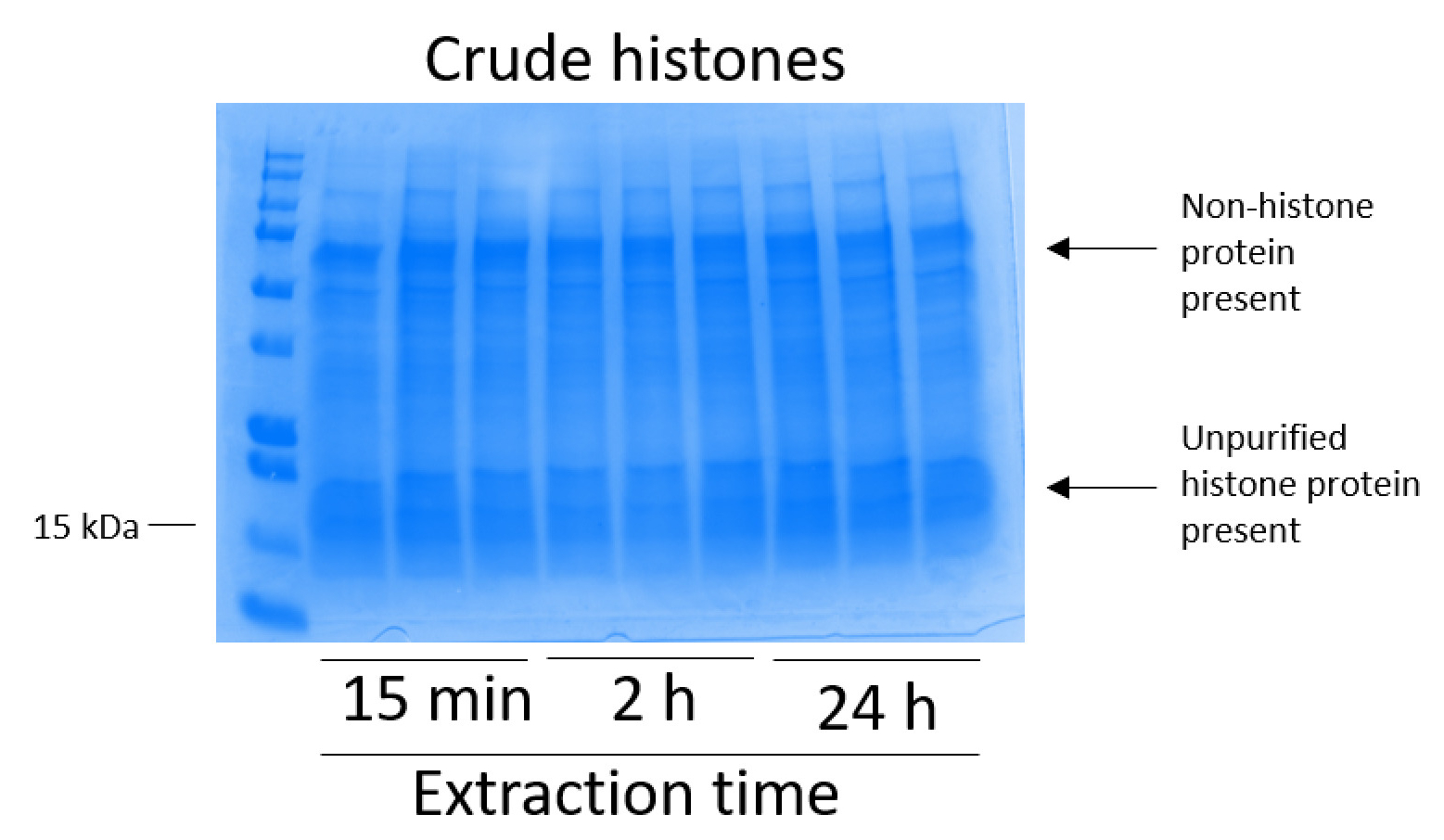
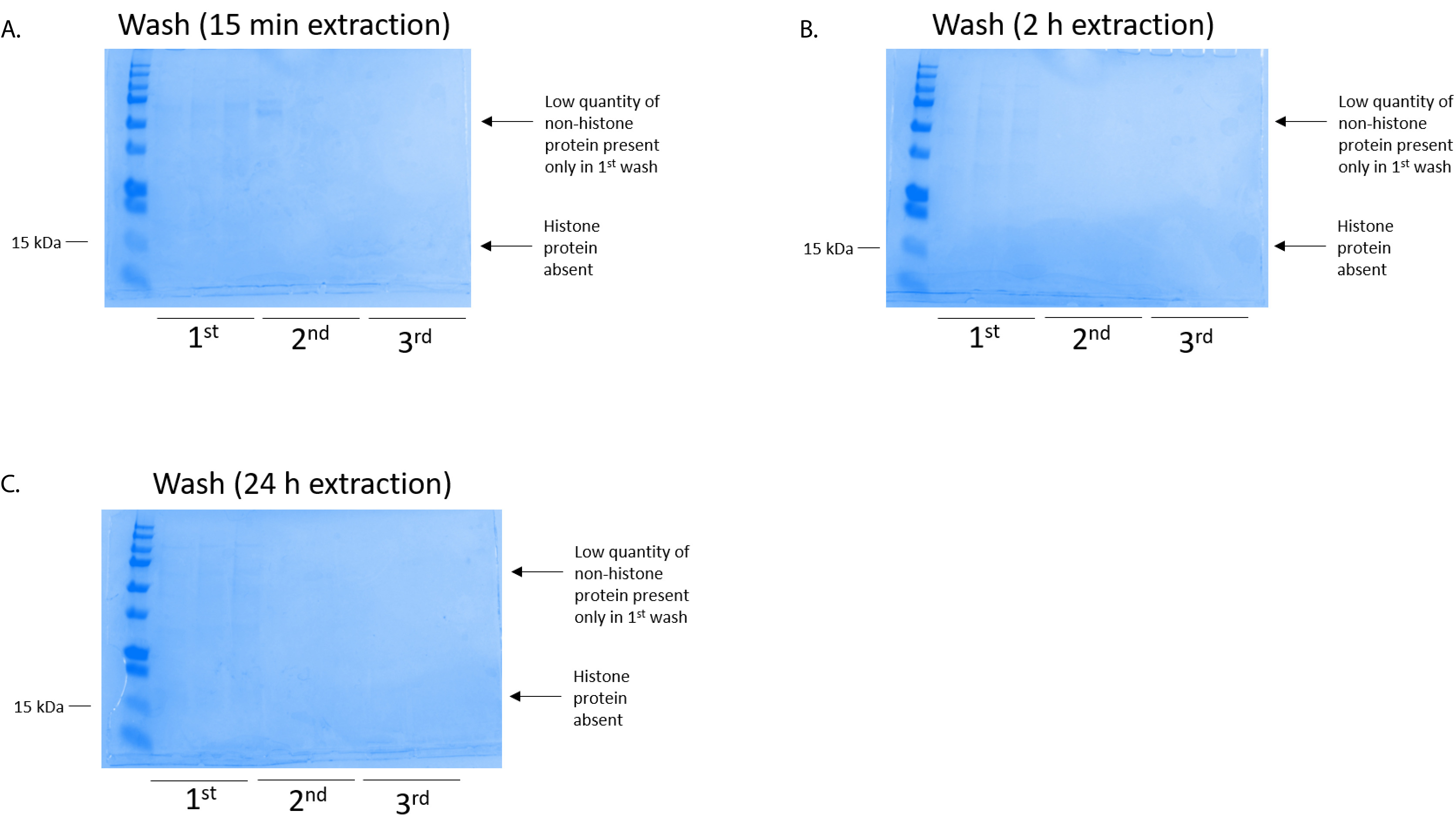
BV2 התאים היו מצופה ב 5 x 106 תאים לכל מאכל במנות שטופלו תרביות רקמה 10 ס מ, מותר כדי לגדול confluency עבור ה 48 תאים ואז נאספו, שינויים היסטוניים שוחררו כרומטין מאת דגירה במאגר החילוץ 0.4 מ' חומצה גופרתית 1 מ"מ אשלגן כלורי, 1 מ MgCl2, 50 מ מ טריס-HCl (pH 8.0), 1 x מעכב פרוטאז. הזמן החילוץ, בין 15 דקות, 24 שעות ביממה, לא השפיע על ההרכב הכולל של תמציות היסטון גולמי כפי שנקבע על ידי Coomassie מבריק כחול מכתים (איור 2). שינויים היסטוניים הבא, גולמי הועברו דרך העמודים equilibrated היסטון וניתח הזרימה דרך היה. יעילות גבוהה מחייב עמודה נקבעת על ידי העדר היסטון חלבון הזרימה דרך כאשר נותחו על ידי צביעת Coomassie כחול מבריק. אנחנו נקבע את היעילות מחייב טור יהיה 100% כפי שהיו לא חלבוני היסטון לזיהוי נוכחות שנותחה הזרימה דרך (איור 3). ממברנות כל עם שינויים היסטוניים מאוגד נשטפו ואז שלוש פעמים עם מאגר לשטוף כדי להסיר כל הטומאות הנותרים, עוזב חלבוני היסטון היחידה קשורה בג'ל סיליקה. אנחנו נקבע, לחילוץ כל היסטון פעמים (כלומר, 15 דקות, שעתיים ו 24 שעות), לשטוף את קרום הראשונה הייתה הדבר החשוב ביותר להסיר nonhistone מציג העמודות, בעוד שוטף השני והשלישי לא להשפיע על טוהר הדגימה. לפיכך, בהתאם לסוג לדוגמה, שוטף שני יכול להיות מושמט. בעקבות • הראשון תנאי היסטון החלבון מן העמודה (באמצעות מאגר • תנאי המכיל 1 מ מ NaCl EDTA), שינויים היסטוניים היו זירז בין לילה עם 4% חומצה על-כלורית מגורען, לשטוף, ואז ניתח את העשרה של שינויים היסטוניים מטוהרים H3 ו H4. הבחנו הזה 24 שעות ביממה של מיצוי הזמן מגביר כמות שינויים היסטוניים H3 ו- H4 ב השבר מטוהרים בהשוואה ל 15 דקות ו- h 2 זמן החילוץ (איור 5A). • השני תנאי מן העמודה לא לגרום שינויים היסטוניים באיכות גבוהה או גבוהה-הכמות (איור 5B).
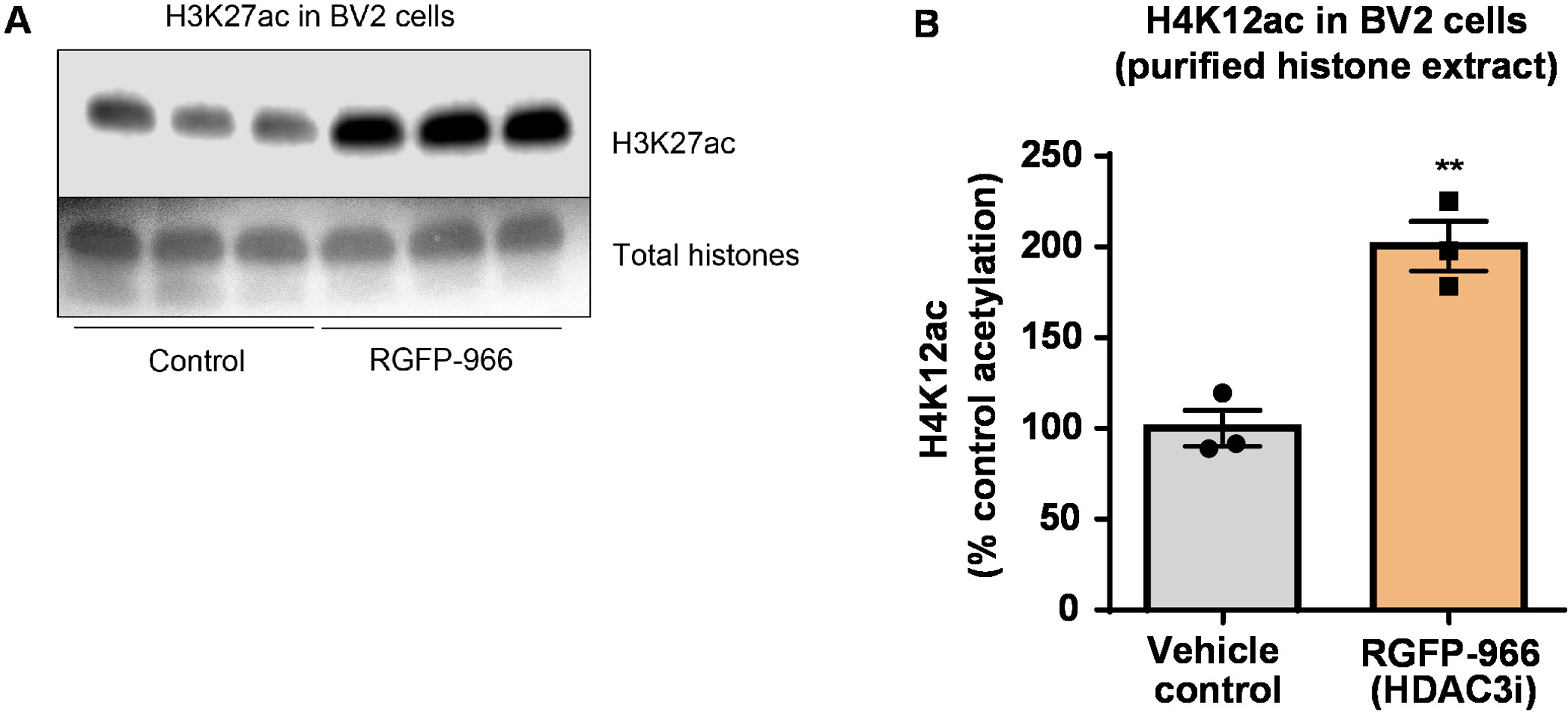
בשלב הבא, השתמשנו homogenates רקמת המוח לכמת היסטון H3 ו- H4 PTMs, כלומר acetylation. פראי-סוג (C57BL6/J) היו עכברים זכרים מנוהל מתנהג באופן כללי HDAC מעכב (tributyrin) במינון של 5 g/kg אוראלית עבור 3 רקמות ד שלם-מוח נאסף ביום 4, שינויים היסטוניים גולמי חולצו לפי הפרוטוקול המתואר. באמצעות של אינטראקצית t-test, קבענו את tributyrin מגביר את acetylation של שינויים היסטוניים תמצית גולמי (t(6) = 6.184, P = 0.0004); עם זאת, זיהומים מזוהים את תמצית (הלהקות היסטון אינם מוגדרים בבירור). לפיכך, נוגדן H4K12ac אין ירידה לפרטים גבוה (איור 6). להעריך עוד יותר את הישימות של הפרוטוקול הציג מקטעים קטנים של רקמות, אספנו את קליפת המוח הקדם חזיתית של מחלת אלצהיימר הטרנסגניים טריפל (3 x Tg-לספירה) עכברים שטופלו מדי יום 10 מ"ג/ק"ג M344, מחלקה, ואני IIb HDAC מעכב, לארבעה . חודשים. טיהור היסטון והמשקעים בוצעה לפי הפרוטוקול המתואר במסמך זה. שימוש מטוהרים היסטון H3 ו- H4 שבר, קבענו כי M344 מגביר H4K12 acetylation 2.4-fold (t(6) 13.03, P = < 0.0001), עם יחודיות גבוהה של נוגדנים H4K12ac (איור 7). באופן דומה, הבחנו עלייה היסטון acetylation H3 בתאים BV2 בתגובה אחרת HDAC מעכב, כלומר סלקטיבי HDAC3 המדכא, RGFP-966. 10 מיקרומטר של גורמים RGFP-966 כ גידול של acetylation-היסטון H3K27 לאחר 24 שעות של טיפול. תלמיד של אינטראקצית t-מבחן שימש כדי להשוות בין תאים שליטה לעומת מטופלים.

איור 1: ציר הזמן של פרוטוקול טיהור היסטון. כל השלבים עבור ניתוחים היסטון מוצגים להלן יחד עם הזמן הדרוש עבור כל שלב. איורים המתארים את התוצאה של צעדים מסוים והציג בתוך היד נקראים בסוגריים. אנא לחץ כאן כדי להציג גירסה גדולה יותר של הדמות הזאת.
{kind=link}

איור 2: נציג ג'ל כחול מבריק Coomassie שהוכתמו להדגים שינויים היסטוניים גולמי שחולצו מן התאים BV2. BV2 התאים היו תרבותי במשך 48 שעות לפני שהחלה פרוטוקול החילוץ היסטון. שינויים היסטוניים גולמי חולצו 15 דקות, שעתיים של 24 שעות, עם שלושה משכפל עבור כל נקודת זמן (זה גם במקרה של איור 3, איור 4, ואיור 5). הן nonhistone והן היסטון חלבונים נמצאים תמצית היסטון גולמי. אנא לחץ כאן כדי להציג גירסה גדולה יותר של הדמות הזאת.
{kind=link}

איור 3: נציג ג'ל כחול מבריק Coomassie שהוכתמו הוכחת עמודה זרימה דרך טיהור היסטון הבאים צעדים מתאי BV2. בעקבות היסטון הגולמי עובר דרך העמודה איגוד היסטון, רק nonhistone חלבונים נמצאים הזרימה דרך. חלבוני היסטון נעדרים בתוך השבר הזה. אנא לחץ כאן כדי להציג גירסה גדולה יותר של הדמות הזאת.
{kind=link}

איור 4: נציג ג'ל כחול מבריק Coomassie שהוכתמו הוכחת מכונת שטיפת העמודה בעקבות צעד טיהור היסטון מתאי BV2. בין הזמנים החילוץ של היסטון, אשר היו (א) 15 דקות, (B) 2 h, או (ג) 24 שעות ביממה, כמויות נמוכות של חלבונים nonhistone נכחו רק בעמודה הראשונה-שטיפת histonebinding. חלבוני היסטון נעדרו ב כל שוטף. אנא לחץ כאן כדי להציג גירסה גדולה יותר של הדמות הזאת.
{kind=link}

איור 5: נציג ג'ל כחול מבריק Coomassie שהוכתמו הוכחת elutions בעקבות צעד טיהור היסטון מתאי BV2. (א) באיכות גבוהה מטוהרים, desalted היסטון H3, H4 אותרו לאחר • הראשון תנאי מן העמודה טיהור היסטון. (B) • השני תנאי מן הטיהור היסטון עמודה לא נכנע באיכות גבוהה או כמות שינויים היסטוניים H3 או H4. אנא לחץ כאן כדי להציג גירסה גדולה יותר של הדמות הזאת.
{kind=link}

איור 6: בהרחבה מתנהג HDAC מעכב, tributyrin, מגבירה את acetylation H4K12 במוח שלם של פראי-סוג עכברים. (א) לוח זה מציג נציג המערבי חשופה המתארת גידול של H4K12 acetylation ב היסטון גסה תמצית אסף כל המוח של עכברים פראי-סוג בתגובה HDAC בהרחבה בפועל החומר המדכא, tributyrin. (B) לוח זה מציג כימות של העלייה H4K12 acetylation vivo בתוך. אינטראקצית t-test שימש כדי להשוות בין קבוצות (t(6) = 6.076, P = 0.0005). העמודות מסמנות את זאת אומרת ± שגיאת התקן של הממוצע (SEM). N = 8. P < 0.0001. אנא לחץ כאן כדי להציג גירסה גדולה יותר של הדמות הזאת.
{kind=link}

איור 7: המחלקה, ואני IIb HDAC מעכב, M344, מגבירה את acetylation H4K12 של קליפת המוח הקדם חזיתית של טריפל הטרנסגניים מחלת אלצהיימר (3 x Tg-לספירה) עכברים. (א) לוח זה מראה נציג המערבי כתם המתארת גידול של H4K12 acetylation ב מטוהרים היסטון desalted תמצית שנאסף קליפת המוח הקדם חזיתית של 3 x Tg-לספירה עכברים בתגובה עיכוב של HDACs על ידי M344. (B) לוח זה מציג כימות של העלייה של H4K12 acetylation בתגובה M344 ניתנת במינון יומי 10 מ"ג/ק"ג במשך ארבעה חודשים. אינטראקצית t-test שימש כדי להשוות בין קבוצות (t(6) 13.30, P = < 0.0001). העמודות מסמנות את זאת אומרת ± ב- SEM. N = 8. P < של 0.00001. אנא לחץ כאן כדי להציג גירסה גדולה יותר של הדמות הזאת.
{kind=link}

איור 8: מעכב סלקטיבי HDAC3, RGFP-966, מגדילה את H3K27 acetylation בתאים microglial BV2. (א) לוח זה מראה תספיג נציג המתאר גידול של H3K27 acetylation תמצית היסטון מטוהרים, desalted שנאספו מתאי BV2 בתגובה עיכוב HDAC3 על ידי RGFP-966. RGFP (B)-966 גורם כ גידול של acetylation-היסטון H3 וליזין (K) 27 לאחר 24 שעות של טיפול. אינטראקצית t-מבחן שימש כדי להשוות בין תאים שליטה לעומת מטופלים (t(4) = 5.981, P = 0.002). העמודות מסמנות את זאת אומרת ± ב- SEM. N = 6. P < 0.01. אנא לחץ כאן כדי להציג גירסה גדולה יותר של הדמות הזאת.
{kind=link}
| נציג רקמות (חצי כדור אחד) * | משקל ממוצע רקמות (מ ג) | מאגר החילוץ (mL) | מספר קווים |
| המוח הקטן העכבר | 40 | 1 | 40 |
| העכבר החזיתית | 30 | 0.3 | 20 |
| העכבר ההיפוקמפוס | 27 | 0.3 | 18 |
| העכבר entorhinal קליפת | 19 | 0.3 | 17 |
| * כל הניסויים בוצעו על עכברים זכרים בוגרים. | |||
| ממוצע גיל: 16 חודשים. משקל ממוצע: 30 גרם. | |||
טבלה 1: אופטימיזציה תנאי המגון רקמת המוח.
Discussion
בשנת העבודה הנוכחית, הפגנו שיטה ממוטבת כדי לטהר חלבוני היסטון הליבה ולכמת היסטון H3 ו- H4 PTMs (למשל, acetylation). פרוטוקול הציג הוא זרימת עבודה מקיפה, המשלבת ממוטב נהלים לגבי תאים, הכנה רקמת המוח, גולמי • תנאי טיהור היסטון, והמשקעים היסטון מפורט, ו כמת, אשר מלוות אלקטרופורזה היסטון, כימות PTM היסטון חזקים. כמות גדולה של פרטים שסופק כאן מאפשר לדור לשכפול של נתונים באיכות גבוהה, למרות הצורך מניפולציות ממושך של הדגימות היסטון.
פרוטוקולים המפורסמת כעת רבים דורשים שימוש HPLC לבודד שברים טהור של שינויים היסטוניים H3 ו- H419. למרות HPLC טכניקה חזקה, את המורכבות שלה ואת בתפוקה נמוכה להרתיע רוב פיצחו, nonexperts מ השימוש בתדירות גבוהה. אכן, HPLC אינה זמינה למעבדות רבים, אנשי צוות מיומנים נדרש להפעיל את הכלי. HPLC הוא לעתים קרובות גוזלת זמן יקר, העלולים להיות מסוכנים. המובאת כאן היא אסטרטגיית זול, תפוקה בינונית על מנת להשיג תוצאות באיכות דומה אשר עוקף HPLC. האסטרטגיה שדווח הוא גם מעשי יותר מתאים לשימוש במעבדה כמעט כל כפי שהיא משתמשת בגישה עמודה ספין פשוטה שאינה דורשת מיומנויות מבצע כלי נגינה מיוחדים. בנוסף, היסטון H3/H4 tetramer חילוץ כמו יחיד, טהור, שבר שופע, המאפשר כימות של אמין PTMs שהשתמרה על כל אחד החלבונים.
PTMs רגישים מאוד לשינויים סטרס חמצוני, שינויים ב- pH20,21. לפיכך, בניגוד שפורסמו בעבר שיטות18, מדווחים אסטרטגיית יעיל של שטיפה התאים בתקשורת ללא סרום כדי להבטיח הפרעה מטבולית המינימלי של התאים וכדי למנוע ההפרעה של PTMs מקורי עם סרום רכיבים. בפרוטוקול הנוכחי לא רק עוקף את הבידוד גרעינים מסורתי, אלא גם מספק פעמים אופטימלית עבור פירוק התא ואת ההליך המגון רקמות מדויק המאפשר השימור של המעטפה גרעיני תוך הימנעות צבירת גרעינית. למרות הזמן החילוץ יכול להיות מתומרן מבוסס על מספר התאים את סוג התא המשמש, רקמות גודל, וכו ', פירוק מורחב אינו רצוי כמו זה עלול להוביל פירוק של גרעינים, שחרור ה-DNA, מקשה על הדגימה לטפל. חשוב, להציב מחסומים מרובים הפרוטוקול קיימות עבור האימות של טיהור היסטון מוצלחת (למשל, שלבים 2.7 ו 3.2.3). אסטרטגיה זו מקלה גם על פתרון בעיות לאורך כל ההליך הממושך.
עוד תכונה חשובה וייחודית של פרוטוקול הציג הוא שלה תאימות מלאה עם כלי ניתוח תספיג במורד הזרם ואחרים אם כל כך השתוקקה. חלבוני היסטון מזוהים ~ 15 kDa13,22,23 ו, באופן דומה חלבונים אחרים משקל מולקולרי קטן, הוכחו מאתגר לזהות על-ידי טכניקות immunoblotting רגיל. השימוש של מערכת העברת ביצועים גבוהים, תפוקה גבוהה בשילוב עם רזולוציה מיטבית חלבון ג'ל מאפשר קיום אישור מקורי חלבון (בהיעדר מרחביות) ופעילות בהיעדר מרחביות ויעילות העברה גבוהים של החלבונים היסטון נמוך משקל מולקולרי, ומבטיח כימות PTM היסטון אמין.
Disclosures
המחברים אין לחשוף.
Acknowledgements
המחברים להביע את תודתם תכנית המחקר משרד הבריאות של פלורידה אד ואלצהיימר אתל מור (מענקים 6AZ08 ו- 7AZ26), NIH-NIAAA (גרנט 5R01AA023781-03) של איגוד הלב האמריקני (גרנט 17PRE33660831).
Materials
| Name | Company | Catalog Number | Comments |
| 1.5 mL microcentrifuge tubes | ThermoFiser Scientific | 05-408-129 | or equivalent from other sources |
| Sterile water | Gibco | 15-230-204 | or equivalent from other sources |
| 70% perchloric acid | Sigma Aldrich | 311421 | or equivalent from other sources |
| 100% acetone | Sigma Aldrich | 270725 | or equivalent from other sources |
| pH-indicator strips, non-bleeding | Milliipore Sigma | 1095310001 | |
| 4x SDS sample buffer | BIO-RAD | 161-0747 | |
| Benchtop rotor | Cole-Parmer | UX-04397-34 | or equivalent from other sources |
| 1.5 mL tube rack | ThermoFiser Scientific | 05-541 | or equivalent from other sources |
| Histone purification mini kit | Active Motif | 40026 | spin columns included in the kit |
| Protease Inhibitor Cocktail | ThermoFiser Scientific | 78430 | or equivalent from other sources |
| Nanodrop instrument | ThermoFiser Scientific | ND-2000 | |
| Tissue culture dishes | VWR | 10062-880 | required for histone extraction from cultured cells |
| Tissue culute media | varies based on cell line used | varies based on cell line used | required for histone extraction from cultured cells |
| Low-serum media | ThermoFiser Scientific | 51985091 | required for histone extraction from cultured cells |
| Plastic cell scraper | Falcon | 353086 | required for histone extraction from cultured cells |
| SDS-PAGE gradient gel | BIO-RAD | 456-9035 | required for histone extraction from cultured cells |
| Coomassie Brilliant Blue R-250 Staining Solution | BIO-RAD | 1610436 | required for histone extraction from cultured cells |
| Coomassie Brilliant Blue R-250 Destaining Solution | BIO-RAD | 1610438 | required for histone extraction from cultured cells |
| Trans-Blot Turbo Mini PVDF Transfer Packs | BIO-RAD | 1704156 | required for histone extraction from cultured cells |
| Trans-Blot Turbo Transfer System | RIO-RAD | 1704150 | required for histone extraction from cultured cells |
| Ponceau S stain | CellSignalling | 59803S | required for histone extraction from cultured cells |
| Dounce homogenizer (size/cap sc 7mL) with a small size clearance | Kimble Chase | 885302-0007 | required for histone extraction from tissues |
| 100% bleach | Clorox | 68973 | required for histone extraction from tissues |
| H4K12ac antibody | Active Motif | 39166 | required for PTMs quantification via WB |
| H3K27ac antibody | Active Motif | 39134 | required for PTMs quantification via WB |
References
- Holliday, R. Is there an Epigenetic Component in Long-term Memory? Journal of Theoretical Biology. 200, 339-341 (1999).
- DeWoskin, V. A., Million, R. P. The epigenetics pipeline. Nature Reviews Drug Discovery. 12, 661-662 (2013).
- Eberharter, A., Becker, P. B. Histone acetylation: a switch between repressive and permissive chromatin. EMBO Reports. 3, 224-229 (2002).
- Grunstein, M. Histone acetylation in chromatin structure and transcription. Nature. 389, 349-352 (1997).
- Sartor, G. C., Powell, S. K., Brothers, S. P., Wahlestedt, C. Epigenetic Readers of Lysine Acetylation Regulate Cocaine-Induced Plasticity. The Journal of Neuroscience. 35, 15062-15072 (2015).
- Komatsu, N., et al. SAHA, a HDAC inhibitor, has profound anti-growth activity against non-small cell lung cancer cells. Oncology Reports. 15, 187-191 (2006).
- Bahari-Javan, S., Sananbenesi, F., Fischer, A. Histone-acetylation: a link between Alzheimer's disease and post-traumatic stress disorder. Frontiers in Neuroscience. 8, 160(2014).
- Roh, T. -Y., Cuddapah, S., Zhao, K. Active chromatin domains are defined by acetylation islands revealed by genome-wide mapping. Genes & Development. 19, 542-552 (2005).
- Mutskov, V., Felsenfeld, G. Silencing of transgene transcription precedes methylation of promoter DNA and histone H3 lysine 9. The EMBO Journal. 23, 138-149 (2004).
- Howe, L., Brown, C. E., Lechner, T., Workman, J. L. Histone acetyltransferase complexes and their link to transcription. Critical Reviews in Eukaryotic Gene Expression. 9, 231-243 (1999).
- Jenuwein, T., Allis, C. D. Translating the histone code. Science. 293, 1074-1080 (2001).
- Bowman, G. D., Poirier, M. G. Post-Translational Modifications of Histones That Influence Nucleosome Dynamics. Chemical Reviews. 115, 2274-2295 (2015).
- Volmar, C. -H., Wahlestedt, C. Histone deacetylases (HDACs) and brain function. Neuroepigenetics. 1, 20-27 (2015).
- Plagg, B., Ehrlich, D., Kniewallner, K. M., Marksteiner, J., Humpel, C. Increased Acetylation of Histone H4 at Lysine 12 (H4K12) in Monocytes of Transgenic Alzheimer's Mice and in Human Patients. Current Alzheimer Research. 12, 752-760 (2015).
- Bhaskara, S., et al. Hdac3 is essential for the maintenance of chromatin structure and genome stability. Cancer Cell. 18, 436-447 (2010).
- Mottamal, M., Zheng, S., Huang, T. L., Wang, G. Histone Deacetylase Inhibitors in Clinical Studies as Templates for New Anticancer Agents. Molecules. 20, 3898-3941 (2015).
- Ramakrishnan, S., et al. HDAC 1 and 6 modulate cell invasion and migration in clear cell renal cell carcinoma. BMC Cancer. 16, 617(2016).
- Wapenaar, H., Dekker, F. J. Histone acetyltransferases: challenges in targeting bi-substrate enzymes. Clinical Epigenetics. 8, 59(2016).
- Klinker, H., Haas, C., Harrer, N., Becker, P. B., Mueller-Planitz, F. Rapid Purification of Recombinant Histones. PLoS ONE. 9, e104029(2014).
- Chen, K., et al. Neurodegenerative Disease Proteinopathies Are Connected to Distinct Histone Post-translational Modification Landscapes. ACS Chemical Neuroscience. 9, 838-848 (2018).
- Simithy, J., Sidoli, S., Garcia, B. A. Integrating Proteomics and Targeted Metabolomics to Understand Global Changes in Histone Modifications. Proteomics. , e1700309 (2018).
- Volmar, C. -H., et al. An Epigenetic Approach for the Modulation of Amyloid Precursor Protein (APP) Processing and Improvement of Memory in Alzheimer's Disease. Neuropsychopharmacology: official publication of the American College of Neuropsychopharmacology. 40, S470(2015).
- Volmar, C. -H., et al. M344 promotes nonamyloidogenic amyloid precursor protein processing while normalizing Alzheimer’s disease genes and improving memory. Proceedings of the National Academy of Sciences. 114 (43), E9135-E9144 (2017).
Reprints and Permissions
Request permission to reuse the text or figures of this JoVE article
Request PermissionExplore More Articles
This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. All rights reserved