
Method Article
אקס ויוו שחרור פפטיד הקשור לגן קלציטונין ממערכת Trigeminovascular במכרסמים
In This Article
Summary
הפרוטוקול הנוכחי מתאר את מודל שחרור הפפטיד הקשור לגן ex vivo calcitonin (CGRP) ואת האסטרטגיה לכמת את ההשפעה של חומרים פרמקולוגיים על כמות CGRP המשתחררת מהמערכת הטריגמינווסקולרית במכרסמים.
Abstract
פפטיד הקשור לגן קלציטונין (CGRP) התגלה לראשונה בשנות ה-80 של המאה ה-20 כגרסת שחבור מהגן קלציטונין. מאז גילויו, תפקידו בפתופיזיולוגיה של מיגרנה התבסס היטב, תחילה על ידי תכונות מרחיב כלי הדם החזקות שלו ולאחר מכן על ידי נוכחותו ותפקודו כמוליך עצבי במערכת הטריגמינווסקולרית החושית. היכולת מעוררת המיגרנה של CGRP נתנה תמיכה לתעשיית הפארמה לפתח נוגדנים חד שבטיים ואנטגוניסטים המעכבים את ההשפעה של CGRP. פרדיגמת טיפול חדשה הוכחה כיעילה בטיפול מניעתי במיגרנה. אחד הכלים השימושיים להבנה נוספת של מנגנוני מיגרנה הוא מודל ex vivo של שחרור CGRP מהמערכת הטריגמינווסקולרית. זוהי שיטה פשוטה יחסית שניתן להשתמש בה עם כלים פרמקולוגיים שונים כדי להשיג ידע כיצד להמשיך ולפתח טיפולים יעילים חדשים במיגרנה. הפרוטוקול הנוכחי מתאר מודל שחרור CGRP ואת הטכניקה לכימות ההשפעה של חומרים פרמקולוגיים על כמות CGRP המשתחררת מהמערכת הטריגמינווסקולרית במכרסמים. הליך המתאר את הגישה הניסויית מהמתת חסד למדידת רמות החלבון מסופק. הבידוד החיוני של הגנגליון הטריגמינלי והגרעין הטריגמינלי caudalis הן מעכברים והן מחולדות והכנת דורה מאטר חולדה מתוארים בפירוט. יתר על כן, מוצגות תוצאות מייצגות משני המינים (חולדות ועכברים). הטכניקה היא כלי מפתח לחקר המנגנונים המולקולריים המעורבים בפתופיזיולוגיה של מיגרנה באמצעות תרכובות פרמקולוגיות שונות ובעלי חיים מהונדסים גנטית.
Introduction
מיגרנה היא הפרעה נוירולוגית, על פי ארגון הבריאות העולמי, מוערך להשפיע על יותר מ -1 מיליארד אנשים והוא אחד הגורמים המובילים של נכות ברחבי העולם1. לפיכך, למיגרנה יש השפעה משמעותית הן על המטופלים והן על החברה. למרות ההצלחה הקלינית האחרונה של תרופות נגד CGRP, חלק גדול מהחולים זקוקים לאפשרויות טיפול משופרות 2,3,4,5. נדרשת הבהרה של פתופיזיולוגיה של מיגרנה המובילה לטיפולים יעילים חדשניים. איתות בתוך המערכת הטריגמינווסקולרית המורכבת מקרום המוח, הגרעינים הטריגמינליים (TG) והגרעין הטריגמינלי caudalis (TNC) הוא מרכזי לפתופיזיולוגיה של מיגרנה 6,7.
37 חומצות האמינו נוירופפטיד קלציטונין הקשורות לגן (CGRP) התגלה לראשונה בתחילת שנות ה-80 של המאה ה-20, כאשר אמארה ועמיתיו הראו כי ניתן לעבד את תעתיק ה-RNA הראשוני של הגן קלציטונין כדי לתת קידוד mRNA ל-CGRP בנוסף לקלציטונין 8,9. המחקר שלאחר מכן הציע קשר לפתופיזיולוגיה של מיגרנה10. CGRP הוא נוירוטרנסמיטר עם תכונות מרחיבות כלי דם חזקות 11,12,13,14,15,16,1 7, והוא מופץ באופן נרחב במערכת העצבים המרכזית וההיקפית 13,14,18,19,20,21,22 . המעורבות של CGRP במיגרנה הודגשה עם גילוי רמות CGRP מוגברות במחזור הדם התוך-מוחי במהלך התקפי מיגרנהבבני אדם 23, וכי עירוי של CGRP גורם לכאב דמוי מיגרנה בחולים24. שנתיים לאחר מכן, פורסם מחקר הוכחת ההיתכנות הראשון של האפקטיביות של אנטגוניסט CGRP olcegepant בטיפול במיגרנה25.
CGRP מצוי בשפע במערכת הטריגמינווסקולרית כפי שהודגם ב-TG21,26, סיבי עצב תחושתיים המעוררים את הדורה מאטר27,28,29 ו-TNC30. במערכת הטריגמינווסקולרית, CGRP נמצא בתאי העצב הקטנים עד בינוניים של ה-TG, בסיבי C לא מיאלינים, ומתבטא בכמעט 50% מהאוכלוסייה העצבית של ה-TG. קולטן CGRP מתבטא בעיקר בתאי עצב גדולים יותר ונמצא בסיבי Aδ31,32. CGRP משתחרר מתאי עצב לאחר גירוי כימי או חשמלי33,34. מחקרים על מסלולים המובילים לשחרור CGRP ומיקום הפעלה זו חיוניים להבנת פתופיזיולוגיה של מיגרנה. במהלך 5 העשורים האחרונים, מחקרים פרה-קליניים תרמו לרכישת ידע נרחב על איתותים הקשורים למיגרנה ותרמו לפיתוח טיפולים חדשים35. שיטות רבות המתחשבות במעורבות וסקולרית ונוירוגנית שונו ויושמו בחקר המיגרנה. ניתן להזכיר מודלים in vivo ו- in vitro של תגובות עורקים לתרכובות ביולוגיות או טיפולים תרופתיים17,36,37, וגירוי עצבי חשמלי38,39. יתר על כן, נוירונים מופעלים ב- TNC יכולים להיות מזוהים על ידי ביטוי c-Fos 40,41,42 והקלטות אלקטרופיזיולוגיות באזור זה 43,44. שתי השיטות מודדות אותות נוסיצפטיביים המועברים למוח מהראש, למשל דורה מאטר. השימוש במודל פרה-קליני אחד בלבד אינו מציג את התמונה המלאה של פתופיזיולוגיה של מיגרנה. לכן, חשוב לשלב מודלים שונים המכסים היבטים רבים ככל האפשר של פתופיזיולוגיה מיגרנה. המשך הפיתוח של מודלים חדשים יכסה היבטים שונים של מנגנוני מיגרנה, ועם הזמן תיחשף תעלומת הפתופיזיולוגיה של המיגרנה.
כאן, פרוטוקול מפורט מוצג של שיטת שחרור CGRP, שבוצעה ex vivo ב TG מבודד ו TNC מעכברים לאחר גירוי כימי. שחרור CGRP ניתן לחקור גם בדורה מאטר מחולדות. לפיכך, בפרוטוקול הניסוי לחולדות, דורה מאטר מתוארת יחד עם TG ו- TNC. הבסיס לשיטת שחרור CGRP תואר לראשונה בשנת 1999, שם אברסברגר ועמיתיו ערכו מחקר חלוצי ומצאו כי CGRP שוחרר מדורא מאטר לאחר גירוי כימי וחשמלי של אפרנטים דוראליים בחולדות45. מאוחר יותר, גישה זו הורחבה לשחרור CGRP מה-TG46 וה-TNC47. לאחר מכן, השיטה שונתה כך שתחול על TG ו- TNC בעכברים. עד כה, שחרור CGRP מהדורה מאטר היה מאתגר בעכברים.
Protocol
כל הליכי הטיפול והניסויים בבעלי חיים בוצעו בהתאם למדריך הקהילה האירופית לטיפול ושימוש בבעלי חיים (2010/63/UE). עכברי C57BL/6JBomTac זכרים, בני 10 שבועות, ועכברי ספראג דאולי זכרים, בני 10 שבועות, שימשו להדגמת פרוטוקול זה.
1. הכנת הנוזל הבין-תאי הסינתטי
- הכינו נוזל אינטרסטיציאלי סינתטי (SIF) על פי המתכון הבא: 108 mM NaCl, 3.48 mM KCl, 3.50 mM MgSO 4, 26 mM NaHCO3, 11.70 mM NaH 2 PO4, 1.50 mM CaCl2, 9.60 mM Na-gluconate, 5.50 mM גלוקוז ו-7.60 mM סוכרוז (ראו טבלת חומרים).
הערה: SIF יכול להיות מגוון בהתאם ליעד הגירוי, למשל, ניתן להשתמש בתמיסה נטולת סידן בעת חקר תעלות סידן. - התאימו את ה-pH ל-7.4 וייצבו את ה-pH על ידי גז קרבוגן (5% CO 2 ו-95% O2)46.
2. המתת חסד
- הרדימו עכברים וחולדות בוגרים בתערובת של 70% CO 2 ו-30% O2. ערפו עכברים באמצעות זוג מספריים וחולדות באמצעות גיליוטינה (ראו טבלת חומרים).
הערה: השתמש בזן ובגיל המתאימים למטרת המחקר. גם זכרים וגם נקבות יכולים לשמש במודל זה. - הפרד את הראש מהגוף ברמת C3-C4 של חוט השדרה.
הערה: המתת חסד יכולה להתבצע גם עם הזרקה תוך-צפקית של פנטוברביטל (100-150 מ"ג/ק"ג).
3. דיסקציה
- הכינו את רקמת החולדה לפי השלבים הבאים.
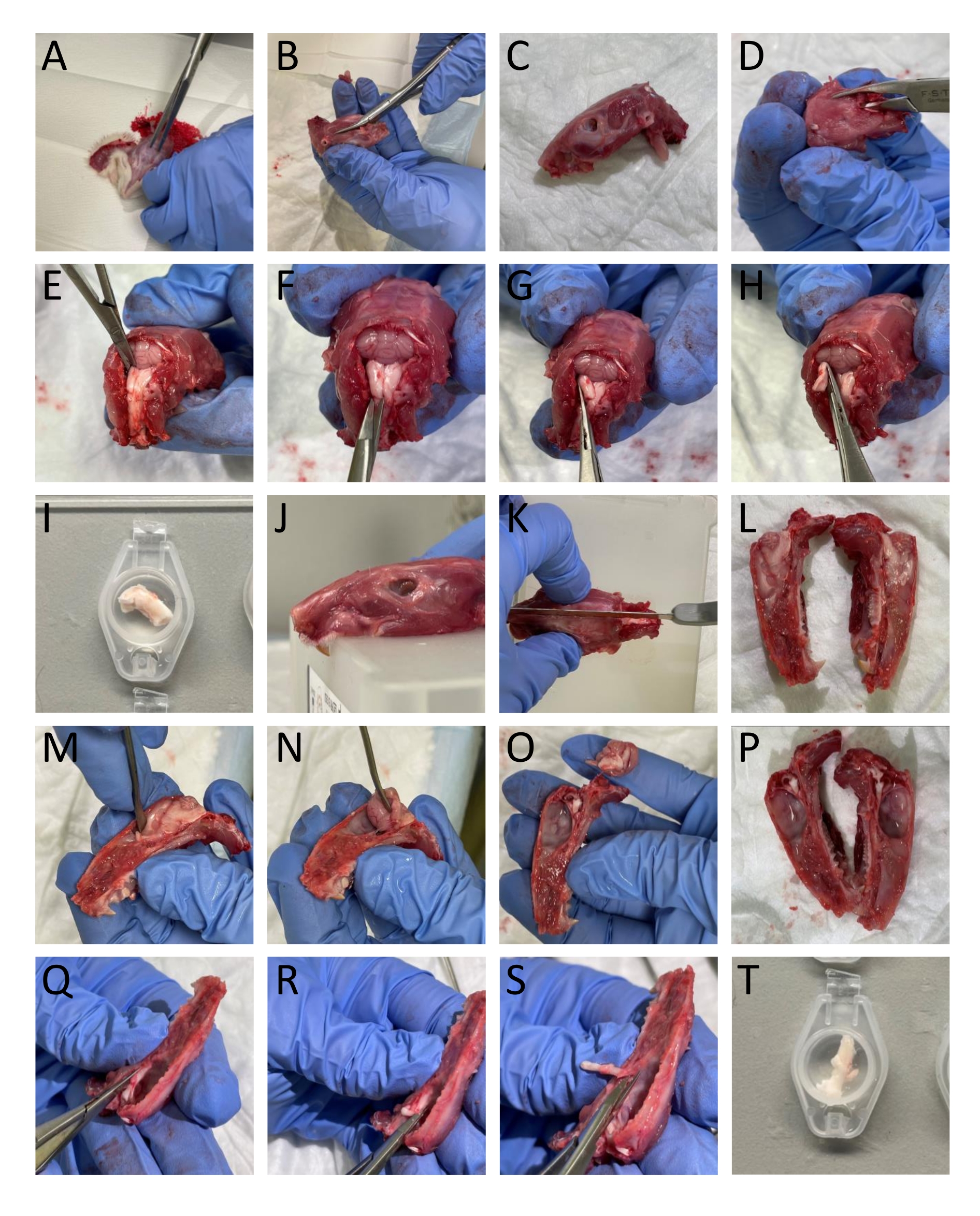
- הסר את העור ואת השריר סביב הראש והצוואר באמצעות זוג מספריים (איור 1A).
- השתמשו בגוזם עצמות ובמספריים כדי להפריד בין הלסתות התחתונות לראש (איור 1B-C).
- פתחו את חוט השדרה על-ידי החדרת גוזם עצם לתוך החלק הגבי של החוליות והסירו את החלק הגבי של החוליות כדי לחשוף את חוט השדרה ואת גזע המוח (איור 1D).
- חתכו את החלק הקאודאלי של הגולגולת בגבולות העצמות העורפיות והבין-פריאטליות כדי להסיר את מבני העצם האלה שחושפים את המוח הקטן (איור 1E).
הערה: חשוב לא לפגוע בגזע המוח ובחוט השדרה בעת חיתוך והסרה של החוליות. - יש לבודד את ה-TNC (Sp5C) הפועל באופן קאודיאלי במרחק של כ-13-16 מ"מ מהברגמה מכל צד על ידי חיתוך החלק הגבי של גזע המוח באמצעות מספריים קפיציים. היסחף את ה-TNC השמאלי והימני לתוך SIF (איור 1E-I).
הערה: התיאור מתאים לחולדות בוגרות. - חתכו את הראש באמצע הגולגולת כדי לחלק את הגולגולת לשניים באמצעות מסור (איור 1J-L).
- הסירו בזהירות את המוח מבלי לגעת בדורה מאטר המחוברת לגולגולת באמצעות מרית וחתכו את העצב הטריגמינלי במקום שבו הוא נכנס לגזע המוח (איור 1M-P).
- כדי לבודד את ה- TG, חתוך אותו, כולל ענפיו סביב הגבולות החזותיים. חותכים את הענף המנדיבולרי שבו הוא נכנס לאובלה של פורמן. חותכים את ענפי העיניים והמקסילריים הנכנסים לגולגולת מכיוון שהם אינם מחולקים באופן מקרוסקופי. בזמן ניתוח ה-TG, הסירו את הדורה מאטר שמכסה את ה-TG (איור 1Q-T).
- לטבול את חצאי הגולגולת ואת ה- TGs ב- SIF.
- הכן את רקמת העכבר לפי השלבים הבאים.
- הסירו את העור והשרירים סביב הראש והצוואר באמצעות מספריים קטנים (איור 2A-B).
- פתח את חוט השדרה על-ידי החדרת זוג מספריים קטנים לתוך החלק הגבי של החוליות והסר את החלק הגבי של החוליות כדי לחשוף את חוט השדרה ואת גזע המוח (איור 2C).
הערה: חשוב לא לפגוע בחוט השדרה בעת חיתוך והסרה של החוליות. - חתכו את הגולגולת בגבולות העצמות העורפיות והבין-פריאטליות כדי להסיר את מבני העצמות האלה שחושפים את המוח הקטן (איור 2D-F).
- לאחר מכן, חתכו את עצם הקודקוד באמצע הקשת והסירו את העצם כדי לחשוף את המוח הגס (איור 2G-I).
- הסירו בזהירות את המוח הקטן בעזרת מרית כדי לחשוף את גזע המוח (איור 2J).
- בודדו את החלק המכיל TNC בגזע המוח באמצעות מספריים קפיציים (איור 2K-N). טבול את גזע המוח עם TNC ב-SIF (איור 2O).
- הסירו את המוח וחתכו את העצב הטריגמינלי במקום שבו הוא נכנס לגזע המוח (איור 2P-Q).
- כדי לבודד את ה- TG, חתוך אותו, כולל ענפיו סביב הגבולות החזותיים. חותכים את הענף המנדיבולרי שבו הוא נכנס לאובלה של פורמן. חותכים את ענפי העיניים והמקסילריים הנכנסים לגולגולת מכיוון שהם אינם מחולקים באופן מקרוסקופי. בזמן ניתוח ה-TG, הסירו את הדורה מאטר שמכסה את ה-TG (איור 2R-S).
- לטבול TGs ב-SIF (איור 2T).
4. כביסה
- שטפו חצאי גולגולת, TNCs ו-TGs ב-SIF למשך 30 דקות תוך החלפת ה-SIF כל 5 דקות בטמפרטורת החדר.
הערה: ניתן לבצע את שלבי הכביסה תוך שמירה על הרקמה במיכלי פלסטיק עם מכסה טול כדי לאפשר החלפת SIF קלה (איור 3A-B). - הכינו את רקמת החולדה לפי השלבים הבאים.
- העבר חצאי TNC למכסי צינורות מיקרוצנטריפוגה נפרדים עם 350 μL של SIF (איור 3C).
- העבר את ה-TGs למכסי צינורות מיקרוצנטריפוגה - TG אחד לכל מכסה צינור מיקרוצנטריפוגה עם 350 μL של SIF (איור 3C).
- הניחו את חצאי הגולגולת על פלטפורמות העשויות מחימר או מצלחת תרבית בעלת 6 בארות ומלאו את הגולגולת ב-400 מיקרו-ליטר של SIF (איור 3C).
- הכן את רקמת העכבר לפי השלבים הבאים.
- העבירו את גזע המוח עם TNC למכסה צינור מיקרוצנטריפוגה עם 250 μL של SIF (איור 3D).
- העבירו את שני ה-TGs למכסה צינור מיקרוצנטריפוגה עם 250 μL של SIF (איור 3D).
הערה: יש להניח שני TGs לכל מכסה צינור מיקרוצנטריפוגה בעת שימוש ברקמה מעכברים.
- הניחו גולגולות חולדות וכובעי צינורות מיקרוצנטריפוגה עם רקמת חולדה ועכבר באינקובטור לח בטמפרטורה של 37 מעלות צלזיוס. החלף SIF באמצעות פיפטה כל 5 דקות למשך 20 דקות.
הערה: חשוב לא לגעת ברקמה בעת הוספה והסרה של SIF.
5. בדיקות סמים
- קבע את רמות השחרור הבסיסיות של CGRP.
- לאחר הכביסה האחרונה, יש להוסיף 250 μL של SIF לעכבר TG ו-TNC. הוסיפו 350 μL לחולדה TG ו-TNC ו-400 μL לכל גולגולת חולדה.
- לאחר 10 דקות של דגירה, אסוף 200 μL של הדגימה בצינור microcentrifuge והוסף 50 μL של 10x EIA buffer (מסופק עם ערכת האנזים CGRP immunoassay, ראה טבלת חומרים) כדי לאפשר מדידה של שחרור CGRP הבסיסי (שלב 6). יש להשליך את הנוזל הנותר.
הערה: זמן הדגירה חייב להיות זהה עבור כל הדגימות. - מיד, לאחסן את הדגימות ב -20 מעלות צלזיוס.
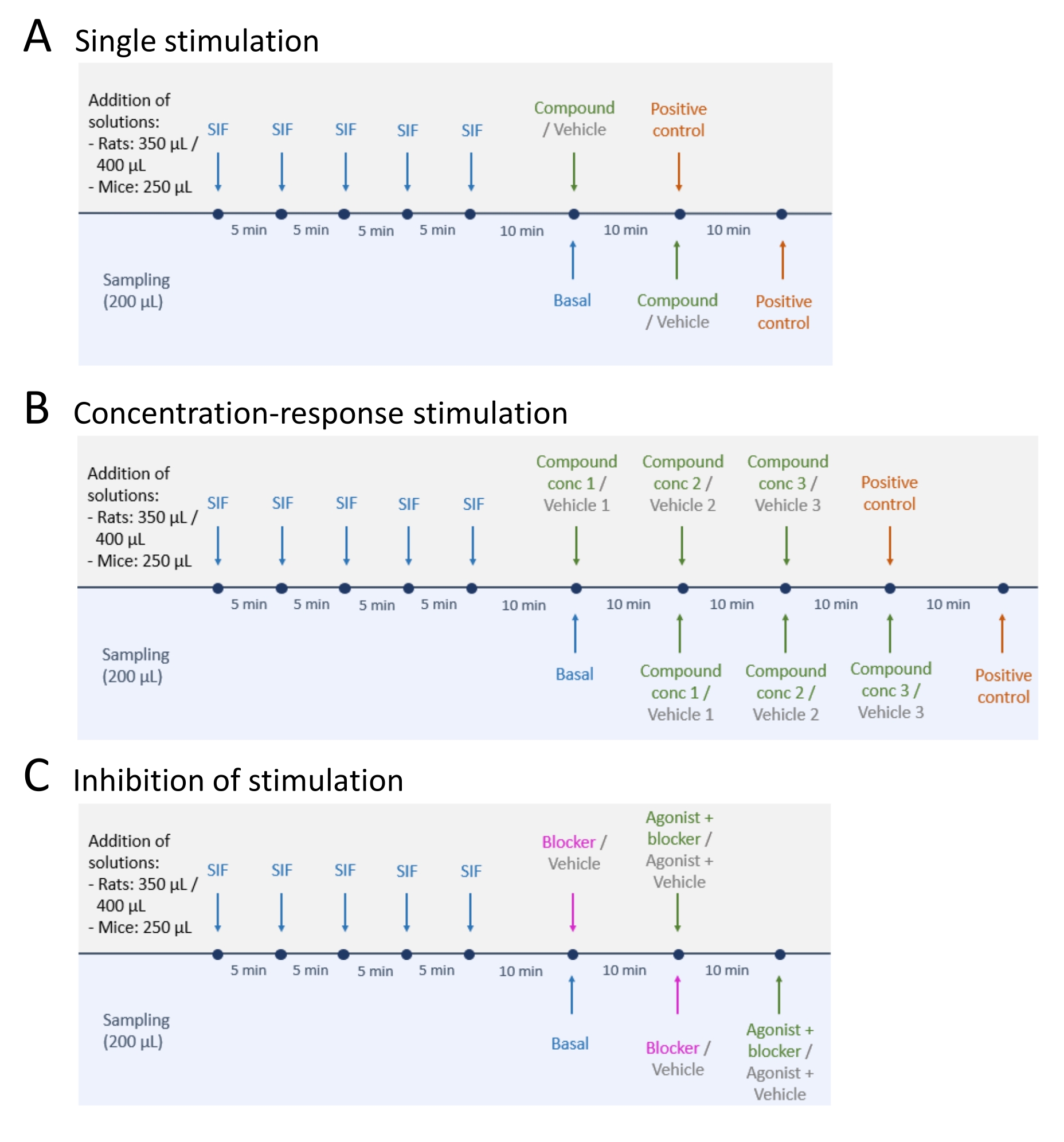
- לאחר דגימה של רמות שחרור CGRP בסיסיות, בצע אחת משלוש השיטות: (A) גירוי יחיד (שלב 5.2); (B) גירוי ריכוז-תגובה (שלב 5.3); ו-(C) עיכוב גירוי (שלב 5.4) (איור 4).
הערה: תרכובת הבדיקה והריכוזים שבהם נעשה שימוש תלויים במטרת המחקר.
- בצע גירוי יחיד בהתאם לשלבים הבאים.
- הוסיפו תרכובת בדיקה או רכב לרקמה והשאירו אותה למשך 10 דקות (נפח: 250 μL עבור עכבר TG ו-TNC, 350 μL עבור TG ו-TNC של חולדה, ו-400 μL עבור כל גולגולת חולדה).
- לאחר 10 דקות של דגירה, לאסוף 200 μL של הדגימה בצינור microcentrifuge עם 50 μL של 10x EIA חיץ. יש להשליך את הנוזל הנותר ולאחסן מיד את הדגימות בטמפרטורה של -20°C.
הערה: זמן הדגירה חייב להיות זהה עבור כל הדגימות.
- בצע גירוי תגובת ריכוז בהתאם לשלבים הבאים.
- יש לדלל את תרכובת הבדיקה או הרכב לריכוזים הרצויים. הוסף את תרכובת הבדיקה בהגדלת הריכוזים החל מהריכוז הנמוך ביותר.
הערה: תרכובת הבדיקה והריכוזים שבהם נעשה שימוש תלויים במטרת המחקר. לדוגמה, 1 μM, 10 μM, ו 100 μM supercinnamaldehyde שימשו במחקר הנוכחי. - הוסף את הריכוז הנמוך ביותר (1 μM עבור המחקר הנוכחי) של תרכובת הבדיקה והרכב המתאים לשני תכשירי רקמה זהים ודגרה במשך 10 דקות (נפח: 250 μL עבור רקמת עכבר, 350 μL עבור חולדה TG ו- TNC, ו 400 μL עבור כל גולגולת חולדה).
- לאחר 10 דקות של דגירה, לאסוף 200 μL של הדגימה בצינור microcentrifuge עם 50 μL של 10x EIA חיץ.
- יש להשליך את הנוזל הנותר ולהוסיף את הריכוז השני הנמוך ביותר (10 מיקרומטר למחקר הנוכחי) לרקמה.
- מיד, לאחסן את הדגימות ב -20 מעלות צלזיוס.
- חזור על הליך זה עם הריכוזים הנותרים (100 μM עבור המחקר הנוכחי).
- יש לדלל את תרכובת הבדיקה או הרכב לריכוזים הרצויים. הוסף את תרכובת הבדיקה בהגדלת הריכוזים החל מהריכוז הנמוך ביותר.
- בצע עיכוב של גירוי בעקבות השלבים הבאים.
- מוסיפים את החוסם או הרכב לרקמה ודוגרים במשך 10 דקות (נפח: 250 μL עבור רקמת עכבר, 350 μL עבור חולדה TG ו- TNC, ו- 400 μL עבור כל גולגולת חולדה).
הערה: החוסם והריכוז שבהם נעשה שימוש תלויים במטרת המחקר. לדוגמה, 3 μM glibenclamide שימשו לתוצאה המייצגת באיור 5. - לאחר 10 דקות של דגירה, לאסוף דגימה של 200 μL בצינור microcentrifuge עם 50 μL של 10x EIA חיץ. יש להשליך את הנוזל הנותר ולאחסן מיד את הדגימות בטמפרטורה של -20°C.
- מוסיפים את האגוניסט או האגוניסט + החוסם (ראו טבלת חומרים) לרקמה ודוגרים במשך 10 דקות.
הערה: האגוניסט, החוסם והריכוזים שבהם נעשה שימוש תלויים במטרת המחקר. במחקר הנוכחי נעשה שימוש ב-3 מיקרומטר של גליבנקלמיד ו-1 מיקרומטר של קפסאיצין, איור 5. - לאחר 10 דקות של דגירה, לאסוף דגימה של 200 μL בצינור microcentrifuge עם 50 μL של 10x EIA חיץ. יש להשליך את הנוזל הנותר ולאחסן מיד את הדגימות בטמפרטורה של -20°C.
- מוסיפים את החוסם או הרכב לרקמה ודוגרים במשך 10 דקות (נפח: 250 μL עבור רקמת עכבר, 350 μL עבור חולדה TG ו- TNC, ו- 400 μL עבור כל גולגולת חולדה).
- בצע בקרה חיובית עבור הניסוי.
- בעת הצורך, הוסף בקרה חיובית (למשל, 1-10 μM של קפסאיצין, ראה טבלת חומרים) לרקמה בסוף הפרוטוקול ואסוף דגימה של 200 μL בצינור microcentrifuge עם 50 μL של 10x EIA buffer לאחר תקופת דגירה של 10 דקות.
הערה: כדאי לכלול בקרה חיובית כדי להבטיח שההתקנה והרקמות מתפקדות. קפסאיצין 48,49,50 או הגירוי הדה-פולריזציה של אשלגן (40-60 מ"מ של KCl)46,47,49 משמשים באופן שגרתי כדי לגרום לשחרור CGRP מהמערכת הטריגמינווסקולרית. 40-60 mM של KCl SIF מוכן כמו SIF, למעט NaCl מוחלף עבור KCl על בסיס שווה.
- בעת הצורך, הוסף בקרה חיובית (למשל, 1-10 μM של קפסאיצין, ראה טבלת חומרים) לרקמה בסוף הפרוטוקול ואסוף דגימה של 200 μL בצינור microcentrifuge עם 50 μL של 10x EIA buffer לאחר תקופת דגירה של 10 דקות.
6. ניתוח ריכוזי CGRP
- מדוד את כמות ה-CGRP המשתחררת באמצעות ערכת בדיקת אנזימים (EIA) בהתאם לפרוטוקול היצרן (ראה טבלת חומרים).
- מדוד את הצפיפות האופטית ב-410 ננומטר באמצעות פוטומטר צלחת. אם נעשה שימוש בערכת CGRP EIA אחרת, מדוד את הצפיפות האופטית באורך הגל המסופק בפרוטוקול היצרן.
הערה: יש לדלל את הדוגמאות כך שיתאימו לעקומה הסטנדרטית. - בצע ניתוח נתונים.
- הצג את הנתונים כריכוזים מוחלטים או נרמל לשחרור CGRP הבסיסי מהרקמה הספציפית.
תוצאות
טכניקה זו היא כלי לחקור את המנגנונים המולקולריים הקשורים ל-CGRP המעורבים במיגרנה. יש לו את היתרון של הערכת שחרור CGRP מרמות שונות של מערכת trigeminovascular והוא יכול להיות מיושם הן על עכברים מסוג בר וטרנסגניים וחולדות בשילוב עם תרכובות פרמקולוגיות שונות. כאן מוצגים ניסויי ריכוז-תגובה וחסימה מחולדות ותוצאות תגובת ריכוז מעכברים מסוג בר ועכברים מהונדסים.
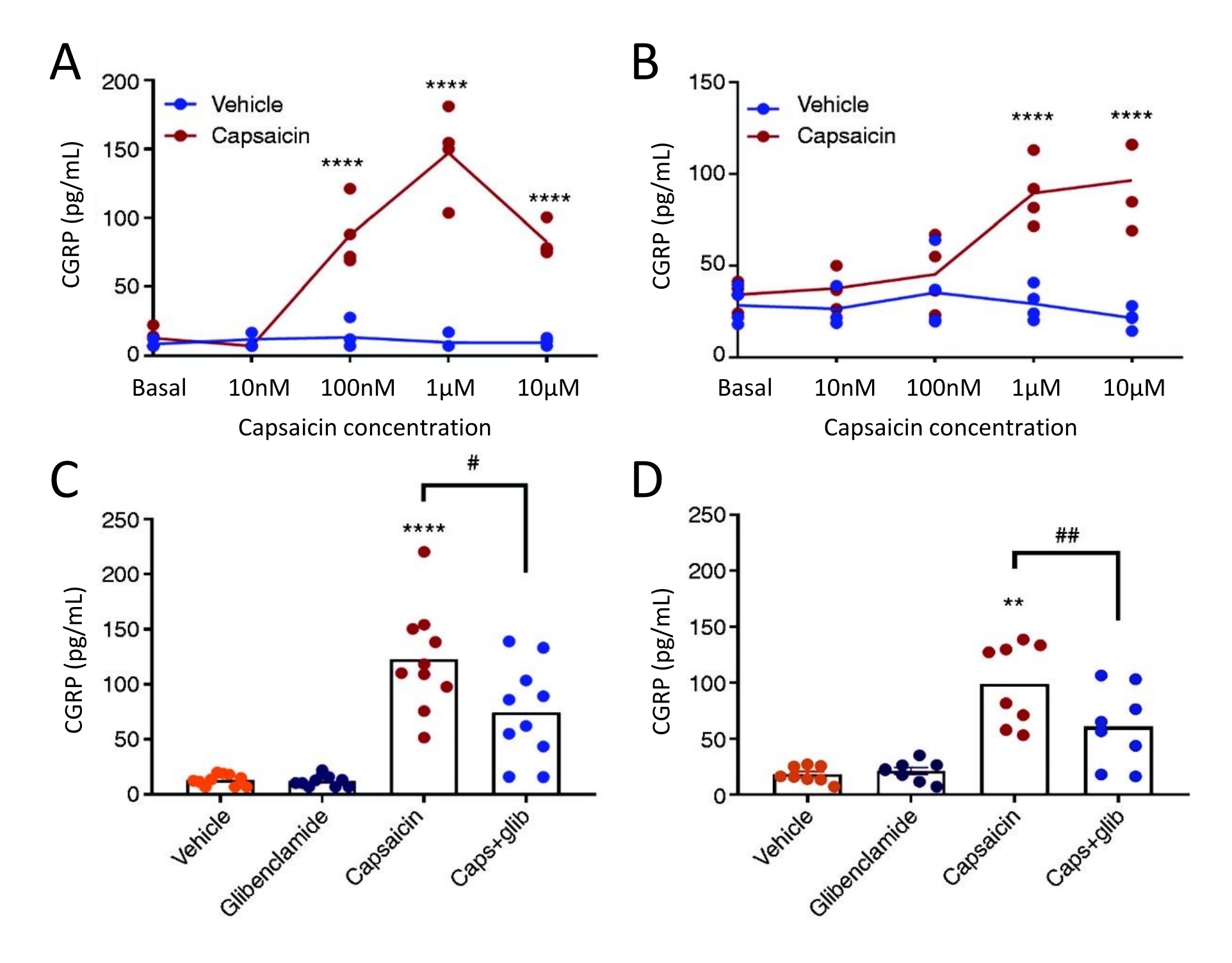
שיטת השחרור של CGRP שימשה כדי לחקור את ההשפעה של מעכב ערוץK ATP גליבנקלמיד על שחרור CGRP מ-TG ודורה מאטר אצל נקבות חולדות אלודינליות טריגמינליות ספונטניות (STA). ראשית, הריכוז האופטימלי של קפסאיצין נמצא באמצעות תכנון מחקר תגובה-ריכוז. חשיפה לקפסאיצין גרמה לשחרור CGRP משמעותי מדורא מאטר ו-TG בהשוואה לרכב (איור 5). בדורה מאטר, השחרור המרבי של CGRP נמצא ב-1 μM של קפסאיצין, וב-TG, השחרור המרבי של CGRP נמצא ב-10 μM של קפסאיצין (איור 5A-B). בהתבסס על ניסויי תגובת הריכוז, 1 μM של קפסאיצין ו-3 μM של גליבנקלמיד שימשו לניסויים חוסמים. גליבנקלמיד לא הראה השפעה על שחרור CGRP בסיסי מדורא מאטר (P = 0.441) ו- TG (P = 0.881) כאשר נותח עם ANOVA51 חד-כיווני. גליבנקלמיד הפחית באופן משמעותי את שחרור ה-CGRP המושרה על-ידי קפסאיצין בדורה מאטר ב-40% (P = 0.031) וב-TG ב-39% (P = 0.003) בהשוואה לקפסאיצין עם הרכב כאשר נותח באמצעות ANOVA חד-כיווני (איור 5C-D)51.
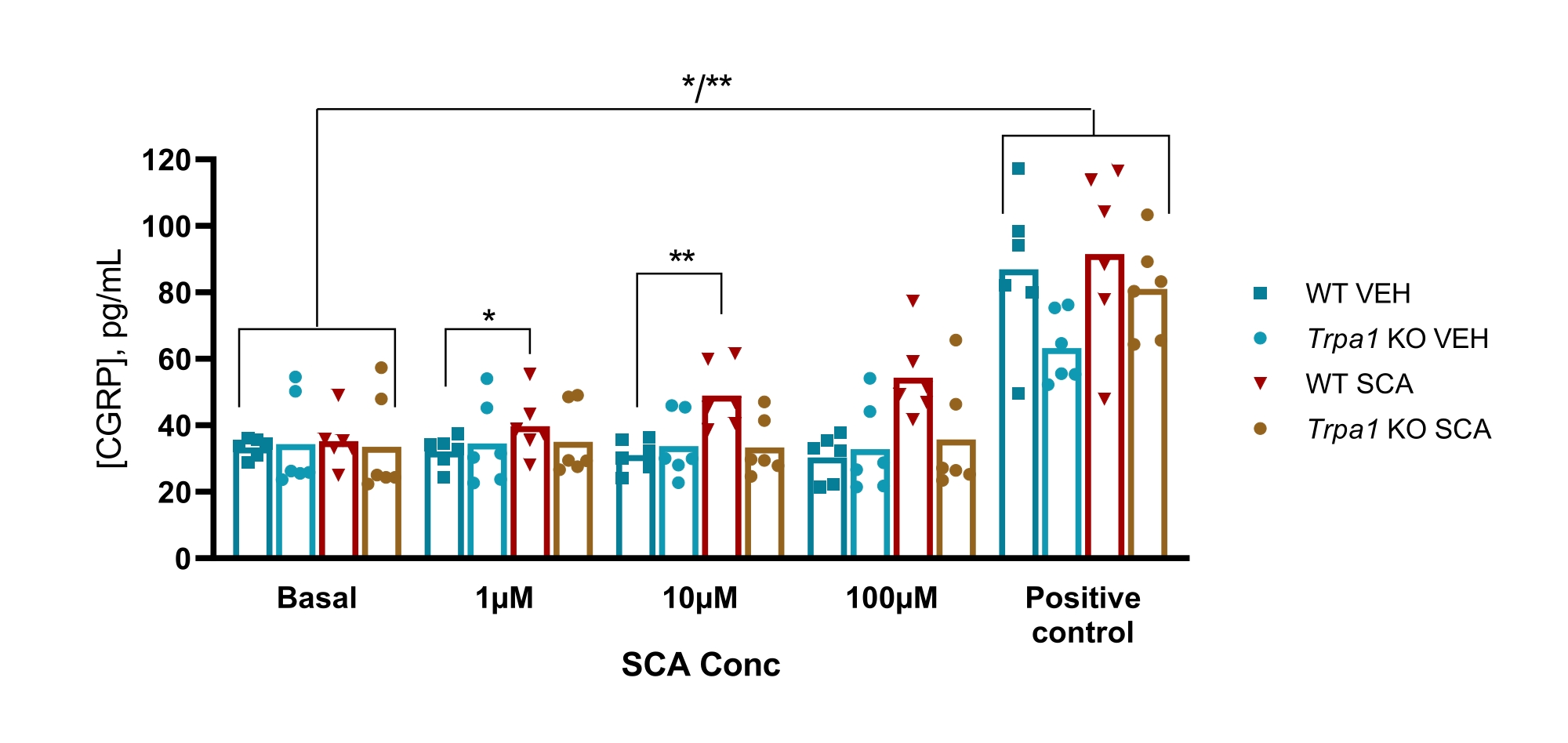
בעכברים, הפרוטוקול שימש לבחינת המעורבות של תעלת היונים הפוטנציאלית של הקולטן החולף אנקירין 1 (TRPA1) במודל עכבר GTN של מיגרנה, שבו רגישות יתר הנגרמת על ידי GTN הייתה תלויה לחלוטין בערוצי TRPA1. נמצא כי אגוניסט TRPA1 supercinnamaldehyde (SCA) משחרר CGRP באופן תלוי מינון מה-TG עם 1 μM, 10 μM ו-100 μM של SCA, וכתוצאה מכך 9% (P = 0.23), 51% (P = 0.011) ו-69% (P = 0.0097) הגדילו את השחרור של CGRP בהשוואה לרכב, בהתאמה כאשר נותחו עם ANOVA דו-כיווני. שחרור זה נעדר ב-TG מעכברי Trpa1 null שבהם חשיפה ל-1 μM, 10 μM ו-100 μM של SCA הביאה ל-11% (P > 0.99), -13% (P > 0.99) ו-9% (P = 0.97) אחוז שינוי בשחרור CGRP בהשוואה לרכב, בהתאמה כאשר נותח עם ANOVA דו-כיווני. הגירוי הבא עם 10 מיקרומטר של קפסאיצין (בקרה חיובית) מראה שכל דגימות הרקמה יכולות לשחרר CGRP (איור 6)50.

איור 1: כריתה שלב אחר שלב של רקמות מחולדות. הפרטים (A-T) מופיעים בסעיף הפרוטוקול (שלב 3.1). אנא לחץ כאן כדי להציג גרסה גדולה יותר של נתון זה.
{kind=link}

איור 2: ניתוח שלב אחר שלב של רקמה מעכברים. הפרטים (A-T) מופיעים בסעיף הפרוטוקול (שלב 3.2). אנא לחץ כאן כדי להציג גרסה גדולה יותר של נתון זה.
{kind=link}

איור 3: שטיפה ודגירה של רקמת מכרסמים. (א-ב) רקמה מבודדת טרייה במיכלי פלסטיק עם SIF. המכלים מכוסים טול כדי לאפשר החלפה קלה של SIF. (C) רקמת חולדה - שני חצאי גולגולת עם דורה מאטר המכסים את הבטנה הפנימית של הגולגולת מונחים על צלחת תרבית 6 בארות. הגרעין הטריגמינלי הימני והשמאלי caudalis במכסי צינורות מיקרוצנטריפוגה נפרדים (שורה עליונה של מכסים). שני הגרעינים הטריגמינליים במכסי צינורות מיקרוצנטריפוגה בודדים (שורה תחתונה של מכסים). (D) רקמת עכבר - הגרעין הטריגמינלי המכיל חלק מגזע המוח במכסה צינור מיקרוצנטריפוגה נפרד (למעלה). שני גרעינים טריגמינליים של עכברים נמצאים במכסה צינור מיקרוצנטריפוגה אחד (למטה). אנא לחץ כאן כדי להציג גרסה גדולה יותר של נתון זה.
{kind=link}

איור 4: תכנוני מחקר לשחרור CGRP. שלושה פרוטוקולים שונים לביצוע ניסויי שחרור CGRP. יש לדלל את התרופות בנוזל אינטרסטיציאלי סינתטי (SIF). (A) גירוי יחיד. (B) גירוי ריכוז-תגובה. (C) עיכוב של גירוי. אנא לחץ כאן כדי להציג גרסה גדולה יותר של נתון זה.
{kind=link}

איור 5: גליבנקלמיד מעכב שחרור CGRP המושרה על-ידי קפסאיצין מחולדות דורה מאטר ומגנגליון טריגמינלי. רמות CGRP מגנגליון טריגמינלי ודורה מאטר שבודדו מנקבות חולדות אלודינליות טריגמינליות ספונטניות (STA) (215-318 גרם) שמקורן במקור באוניברסיטת תומאס ג'פרסון52 נמדדו באמצעות ערכות CGRP EIA אנושיות מסחריות. (א-ב) שחרור CGRP מ-(A) דורה מאטר ו-(B) גנגליון טריגמינלי לאחר הגדלת ריכוזי קפסאיצין (10 ננומטר, 100 ננומטר, 1 מיקרומטר ו-10 מיקרומטר) (n = 4). הנתונים מוצגים כנקודות בודדות ונותחו עם ANOVA דו-כיווני. p < 0.0001 (C-D) רמות של CGRP ששוחררו מ-(C) דורה מאטר ו-(D) גנגליון טריגמינלי לאחר חשיפה של 10 דקות לרכב, 3 μM glibenclamide (glib), 1 μM קפסאיצין ו-1 μM קפסאיצין + 3 μM glib (n = 6-11). הנתונים מוצגים כנקודות בודדות וערכים ממוצעים ונותחו עם ANOVA חד כיווני. *בהשוואה לרכב. #קפסאיצין לעומת קפסאיצין + גליב. #P < 0.05, ** ו##P < 0.01, ****P < 0.0001. הניתוחים בוצעו באמצעות מבחן ההשוואה המרובה של בונפרוני. רמה משמעותית של α = 0.05 שימשה לכל הבדיקות. נתון זה שונה מכריסטנסן ואחרים. 51. אנא לחץ כאן כדי להציג גרסה גדולה יותר של נתון זה.
{kind=link}

איור 6: חשיפה ל-SCA גורמת לשחרור CGRP תלוי TRPA1 מגנגליון טריגמינלי של עכברים. רמות CGRP ששוחררו מגנגליון טריגמינלי שבודד מ-WT ו-Trpa1 null (Trpa1tm1/Dpc)53 עכברים זכרים (8-10 שבועות) נמדדו עם ערכות CGRP EIA של חולדות לאחר חשיפה לסופר-סינמאלדהיד (SCA) ב-1 מיקרומטר, 10 מיקרומטר ו-100 מיקרומטר וקפסאיצין ב-10 מיקרומטר כבקרה חיובית (n = 6). הנתונים מכל עכבר מוצגים כנקודות נפרדות, ופסים מציינים את הערכים הממוצעים. סטטיסטיקה: ההשוואה בין SCA לרכב בכל ריכוז ובין בקרה בסיסית לחיובית בוצעה עם ANOVA דו-כיווני חוזר. רמה משמעותית של α = 0.05. *P < 0.05, **P < 0.01. נתון זה שונה מכריסטנסן ואחרים. 50. אנא לחץ כאן כדי להציג גרסה גדולה יותר של נתון זה.
{kind=link}
Discussion
השיטה המתוארת פותחה בעקבות מחקרים שהראו את החשיבות של CGRP בפתופיזיולוגיה של מיגרנה. הוא מתאים היטב לחקר המנגנונים המעורבים בשחרור CGRP מהמערכת הטריגמינווסקולרית, שהיא חיונית לאיתות כאב באזור הראש. כמות ה-CGRP המתקבלת במודל זה מודדת ישירות את שחרור ה-CGRP מהעצבים הטריגמינליים הפנימיים של דורה מאטר, TG ו-TNC. כמות שחרור ה-CGRP גדולה יותר מבחינה כמותיתמ-45,54 מהשחרור שנמדד בפלזמה לאחר התרמוקו-קרישה בבני אדם, גירוי טריגמינלי בחתול 39,55,56 ובמהלך התקפי מיגרנה 23. הסבר אחד יכול להיות כי CGRP הוא מדולל ומושפל בדם54. עם זאת, יש לציין כי גירוי ישיר עם כימיקלים עשוי להיות עדיף על הפעלה פתופיזיולוגית. יתרונות נוספים הם שניתן לאתר את השחרור משלושה אתרים שונים בתוך המערכת הטריגמינווסקולרית וכי ניתן להשתמש בו יחד עם מניפולציה פרמקולוגית וברקמות ממכרסמים מהונדסים גנטית.
לאחרונה, מודלים פרה-קליניים רבים של מכרסמים מתמקדים במתן סיסטמי של חומרים וקריאות הקשורות לכאב או מיגרנה לאחר מכן באמצעות בדיקת פון פריי57,58, 59,60,61 או סלידה קלה62,63,64. שיטות אלה שימושיות בהבנת התכונות של גרימת כאב ושיכוך כאבים של חומרים שונים. עם זאת, גישות אלה אינן מספקות מידע על רקמות מטרה ספציפיות המעורבות. בשיטה הנוכחית, המערכת הטריגמינובסקולרית מחולקת לשלושה מבנים: הדורה מאטר, TG ו- TNC. הדבר מאפשר חשיפה מקומית של כל מבנה והערכת מיקום הפעולה של חומר מסוים. זה נוצל במחקר משנת 2011, שבו נחקר תפקידן של תעלות סידן מגודרות מתח בחולדות, והעיכוב של תעלות אלה נמצא שונה בשלושת המבנים של המסלול הטריגמינווסקולרי47. כאשר לנתח מבנים של מערכת העצבים, אקסוטומיה היא בלתי נמנעת. אקסוטומיה הוכחה כמשנה את השעתוק של גנים שונים65. שינויי שעתוק אלה איטיים מכדי להשפיע על התוצאות משיטה זו, אך לא ניתן לשלול שינויים בזרחון בהשוואה למצבי in vivo 46. למרות נוירופפטידים כגון CGRP נוצרים בסומה התא, שחרור ופעולה של נוירופפטידים הם בדרך כלל בקצות העצבים המרכזיים או ההיקפיים. לכן, מחקרים של נוירונים שלמים, כולל מסופים, מעניינים כאשר חוקרים שחרור נוירופפטידים. לכן, שיטות של לימוד תרבויות של נוירונים מגרעינים מבודדים הוקמו כדי לשמש מודל של הטרמינלים. עם זאת, תרביות תאים עצביים חשופות למספר בעיות שכן דיסוציאציה מכנית יכולה להרוס נוירונים בתרבית46. מסגרת הזמן הארוכה יותר הקשורה להתרבות תאים משאירה שיטה זו רגישה לשינויים תעתיקיים עקב תנאי אקסוטומיה ותרבית65. יתר על כן, תוספת של גורמי גדילה והתרבות על ציפויי פני השטח שינו את התכונות העצביות כביטוי משדר וקולטן66,67,68,69. בעיות אלה נמנעות כאשר חוקרים גרעינים שלמים שבודדו זה עתה במקום תרביות תאים עצביים.
אחד האתגרים בשיטת השחרור ex vivo CGRP הוא הדיסקציה המדויקת של הרקמה הנדרשת לתוצאות הניתנות לשחזור. דיסקציה מדויקת במיוחד של TNC היא מאתגרת מכיוון שמדובר במבנה בתוך גזע המוח ללא גבולות נראים לעין. יתר על כן, הדורה מאטר שברירית, והסרת המוח צריכה להתבצע בזהירות כדי להבטיח מבנה שלם. מכשולים אלה יכולים לגרום לגודל רקמה משתנה, ולכן, רמות CGRP משתנות של הבסיס והגירוי. עם זאת, וריאציה זו יכולה להיות מוסברת על ידי נורמליזציה לשחרור CGRP הבסיסי. כמו כן יש לציין כי בעת בידוד TNC מעכברים, כל החלק התחתון של גזע המוח מבודד ולא החלק הספציפי יותר המכיל TNC כפי שנעשה בחולדות. באופן כללי, זה יכול להיות יתרון להשתמש ברקמת חולדה, שכן זה מאפשר מדידה של שחרור CGRP מדורא מאטר ודיסקציה מדויקת יותר של TNC. יתר על כן, גודל הרקמה מאפשר גם שימוש בחולדה כבקרת הרכב שלה, שכן חולדה אחת גורמת לשני חצאי גולגולת, שני TGs ושני TNCs כאשר חתיכה אחת של הרקמה משמשת לגירוי החומר והשנייה לרכב. בעת שימוש בעכברים, יש צורך בשני בעלי חיים לניסוי אחד, שכן שני ה-TGs מרוכזים בדגימה אחת, וה-TNCs מנותחים כגזע מוח אחד. לכן, שני TGs וגזע מוח אחד משמשים לגירוי חומרים, ושני TGs וגזע מוח אחד מעכבר אחר משמשים לבקרת רכב. התוצאה היא שימוש בכמות כפולה של עכברים בהשוואה לחולדות כדי לקבל את אותו מספר של שכפולים. כדי להפחית את מספר העכברים שבהם משתמשים, הוצעה שיטה למדידת שחרור CGRP מפרוסות גזע המוח49. זה יתרון כי השיטה שונתה כדי לאפשר את השימוש בעכברים. זה מאפשר שימוש בזני עכברים מהונדסים רבים שכבר זמינים, כלי שימושי ללימוד, למשל, מסלולי איתות. יש לכלול בקרה חיובית בסוף הניסוי כדי להבטיח שהרקמה ששימשה בניסוי תוכל לשחרר CGRP. הבקרה החיובית יכולה להיות קפסאיצין אגוניסט TRPV1 או אשלגן הגירוי הדפולריזציה (KCl), אשר נמצאו כמשחררים CGRP מהמערכת הטריגמינווסקולרית הן בעכברים והן בחולדות 46,47,48,49,50. יתר על כן, השיטה הותאמה גם למדידת שחרורם של פפטידים רלוונטיים אחרים כפפטיד מפעיל אדנילט ציקלאז (PACAP) - פפטיד נוסף בעל עניין רב במחקר מיגרנה70.
השיטה מספקת כלי שימושי לחקר שחרור CGRP מרקמות מטרה ספציפיות בחולדות ובעכברים. זוהי שיטה מהירה יחסית המונעת בעיות הקשורות culturing נוירונים. ניתן לשנות בקלות את פרוטוקול השיטה כדי לחקור את הקשר בין ריכוז לתגובה או עיכוב תגובה על ידי תרכובות פרמקולוגיות שונות. שיטת השחרור ex vivo CGRP היא אחת מכמה שיטות פרה-קליניות שימושיות לחקר התפקיד של CGRP ומנגנונים אחרים הקשורים לשחרור CGRP בפתופיזיולוגיה של מיגרנה.
Disclosures
למחברים אין מה לחשוף.
Acknowledgements
עבודה זו מומנה על ידי קרן קנדיס.
Materials
| Name | Company | Catalog Number | Comments |
| 6-well culture plate | NUNC | 140675 | |
| Calcium chloride dihydrate | Merck | 1.02382.1000 | For SIF buffer |
| Caps for plastic containers | ThermoFisher Scientific | 536617 | |
| Capsaicin | Merck | M2028 | |
| CGRP kits | AH Diagnostics | A05482.96 | |
| CO2 | Strandmøllen | 4.6 | For carbogen gassing of SIF |
| Delicate Bone Trimmer | Fine Science Tools | 16109-14 | |
| Glibenclamide | Tocris | 911 | |
| Glucose | Merck | G7021 | For SIF buffer |
| Guillotine for rats | Scandidact | NS-802 | |
| Magnesium sulfate heptahydrate | Merck | M5921 | For SIF buffer |
| Microcenrifuge tubes + lids/caps | VWR | 700-5239 | |
| Mini Hacksaw | BAHCO | 208 | |
| O2 | Strandmøllen | 4.5 | For carbogen gassing of SIF |
| Pentobarbital | Glostrup pharmacy | NA | Magistral formula |
| Plastic containers | ThermoFisher Scientific | 536455 | |
| Plate photometer - Infinite M200 | Tecan | NA | Infinite M200 is discontinued. A Infinite 200 PRO is available at Tecan. Software: SW Magellan v.6.3 |
| Potassium chloride | Merck | P9333 | For SIF buffer |
| Scissor | Allgaier Instruments | 307-156-170 | |
| Small scissor | Allgaier Instruments | 04-520-115 | |
| Sodium bicarbonate | Merck | S6014 | For SIF buffer |
| Sodium chloride | Merck | S9888 | For SIF buffer |
| Sodium dihydrogen phosphate monohydrate | Merck | 1.06346.1000 | For SIF buffer |
| Sodium gluconate | Merck | S2054 | For SIF buffer |
| Spatula | Bochem Lab Supply | 3018 | |
| Spring scissor | Fine Science Tools | 15024-10 | |
| Sucrose | Merck | 84097 | For SIF buffer |
| Supercinnamaldehyde | Merck | S3322 | |
| Tulle (fabrics) | NA | NA | Bought in the local fabrics store |
References
- GBD 2016 Collaborators. Global, regional, and national incidence, prevalence, and years lived with disability for 328 diseases and injuries for 195 countries, 1990-2016: A systematic analysis for the Global Burden of Disease Study. The Lancet. 390 (10100), 1211-1259 (2017).
- Dodick, D. W. CGRP ligand and receptor monoclonal antibodies for migraine prevention: Evidence review and clinical implications. Cephalalgia: An International Journal of Headache. 39 (3), 445-458 (2019).
- Sacco, S., et al. European headache federation guideline on the use of monoclonal antibodies acting on the calcitonin gene related peptide or its receptor for migraine prevention. The Journal of Headache and Pain. 20 (1), 6 (2019).
- Moreno-Ajona, D., Pérez-Rodríguez, A., Goadsby, P. J. Gepants, calcitonin-gene-related peptide receptor antagonists: what could be their role in migraine treatment. Current Opinion in Neurology. 33 (3), 309-315 (2020).
- Khan, S., Olesen, A., Ashina, M. CGRP, a target for preventive therapy in migraine and cluster headache: Systematic review of clinical data. Cephalalgia: An International Journal of Headache. 39 (3), 374-389 (2019).
- Edvinsson, L., Haanes, K. A., Warfvinge, K., Krause, D. N. CGRP as the target of new migraine therapies - successful translation from bench to clinic. Nature Reviews Neurology. 14 (6), 338 (2018).
- Goadsby, P. J., et al. Pathophysiology of migraine: A disorder of sensory processing. Physiological Reviews. 97 (2), 553-622 (2017).
- Amara, S. G., Jonas, V., Rosenfeld, M. G., Ong, E. S., Evans, R. M. Alternative RNA processing in calcitonin gene expression generates mRNAs encoding different polypeptide products. Nature. 298 (5871), 240-244 (1982).
- Rosenfeld, M. G., et al. Production of a novel neuropeptide encoded by the calcitonin gene via tissue-specific RNA processing. Nature. 304 (5922), 129-135 (1983).
- Edvinsson, L., Goadsby, P. J. Discovery of CGRP in relation to migraine. Cephalalgia: An International Journal of Headache. 39 (3), 331-332 (2019).
- Brain, S. D., Williams, T. J., Tippins, J. R., Morris, H. R., MacIntyre, I. Calcitonin gene-related peptide is a potent vasodilator. Nature. 313 (5997), 54-56 (1985).
- Fisher, L. A., et al. Stimulation of noradrenergic sympathetic outflow by calcitonin gene-related peptide. Nature. 305 (5934), 534-536 (1983).
- Hanko, J., Hardebo, J. E., Kåhrström, J., Owman, C., Sundler, F. Calcitonin gene-related peptide is present in mammalian cerebrovascular nerve fibres and dilates pial and peripheral arteries. Neuroscience Letters. 57 (1), 91-95 (1985).
- Uddman, R., Edvinsson, L., Ekblad, E., Håkanson, R., Sundler, F. Calcitonin gene-related peptide (CGRP): perivascular distribution and vasodilatory effects. Regulatory Peptides. 15 (1), 1-23 (1986).
- Edvinsson, L. Functional role of perivascular peptides in the control of cerebral circulation. Trends in Neurosciences. 8, 126-131 (1985).
- Edvinsson, L., Ekman, R., Jansen, I., Ottosson, A., Uddman, R. Peptide-containing nerve fibers in human cerebral arteries: Immunocytochemistry, radioimmunoassay and in vitro pharmacology. Annals of Neurology. 21 (5), 431-437 (1987).
- Edvinsson, L., Fredholm, B. B., Hamel, E., Jansen, I., Verrecchia, C. Perivascular peptides relax cerebral arteries concomitant with stimulation of cyclic adenosine monophosphate accumulation or release of an endothelium-derived relaxing factor in the cat. Neuroscience Letters. 58 (2), 213-217 (1985).
- Brain, S. D., Grant, A. D. Vascular actions of calcitonin gene-related peptide and adrenomedullin. Physiological Reviews. 84 (3), 903-934 (2004).
- Skofitsch, G., Jacobowitz, D. M. Calcitonin gene-related peptide: Detailed immunohistochemical distribution in the central nervous system. Peptides. 6 (4), 721-745 (1985).
- Suzuki, N., Hardebo, J. E., Owman, C. Origins and pathways of cerebrovascular nerves storing substance P and calcitonin gene-related peptide in rat. Neuroscience. 31 (2), 427-438 (1989).
- Uddman, R., Edvinsson, L., Ekman, R., Kingman, T., McCulloch, J. Innervation of the feline cerebral vasculature by nerve fibers containing calcitonin gene-related peptide: trigeminal origin and co-existence with substance P. Neuroscience Letters. 62 (1), 131-136 (1985).
- Warfvinge, K., Edvinsson, L. Distribution of CGRP and CGRP receptor components in the rat brain. Cephalalgia: An International Journal of Headache. 39 (3), 342-353 (2019).
- Goadsby, P. J., Edvinsson, L., Ekman, R. Vasoactive peptide release in the extracerebral circulation of humans during migraine headache. Annals of Neurology. 28 (2), 183-187 (1990).
- Lassen, L. H., et al. CGRP may play a causative role in migraine. Cephalalgia: An International Journal of Headache. 22 (1), 54-61 (2002).
- Olesen, J., et al. Calcitonin gene-related peptide receptor antagonist BIBN 4096 BS for the acute treatment of migraine. New England Journal of Medicine. 350 (11), 1104-1110 (2004).
- Eftekhari, S., et al. Localization of CGRP, CGRP receptor, PACAP and glutamate in trigeminal ganglion. Relation to the blood-brain barrier. Brain Research. 1600, 93-109 (2015).
- Keller, J. T., Marfurt, C. F. Peptidergic and serotoninergic innervation of the rat dura mater. Journal of Comparative Neurology. 309 (4), 515-534 (1991).
- Edvinsson, L., et al. Innervation of the human middle meningeal artery: immunohistochemistry, ultrastructure, and role of endothelium for vasomotility. Peptides. 19 (7), 1213-1225 (1998).
- Lennerz, J. K., et al. Calcitonin receptor-like receptor (CLR), receptor activity-modifying protein 1 (RAMP1), and calcitonin gene-related peptide (CGRP) immunoreactivity in the rat trigeminovascular system: Differences between peripheral and central CGRP receptor distribution. Journal of Comparative Neurology. 507 (3), 1277-1299 (2008).
- Eftekhari, S., Edvinsson, L. Calcitonin gene-related peptide (CGRP) and its receptor components in human and rat spinal trigeminal nucleus and spinal cord at C1-level. BMC Neuroscience. 12, 112 (2011).
- Eftekhari, S., et al. Differential distribution of calcitonin gene-related peptide and its receptor components in the human trigeminal ganglion. Neuroscience. 169 (2), 683-696 (2010).
- Eftekhari, S., Warfvinge, K., Blixt, F. W., Edvinsson, L. Differentiation of nerve fibers storing CGRP and CGRP receptors in the peripheral trigeminovascular system. The Journal of Pain. 14 (11), 1289-1303 (2013).
- Spitzer, M. J. S., Reeh, P. W., Sauer, S. K. Mechanisms of potassium- and capsaicin-induced axonal calcitonin gene-related peptide release: Involvement of L- and T-type calcium channels and TRPV1 but not sodium channels. Neuroscience. 151 (3), 836-842 (2008).
- Evans, A. R., Nicol, G. D., Vasko, M. R. Differential regulation of evoked peptide release by voltage-sensitive calcium channels in rat sensory neurons. Brain Research. 712 (2), 265-273 (1996).
- Gupta, S., Villalón, C. M. The relevance of preclinical research models for the development of antimigraine drugs: Focus on 5-HT 1B/1D and CGRP receptors. Pharmocology & Therapeutics. 128 (1), 170-190 (2010).
- Williamson, D. J., Hargreaves, R. J., Hill, R. G., Shepheard, S. L. Intravital microscope studies on the effects of neurokinin agonists and calcitonin gene-related peptide on dural vessel diameter in the anaesthetized rat. Cephalalgia: An International Journal of Headache. 17 (4), 518-524 (1997).
- Gupta, S., Bhatt, D. K., Boni, L. J., Olesen, J. Improvement of the closed cranial window model in rats by intracarotid infusion of signalling molecules implicated in migraine. Cephalalgia: An International Journal of Headache. 30 (1), 27-36 (2010).
- Knight, Y. E., Edvinsson, L., Goadsby, P. J. Blockade of calcitonin gene-related peptide release after superior sagittal sinus stimulation in cat: a comparison of avitriptan and CP122, 288. Neuropeptides. 33 (1), 41-46 (1999).
- Goadsby, P. J., Edvinsson, L., Ekman, R. Release of vasoactive peptides in the extracerebral circulation of humans and the cat during activation of the trigeminovascular system. Annals of Neurology. 23 (2), 193-196 (1988).
- Tassorelli, C., Joseph, S. A. Systemic nitroglycerin induces Fos immunoreactivity in brainstem and forebrain structures of the rat. Brain Research. 682 (1-2), 167-181 (1995).
- Ramachandran, R., et al. A naturalistic glyceryl trinitrate infusion migraine model in the rat. Cephalalgia: An International Journal of Headache. 32 (1), 73-84 (2012).
- Hoskin, K. L., Zagami, A. S., Goadsby, P. J. Stimulation of the middle meningeal artery leads to Fos expression in the trigeminocervical nucleus: a comparative study of monkey and cat. Journal of Anatomy. 194, 579-588 (1999).
- Charbit, A. R., Akerman, S., Goadsby, P. J. Comparison of the Effects of Central and Peripheral Dopamine Receptor Activation on Evoked Firing in the Trigeminocervical Complex. Journal of Pharmacology and Experimental Therapeutics. 331 (2), 752-763 (2009).
- Koulchitsky, S., Fischer, M., Messlinger, K. Calcitonin gene-related peptide receptor inhibition reduces neuronal activity induced by prolonged increase in nitric oxide in the rat spinal trigeminal nucleus. Cephalalgia: An International Journal of Headache. 29 (4), 408-417 (2009).
- Ebersberger, A., Averbeck, B., Messlinger, K., Reeh, P. W. Release of substance P, calcitonin gene-related peptide and prostaglandin E2 from rat dura mater encephali following electrical and chemical stimulation in vitro. Neuroscience. 89 (3), 901-907 (1999).
- Eberhardt, M., et al. Calcitonin gene-related peptide release from intact isolated dorsal root and trigeminal ganglia. Neuropeptides. 42 (3), 311-317 (2008).
- Amrutkar, D. V., Ploug, K. B., Olesen, J., Jansen-Olesen, I. Role for voltage gated calcium channels in calcitonin gene-related peptide release in the rat trigeminovascular system. Neuroscience. 172, 510-517 (2011).
- Gupta, S., et al. Evidence for CGRP re-uptake in rat dura mater encephali. British Journal of Pharmacology. 161 (8), 1885-1898 (2010).
- Kageneck, C., Nixdorf-Bergweiler, B. E., Messlinger, K., Fischer, M. J. M. Release of CGRP from mouse brainstem slices indicates central inhibitory effect of triptans and kynurenate. Journal of Headache and Pain. 15 (1), 1-9 (2014).
- Christensen, S. L., et al. CGRP-dependent signalling pathways involved in mouse models of GTN- cilostazol- and levcromakalim-induced migraine. Cephalalgia: An International Journal of Headache. 41 (14), 1413-1426 (2021).
- Christensen, S. L., et al. ATP sensitive potassium (KATP) channel inhibition: A promising new drug target for migraine. Cephalalgia: An International Journal of Headache. 40 (7), 650-664 (2020).
- Oshinsky, M. L., et al. Spontaneous trigeminal allodynia in rats: A model of primary headache. Headache. 52 (9), 1336 (2012).
- Kwan, K. Y., et al. TRPA1 contributes to cold, mechanical, and chemical nociception but is not essential for hair-cell transduction. Neuron. 50 (2), 277-289 (2006).
- Eltorp, C., Jansen-Olesen, I., Hansen, A. J. Release of calcitonin gene-related peptide (CGRP) from guinea pig dura mater in vitro is inhibited by sumatriptan but unaffected by nitric oxide. Cephalalgia: An International Journal of Headache. 20 (9), 838-844 (2000).
- Zagami, A. S., Goadsby, P. J., Edvinsson, L. Stimulation of the superior sagittal sinus in the cat causes release of vasoactive peptides. Neuropeptides. 16 (2), 69-75 (1990).
- Goadsby, P. J., Edvinsson, L. The trigeminovascular system and migraine: studies characterizing cerebrovascular and neuropeptide changes seen in humans and cats. Annals of Neurology. 33 (1), 48-56 (1993).
- Bates, E. A., et al. Sumatriptan alleviates nitroglycerin-induced mechanical and thermal allodynia in mice. Cephalalgia: An International Journal of Headache. 30 (2), 170-178 (2010).
- Pradhan, A. A., et al. Characterization of a novel model of chronic migraine. Pain. 155 (2), 269-274 (2014).
- Mogil, J. S., Pang, D. S. J., Silva Dutra, G. G., Chambers, C. T. The development and use of facial grimace scales for pain measurement in animals. Neuroscience and Biobehavioral Reviews. 116, 480-493 (2020).
- Sotocinal, S. G., et al. The rat grimace scale: A partially automated method for quantifying pain in the laboratory rat via facial expressions. Molecular Pain. 7, 55 (2011).
- Langford, D. J., et al. Coding of facial expressions of pain in the laboratory mouse. Nature Methods. 7 (6), 447-449 (2010).
- Mahmoudi, J., et al. Cerebrolysin attenuates hyperalgesia, photophobia, and neuroinflammation in a nitroglycerin-induced migraine model in rats. Brain Research Bulletin. 140, 197-204 (2018).
- Farajdokht, F., Babri, S., Karimi, P., Mohaddes, G. Ghrelin attenuates hyperalgesia and light aversion-induced by nitroglycerin in male rats. Neuroscience Letters. 630, 30-37 (2016).
- Kuburas, A., et al. PACAP Induces Light Aversion in Mice by an Inheritable Mechanism Independent of CGRP. Journal of Neuroscience. 41 (21), 4697-4715 (2021).
- Buschmann, T., et al. Expression of Jun, Fos, and ATF-2 proteins in axotomized explanted and cultured adult rat dorsal root ganglia. Neuroscience. 84 (1), 163-176 (1998).
- Lee, Y. J., Zachrisson, O., Tonge, D. A., McNaughton, P. A. Upregulation of bradykinin B2 receptor expression by neurotrophic factors and nerve injury in mouse sensory neurons. Molecular and Cellular Neuroscience. 19 (2), 186-200 (2002).
- Hari, A., Djohar, B., Skutella, T., Montazeri, S. Neurotrophins and extracellular matrix molecules modulate sensory axon outgrowth. International Journal of Developmental Neuroscience. 22 (2), 113-117 (2004).
- Skoff, A. M., Resta, C., Swamydas, M., Adler, J. E. Nerve Growth Factor (NGF) and Glial Cell Line-Derived Neurotrophic Factor (GDNF) regulate substance P release in adult spinal sensory neurons. Neurochemical Research. 28 (6), 847-854 (2003).
- Lindsay, R. M., Harmar, A. J. Nerve growth factor regulates expression of neuropeptide genes in adult sensory neurons. Nature. 337 (6205), 362-364 (1989).
- Edvinsson, J. C. A., et al. Differences in pituitary adenylate cyclase-activating peptide and calcitonin gene-related peptide release in the trigeminovascular system. Cephalalgia: An International Journal of Headache. 40 (12), 1296-1309 (2020).
Reprints and Permissions
Request permission to reuse the text or figures of this JoVE article
Request PermissionExplore More Articles
This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. All rights reserved