
Method Article
Ex vivo Freisetzung von Calcitonin-Gen-verwandtem Peptid aus dem trigeminovaskulären System bei Nagetieren
In diesem Artikel
Zusammenfassung
Das vorliegende Protokoll beschreibt das ex vivo Calcitonin Gene-Related Peptide (CGRP) Freisetzungsmodell und die Strategie zur Quantifizierung der Wirkung pharmakologischer Wirkstoffe auf die Menge an CGRP, die aus dem trigeminovaskulären System bei Nagetieren freigesetzt wird.
Zusammenfassung
Calcitonin Gene-Related Peptide (CGRP) wurde erstmals in den 1980er Jahren als Spleißvariante aus dem Calcitonin-Gen entdeckt. Seit seiner Entdeckung ist seine Rolle in der Pathophysiologie der Migräne gut etabliert, zunächst durch seine starken gefäßerweiternden Eigenschaften und später durch seine Anwesenheit und Funktion als Neurotransmitter im sensorischen trigeminovaskulären System. Die Migräne-auslösende Fähigkeit von CGRP unterstützte die Pharmaindustrie bei der Entwicklung monoklonaler Antikörper und Antagonisten, die die Wirkung von CGRP hemmen. Ein neues Behandlungsparadigma hat sich bei der prophylaktischen Behandlung von Migräne bewährt. Eines der nützlichen Werkzeuge, um die Mechanismen der Migräne besser zu verstehen, ist das Ex-vivo-Modell der CGRP-Freisetzung aus dem trigeminovaskulären System. Es handelt sich um eine relativ einfache Methode, die mit verschiedenen pharmakologischen Instrumenten eingesetzt werden kann, um Know-how für die Weiterentwicklung neuer wirksamer Migränebehandlungen zu erlangen. Das vorliegende Protokoll beschreibt ein CGRP-Freisetzungsmodell und die Technik zur Quantifizierung der Wirkung pharmakologischer Wirkstoffe auf die Menge an CGRP, die aus dem trigeminovaskulären System bei Nagetieren freigesetzt wird. Ein Verfahren, das den experimentellen Ansatz von der Euthanasie bis zur Messung des Proteingehalts beschreibt, wird zur Verfügung gestellt. Die essentielle Isolierung des Trigeminusganglions und des Trigeminuskerns caudalis sowohl von Mäusen als auch von Ratten sowie die Herstellung von Rattendura mater werden ausführlich beschrieben. Darüber hinaus werden repräsentative Ergebnisse beider Spezies (Ratten und Mäuse) vorgestellt. Die Technik ist ein Schlüsselwerkzeug, um die molekularen Mechanismen der Migräne-Pathophysiologie unter Verwendung verschiedener pharmakologischer Verbindungen und genetisch veränderter Tiere zu untersuchen.
Einleitung
Migräne ist eine neurologische Erkrankung, von der laut WHO schätzungsweise mehr als 1 Milliarde Menschen betroffen sind unddie weltweit eine der Hauptursachen für Behinderungen ist1. Somit hat Migräne erhebliche Auswirkungen sowohl auf die Patienten als auch auf die Gesellschaft. Trotz des jüngsten klinischen Erfolgs von CGRP-antagonisierenden Medikamenten benötigt ein großer Teil der Patienten verbesserte Behandlungsmöglichkeiten 2,3,4,5. Die Aufklärung der Migräne-Pathophysiologie, die zu neuartigen wirksamen Behandlungen führt, ist erforderlich. Die Signalübertragung innerhalb des trigeminovaskulären Systems, bestehend aus den Hirnhäuten, den Trigeminusganglien (TG) und dem Trigeminuskern caudalis (TNC), ist für die Pathophysiologie der Migräne von zentraler Bedeutung 6,7.
Das 37-Aminosäuren-Neuropeptid Calcitonin Gene-Related Peptide (CGRP) wurde erstmals in den frühen 1980er Jahren entdeckt, als Amara und Mitarbeiter zeigten, dass das primäre RNA-Transkript des Calcitonin-Gens verarbeitet werden kann, um mRNA zu erhalten, die zusätzlich zu Calcitonin 8,9 für CGRP kodiert. Die anschließende Forschung deutete auf eine Verbindung zur Migräne-Pathophysiologiehin 10. CGRP ist ein Neurotransmitter mit starken gefäßerweiternden Eigenschaften 11,12,13,14,15,16,1 7 und ist im zentralen und peripheren Nervensystem weit verbreitet 13,14,18,19,20,21,22 . Die Beteiligung von CGRP an Migräne wurde durch die Entdeckung erhöhter CGRP-Spiegel in der exzerebralen Zirkulation während Migräneattacken beim Menschen unterstrichen23 und dass die Infusion von CGRP bei Patienten migräneähnliche Schmerzen verursacht24. Zwei Jahre später wurde die erste Proof-of-Concept-Studie zur Wirksamkeit des CGRP-Antagonisten Olcegepant bei der Behandlung von Migräne veröffentlicht25.
CGRP ist im trigeminovaskulären System reichlich vorhanden, wie in den TG21,26 gezeigt, sensorischen Nervenfasern, die die Dura mater27,28,29 und TNC30 innervieren. Im trigeminovaskulären System findet sich CGRP in den kleinen bis mittelgroßen Neuronen der TG, in unmyelinisierten C-Fasern, und wird in fast 50% der neuronalen Population der TG exprimiert. Der CGRP-Rezeptor wird hauptsächlich in größeren Neuronen exprimiert und findet sich in myelinisierten Aδ-Fasern31,32. CGRP wird bei chemischer oder elektrischer Stimulation aus Neuronen freigesetzt33,34. Untersuchungen von Signalwegen, die zur Freisetzung von CGRP führen, und der Ort dieser Aktivierung sind entscheidend für das Verständnis der Pathophysiologie von Migräne. In den letzten 5 Jahrzehnten haben präklinische Studien dazu beigetragen, umfassende Erkenntnisse über Migräne-bezogene Signalwege zu gewinnen und neue Therapien zu entwickeln35. Viele Methoden, die die vaskuläre und neurogene Beteiligung berücksichtigen, wurden modifiziert und in der Migräneforschung angewendet. In-vivo- und In-vitro-Modelle der arteriellen Reaktionen auf biologische Verbindungen oder pharmakologische Behandlungen17,36,37 und die elektrische Nervenstimulation38,39 können erwähnt werden. Darüber hinaus können aktivierte Neuronen im TNC durch c-Fos-Expression 40,41,42 und elektrophysiologische Aufnahmen in diesem Bereich 43,44 nachgewiesen werden. Beide Methoden messen nozizeptive Signale, die vom Kopf an das Gehirn übertragen werden, z.B. Dura mater. Die Verwendung von nur einem präklinischen Modell stellt nicht das vollständige Bild der Migräne-Pathophysiologie dar. Daher ist es wichtig, verschiedene Modelle zu kombinieren, die so viele Aspekte der Migräne-Pathophysiologie wie möglich abdecken. Die Weiterentwicklung neuer Modelle wird verschiedene Aspekte der Migränemechanismen abdecken, und mit der Zeit wird das Geheimnis der Migräne-Pathophysiologie aufgedeckt werden.
Hier wird ein detailliertes Protokoll der CGRP-Freisetzungsmethode vorgestellt, die ex vivo in isolierten TG und TNC von Mäusen nach chemischer Stimulation durchgeführt wurde. Die CGRP-Freisetzung kann auch in der Dura mater von Ratten untersucht werden. So wird im Versuchsprotokoll für Ratten die Dura mater zusammen mit TG und TNC beschrieben. Die Grundlage für die CGRP-Freisetzungsmethode wurde erstmals 1999 beschrieben, als Ebersberger und Mitarbeiter bahnbrechende Forschungen durchführten und herausfanden, dass CGRP nach chemischer und elektrischer Stimulation von duralen Afferenzen bei Ratten aus der Dura mater freigesetzt wurde45. Später wurde dieser Ansatz auf die CGRP-Freigabe von der TG46 und der TNC47 erweitert. Anschließend wurde die Methode modifiziert, um sie auf TG und TNC in Mäusen anzuwenden. Bisher war die Freisetzung von CGRP aus der Dura mater bei Mäusen eine Herausforderung.
Protokoll
Alle Tierpflege- und Versuchsverfahren wurden in Übereinstimmung mit dem Leitfaden der Europäischen Gemeinschaft für die Pflege und Verwendung von Tieren (2010/63/EU) durchgeführt. Männliche C57BL/6JBomTac-Mäuse im Alter von 10 Wochen und männliche Sprague-Dawley-Ratten im Alter von 10 Wochen wurden verwendet, um dieses Protokoll zu demonstrieren.
1. Herstellung der synthetischen interstitiellen Flüssigkeit
- Synthetische interstitielle Flüssigkeit (SIF) nach folgendem Rezept herstellen: 108 mM NaCl, 3,48 mM KCl, 3,50 mM MgSO4, 26 mMNaHCO3, 11,70 mM NaH2PO4, 1,50 mM CaCl2, 9,60 mM Na-Gluconat, 5,50 mM Glucose und 7,60 mM Saccharose (siehe Materialtabelle).
HINWEIS: SIF kann je nach Stimulationsziel variiert werden, z. B. kann eine kalziumfreie Lösung verwendet werden, wenn Kalziumkanäle untersucht werden. - Stellen Sie den pH-Wert auf 7,4 ein und stabilisieren Sie den pH-Wert durch Carbogen-Begasung (5% CO2 und 95%O2)46.
2. Euthanasie
- Betäuben Sie erwachsene Mäuse und Ratten mit einer Mischung aus 70% CO2 und 30%O2. Enthaupten Sie Mäuse mit einer Schere und Ratten mit einer Guillotine (siehe Materialtabelle).
HINWEIS: Verwenden Sie eine Sorte und ein Alter, die dem Ziel der Forschung entsprechen. Sowohl Männchen als auch Weibchen können in diesem Modell verwendet werden. - Trennen Sie den Kopf vom Körper auf der C3-C4-Ebene des Rückenmarks.
HINWEIS: Euthanasie kann auch mit einer intraperitonealen Injektion von Pentobarbital (100-150 mg / kg) durchgeführt werden.
3. Sektion
- Bereiten Sie das Rattengewebe nach den folgenden Schritten vor.
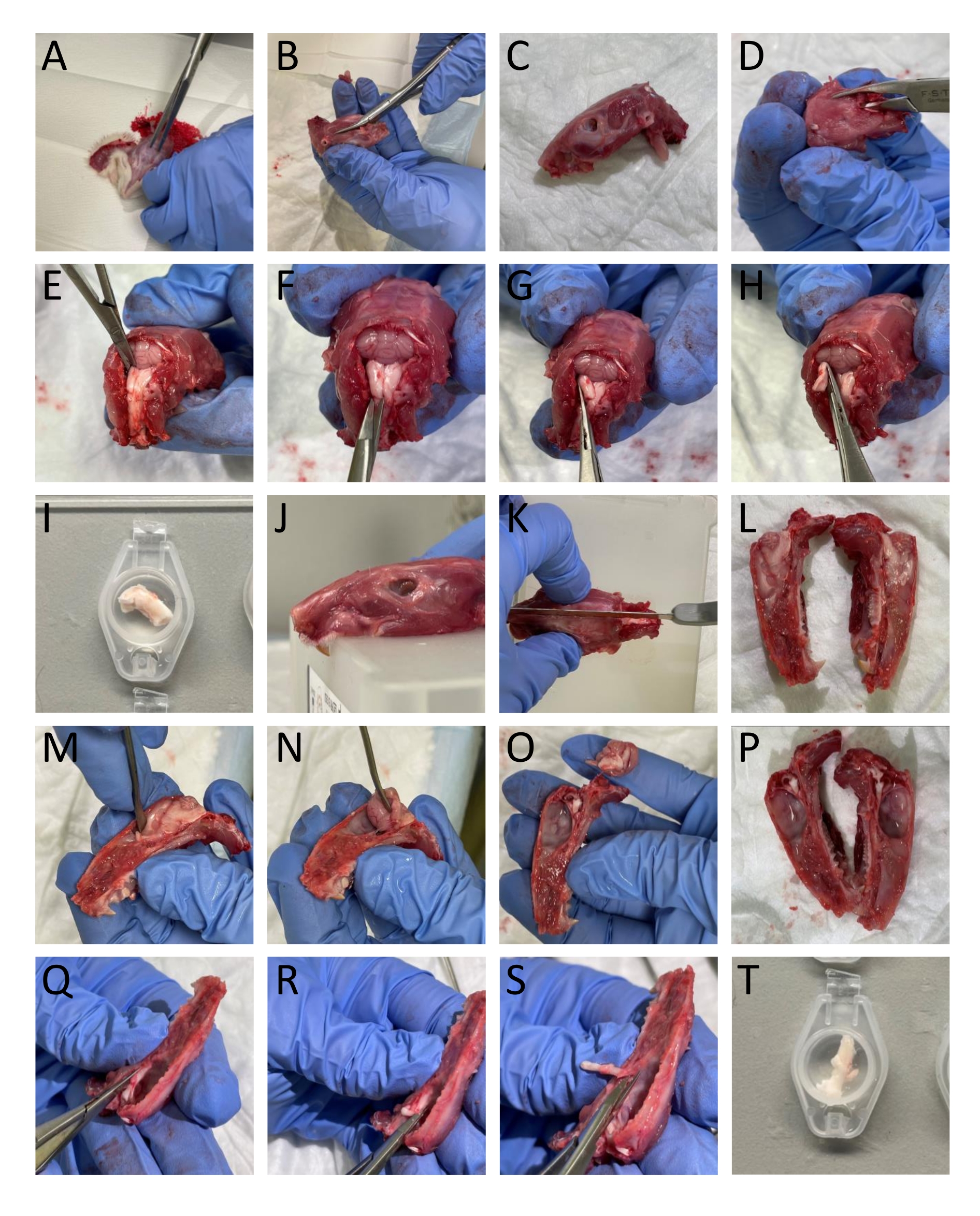
- Entfernen Sie die Haut und den Muskel um Kopf und Hals mit einer Schere (Abbildung 1A).
- Verwenden Sie einen Knochentrimmer und eine Schere, um den Unterkiefer vom Kopf zu trennen (Abbildung 1B-C).
- Öffnen Sie das Rückenmark, indem Sie einen Knochentrimmer kaudal in den dorsalen Teil der Wirbel einführen und den dorsalen Teil der Wirbel entfernen, um das Rückenmark und den Hirnstamm freizulegen (Abbildung 1D).
- Schneiden Sie den kaudalen Teil des Schädels an den Rändern des Hinterhauptbeins und des interparietalen Knochens auf, um diese Knochenstrukturen zu entfernen, die das Kleinhirn freilegen (Abbildung 1E).
HINWEIS: Es ist wichtig, den Hirnstamm und das Rückenmark beim Schneiden und Entfernen der Wirbel nicht zu beschädigen. - Isolieren Sie das TNC (Sp5C), das ca. 13-16 mm von der Bregma entfernt auf jeder Seite kaudal verläuft, indem Sie den dorsolateralen Teil des Hirnstamms mit einer Federschere abschneiden. Tauchen Sie den TNC auf der linken und rechten Seite in SIF ein (Abbildung 1E-I).
HINWEIS: Die Beschreibung bezieht sich auf erwachsene Ratten. - Schneiden Sie den Kopf in der Mitte sagital, um den Schädel mit einer Säge in zwei Teile zu teilen (Abbildung 1J-L).
- Entfernen Sie das Gehirn vorsichtig, ohne die am Schädel befestigte Dura mater mit einem Spatel zu berühren und den Trigeminusnerv dort zu schneiden, wo er in den Hirnstamm eintritt (Abbildung 1M-P).
- Um die TG zu isolieren, schneiden Sie sie ab, einschließlich ihrer Äste an den visuellen Rändern. Schneiden Sie den Unterkieferast dort ab, wo er in das Foramen ovale eintritt. Schneiden Sie die Augen- und Oberkieferäste ab, die in den Schädel eintreten, da sie nicht makroskopisch geteilt sind. Entfernen Sie beim Sezieren der TG die Dura mater, die die TG bedeckt (Abbildung 1Q-T).
- Tauchen Sie die Schädelhälften und die TGs in SIF.
- Bereiten Sie das Mausgewebe vor, indem Sie die folgenden Schritte ausführen.
- Entfernen Sie Haut und Muskeln um Kopf und Hals mit einer kleinen Schere (Abbildung 2A-B).
- Öffnen Sie das Rückenmark, indem Sie eine kleine Schere kaudal in den dorsalen Teil der Wirbel einführen und den dorsalen Teil der Wirbel entfernen, um das Rückenmark und den Hirnstamm freizulegen (Abbildung 2C).
HINWEIS: Es ist wichtig, das Rückenmark beim Schneiden und Entfernen der Wirbel nicht zu beschädigen. - Schneiden Sie den Schädel an den Rändern der Hinterhaupts- und Hälfteknochen auf, um diese Knochenstrukturen zu entfernen, die das Kleinhirn freilegen (Abbildung 2D-F).
- Schneiden Sie dann den Scheitelbein in der Mitte des Sagitals und entfernen Sie den Knochen, um das Großhirn freizulegen (Abbildung 2G-I).
- Entfernen Sie das Kleinhirn vorsichtig mit einem Spatel, um den Hirnstamm freizulegen (Abbildung 2J).
- Isolieren Sie den TNC-haltigen Teil des Hirnstamms mit einer Federschere (Abbildung 2K-N). Tauchen Sie den Hirnstamm mit TNC in SIF ein (Abbildung 2O).
- Entfernen Sie das Gehirn und schneiden Sie den Trigeminusnerv dort, wo er in den Hirnstamm eintritt (Abbildung 2P-Q).
- Um die TG zu isolieren, schneiden Sie sie ab, einschließlich ihrer Äste an den visuellen Rändern. Schneiden Sie den Unterkieferast dort ab, wo er in das Foramen ovale eintritt. Schneiden Sie die Augen- und Oberkieferäste ab, die in den Schädel eintreten, da sie nicht makroskopisch geteilt sind. Entfernen Sie beim Sezieren der TG die Dura mater, die die TG bedeckt (Abbildung 2R-S).
- Tauchen Sie TGs in SIF ein (Abbildung 2T).
4. Waschen
- Waschen Sie Schädelhälften, TNCs und TGs 30 Minuten lang in SIF, während Sie das SIF alle 5 Minuten bei Raumtemperatur ersetzen.
HINWEIS: Die Waschschritte können durchgeführt werden, während das Gewebe in Kunststoffbehältern mit Tülldeckel aufbewahrt wird, um einen einfachen SIF-Austausch zu ermöglichen (Abbildung 3A-B). - Bereiten Sie das Rattengewebe nach den folgenden Schritten vor.
- Überführen Sie TNC-Hälften in separate Mikrozentrifugenröhrchenkappen mit 350 μL SIF (Abbildung 3C).
- Übertragen Sie die TGs auf Mikrozentrifugenröhrchenkappen - ein TG pro Mikrozentrifugenröhrchenkappe mit 350 μL SIF (Abbildung 3C).
- Legen Sie die Schädelhälften auf Plattformen aus Ton oder einer 6-Well-Kulturplatte und füllen Sie den Schädel mit 400 μL SIF (Abbildung 3C).
- Bereiten Sie das Mausgewebe vor, indem Sie die folgenden Schritte ausführen.
- Übertragen Sie den Hirnstamm mit TNC in eine Mikrozentrifugenröhrchenkappe mit 250 μL SIF (Abbildung 3D).
- Die beiden TGs werden in einen Mikrozentrifugenverschluss mit 250 μL SIF gegeben (Abbildung 3D).
HINWEIS: Legen Sie zwei TGs pro Kappe des Mikrozentrifugenröhrchens ein, wenn Sie Gewebe von Mäusen verwenden.
- Rattenschädel und Mikrozentrifugenröhrchenkappen mit Ratten- und Mausgewebe in einen befeuchteten Inkubator bei 37 °C legen. Ersetzen Sie SIF mit einer Pipette alle 5 Minuten für 20 Minuten.
HINWEIS: Es ist wichtig, das Gewebe beim Hinzufügen und Entfernen von SIF nicht zu berühren.
5. Drogentests
- Bestimmen Sie die basalen CGRP-Freisetzungsniveaus.
- Nach der letzten Wäsche 250 μL SIF in Maus TG und TNC geben. Geben Sie 350 μl zu Ratten-TG und TNC und 400 μl zu jedem Rattenschädel.
- Nach 10 Minuten Inkubation sammeln Sie 200 μL der Probe in einem Mikrozentrifugenröhrchen und fügen Sie 50 μL 10x EIA-Puffer hinzu (im Lieferumfang des CGRP-Enzymimmunoassay-Kits enthalten, siehe Materialtabelle), um die Messung der basalen CGRP-Freisetzung zu ermöglichen (Schritt 6). Entsorgen Sie die verbleibende Flüssigkeit.
HINWEIS: Die Inkubationszeit muss für alle Proben gleich sein. - Lagern Sie die Proben sofort bei -20 °C.
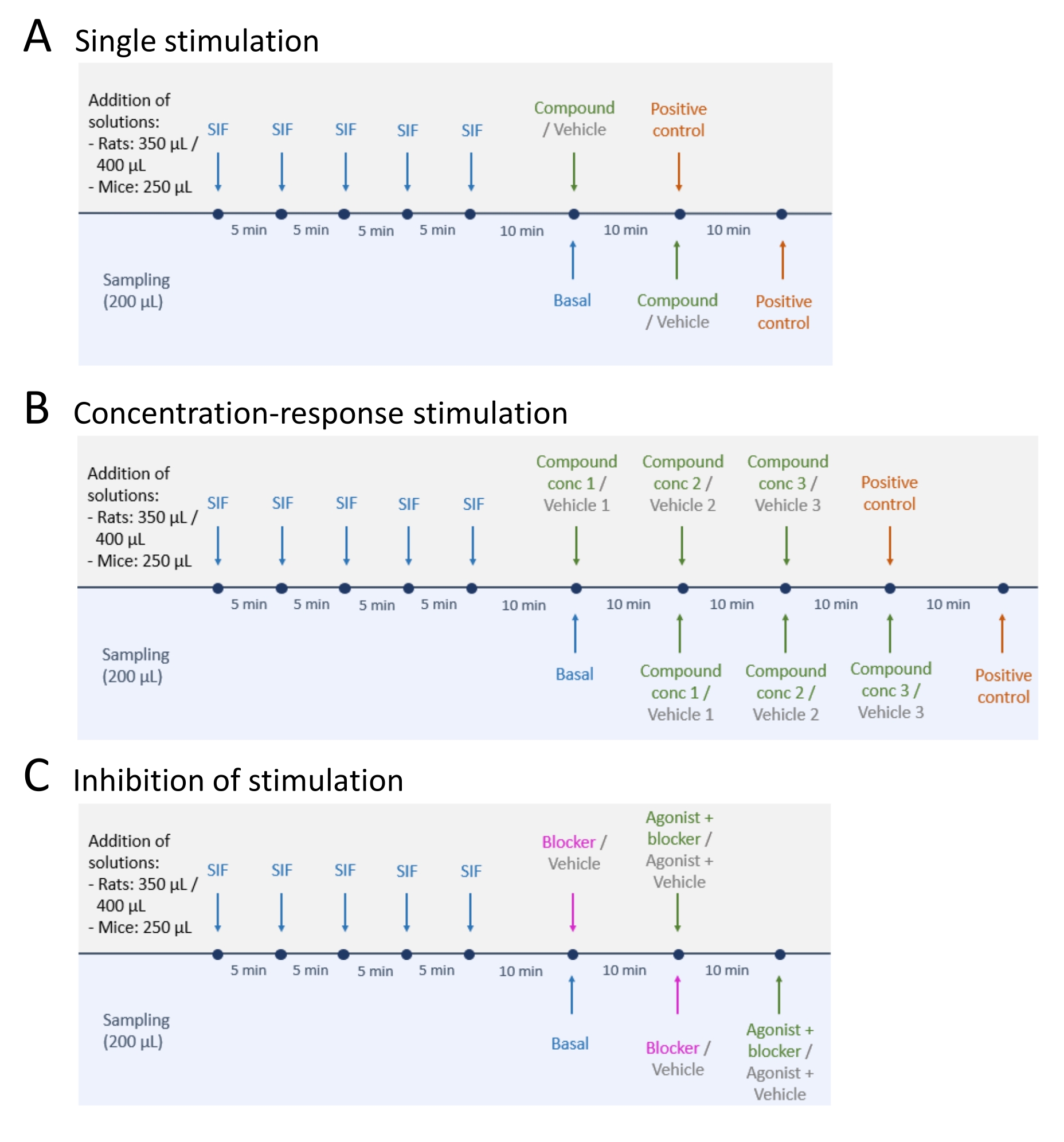
- Nach der Probenahme der basalen CGRP-Freisetzungswerte ist eine der drei Methoden anzuwenden: (A) Einmalige Stimulation (Schritt 5.2); (B) Konzentrations-Wirkungs-Stimulation (Schritt 5.3); und (C) Hemmung der Stimulation (Schritt 5.4) (Abbildung 4).
ANMERKUNG: Die verwendete Testverbindung und die verwendeten Konzentrationen hängen vom Ziel der Studie ab.
- Führen Sie eine einzelne Stimulation durch, indem Sie die folgenden Schritte ausführen.
- Geben Sie die Testsubstanz oder das Vehikel in das Gewebe und lassen Sie es 10 Minuten stehen (Volumen: 250 μl für TG und TNC der Maus, 350 μl für TG und TNC der Ratte und 400 μl für jeden Rattenschädel).
- Nach 10 Minuten Inkubation werden 200 μl der Probe in einem Mikrozentrifugenröhrchen mit 50 μl 10x EIA-Puffer gesammelt. Die restliche Flüssigkeit wird entsorgt und die Proben sofort bei -20 °C gelagert.
HINWEIS: Die Inkubationszeit muss für alle Proben gleich sein.
- Führen Sie eine Konzentrations-Reaktions-Stimulation durch, indem Sie die folgenden Schritte ausführen.
- Die Prüfsubstanz oder das Prüfvehikel wird auf die gewünschten Konzentrationen verdünnt. Die Prüfsubstanz wird in aufsteigenden Konzentrationen, beginnend mit der niedrigsten Konzentration, zugegeben.
ANMERKUNG: Die verwendete Testverbindung und die verwendeten Konzentrationen hängen vom Ziel der Studie ab. Zum Beispiel wurden 1 μM, 10 μM und 100 μM Superzimtaldehyd für die vorliegende Studie verwendet. - Die niedrigste Konzentration (1 μM für die vorliegende Studie) der Testverbindung und des entsprechenden Vehikels wird zu zwei identischen Gewebepräparaten gegeben und 10 Minuten lang inkubiert (Volumen: 250 μl für Mausgewebe, 350 μl für TG und TNC der Ratte und 400 μl für jeden Rattenschädel).
- Nach 10 Minuten Inkubation werden 200 μl der Probe in einem Mikrozentrifugenröhrchen mit 50 μl 10x EIA-Puffer gesammelt.
- Die verbleibende Flüssigkeit wird verworfen und die zweitniedrigste Konzentration (10 μM für die vorliegende Studie) in das Gewebe gegeben.
- Lagern Sie die Proben sofort bei -20 °C.
- Wiederholen Sie diesen Vorgang mit den verbleibenden Konzentrationen (100 μM für die vorliegende Studie).
- Die Prüfsubstanz oder das Prüfvehikel wird auf die gewünschten Konzentrationen verdünnt. Die Prüfsubstanz wird in aufsteigenden Konzentrationen, beginnend mit der niedrigsten Konzentration, zugegeben.
- Führen Sie die Hemmung der Stimulation durch, indem Sie die folgenden Schritte ausführen.
- Fügen Sie den Blocker oder das Vehikel dem Gewebe hinzu und inkubieren Sie für 10 Minuten (Volumen: 250 μL für Mausgewebe, 350 μL für Ratten-TG und TNC und 400 μL für jeden Rattenschädel).
HINWEIS: Der verwendete Blocker und die verwendete Konzentration hängen vom Ziel der Studie ab. Zum Beispiel wurde 3 μM Glibenclamid für das repräsentative Ergebnis in Abbildung 5 verwendet. - Nach 10 Minuten Inkubation wird eine 200-μl-Probe in einem Mikrozentrifugenröhrchen mit 50 μl 10-fachem EIA-Puffer entnommen. Die restliche Flüssigkeit wird entsorgt und die Proben sofort bei -20 °C gelagert.
- Geben Sie den Agonisten oder Agonisten + Blocker (siehe Materialtabelle) in das Gewebe und inkubieren Sie für 10 min.
HINWEIS: Der verwendete Agonist, Blocker und die verwendeten Konzentrationen hängen vom Ziel der Studie ab. In der vorliegenden Studie wurden 3 μM Glibenclamid und 1 μM Capsaicin verwendet, Abbildung 5. - Nach 10 Minuten Inkubation wird eine 200-μl-Probe in einem Mikrozentrifugenröhrchen mit 50 μl 10-fachem EIA-Puffer entnommen. Die restliche Flüssigkeit wird entsorgt und die Proben sofort bei -20 °C gelagert.
- Fügen Sie den Blocker oder das Vehikel dem Gewebe hinzu und inkubieren Sie für 10 Minuten (Volumen: 250 μL für Mausgewebe, 350 μL für Ratten-TG und TNC und 400 μL für jeden Rattenschädel).
- Führen Sie eine Positivkontrolle für das Experiment durch.
- Gegebenenfalls wird am Ende des Protokolls eine Positivkontrolle (z. B. 1-10 μM Capsaicin, siehe Materialtabelle) in das Gewebe gegeben und nach einer Inkubationszeit von 10 Minuten eine 200-μl-Probe in einem Mikrozentrifugenröhrchen mit 50 μl 10x EIA-Puffer entnommen.
HINWEIS: Es ist vorteilhaft, eine Positivkontrolle einzuschließen, um sicherzustellen, dass das Setup und das Gewebe funktionieren. Capsaicin 48,49,50 oder der depolarisierende Stimulus von Kalium (40-60 mM KCl)46,47,49 werden routinemäßig verwendet, um die Freisetzung von CGRP aus dem trigeminovaskulären System zu bewirken. 40-60 mM KCl SIF werden als SIF hergestellt, mit der Ausnahme, dass NaCl auf äquimolarer Basis gegen KCl ausgetauscht wird.
- Gegebenenfalls wird am Ende des Protokolls eine Positivkontrolle (z. B. 1-10 μM Capsaicin, siehe Materialtabelle) in das Gewebe gegeben und nach einer Inkubationszeit von 10 Minuten eine 200-μl-Probe in einem Mikrozentrifugenröhrchen mit 50 μl 10x EIA-Puffer entnommen.
6. Analyse der CGRP-Konzentrationen
- Messen Sie die freigesetzte Menge an CGRP mit einem Enzymimmunoassay-Kit (EIA) gemäß dem Protokoll des Herstellers (siehe Materialtabelle).
- Die optische Dichte bei 410 nm wird mit einem Plattenphotometer gemessen. Wenn ein anderes CGRP-EIA-Kit verwendet wird, messen Sie die optische Dichte bei der Wellenlänge, die im Protokoll des Herstellers angegeben ist.
HINWEIS: Die Proben müssen verdünnt werden, um der Standardkurve zu entsprechen. - Führen Sie eine Datenanalyse durch.
- Präsentieren Sie die Daten entweder als absolute Konzentrationen oder normalisieren Sie sie auf die basale CGRP-Freisetzung aus dem spezifischen Gewebe.
Ergebnisse
Diese Technik ist ein Werkzeug, um die CGRP-bezogenen molekularen Mechanismen zu untersuchen, die an Migräne beteiligt sind. Es hat den Vorteil, die CGRP-Freisetzung aus verschiedenen Ebenen des trigeminovaskulären Systems zu bewerten und kann sowohl bei Wildtyp- als auch bei transgenen Mäusen und Ratten in Kombination mit verschiedenen pharmakologischen Verbindungen angewendet werden. Hier werden Konzentrations-Response- und Blocking-Experimente von Ratten sowie Konzentrations-Response-Ergebnisse von Wildtyp- und transgenen Mäusen vorgestellt.
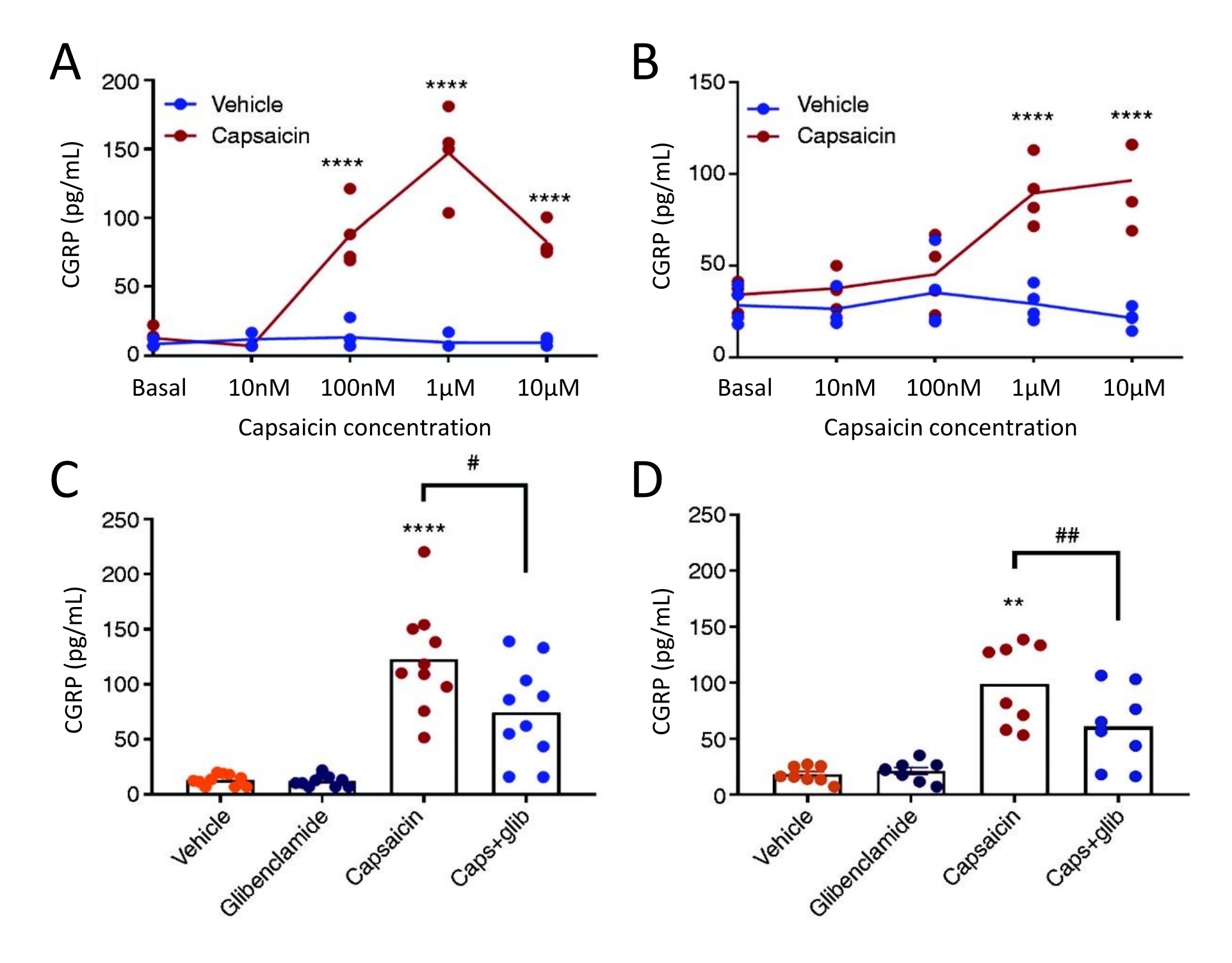
Die CGRP-Freisetzungsmethode wurde verwendet, um die Wirkung desK-ATP-Kanal-Inhibitors Glibenclamid auf die CGRP-Freisetzung aus TG und Dura mater bei weiblichen spontanen trigeminalen allodynischen (STA) Ratten zu untersuchen. Zunächst wurde die optimale Konzentration von Capsaicin unter Verwendung eines Konzentrations-Wirkungs-Studiendesigns ermittelt. Die Capsaicin-Exposition induzierte im Vergleich zum Vehikel eine signifikante CGRP-Freisetzung aus Dura mater und TG (Abbildung 5). In der Dura mater wurde die maximale Freisetzung von CGRP bei 1 μM Capsaicin und in TG die maximale CGRP-Freisetzung bei 10 μM Capsaicin gefunden (Abbildung 5A-B). Basierend auf den Konzentrations-Wirkungs-Experimenten wurden 1 μM Capsaicin und 3 μM Glibenclamid für Blockierungsexperimente verwendet. Glibenclamid zeigte keine Wirkung auf die basale CGRP-Freisetzung aus Dura mater (P = 0,441) und TG (P = 0,881), wenn es mit einer Einweg-ANOVA51 analysiert wurde. Glibenclamid reduzierte signifikant die Capsaicin-induzierte CGRP-Freisetzung in Dura mater um 40 % (P = 0,031) und TG um 39 % (P = 0,003) im Vergleich zu Capsaicin mit dem Vehikel, wenn es mit einer Einweg-ANOVA analysiert wurde (Abbildung 5C-D)51.
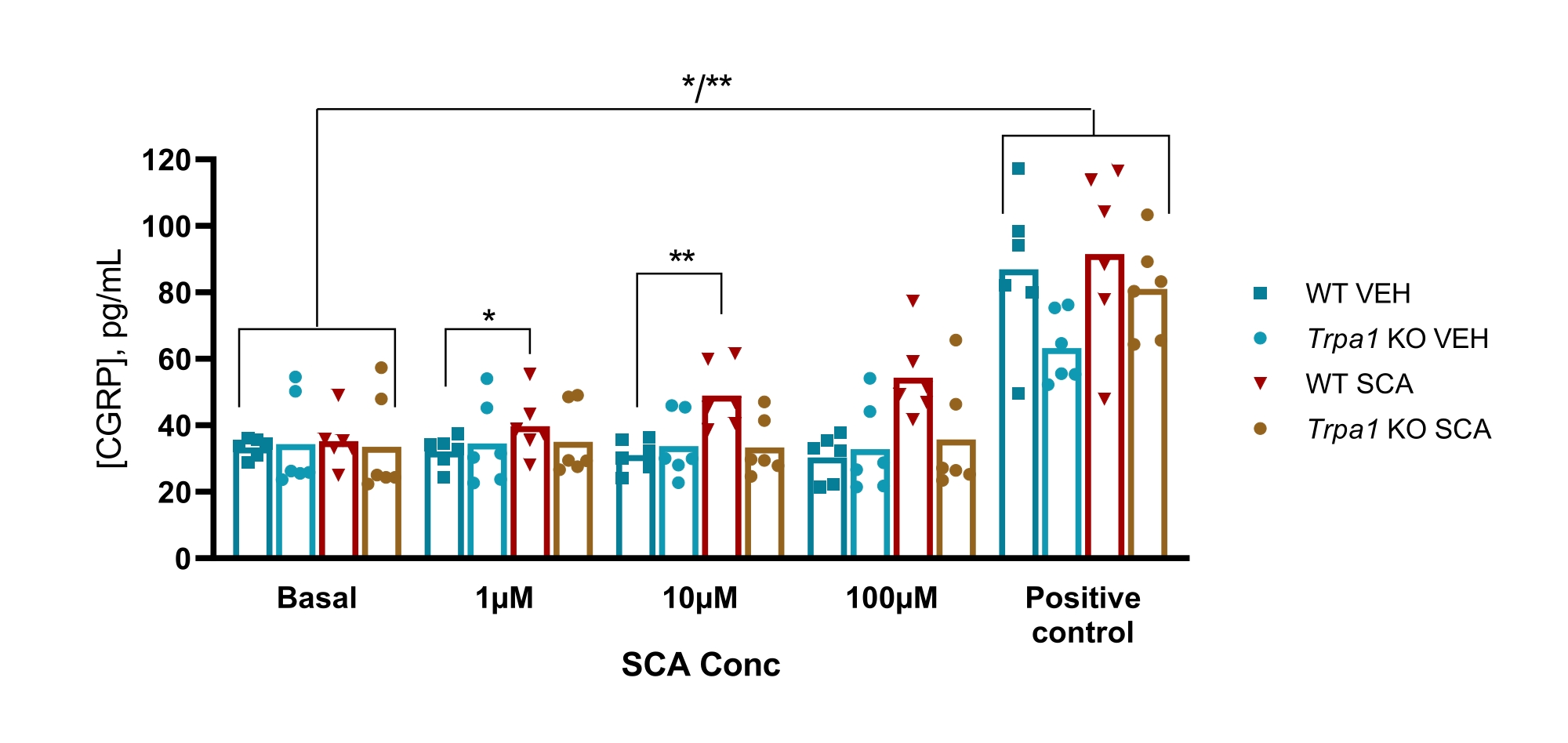
Bei Mäusen wurde das Protokoll verwendet, um die Beteiligung des transienten Rezeptorpotentials Ankyrin 1 (TRPA1) -Ionenkanals in einem GTN-Mausmodell für Migräne zu untersuchen, bei dem die GTN-induzierte Überempfindlichkeit vollständig von TRPA1-Kanälen abhängig war. Es wurde festgestellt, dass der TRPA1-Agonist Supercinnamaldehyd (SCA) CGRP in einer dosisabhängigen Weise aus der TG mit 1 μM, 10 μM und 100 μM SCA freisetzt, was zu einer um 9% (P = 0,23), 51% (P = 0,011) und 69% (P = 0,0097) erhöhten Freisetzung von CGRP im Vergleich zum Vehikel führt, wenn es mit Zwei-Wege-ANOVA analysiert wurde. Diese Freisetzung fehlte bei TG von Trpa1-Nullmäusen, bei denen die Exposition bei 1 μM, 10 μM und 100 μM SCA zu einer 11% (P > 0,99), -13% (P > 0,99) und 9% (P = 0,97) Prozent Veränderung der Freisetzung von CGRP im Vergleich zum Vehikel führte, wenn sie mit Zwei-Wege-ANOVA analysiert wurde. Die anschließende Stimulation mit 10 μM Capsaicin (Positivkontrolle) zeigt, dass alle Gewebeproben CGRP freisetzen konnten (Abbildung 6)50.

Abbildung 1: Schritt-für-Schritt-Dissektion von Gewebe von Ratten. Die Details (A-T) finden Sie im Protokollabschnitt (Schritt 3.1). Bitte klicken Sie hier, um eine größere Version dieser Abbildung zu sehen.
{kind=link}

Abbildung 2: Schritt-für-Schritt-Dissektion von Gewebe von Mäusen. Die Details (A-T) finden Sie im Protokollabschnitt (Schritt 3.2). Bitte klicken Sie hier, um eine größere Version dieser Abbildung zu sehen.
{kind=link}

Abbildung 3: Waschen und Inkubieren von Nagetiergewebe. (A-B) Frisch isoliertes Gewebe in Kunststoffbehältern mit SIF. Die Behälter sind mit Tüll überzogen, um einen einfachen SIF-Wechsel zu ermöglichen. (C) Rattengewebe - Zwei Schädelhälften mit Dura mater, die die inneren Auskleidungen des Schädels bedecken, die auf einer 6-Well-Kulturplatte platziert sind. Der rechte und linke Trigeminuskern caudalis in getrennten Mikrozentrifugenröhrchendeckeln (obere Deckelreihe). Die beiden Trigeminusganglien in einzelnen Mikrozentrifugen-Röhrchendeckeln (untere Deckelreihe). (D) Mausgewebe - Der Trigeminuskern Caudalis-enthaltender Teil des Hirnstamms in einem separaten Mikrozentrifugen-Röhrchendeckel (oben). Zwei Maus-Trigeminusganglien befinden sich in einem Mikrozentrifugen-Röhrchendeckel (unten). Bitte klicken Sie hier, um eine größere Version dieser Abbildung zu sehen.
{kind=link}

Abbildung 4: Studiendesigns für die CGRP-Freigabe. Drei verschiedene Protokolle für die Durchführung von CGRP-Freisetzungsexperimenten. Medikamente sollen in synthetischer interstitieller Flüssigkeit (SIF) verdünnt werden. (A) Einmalige Stimulation. (B) Stimulation der Konzentrationsreaktion. (C) Hemmung einer Stimulation. Bitte klicken Sie hier, um eine größere Version dieser Abbildung zu sehen.
{kind=link}

Abbildung 5: Glibenclamid hemmt die Capsaicin-induzierte CGRP-Freisetzung aus der Rattendura mater und dem Trigeminusganglion. Die CGRP-Konzentrationen von Trigeminusganglion und Dura mater, die von weiblichen spontanen trigeminalen allodynischen (STA) Ratten isoliert wurden (215-318 g), die ursprünglich von der Thomas Jefferson University52 stammten, wurden mit kommerziellen menschlichen CGRP-EIA-Kits gemessen. (A-B) Freisetzung von CGRP aus (A) dura mater und (B) Ganglion trigeminus nach steigenden Konzentrationen von Capsaicin (10 nM, 100 nM, 1 μM und 10 μM) (n = 4). Die Daten werden als einzelne Punkte dargestellt und mit einer Zwei-Wege-ANOVA analysiert. p < 0,0001 (C-D) Konzentrationen von CGRP, die aus (C) Dura mater und (D) Trigeminusganglion nach 10-minütiger Exposition gegenüber Vehikel, 3 μM Glibenclamid (glib), 1 μM Capsaicin und 1 μM Capsaicin + 3 μM glib (n = 6-11) freigesetzt werden. Die Daten werden als Einzelpunkte und Mittelwerte dargestellt und mit einer Einweg-ANOVA analysiert. *im Vergleich zum Fahrzeug. #Capsaicin im Vergleich zu Capsaicin + GLIB. #P < 0,05, ** und ##P < 0,01, ****P < 0,0001. Den Analysen folgte der Mehrfachvergleichstest von Bonferroni. Für alle Tests wurde ein signifikanter Wert von α = 0,05 verwendet. Diese Abbildung wurde von Christensen et al. modifiziert. 51. Bitte klicken Sie hier, um eine größere Version dieser Abbildung anzuzeigen.
{kind=link}

Abbildung 6: Die SCA-Exposition führt zu einer TRPA1-abhängigen CGRP-Freisetzung aus dem Trigeminusganglion der Maus. CGRP-Konzentrationen, die aus dem Trigeminusganglion freigesetzt wurden, das aus WT und Trpa1 null (Trpa1tm1/Dpc)53 männlichen Mäusen (8-10 Wochen) isoliert wurde, wurden mit Ratten-CGRP-EIA-Kits nach Exposition bei Supercinnamaldehyd (SCA) bei 1 μM, 10 μM und 100 μM und Capsaicin bei 10 μM als Positivkontrolle (n = 6) gemessen. Die Daten jeder Maus werden als einzelne Punkte dargestellt, und Balken zeigen die Mittelwerte an. Statistik: Der Vergleich zwischen SCA und Vehikel bei jeder Konzentration und zwischen Basal- und Positivkontrolle wurde mit einer bidirektional wiederholten ANOVA durchgeführt. Ein signifikantes Niveau von α = 0,05. *P < 0,05, **P < 0,01. Diese Abbildung wurde von Christensen et al. modifiziert. 50. Bitte klicken Sie hier, um eine größere Version dieser Abbildung anzuzeigen.
{kind=link}
Diskussion
Die beschriebene Methode wurde nach Studien entwickelt, die die Bedeutung von CGRP in der Pathophysiologie von Migräne zeigen. Es eignet sich gut, um die Mechanismen zu untersuchen, die an der Freisetzung von CGRP aus dem trigeminovaskulären System beteiligt sind, das für die Schmerzsignalisierung in der Kopfregion entscheidend ist. Die Menge an CGRP, die in diesem Modell erhalten wird, misst direkt die CGRP-Freisetzung aus den Trigeminusnerven, die Dura mater, TG und TNC innervieren. Die Menge der CGRP-Freisetzung ist quantitativ größer 45,54 als die Freisetzung, die im Plasma nach Thermokoagulation beim Menschen, Trigeminusstimulation bei Katzen 39,55,56 und bei Migräneattacken23 gemessen wurde. Eine Erklärung könnte sein, dass CGRP im Blut verdünnt und abgebaut wird54. Es ist jedoch zu beachten, dass die direkte Stimulation mit Chemikalien der pathophysiologischen Aktivierung überlegen sein kann. Weitere Vorteile sind, dass es möglich ist, die Freisetzung von drei verschiedenen Stellen innerhalb des trigeminovaskulären Systems zu lokalisieren und dass sie zusammen mit pharmakologischer Manipulation und in Gewebe von gentechnisch veränderten Nagetieren verwendet werden kann.
In letzter Zeit konzentrieren sich viele präklinische Nagetiermodelle auf die systemische Verabreichung von Substanzen und die anschließende Schmerz- oder Migräne-bezogene Messungen unter Verwendung von Frey-Tests57,58, Grimassen 59,60,61 oder leichter Aversion62,63,64. Diese Methoden sind nützlich, um die schmerzauslösenden und schmerzlindernden Eigenschaften verschiedener Substanzen zu verstehen. Diese Ansätze liefern jedoch keine Informationen über spezifische Zielgewebe. Bei der vorliegenden Methode wird das trigeminovaskuläre System in drei Strukturen unterteilt: die Dura mater, TG und TNC. Dies ermöglicht die lokale Exposition jedes Bauwerks und die Beurteilung des Wirkungsortes eines bestimmten Stoffes. Dies wurde in einer Studie aus dem Jahr 2011 verwendet, in der die Rolle von spannungsgesteuerten Kalziumkanälen bei Ratten untersucht wurde und die Hemmung dieser Kanäle in den drei Strukturen des trigeminovaskulären Weges47 unterschiedlich war. Bei der Analyse von Strukturen des Nervensystems ist eine Axotomie unvermeidlich. Es wurde gezeigt, dass die Axotomie die Transkription verschiedener Gene verändert65. Diese transkriptionellen Veränderungen sind zu langsam, um die Ergebnisse dieser Methode zu beeinflussen, aber Veränderungen in der Phosphorylierung können im Vergleich zu In-vivo-Situationen nicht ausgeschlossen werden46. Obwohl Neuropeptide wie CGRP im Zellsoma gebildet werden, erfolgt die Freisetzung und Wirkung von Neuropeptiden in der Regel an den zentralen oder peripheren Nervenenden. Daher sind Untersuchungen von intakten Neuronen, einschließlich Terminals, interessant, wenn die Freisetzung von Neuropeptiden untersucht wird. Daher wurden Methoden zur Untersuchung von Kulturen von Neuronen aus isolierten Ganglien etabliert, die als Modell der Terminals dienen. Neuronale Zellkulturen unterliegen jedoch mehreren Problemen, da mechanische Dissoziation Neuronen in einer Kultur zerstören kann46. Der längere Zeitrahmen, der mit der Kultivierung von Zellen verbunden ist, macht diese Methode anfällig für transkriptomische Veränderungen aufgrund von Axotomie und Kulturbedingungen65. Darüber hinaus haben die Zugabe von Wachstumsfaktoren und die Kultivierung auf Oberflächenbeschichtungen die neuronalen Eigenschaften als Transmitter- und Rezeptorexpressionverändert 66,67,68,69. Diese Probleme werden vermieden, wenn frisch isolierte intakte Ganglien anstelle von neuronalen Zellkulturen untersucht werden.
Eine Herausforderung bei der ex vivo CGRP-Freisetzungsmethode ist die präzise Dissektion des Gewebes, die für reproduzierbare Ergebnisse erforderlich ist. Eine besonders genaue Dissektion des TNC ist eine Herausforderung, da es sich um eine Struktur innerhalb des Hirnstamms ohne sichtbare Grenzen handelt. Darüber hinaus ist die Dura mater zerbrechlich, und die Entfernung des Gehirns muss sorgfältig durchgeführt werden, um eine intakte Struktur zu gewährleisten. Diese Hindernisse können zu unterschiedlichen Gewebegrößen und damit zu unterschiedlichen basalen und stimulationsinduzierten CGRP-Spiegeln führen. Diese Variation kann jedoch durch Normalisierung auf die basale CGRP-Freisetzung erklärt werden. Es sollte auch beachtet werden, dass bei der Isolierung des TNC von Mäusen der gesamte untere Teil des Hirnstamms isoliert wird und nicht der spezifischere TNC-haltige Teil wie bei Ratten. Im Allgemeinen kann es von Vorteil sein, Rattengewebe zu verwenden, da dies die Messung der CGRP-Freisetzung aus der Dura mater und eine genauere Dissektion des TNC ermöglicht. Darüber hinaus ermöglicht die Größe des Gewebes auch die Verwendung einer Ratte als Vehikelkontrolle, da eine Ratte zu zwei Schädelhälften, zwei TGs und zwei TNCs führt, wobei ein Teil des Gewebes für die Substanzstimulation und das andere für das Vehikel verwendet wird. Bei der Verwendung von Mäusen werden zwei Tiere für ein Experiment benötigt, da beide TGs in einer Probe zusammengefasst werden und die TNCs als ein Hirnstamm seziert werden. Daher werden zwei TGs und ein Hirnstamm für die Substanzstimulation verwendet, und zwei TGs und ein Hirnstamm von einer anderen Maus werden für die Vehikelkontrolle verwendet. Dies führt dazu, dass doppelt so viele Mäuse im Vergleich zu Ratten verwendet werden, um die gleiche Anzahl von Replikaten zu erhalten. Um die Anzahl der verwendeten Mäuse zu reduzieren, wurde eine Methode zur Messung der CGRP-Freisetzung aus Hirnstammschnitten vorgeschlagen49. Es ist von Vorteil, dass die Methode modifiziert wurde, um den Einsatz von Mäusen zu ermöglichen. Dies ermöglicht die Verwendung vieler bereits verfügbarer transgener Mausstämme, ein nützliches Werkzeug für die Untersuchung von z.B. Signalwegen. Eine Positivkontrolle am Ende eines Experiments sollte enthalten sein, um sicherzustellen, dass das im Experiment verwendete Gewebe CGRP freisetzen kann. Die Positivkontrolle könnte der TRPV1-Agonist Capsaicin oder das depolarisierende Stimulus-Kalium (KCl) sein, von denen festgestellt wurde, dass sie CGRP aus dem trigeminovaskulären System sowohl bei Mäusen als auch bei Ratten freisetzen 46,47,48,49,50. Darüber hinaus wurde die Methode auch angepasst, um die Freisetzung anderer relevanter Peptide wie Hypophysenadenylatcyclase-aktivierendes Peptid (PACAP) zu messen - ein weiteres Peptid, das in der Migräneforschung von großem Interesse ist70.
Die Methode bietet ein nützliches Werkzeug zur Untersuchung der CGRP-Freisetzung aus spezifischen Zielgeweben bei Ratten und Mäusen. Es ist eine relativ schnelle Methode, die Probleme vermeidet, die mit der Kultivierung von Neuronen verbunden sind. Das Methodenprotokoll kann leicht modifiziert werden, um die Konzentrations-Wirkungs-Beziehung oder die Hemmung einer Reaktion durch verschiedene pharmakologische Verbindungen zu untersuchen. Die Ex-vivo-CGRP-Freisetzungsmethode ist eine von mehreren präklinischen Methoden, die für die Untersuchung der Rolle von CGRP und anderer Mechanismen im Zusammenhang mit der CGRP-Freisetzung in der Pathophysiologie von Migräne nützlich sind.
Offenlegungen
Die Autoren haben nichts zu offenbaren.
Danksagungen
Diese Arbeit wurde von der Candys Foundation finanziert.
Materialien
| Name | Company | Catalog Number | Comments |
| 6-well culture plate | NUNC | 140675 | |
| Calcium chloride dihydrate | Merck | 1.02382.1000 | For SIF buffer |
| Caps for plastic containers | ThermoFisher Scientific | 536617 | |
| Capsaicin | Merck | M2028 | |
| CGRP kits | AH Diagnostics | A05482.96 | |
| CO2 | Strandmøllen | 4.6 | For carbogen gassing of SIF |
| Delicate Bone Trimmer | Fine Science Tools | 16109-14 | |
| Glibenclamide | Tocris | 911 | |
| Glucose | Merck | G7021 | For SIF buffer |
| Guillotine for rats | Scandidact | NS-802 | |
| Magnesium sulfate heptahydrate | Merck | M5921 | For SIF buffer |
| Microcenrifuge tubes + lids/caps | VWR | 700-5239 | |
| Mini Hacksaw | BAHCO | 208 | |
| O2 | Strandmøllen | 4.5 | For carbogen gassing of SIF |
| Pentobarbital | Glostrup pharmacy | NA | Magistral formula |
| Plastic containers | ThermoFisher Scientific | 536455 | |
| Plate photometer - Infinite M200 | Tecan | NA | Infinite M200 is discontinued. A Infinite 200 PRO is available at Tecan. Software: SW Magellan v.6.3 |
| Potassium chloride | Merck | P9333 | For SIF buffer |
| Scissor | Allgaier Instruments | 307-156-170 | |
| Small scissor | Allgaier Instruments | 04-520-115 | |
| Sodium bicarbonate | Merck | S6014 | For SIF buffer |
| Sodium chloride | Merck | S9888 | For SIF buffer |
| Sodium dihydrogen phosphate monohydrate | Merck | 1.06346.1000 | For SIF buffer |
| Sodium gluconate | Merck | S2054 | For SIF buffer |
| Spatula | Bochem Lab Supply | 3018 | |
| Spring scissor | Fine Science Tools | 15024-10 | |
| Sucrose | Merck | 84097 | For SIF buffer |
| Supercinnamaldehyde | Merck | S3322 | |
| Tulle (fabrics) | NA | NA | Bought in the local fabrics store |
Referenzen
- GBD 2016 Collaborators. Global, regional, and national incidence, prevalence, and years lived with disability for 328 diseases and injuries for 195 countries, 1990-2016: A systematic analysis for the Global Burden of Disease Study. The Lancet. 390 (10100), 1211-1259 (2017).
- Dodick, D. W. CGRP ligand and receptor monoclonal antibodies for migraine prevention: Evidence review and clinical implications. Cephalalgia: An International Journal of Headache. 39 (3), 445-458 (2019).
- Sacco, S., et al. European headache federation guideline on the use of monoclonal antibodies acting on the calcitonin gene related peptide or its receptor for migraine prevention. The Journal of Headache and Pain. 20 (1), 6 (2019).
- Moreno-Ajona, D., Pérez-Rodríguez, A., Goadsby, P. J. Gepants, calcitonin-gene-related peptide receptor antagonists: what could be their role in migraine treatment. Current Opinion in Neurology. 33 (3), 309-315 (2020).
- Khan, S., Olesen, A., Ashina, M. CGRP, a target for preventive therapy in migraine and cluster headache: Systematic review of clinical data. Cephalalgia: An International Journal of Headache. 39 (3), 374-389 (2019).
- Edvinsson, L., Haanes, K. A., Warfvinge, K., Krause, D. N. CGRP as the target of new migraine therapies - successful translation from bench to clinic. Nature Reviews Neurology. 14 (6), 338 (2018).
- Goadsby, P. J., et al. Pathophysiology of migraine: A disorder of sensory processing. Physiological Reviews. 97 (2), 553-622 (2017).
- Amara, S. G., Jonas, V., Rosenfeld, M. G., Ong, E. S., Evans, R. M. Alternative RNA processing in calcitonin gene expression generates mRNAs encoding different polypeptide products. Nature. 298 (5871), 240-244 (1982).
- Rosenfeld, M. G., et al. Production of a novel neuropeptide encoded by the calcitonin gene via tissue-specific RNA processing. Nature. 304 (5922), 129-135 (1983).
- Edvinsson, L., Goadsby, P. J. Discovery of CGRP in relation to migraine. Cephalalgia: An International Journal of Headache. 39 (3), 331-332 (2019).
- Brain, S. D., Williams, T. J., Tippins, J. R., Morris, H. R., MacIntyre, I. Calcitonin gene-related peptide is a potent vasodilator. Nature. 313 (5997), 54-56 (1985).
- Fisher, L. A., et al. Stimulation of noradrenergic sympathetic outflow by calcitonin gene-related peptide. Nature. 305 (5934), 534-536 (1983).
- Hanko, J., Hardebo, J. E., Kåhrström, J., Owman, C., Sundler, F. Calcitonin gene-related peptide is present in mammalian cerebrovascular nerve fibres and dilates pial and peripheral arteries. Neuroscience Letters. 57 (1), 91-95 (1985).
- Uddman, R., Edvinsson, L., Ekblad, E., Håkanson, R., Sundler, F. Calcitonin gene-related peptide (CGRP): perivascular distribution and vasodilatory effects. Regulatory Peptides. 15 (1), 1-23 (1986).
- Edvinsson, L. Functional role of perivascular peptides in the control of cerebral circulation. Trends in Neurosciences. 8, 126-131 (1985).
- Edvinsson, L., Ekman, R., Jansen, I., Ottosson, A., Uddman, R. Peptide-containing nerve fibers in human cerebral arteries: Immunocytochemistry, radioimmunoassay and in vitro pharmacology. Annals of Neurology. 21 (5), 431-437 (1987).
- Edvinsson, L., Fredholm, B. B., Hamel, E., Jansen, I., Verrecchia, C. Perivascular peptides relax cerebral arteries concomitant with stimulation of cyclic adenosine monophosphate accumulation or release of an endothelium-derived relaxing factor in the cat. Neuroscience Letters. 58 (2), 213-217 (1985).
- Brain, S. D., Grant, A. D. Vascular actions of calcitonin gene-related peptide and adrenomedullin. Physiological Reviews. 84 (3), 903-934 (2004).
- Skofitsch, G., Jacobowitz, D. M. Calcitonin gene-related peptide: Detailed immunohistochemical distribution in the central nervous system. Peptides. 6 (4), 721-745 (1985).
- Suzuki, N., Hardebo, J. E., Owman, C. Origins and pathways of cerebrovascular nerves storing substance P and calcitonin gene-related peptide in rat. Neuroscience. 31 (2), 427-438 (1989).
- Uddman, R., Edvinsson, L., Ekman, R., Kingman, T., McCulloch, J. Innervation of the feline cerebral vasculature by nerve fibers containing calcitonin gene-related peptide: trigeminal origin and co-existence with substance P. Neuroscience Letters. 62 (1), 131-136 (1985).
- Warfvinge, K., Edvinsson, L. Distribution of CGRP and CGRP receptor components in the rat brain. Cephalalgia: An International Journal of Headache. 39 (3), 342-353 (2019).
- Goadsby, P. J., Edvinsson, L., Ekman, R. Vasoactive peptide release in the extracerebral circulation of humans during migraine headache. Annals of Neurology. 28 (2), 183-187 (1990).
- Lassen, L. H., et al. CGRP may play a causative role in migraine. Cephalalgia: An International Journal of Headache. 22 (1), 54-61 (2002).
- Olesen, J., et al. Calcitonin gene-related peptide receptor antagonist BIBN 4096 BS for the acute treatment of migraine. New England Journal of Medicine. 350 (11), 1104-1110 (2004).
- Eftekhari, S., et al. Localization of CGRP, CGRP receptor, PACAP and glutamate in trigeminal ganglion. Relation to the blood-brain barrier. Brain Research. 1600, 93-109 (2015).
- Keller, J. T., Marfurt, C. F. Peptidergic and serotoninergic innervation of the rat dura mater. Journal of Comparative Neurology. 309 (4), 515-534 (1991).
- Edvinsson, L., et al. Innervation of the human middle meningeal artery: immunohistochemistry, ultrastructure, and role of endothelium for vasomotility. Peptides. 19 (7), 1213-1225 (1998).
- Lennerz, J. K., et al. Calcitonin receptor-like receptor (CLR), receptor activity-modifying protein 1 (RAMP1), and calcitonin gene-related peptide (CGRP) immunoreactivity in the rat trigeminovascular system: Differences between peripheral and central CGRP receptor distribution. Journal of Comparative Neurology. 507 (3), 1277-1299 (2008).
- Eftekhari, S., Edvinsson, L. Calcitonin gene-related peptide (CGRP) and its receptor components in human and rat spinal trigeminal nucleus and spinal cord at C1-level. BMC Neuroscience. 12, 112 (2011).
- Eftekhari, S., et al. Differential distribution of calcitonin gene-related peptide and its receptor components in the human trigeminal ganglion. Neuroscience. 169 (2), 683-696 (2010).
- Eftekhari, S., Warfvinge, K., Blixt, F. W., Edvinsson, L. Differentiation of nerve fibers storing CGRP and CGRP receptors in the peripheral trigeminovascular system. The Journal of Pain. 14 (11), 1289-1303 (2013).
- Spitzer, M. J. S., Reeh, P. W., Sauer, S. K. Mechanisms of potassium- and capsaicin-induced axonal calcitonin gene-related peptide release: Involvement of L- and T-type calcium channels and TRPV1 but not sodium channels. Neuroscience. 151 (3), 836-842 (2008).
- Evans, A. R., Nicol, G. D., Vasko, M. R. Differential regulation of evoked peptide release by voltage-sensitive calcium channels in rat sensory neurons. Brain Research. 712 (2), 265-273 (1996).
- Gupta, S., Villalón, C. M. The relevance of preclinical research models for the development of antimigraine drugs: Focus on 5-HT 1B/1D and CGRP receptors. Pharmocology & Therapeutics. 128 (1), 170-190 (2010).
- Williamson, D. J., Hargreaves, R. J., Hill, R. G., Shepheard, S. L. Intravital microscope studies on the effects of neurokinin agonists and calcitonin gene-related peptide on dural vessel diameter in the anaesthetized rat. Cephalalgia: An International Journal of Headache. 17 (4), 518-524 (1997).
- Gupta, S., Bhatt, D. K., Boni, L. J., Olesen, J. Improvement of the closed cranial window model in rats by intracarotid infusion of signalling molecules implicated in migraine. Cephalalgia: An International Journal of Headache. 30 (1), 27-36 (2010).
- Knight, Y. E., Edvinsson, L., Goadsby, P. J. Blockade of calcitonin gene-related peptide release after superior sagittal sinus stimulation in cat: a comparison of avitriptan and CP122, 288. Neuropeptides. 33 (1), 41-46 (1999).
- Goadsby, P. J., Edvinsson, L., Ekman, R. Release of vasoactive peptides in the extracerebral circulation of humans and the cat during activation of the trigeminovascular system. Annals of Neurology. 23 (2), 193-196 (1988).
- Tassorelli, C., Joseph, S. A. Systemic nitroglycerin induces Fos immunoreactivity in brainstem and forebrain structures of the rat. Brain Research. 682 (1-2), 167-181 (1995).
- Ramachandran, R., et al. A naturalistic glyceryl trinitrate infusion migraine model in the rat. Cephalalgia: An International Journal of Headache. 32 (1), 73-84 (2012).
- Hoskin, K. L., Zagami, A. S., Goadsby, P. J. Stimulation of the middle meningeal artery leads to Fos expression in the trigeminocervical nucleus: a comparative study of monkey and cat. Journal of Anatomy. 194, 579-588 (1999).
- Charbit, A. R., Akerman, S., Goadsby, P. J. Comparison of the Effects of Central and Peripheral Dopamine Receptor Activation on Evoked Firing in the Trigeminocervical Complex. Journal of Pharmacology and Experimental Therapeutics. 331 (2), 752-763 (2009).
- Koulchitsky, S., Fischer, M., Messlinger, K. Calcitonin gene-related peptide receptor inhibition reduces neuronal activity induced by prolonged increase in nitric oxide in the rat spinal trigeminal nucleus. Cephalalgia: An International Journal of Headache. 29 (4), 408-417 (2009).
- Ebersberger, A., Averbeck, B., Messlinger, K., Reeh, P. W. Release of substance P, calcitonin gene-related peptide and prostaglandin E2 from rat dura mater encephali following electrical and chemical stimulation in vitro. Neuroscience. 89 (3), 901-907 (1999).
- Eberhardt, M., et al. Calcitonin gene-related peptide release from intact isolated dorsal root and trigeminal ganglia. Neuropeptides. 42 (3), 311-317 (2008).
- Amrutkar, D. V., Ploug, K. B., Olesen, J., Jansen-Olesen, I. Role for voltage gated calcium channels in calcitonin gene-related peptide release in the rat trigeminovascular system. Neuroscience. 172, 510-517 (2011).
- Gupta, S., et al. Evidence for CGRP re-uptake in rat dura mater encephali. British Journal of Pharmacology. 161 (8), 1885-1898 (2010).
- Kageneck, C., Nixdorf-Bergweiler, B. E., Messlinger, K., Fischer, M. J. M. Release of CGRP from mouse brainstem slices indicates central inhibitory effect of triptans and kynurenate. Journal of Headache and Pain. 15 (1), 1-9 (2014).
- Christensen, S. L., et al. CGRP-dependent signalling pathways involved in mouse models of GTN- cilostazol- and levcromakalim-induced migraine. Cephalalgia: An International Journal of Headache. 41 (14), 1413-1426 (2021).
- Christensen, S. L., et al. ATP sensitive potassium (KATP) channel inhibition: A promising new drug target for migraine. Cephalalgia: An International Journal of Headache. 40 (7), 650-664 (2020).
- Oshinsky, M. L., et al. Spontaneous trigeminal allodynia in rats: A model of primary headache. Headache. 52 (9), 1336 (2012).
- Kwan, K. Y., et al. TRPA1 contributes to cold, mechanical, and chemical nociception but is not essential for hair-cell transduction. Neuron. 50 (2), 277-289 (2006).
- Eltorp, C., Jansen-Olesen, I., Hansen, A. J. Release of calcitonin gene-related peptide (CGRP) from guinea pig dura mater in vitro is inhibited by sumatriptan but unaffected by nitric oxide. Cephalalgia: An International Journal of Headache. 20 (9), 838-844 (2000).
- Zagami, A. S., Goadsby, P. J., Edvinsson, L. Stimulation of the superior sagittal sinus in the cat causes release of vasoactive peptides. Neuropeptides. 16 (2), 69-75 (1990).
- Goadsby, P. J., Edvinsson, L. The trigeminovascular system and migraine: studies characterizing cerebrovascular and neuropeptide changes seen in humans and cats. Annals of Neurology. 33 (1), 48-56 (1993).
- Bates, E. A., et al. Sumatriptan alleviates nitroglycerin-induced mechanical and thermal allodynia in mice. Cephalalgia: An International Journal of Headache. 30 (2), 170-178 (2010).
- Pradhan, A. A., et al. Characterization of a novel model of chronic migraine. Pain. 155 (2), 269-274 (2014).
- Mogil, J. S., Pang, D. S. J., Silva Dutra, G. G., Chambers, C. T. The development and use of facial grimace scales for pain measurement in animals. Neuroscience and Biobehavioral Reviews. 116, 480-493 (2020).
- Sotocinal, S. G., et al. The rat grimace scale: A partially automated method for quantifying pain in the laboratory rat via facial expressions. Molecular Pain. 7, 55 (2011).
- Langford, D. J., et al. Coding of facial expressions of pain in the laboratory mouse. Nature Methods. 7 (6), 447-449 (2010).
- Mahmoudi, J., et al. Cerebrolysin attenuates hyperalgesia, photophobia, and neuroinflammation in a nitroglycerin-induced migraine model in rats. Brain Research Bulletin. 140, 197-204 (2018).
- Farajdokht, F., Babri, S., Karimi, P., Mohaddes, G. Ghrelin attenuates hyperalgesia and light aversion-induced by nitroglycerin in male rats. Neuroscience Letters. 630, 30-37 (2016).
- Kuburas, A., et al. PACAP Induces Light Aversion in Mice by an Inheritable Mechanism Independent of CGRP. Journal of Neuroscience. 41 (21), 4697-4715 (2021).
- Buschmann, T., et al. Expression of Jun, Fos, and ATF-2 proteins in axotomized explanted and cultured adult rat dorsal root ganglia. Neuroscience. 84 (1), 163-176 (1998).
- Lee, Y. J., Zachrisson, O., Tonge, D. A., McNaughton, P. A. Upregulation of bradykinin B2 receptor expression by neurotrophic factors and nerve injury in mouse sensory neurons. Molecular and Cellular Neuroscience. 19 (2), 186-200 (2002).
- Hari, A., Djohar, B., Skutella, T., Montazeri, S. Neurotrophins and extracellular matrix molecules modulate sensory axon outgrowth. International Journal of Developmental Neuroscience. 22 (2), 113-117 (2004).
- Skoff, A. M., Resta, C., Swamydas, M., Adler, J. E. Nerve Growth Factor (NGF) and Glial Cell Line-Derived Neurotrophic Factor (GDNF) regulate substance P release in adult spinal sensory neurons. Neurochemical Research. 28 (6), 847-854 (2003).
- Lindsay, R. M., Harmar, A. J. Nerve growth factor regulates expression of neuropeptide genes in adult sensory neurons. Nature. 337 (6205), 362-364 (1989).
- Edvinsson, J. C. A., et al. Differences in pituitary adenylate cyclase-activating peptide and calcitonin gene-related peptide release in the trigeminovascular system. Cephalalgia: An International Journal of Headache. 40 (12), 1296-1309 (2020).
Nachdrucke und Genehmigungen
Genehmigung beantragen, um den Text oder die Abbildungen dieses JoVE-Artikels zu verwenden
Genehmigung beantragenWeitere Artikel entdecken
This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. Alle Rechte vorbehalten