
Method Article
Ex Vivo Release of Calcitonin Gene-Related Peptide from the Trigeminovascular System in Rodents
In This Article
Summary
The present protocol describes the ex vivo calcitonin gene-related peptide (CGRP) release model and the strategy to quantify the effect of pharmacological agents on the amount of CGRP released from the trigeminovascular system in rodents.
Abstract
Calcitonin gene-related peptide (CGRP) was first discovered in the 1980s as a splice variant from the calcitonin gene. Since its discovery, its role in migraine pathophysiology has been well established, first by its potent vasodilator properties and subsequently by its presence and function as a neurotransmitter in the sensory trigeminovascular system. The migraine-provoking ability of CGRP gave support to the pharma industry to develop monoclonal antibodies and antagonists inhibiting the effect of CGRP. A new treatment paradigm has proven effective in the prophylactic treatment of migraine. One of the useful tools to further understand migraine mechanisms is the ex vivo model of CGRP release from the trigeminovascular system. It is a relatively simple method that can be used with various pharmacological tools to achieve know-how to further develop new effective migraine treatments. The present protocol describes a CGRP release model and the technique to quantify the effect of pharmacological agents on the amount of CGRP released from the trigeminovascular system in rodents. A procedure describing the experimental approach from euthanasia to the measurement of protein levels is provided. The essential isolation of the trigeminal ganglion and the trigeminal nucleus caudalis from both mice and rats and the preparation of rat dura mater are described in detail. Furthermore, representative results from both species (rats and mice) are presented. The technique is a key tool to investigate the molecular mechanisms involved in migraine pathophysiology by using various pharmacological compounds and genetically modified animals.
Introduction
Migraine is a neurological disorder that, according to WHO, is estimated to affect more than 1 billion people and is one of the leading causes of disability worldwide1. Thus, migraine has a significant impact on both patients and society. Despite the recent clinical success of CGRP antagonizing drugs, a large proportion of patients need improved treatment options2,3,4,5. Elucidation of migraine pathophysiology leading to novel effective treatments is required. Signaling within the trigeminovascular system consisting of the meninges, trigeminal ganglia (TG), and trigeminal nucleus caudalis (TNC) is central for migraine pathophysiology6,7.
The 37 amino acid neuropeptide calcitonin gene-related peptide (CGRP) was first discovered in the early 1980s when Amara and co-workers demonstrated that the primary RNA-transcript of the calcitonin gene could be processed to give mRNA coding for CGRP in addition to calcitonin8,9. The subsequent research suggested a link to migraine pathophysiology10. CGRP is a neurotransmitter with potent vasodilating properties11,12,13,14,15,16,17, and it is widely distributed in the central and peripheral nervous system13,14,18,19,20,21,22. The involvement of CGRP in migraine was underlined with the discovery of increased CGRP levels in the extracerebral circulation during migraine attacks in humans23, and that infusion of CGRP causes migraine-like pain in patients24. Two years later, the first proof-of-concept study of the effectivity of the CGRP antagonist olcegepant in treating migraine was published25.
CGRP is abundant in the trigeminovascular system as demonstrated in the TG21,26, sensory nerve fibers innervating the dura mater27,28,29, and TNC30. In the trigeminovascular system, CGRP is found in the small-to-medium-sized neurons of the TG, in unmyelinated C-fibers, and is expressed in nearly 50% of the neuronal population of the TG. The CGRP receptor is expressed mainly in larger neurons and found in myelinated Aδ-fibers31,32. CGRP is released from neurons upon chemical or electrical stimulation33,34. Studies of pathways leading to the release of CGRP and the location of this activation are crucial to understand migraine pathophysiology. Throughout the last 5 decades, preclinical studies have contributed to gaining extensive knowledge on migraine-related signaling and have contributed to development of new treatments35. Many methods considering the vascular and neurogenic involvement have been modified and applied in migraine research. In vivo and in vitro models of arterial responses to biological compounds or pharmacological treatments17,36,37, and electrical nerve-stimulation38,39 can be mentioned. Furthermore, activated neurons in the TNC can be detected by c-Fos expression40,41,42 and electrophysiological recordings in this area43,44. Both methods measure nociceptive signals transmitted to the brain from the head, e.g., dura mater. The use of only one preclinical model does not present the full picture of migraine pathophysiology. Therefore, it is important to combine different models covering as many aspects of migraine pathophysiology as possible. The continued development of new models will cover various aspects of migraine mechanisms, and in time the mystery of migraine pathophysiology will be uncovered.
Here, a detailed protocol is presented of the CGRP release method, performed ex vivo in isolated TG and TNC from mice after chemical stimulation. CGRP release can also be studied in the dura mater from rats. Thus, in the experimental protocol for rats, dura mater is described together with TG and TNC. The basis for the CGRP release method was first described in 1999, where Ebersberger and co-workers conducted pioneering research and found that CGRP was released from dura mater after chemical and electrical stimulation of dural afferents in rats45. Later, this approach was extended to CGRP release from the TG46 and the TNC47. Subsequently, the method was modified to apply to TG and TNC in mice. So far, CGRP release from the dura mater has been challenging in mice.
Protocol
All animal care and experimental procedures were performed in compliance with the European Community guide for the care and use of animals (2010/63/UE). Male C57BL/6JBomTac mice, aged 10 weeks, and male Sprague Dawley rats, aged 10 weeks, were used to demonstrate this protocol.
1. Preparation of the synthetic interstitial fluid
- Prepare synthetic interstitial fluid (SIF) according to the following recipe: 108 mM NaCl, 3.48 mM KCl, 3.50 mM MgSO4, 26 mM NaHCO3, 11.70 mM NaH2PO4, 1.50 mM CaCl2, 9.60 mM Na-gluconate, 5.50 mM glucose and 7.60 mM sucrose (see Table of Materials).
NOTE: SIF can be varied depending on the stimulation target, e.g., a calcium-free solution can be used when studying calcium channels. - Adjust pH to 7.4 and stabilize the pH by carbogen gassing (5% CO2 and 95% O2)46.
2. Euthanasia
- Anesthetize adult mice and rats with a mixture of 70% CO2 and 30% O2. Decapitate mice using a pair of scissors and rats using a guillotine (see Table of Materials).
NOTE: Use a strain and age that fits the aim of the research. Both males and females can be used in this model. - Separate the head from the body at the C3-C4 level of the spinal cord.
NOTE: Euthanasia can also be performed with an intraperitoneal injection of pentobarbital (100-150 mg/kg).
3. Dissection
- Prepare the rat tissue following the steps below.
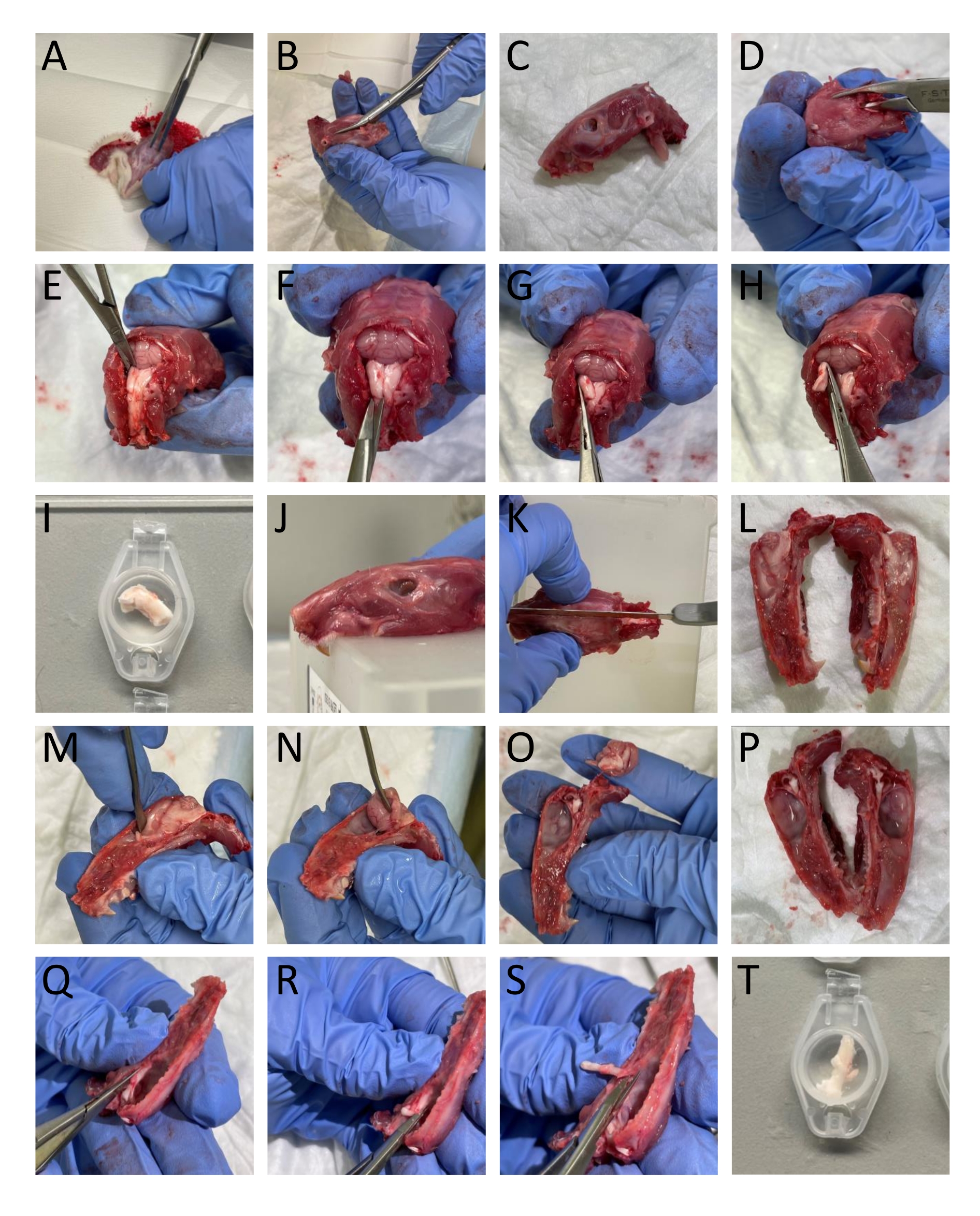
- Remove the skin and the muscle around the head and neck using a pair of scissors (Figure 1A).
- Use a bone trimmer and scissors to separate the lower jaws from the head (Figure 1B-C).
- Open the spinal cord by inserting a bone trimmer caudally into the vertebrae's dorsal part and remove the vertebrae's dorsal part to expose the spinal cord and brainstem (Figure 1D).
- Cut the caudal part of the cranium by the borders of the occipital and interparietal bones to remove these bone structures exposing the cerebellum (Figure 1E).
NOTE: It is important not to damage the brainstem and spinal cord while cutting and removing the vertebras. - Isolate the TNC (Sp5C) running caudally approximately 13-16 mm from the bregma on each side by cutting the dorsolateral part of the brainstem with spring scissors. Immerse the left and the right side TNC into SIF (Figure 1E-I).
NOTE: The description corresponds to adult rats. - Cut the head mid-sagitally to divide the cranium in two using a saw (Figure 1J-L).
- Carefully remove the brain without touching the dura mater attached to the cranium using a spatula and cutting the trigeminal nerve where it enters the brainstem (Figure 1M-P).
- To isolate the TG, cut it, including its branches around the visual borders. Cut the mandibular branch where it enters the foramen ovale. Cut the ophthalmic and maxillary branches entering the skull as they are not divided macroscopically. While dissecting the TG, remove the dura mater covering the TG (Figure 1Q-T).
- Immerse the cranium halves and the TGs in SIF.
- Prepare the mouse tissue following the steps below.
- Remove skin and muscle around the head and neck using small scissors (Figure 2A-B).
- Open the spinal cord by inserting a pair of small scissors caudally into the vertebrae's dorsal part and remove the vertebrae's dorsal part to expose the spinal cord and brainstem (Figure 2C).
NOTE: It is important not to damage the spinal cord while cutting and removing the vertebras. - Cut the cranium by the borders of the occipital and interparietal bones to remove these bone structures exposing the cerebellum (Figure 2D-F).
- Then, cut the parietal bone mid-sagitally and remove the bone to expose the cerebrum (Figure 2G-I).
- Carefully remove the cerebellum with a spatula to expose the brainstem (Figure 2J).
- Isolate the TNC-containing part of the brainstem using spring scissors (Figure 2K-N). Immerse the brainstem with TNC in SIF (Figure 2O).
- Remove the brain and cut the trigeminal nerve where it enters the brainstem (Figure 2P-Q).
- To isolate the TG, cut it, including its branches around the visual borders. Cut the mandibular branch where it enters the foramen ovale. Cut the ophthalmic and maxillary branches entering the skull as they are not divided macroscopically. While dissecting the TG, remove the dura mater covering the TG (Figure 2R-S).
- Immerse TGs in SIF (Figure 2T).
4. Washing
- Wash skull halves, TNCs, and TGs in SIF for 30 min while replacing the SIF every 5 min at room temperature.
NOTE: The washing steps can be performed while keeping the tissue in plastic containers with a tulle lid to enable easy SIF exchange (Figure 3A-B). - Prepare the rat tissue following the steps below.
- Transfer TNC halves to separate microcentrifuge tube caps with 350 µL of SIF (Figure 3C).
- Transfer the TGs to microcentrifuge tube caps - one TG per microcentrifuge tube cap with 350 µL of SIF (Figure 3C).
- Place the skull halves on platforms made of clay or a 6-well culture plate and fill the skull with 400 µL of SIF (Figure 3C).
- Prepare the mouse tissue following the steps below.
- Transfer the brainstem with TNC to a microcentrifuge tube cap with 250 µL of SIF (Figure 3D).
- Transfer the two TGs to a microcentrifuge tube cap with 250 µL of SIF (Figure 3D).
NOTE: Place two TGs per microcentrifuge tube cap when using tissue from mice.
- Place rat skulls and microcentrifuge tube caps with rat and mouse tissue in a humidified incubator at 37 °C. Replace SIF using a pipette every 5 min for 20 min.
NOTE: It is important not to touch the tissue when adding and removing SIF.
5. Drug testing
- Determine the basal CGRP release levels.
- Following the last wash, add 250 µL of SIF to mouse TG and TNC. Add 350 µL to rat TG and TNC and 400 µL to each rat skull.
- After 10 min of incubation, collect 200 µL of the sample in a microcentrifuge tube and add 50 µL of 10x EIA buffer (provided with the CGRP enzyme immunoassay kit, see Table of Materials) to allow measurement of the basal CGRP release (step 6). Discard the remaining liquid.
NOTE: The incubation time must be the same for all samples. - Immediately, store the samples at -20 °C.
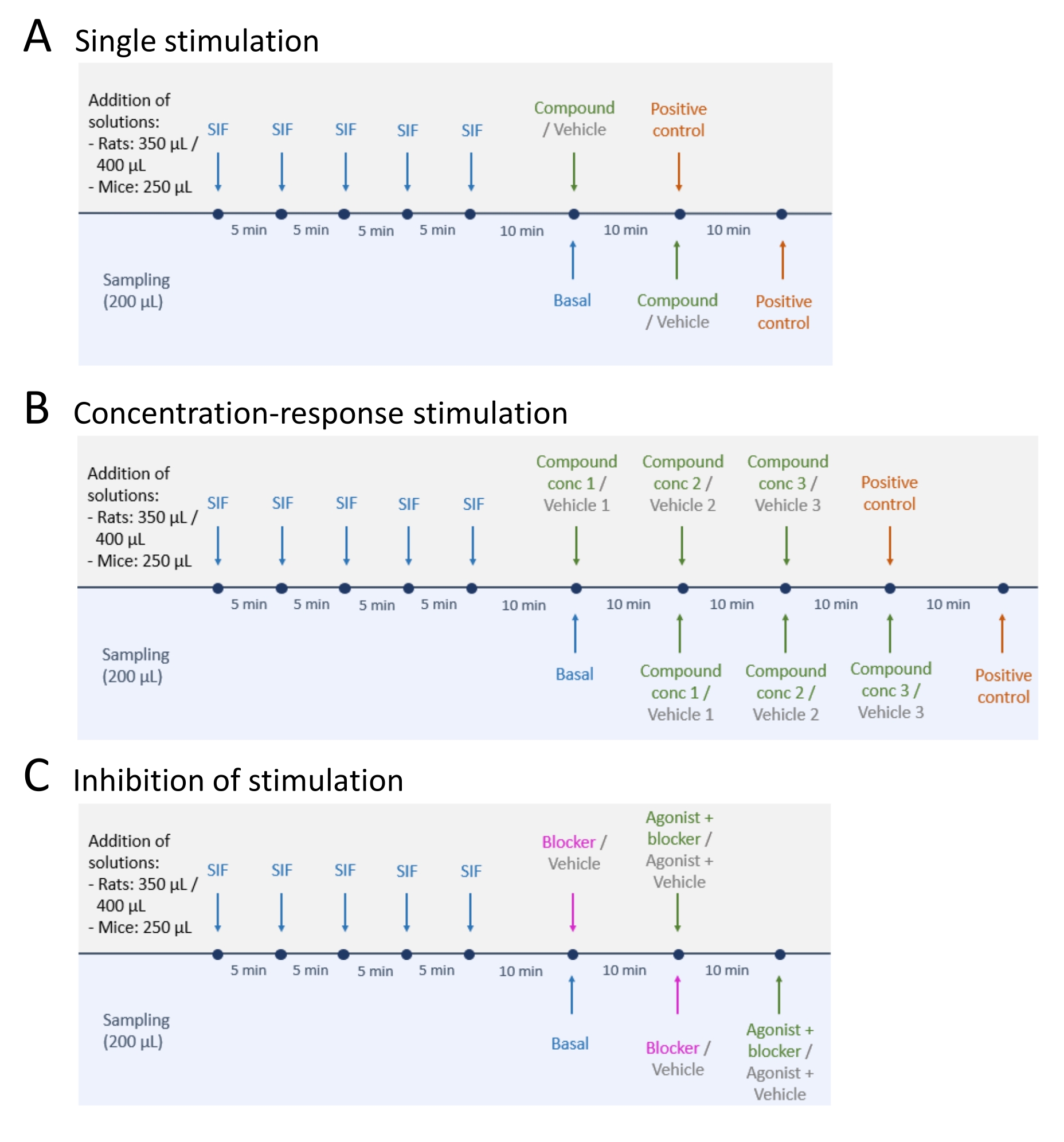
- After sampling of basal CGRP release levels, follow one of the three methods: (A) Single stimulation (step 5.2); (B) Concentration-response stimulation (step 5.3); and (C) Inhibition of stimulation (step 5.4) (Figure 4).
NOTE: The test compound and concentrations used depend on the aim of the study.
- Perform single stimulation following the steps below.
- Add test compound or vehicle to the tissue and leave it for 10 min (volume: 250 µL for mouse TG and TNC, 350 µL for rat TG and TNC, and 400 µL for each rat skull).
- After 10 min of incubation, collect 200 µL of the sample in a microcentrifuge tube with 50 µL of 10x EIA buffer. Discard the remaining liquid and immediately store the samples at -20 °C.
NOTE: The incubation time must be the same for all samples.
- Perform concentration-response stimulation following the steps below.
- Dilute the test compound or vehicle to the desired concentrations. Add the test compound in increasing concentrations starting with the lowest concentration.
NOTE: The test compound and concentrations used depend on the aim of the study. For example, 1 µM, 10 µM, and 100 µM supercinnamaldehyde were used for the present study. - Add the lowest concentration (1 µM for the present study) of the test compound and corresponding vehicle to two identical tissue preparations and incubate for 10 min (volume: 250 µL for mouse tissue, 350 µL for rat TG and TNC, and 400 µL for each rat skull).
- After 10 min of incubation, collect 200 µL of the sample in a microcentrifuge tube with 50 µL of 10x EIA buffer.
- Discard the remaining liquid and add the second lowest concentration (10 µM for the present study) to the tissue.
- Immediately, store the samples at -20 °C.
- Repeat this procedure with the remaining concentrations (100 µM for the present study).
- Dilute the test compound or vehicle to the desired concentrations. Add the test compound in increasing concentrations starting with the lowest concentration.
- Perform inhibition of stimulation following the steps below.
- Add the blocker or vehicle to the tissue and incubate for 10 min (volume: 250 µL for mouse tissue, 350 µL for rat TG and TNC, and 400 µL for each rat skull).
NOTE: The blocker and concentration used depend on the aim of the study. For example, 3 µM glibenclamide was used for the representative result in Figure 5. - After 10 min of incubation, collect a 200 µL sample in a microcentrifuge tube with 50 µL of 10x EIA buffer. Discard the remaining liquid and immediately store the samples at -20 °C.
- Add the agonist or agonist + blocker (see Table of Materials) to the tissue and incubate for 10 min.
NOTE: The agonist, blocker, and concentrations used depend on the aim of the study. In the present study, 3 µM of glibenclamide and 1 µM of capsaicin were used, Figure 5. - After 10 min of incubation, collect a 200 µL sample in a microcentrifuge tube with 50 µL of 10x EIA buffer. Discard the remaining liquid and immediately store the samples at -20 °C.
- Add the blocker or vehicle to the tissue and incubate for 10 min (volume: 250 µL for mouse tissue, 350 µL for rat TG and TNC, and 400 µL for each rat skull).
- Perform a positive control for the experiment.
- When appropriate, add a positive control (e.g., 1-10 µM of capsaicin, see Table of Materials) to the tissue at the end of the protocol and collect a 200 µL sample in a microcentrifuge tube with 50 µL of 10x EIA buffer after an incubation period of 10 min.
NOTE: It is advantageous to include a positive control to ensure the setup and tissues are functioning. Capsaicin48,49,50 or the depolarizing stimulus of potassium (40-60 mM of KCl)46,47,49 are routinely used to cause the release of CGRP from the trigeminovascular system. 40-60 mM of KCl SIF is prepared as SIF, except NaCl is exchanged for KCl on an equimolar basis.
- When appropriate, add a positive control (e.g., 1-10 µM of capsaicin, see Table of Materials) to the tissue at the end of the protocol and collect a 200 µL sample in a microcentrifuge tube with 50 µL of 10x EIA buffer after an incubation period of 10 min.
6. Analysis of CGRP concentrations
- Measure the amount of CGRP released using an enzyme immunoassay (EIA) kit following the manufacturer's protocol (see Table of Materials).
- Measure the optical density at 410 nm using a plate photometer. If a different CGRP EIA kit is used, measure the optical density at the wavelength provided in the manufacturer's protocol.
NOTE: Samples must be diluted to match the standard curve. - Perform data analysis.
- Present the data either as absolute concentrations or normalize to the basal CGRP release from the specific tissue.
Results
This technique is a tool to investigate the CGRP-related molecular mechanisms involved in migraine. It has the advantage of assessing CGRP release from different levels of the trigeminovascular system and can be applied both on wild-type and transgenic mice and rats in combination with various pharmacological compounds. Here, concentration-response and blocking experiments from rats and concentration-response results from wild-type and transgenic mice are presented.
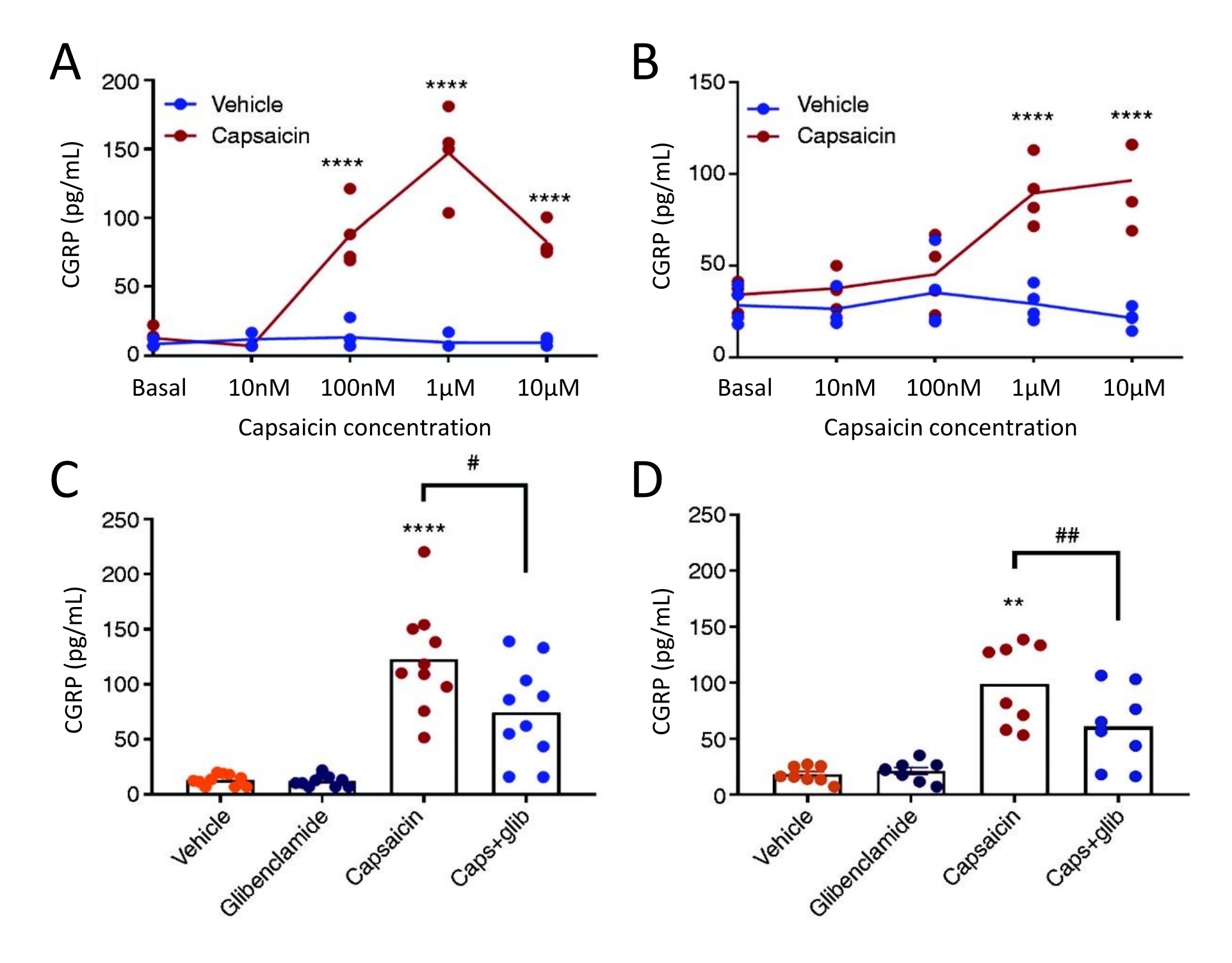
The CGRP release method was used to study the effect of the KATP channel inhibitor glibenclamide on CGRP release from TG and dura mater in female spontaneous trigeminal allodynic (STA) rats. First, the optimal concentration of capsaicin was found using a concentration-response study design. Capsaicin exposure induced a significant CGRP release from dura mater and TG compared to the vehicle (Figure 5). In the dura mater, the maximum release of CGRP was found at 1 µM of capsaicin, and in TG, the maximum CGRP release was found at 10 µM of capsaicin (Figure 5A-B). Based on the concentration-response experiments, 1 µM of capsaicin and 3 µM of glibenclamide were used for blocking experiments. Glibenclamide showed no effect on basal CGRP release from dura mater (P = 0.441) and TG (P = 0.881) when analyzed with a one-way ANOVA51. Glibenclamide significantly reduced the capsaicin-induced CGRP release in dura mater by 40% (P = 0.031) and TG by 39% (P = 0.003) compared to capsaicin with the vehicle when analyzed with a one-way ANOVA (Figure 5C-D)51.
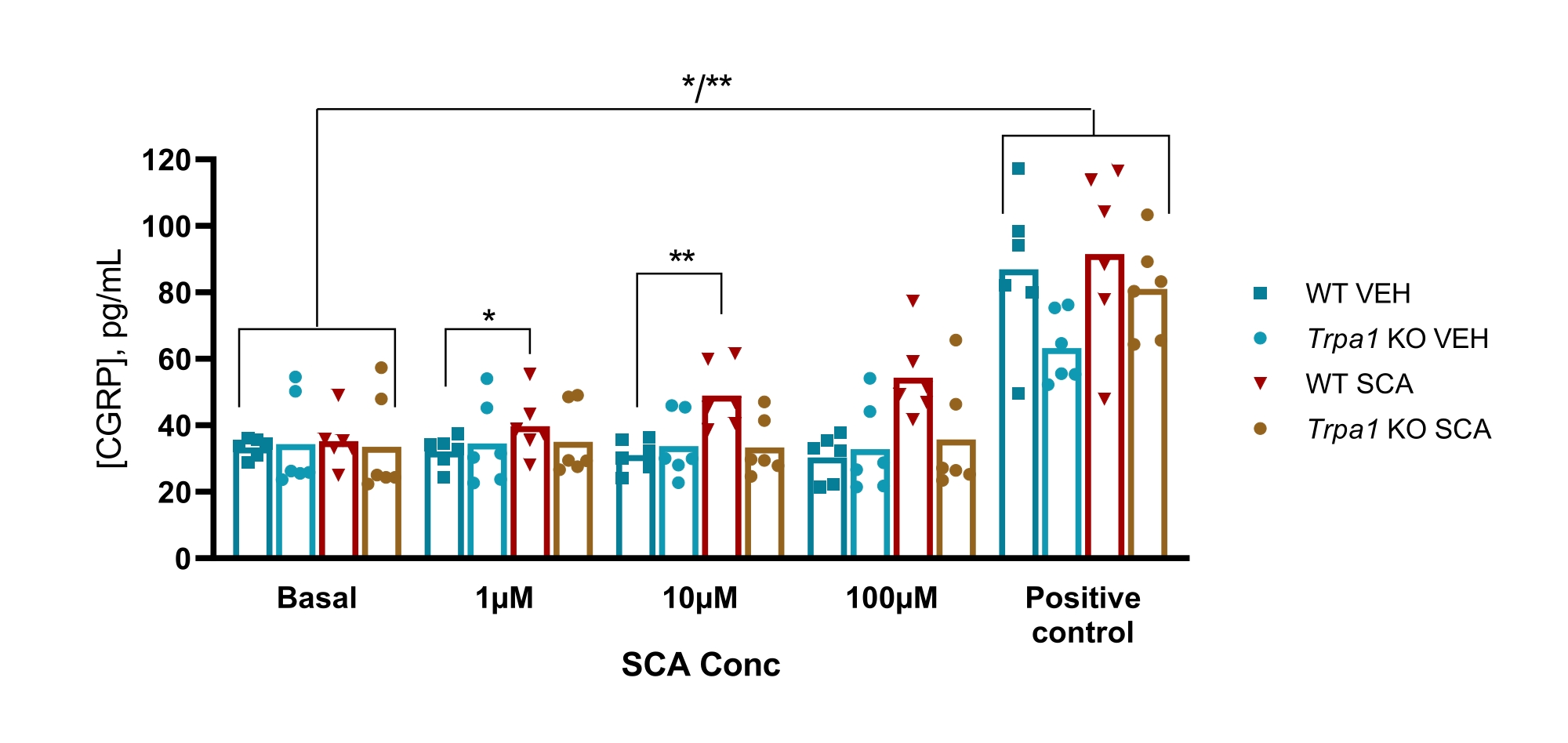
In mice, the protocol was used to examine the involvement of the transient receptor potential ankyrin 1 (TRPA1) ion channel in a GTN mouse model of migraine, where GTN-induced hypersensitivity was fully dependent on TRPA1 channels. The TRPA1 agonist supercinnamaldehyde (SCA) was found to release CGRP in a dose-dependent manner from the TG with 1 µM, 10 µM, and 100 µM of SCA, resulting in 9% (P = 0.23), 51% (P = 0.011), and 69% (P = 0.0097) increased release of CGRP compared to vehicle, respectively when analyzed with two-way ANOVA. This release was absent in TG from Trpa1 null mice where exposure to 1 µM, 10 µM, and 100 µM of SCA resulted in 11% (P > 0.99), -13% (P > 0.99) and 9% (P = 0.97) percent change in the release of CGRP compared to vehicle, respectively when analyzed with two-way ANOVA. The subsequent stimulation with 10 µM of capsaicin (positive control) shows that all tissue samples could release CGRP (Figure 6)50.

Figure 1: Step-by-step dissection of tissue from rats. The (A-T) details are provided in the protocol section (step 3.1). Please click here to view a larger version of this figure.
{kind=link}

Figure 2: Step-by-step dissection of tissue from mice. The (A-T) details are provided in the protocol section (step 3.2). Please click here to view a larger version of this figure.
{kind=link}

Figure 3: Washing and incubation of rodent tissue. (A-B) Freshly isolated tissue in plastic containers with SIF. The containers are covered with tulle to enable easy change of SIF. (C) Rat tissue - Two skull halves with dura mater covering the inner linings of the skull placed on a 6-well culture plate. The right and left trigeminal nucleus caudalis in separate microcentrifuge tube lids (top row of lids). The two trigeminal ganglia in individual microcentrifuge tube lids (bottom row of lids). (D) Mouse tissue - The trigeminal nucleus caudalis-containing part of the brainstem in a separate microcentrifuge tube lid (top). Two mouse trigeminal ganglia are in one microcentrifuge tube lid (bottom). Please click here to view a larger version of this figure.
{kind=link}

Figure 4: Study designs for CGRP release. Three different protocols for performing CGRP release experiments. Drugs are to be diluted in synthetic interstitial fluid (SIF). (A) Single stimulation. (B) Concentration-response stimulation. (C) Inhibition of a stimulation. Please click here to view a larger version of this figure.
{kind=link}

Figure 5: Glibenclamide inhibits capsaicin-induced CGRP release from rat dura mater and trigeminal ganglion. CGRP levels from trigeminal ganglion and dura mater isolated from female spontaneous trigeminal allodynic (STA) rats (215-318 g) originally sourced from the Thomas Jefferson University52 were measured with commercial human CGRP EIA kits. (A-B) CGRP release from (A) dura mater and (B) trigeminal ganglion after increasing concentrations of capsaicin (10 nM, 100 nM, 1 µM and 10 µM) (n = 4). Data are presented as individual points and were analyzed with two-way ANOVA. ****p < 0.0001 (C-D) Levels of CGRP released from (C) dura mater and (D) trigeminal ganglion after 10 min exposure to vehicle, 3 µM glibenclamide (glib), 1 µM capsaicin and 1 µM capsaicin + 3 µM glib (n = 6-11). Data are presented as individual points and mean values and were analyzed with one-way ANOVA. *compared to vehicle. #capsaicin compared to capsaicin + glib. #P < 0.05, ** and ##P < 0.01, ****P < 0.0001. The analyses were followed with Bonferroni's multiple comparison test. A significant level of α = 0.05 was used for all the tests. This figure has been modified from Christensen et al.51. Please click here to view a larger version of this figure.
{kind=link}

Figure 6: SCA exposure results in TRPA1 dependent CGRP release from mouse trigeminal ganglion. CGRP levels released from trigeminal ganglion isolated from WT and Trpa1 null (Trpa1tm1/Dpc)53 male mice (8-10 weeks) were measured with rat CGRP EIA kits after exposure to supercinnamaldehyde (SCA) at 1 µM, 10 µM, and 100 µM and capsaicin at 10 µM as a positive control (n = 6). Data from each mouse is presented as individual points, and bars indicate the mean values. Statistics: Comparison between SCA and vehicle at each concentration and between basal and positive control was performed with a two-way repeated ANOVA. A significant level of α = 0.05. *P < 0.05, **P < 0.01. This figure has been modified from Christensen et al.50. Please click here to view a larger version of this figure.
{kind=link}
Discussion
The method described was developed following studies showing the importance of CGRP in migraine pathophysiology. It is well suited for investigating the mechanisms involved in the release of CGRP from the trigeminovascular system, which is crucial for pain signaling in the head region. The amount of CGRP obtained in this model directly measures CGRP release from the trigeminal nerves innervating dura mater, TG, and TNC. The amount of CGRP release is quantitatively larger45,54 than the release measured in plasma after thermocoagulation in humans, trigeminal stimulation in cat39,55,56, and during migraine attacks23. One explanation could be that CGRP is diluted and degraded in the blood54. However, it is to be noted that direct stimulation with chemicals might be superior to pathophysiological activation. Further advantages are that it is possible to locate the release from three different sites within the trigeminovascular system and that it can be used together with pharmacological manipulation and in tissue from genetically modified rodents.
Lately, many preclinical rodent models focus on systemic administration of substances and subsequent pain or migraine-related read-outs using von Frey testing57,58, grimacing59,60,61 or light aversion62,63,64. These methods are useful in understanding the pain-inducing and pain-relieving properties of different substances. However, these approaches give no information on specific target tissues involved. In the present method, the trigeminovascular system is divided into three structures: the dura mater, TG, and TNC. This enables local exposure of each structure and the assessment of the location of action of a specific substance. This was utilized in a study from 2011, where the role of voltage-gated calcium channels in rats was explored, and the inhibition of these channels was found to be different in the three structures of the trigeminovascular pathway47. When dissecting structures of the nervous system, axotomy is unavoidable. Axotomy has been shown to alter the transcription of various genes65. These transcriptional changes are too slow to impact results from this method, but changes in phosphorylation cannot be excluded when compared to in vivo situations46. Although neuropeptides such as CGRP are formed in the cell soma, the release and action of neuropeptides are usually at the central or peripheral nerve terminals. Thus, studies of intact neurons, including terminals, are interesting when studying neuropeptide release. Therefore, methods of studying cultures of neurons from isolated ganglia have been established to serve as a model of the terminals. However, neuronal cell cultures are subject to several problems as mechanical dissociation can destroy neurons in a culture46. The longer time frame associated with culturing of cells leaves this method sensitive to transcriptomic changes due to axotomy and culture conditions65. Furthermore, the addition of growth factors and culturing on surface coatings have altered neuronal properties as transmitter and receptor expression66,67,68,69. These problems are avoided when studying freshly isolated intact ganglia instead of neuronal cell cultures.
One challenge with the ex vivo CGRP release method is the precise dissection of the tissue required for reproducible results. Especially accurate dissection of the TNC is challenging as this is a structure within the brainstem with no visible borders. Furthermore, the dura mater is fragile, and removal of the brain needs to be carefully carried out to ensure an intact structure. These obstacles can result in varying tissue size and, thus, varying basal and stimulation-induced CGRP levels. However, this variation can be accounted for by normalization to the basal CGRP release. It should also be noted that when isolating the TNC from mice, the entire lower part of the brainstem is isolated and not the more specific TNC-containing part as done in rats. In general, it can be an advantage to use rat tissue, as this allows measurement of CGRP release from dura mater and more precise dissection of the TNC. Furthermore, the size of the tissue also enables the use of a rat as its vehicle control, as one rat results in two cranium halves, two TGs, and two TNCs where one piece of the tissue is used for substance stimulation and the other for the vehicle. When using mice, two animals are needed for one experiment as both TGs are pooled in one sample, and the TNCs are dissected as one brainstem. Therefore, two TGs and one brainstem are used for substance stimulation, and two TGs and one brainstem from another mouse are used for vehicle control. This results in using twice as many mice compared to rats to obtain the same number of replicates. To reduce the number of mice used, a method to measure CGRP release from brainstem slices has been suggested49. It is an advantage that the method has been modified to enable the use of mice. This allows the use of many already available transgenic mouse strains, a useful tool for studying, e.g., signaling pathways. A positive control at the end of an experiment should be included to ensure that the tissue used in the experiment can release CGRP. The positive control could be TRPV1 agonist capsaicin or the depolarizing stimulus potassium (KCl), which have been found to release CGRP from the trigeminovascular system in both mice and rats46,47,48,49,50. Furthermore, the method has also been adapted to measure the release of other relevant peptides as pituitary adenylate cyclase-activating peptide (PACAP) - another peptide of great interest within migraine research70.
The method provides a useful tool for investigating CGRP release from specific target tissues in rats and mice. It is a relatively fast method that avoids problems associated with culturing neurons. The method protocol can easily be modified to study the concentration-response relationship or inhibition of a response by various pharmacological compounds. The ex vivo CGRP release method is one of the several preclinical methods useful for studying the role of CGRP and other mechanisms related to CGRP release in migraine pathophysiology.
Disclosures
The authors have nothing to disclose.
Acknowledgements
This work was funded by Candys Foundation.
Materials
| Name | Company | Catalog Number | Comments |
| 6-well culture plate | NUNC | 140675 | |
| Calcium chloride dihydrate | Merck | 1.02382.1000 | For SIF buffer |
| Caps for plastic containers | ThermoFisher Scientific | 536617 | |
| Capsaicin | Merck | M2028 | |
| CGRP kits | AH Diagnostics | A05482.96 | |
| CO2 | Strandmøllen | 4.6 | For carbogen gassing of SIF |
| Delicate Bone Trimmer | Fine Science Tools | 16109-14 | |
| Glibenclamide | Tocris | 911 | |
| Glucose | Merck | G7021 | For SIF buffer |
| Guillotine for rats | Scandidact | NS-802 | |
| Magnesium sulfate heptahydrate | Merck | M5921 | For SIF buffer |
| Microcenrifuge tubes + lids/caps | VWR | 700-5239 | |
| Mini Hacksaw | BAHCO | 208 | |
| O2 | Strandmøllen | 4.5 | For carbogen gassing of SIF |
| Pentobarbital | Glostrup pharmacy | NA | Magistral formula |
| Plastic containers | ThermoFisher Scientific | 536455 | |
| Plate photometer - Infinite M200 | Tecan | NA | Infinite M200 is discontinued. A Infinite 200 PRO is available at Tecan. Software: SW Magellan v.6.3 |
| Potassium chloride | Merck | P9333 | For SIF buffer |
| Scissor | Allgaier Instruments | 307-156-170 | |
| Small scissor | Allgaier Instruments | 04-520-115 | |
| Sodium bicarbonate | Merck | S6014 | For SIF buffer |
| Sodium chloride | Merck | S9888 | For SIF buffer |
| Sodium dihydrogen phosphate monohydrate | Merck | 1.06346.1000 | For SIF buffer |
| Sodium gluconate | Merck | S2054 | For SIF buffer |
| Spatula | Bochem Lab Supply | 3018 | |
| Spring scissor | Fine Science Tools | 15024-10 | |
| Sucrose | Merck | 84097 | For SIF buffer |
| Supercinnamaldehyde | Merck | S3322 | |
| Tulle (fabrics) | NA | NA | Bought in the local fabrics store |
References
- GBD 2016 Collaborators. Global, regional, and national incidence, prevalence, and years lived with disability for 328 diseases and injuries for 195 countries, 1990-2016: A systematic analysis for the Global Burden of Disease Study. The Lancet. 390 (10100), 1211-1259 (2017).
- Dodick, D. W. CGRP ligand and receptor monoclonal antibodies for migraine prevention: Evidence review and clinical implications. Cephalalgia: An International Journal of Headache. 39 (3), 445-458 (2019).
- Sacco, S., et al. European headache federation guideline on the use of monoclonal antibodies acting on the calcitonin gene related peptide or its receptor for migraine prevention. The Journal of Headache and Pain. 20 (1), 6 (2019).
- Moreno-Ajona, D., Pérez-Rodríguez, A., Goadsby, P. J. Gepants, calcitonin-gene-related peptide receptor antagonists: what could be their role in migraine treatment. Current Opinion in Neurology. 33 (3), 309-315 (2020).
- Khan, S., Olesen, A., Ashina, M. CGRP, a target for preventive therapy in migraine and cluster headache: Systematic review of clinical data. Cephalalgia: An International Journal of Headache. 39 (3), 374-389 (2019).
- Edvinsson, L., Haanes, K. A., Warfvinge, K., Krause, D. N. CGRP as the target of new migraine therapies - successful translation from bench to clinic. Nature Reviews Neurology. 14 (6), 338 (2018).
- Goadsby, P. J., et al. Pathophysiology of migraine: A disorder of sensory processing. Physiological Reviews. 97 (2), 553-622 (2017).
- Amara, S. G., Jonas, V., Rosenfeld, M. G., Ong, E. S., Evans, R. M. Alternative RNA processing in calcitonin gene expression generates mRNAs encoding different polypeptide products. Nature. 298 (5871), 240-244 (1982).
- Rosenfeld, M. G., et al. Production of a novel neuropeptide encoded by the calcitonin gene via tissue-specific RNA processing. Nature. 304 (5922), 129-135 (1983).
- Edvinsson, L., Goadsby, P. J. Discovery of CGRP in relation to migraine. Cephalalgia: An International Journal of Headache. 39 (3), 331-332 (2019).
- Brain, S. D., Williams, T. J., Tippins, J. R., Morris, H. R., MacIntyre, I. Calcitonin gene-related peptide is a potent vasodilator. Nature. 313 (5997), 54-56 (1985).
- Fisher, L. A., et al. Stimulation of noradrenergic sympathetic outflow by calcitonin gene-related peptide. Nature. 305 (5934), 534-536 (1983).
- Hanko, J., Hardebo, J. E., Kåhrström, J., Owman, C., Sundler, F. Calcitonin gene-related peptide is present in mammalian cerebrovascular nerve fibres and dilates pial and peripheral arteries. Neuroscience Letters. 57 (1), 91-95 (1985).
- Uddman, R., Edvinsson, L., Ekblad, E., Håkanson, R., Sundler, F. Calcitonin gene-related peptide (CGRP): perivascular distribution and vasodilatory effects. Regulatory Peptides. 15 (1), 1-23 (1986).
- Edvinsson, L. Functional role of perivascular peptides in the control of cerebral circulation. Trends in Neurosciences. 8, 126-131 (1985).
- Edvinsson, L., Ekman, R., Jansen, I., Ottosson, A., Uddman, R. Peptide-containing nerve fibers in human cerebral arteries: Immunocytochemistry, radioimmunoassay and in vitro pharmacology. Annals of Neurology. 21 (5), 431-437 (1987).
- Edvinsson, L., Fredholm, B. B., Hamel, E., Jansen, I., Verrecchia, C. Perivascular peptides relax cerebral arteries concomitant with stimulation of cyclic adenosine monophosphate accumulation or release of an endothelium-derived relaxing factor in the cat. Neuroscience Letters. 58 (2), 213-217 (1985).
- Brain, S. D., Grant, A. D. Vascular actions of calcitonin gene-related peptide and adrenomedullin. Physiological Reviews. 84 (3), 903-934 (2004).
- Skofitsch, G., Jacobowitz, D. M. Calcitonin gene-related peptide: Detailed immunohistochemical distribution in the central nervous system. Peptides. 6 (4), 721-745 (1985).
- Suzuki, N., Hardebo, J. E., Owman, C. Origins and pathways of cerebrovascular nerves storing substance P and calcitonin gene-related peptide in rat. Neuroscience. 31 (2), 427-438 (1989).
- Uddman, R., Edvinsson, L., Ekman, R., Kingman, T., McCulloch, J. Innervation of the feline cerebral vasculature by nerve fibers containing calcitonin gene-related peptide: trigeminal origin and co-existence with substance P. Neuroscience Letters. 62 (1), 131-136 (1985).
- Warfvinge, K., Edvinsson, L. Distribution of CGRP and CGRP receptor components in the rat brain. Cephalalgia: An International Journal of Headache. 39 (3), 342-353 (2019).
- Goadsby, P. J., Edvinsson, L., Ekman, R. Vasoactive peptide release in the extracerebral circulation of humans during migraine headache. Annals of Neurology. 28 (2), 183-187 (1990).
- Lassen, L. H., et al. CGRP may play a causative role in migraine. Cephalalgia: An International Journal of Headache. 22 (1), 54-61 (2002).
- Olesen, J., et al. Calcitonin gene-related peptide receptor antagonist BIBN 4096 BS for the acute treatment of migraine. New England Journal of Medicine. 350 (11), 1104-1110 (2004).
- Eftekhari, S., et al. Localization of CGRP, CGRP receptor, PACAP and glutamate in trigeminal ganglion. Relation to the blood-brain barrier. Brain Research. 1600, 93-109 (2015).
- Keller, J. T., Marfurt, C. F. Peptidergic and serotoninergic innervation of the rat dura mater. Journal of Comparative Neurology. 309 (4), 515-534 (1991).
- Edvinsson, L., et al. Innervation of the human middle meningeal artery: immunohistochemistry, ultrastructure, and role of endothelium for vasomotility. Peptides. 19 (7), 1213-1225 (1998).
- Lennerz, J. K., et al. Calcitonin receptor-like receptor (CLR), receptor activity-modifying protein 1 (RAMP1), and calcitonin gene-related peptide (CGRP) immunoreactivity in the rat trigeminovascular system: Differences between peripheral and central CGRP receptor distribution. Journal of Comparative Neurology. 507 (3), 1277-1299 (2008).
- Eftekhari, S., Edvinsson, L. Calcitonin gene-related peptide (CGRP) and its receptor components in human and rat spinal trigeminal nucleus and spinal cord at C1-level. BMC Neuroscience. 12, 112 (2011).
- Eftekhari, S., et al. Differential distribution of calcitonin gene-related peptide and its receptor components in the human trigeminal ganglion. Neuroscience. 169 (2), 683-696 (2010).
- Eftekhari, S., Warfvinge, K., Blixt, F. W., Edvinsson, L. Differentiation of nerve fibers storing CGRP and CGRP receptors in the peripheral trigeminovascular system. The Journal of Pain. 14 (11), 1289-1303 (2013).
- Spitzer, M. J. S., Reeh, P. W., Sauer, S. K. Mechanisms of potassium- and capsaicin-induced axonal calcitonin gene-related peptide release: Involvement of L- and T-type calcium channels and TRPV1 but not sodium channels. Neuroscience. 151 (3), 836-842 (2008).
- Evans, A. R., Nicol, G. D., Vasko, M. R. Differential regulation of evoked peptide release by voltage-sensitive calcium channels in rat sensory neurons. Brain Research. 712 (2), 265-273 (1996).
- Gupta, S., Villalón, C. M. The relevance of preclinical research models for the development of antimigraine drugs: Focus on 5-HT 1B/1D and CGRP receptors. Pharmocology & Therapeutics. 128 (1), 170-190 (2010).
- Williamson, D. J., Hargreaves, R. J., Hill, R. G., Shepheard, S. L. Intravital microscope studies on the effects of neurokinin agonists and calcitonin gene-related peptide on dural vessel diameter in the anaesthetized rat. Cephalalgia: An International Journal of Headache. 17 (4), 518-524 (1997).
- Gupta, S., Bhatt, D. K., Boni, L. J., Olesen, J. Improvement of the closed cranial window model in rats by intracarotid infusion of signalling molecules implicated in migraine. Cephalalgia: An International Journal of Headache. 30 (1), 27-36 (2010).
- Knight, Y. E., Edvinsson, L., Goadsby, P. J. Blockade of calcitonin gene-related peptide release after superior sagittal sinus stimulation in cat: a comparison of avitriptan and CP122, 288. Neuropeptides. 33 (1), 41-46 (1999).
- Goadsby, P. J., Edvinsson, L., Ekman, R. Release of vasoactive peptides in the extracerebral circulation of humans and the cat during activation of the trigeminovascular system. Annals of Neurology. 23 (2), 193-196 (1988).
- Tassorelli, C., Joseph, S. A. Systemic nitroglycerin induces Fos immunoreactivity in brainstem and forebrain structures of the rat. Brain Research. 682 (1-2), 167-181 (1995).
- Ramachandran, R., et al. A naturalistic glyceryl trinitrate infusion migraine model in the rat. Cephalalgia: An International Journal of Headache. 32 (1), 73-84 (2012).
- Hoskin, K. L., Zagami, A. S., Goadsby, P. J. Stimulation of the middle meningeal artery leads to Fos expression in the trigeminocervical nucleus: a comparative study of monkey and cat. Journal of Anatomy. 194, 579-588 (1999).
- Charbit, A. R., Akerman, S., Goadsby, P. J. Comparison of the Effects of Central and Peripheral Dopamine Receptor Activation on Evoked Firing in the Trigeminocervical Complex. Journal of Pharmacology and Experimental Therapeutics. 331 (2), 752-763 (2009).
- Koulchitsky, S., Fischer, M., Messlinger, K. Calcitonin gene-related peptide receptor inhibition reduces neuronal activity induced by prolonged increase in nitric oxide in the rat spinal trigeminal nucleus. Cephalalgia: An International Journal of Headache. 29 (4), 408-417 (2009).
- Ebersberger, A., Averbeck, B., Messlinger, K., Reeh, P. W. Release of substance P, calcitonin gene-related peptide and prostaglandin E2 from rat dura mater encephali following electrical and chemical stimulation in vitro. Neuroscience. 89 (3), 901-907 (1999).
- Eberhardt, M., et al. Calcitonin gene-related peptide release from intact isolated dorsal root and trigeminal ganglia. Neuropeptides. 42 (3), 311-317 (2008).
- Amrutkar, D. V., Ploug, K. B., Olesen, J., Jansen-Olesen, I. Role for voltage gated calcium channels in calcitonin gene-related peptide release in the rat trigeminovascular system. Neuroscience. 172, 510-517 (2011).
- Gupta, S., et al. Evidence for CGRP re-uptake in rat dura mater encephali. British Journal of Pharmacology. 161 (8), 1885-1898 (2010).
- Kageneck, C., Nixdorf-Bergweiler, B. E., Messlinger, K., Fischer, M. J. M. Release of CGRP from mouse brainstem slices indicates central inhibitory effect of triptans and kynurenate. Journal of Headache and Pain. 15 (1), 1-9 (2014).
- Christensen, S. L., et al. CGRP-dependent signalling pathways involved in mouse models of GTN- cilostazol- and levcromakalim-induced migraine. Cephalalgia: An International Journal of Headache. 41 (14), 1413-1426 (2021).
- Christensen, S. L., et al. ATP sensitive potassium (KATP) channel inhibition: A promising new drug target for migraine. Cephalalgia: An International Journal of Headache. 40 (7), 650-664 (2020).
- Oshinsky, M. L., et al. Spontaneous trigeminal allodynia in rats: A model of primary headache. Headache. 52 (9), 1336 (2012).
- Kwan, K. Y., et al. TRPA1 contributes to cold, mechanical, and chemical nociception but is not essential for hair-cell transduction. Neuron. 50 (2), 277-289 (2006).
- Eltorp, C., Jansen-Olesen, I., Hansen, A. J. Release of calcitonin gene-related peptide (CGRP) from guinea pig dura mater in vitro is inhibited by sumatriptan but unaffected by nitric oxide. Cephalalgia: An International Journal of Headache. 20 (9), 838-844 (2000).
- Zagami, A. S., Goadsby, P. J., Edvinsson, L. Stimulation of the superior sagittal sinus in the cat causes release of vasoactive peptides. Neuropeptides. 16 (2), 69-75 (1990).
- Goadsby, P. J., Edvinsson, L. The trigeminovascular system and migraine: studies characterizing cerebrovascular and neuropeptide changes seen in humans and cats. Annals of Neurology. 33 (1), 48-56 (1993).
- Bates, E. A., et al. Sumatriptan alleviates nitroglycerin-induced mechanical and thermal allodynia in mice. Cephalalgia: An International Journal of Headache. 30 (2), 170-178 (2010).
- Pradhan, A. A., et al. Characterization of a novel model of chronic migraine. Pain. 155 (2), 269-274 (2014).
- Mogil, J. S., Pang, D. S. J., Silva Dutra, G. G., Chambers, C. T. The development and use of facial grimace scales for pain measurement in animals. Neuroscience and Biobehavioral Reviews. 116, 480-493 (2020).
- Sotocinal, S. G., et al. The rat grimace scale: A partially automated method for quantifying pain in the laboratory rat via facial expressions. Molecular Pain. 7, 55 (2011).
- Langford, D. J., et al. Coding of facial expressions of pain in the laboratory mouse. Nature Methods. 7 (6), 447-449 (2010).
- Mahmoudi, J., et al. Cerebrolysin attenuates hyperalgesia, photophobia, and neuroinflammation in a nitroglycerin-induced migraine model in rats. Brain Research Bulletin. 140, 197-204 (2018).
- Farajdokht, F., Babri, S., Karimi, P., Mohaddes, G. Ghrelin attenuates hyperalgesia and light aversion-induced by nitroglycerin in male rats. Neuroscience Letters. 630, 30-37 (2016).
- Kuburas, A., et al. PACAP Induces Light Aversion in Mice by an Inheritable Mechanism Independent of CGRP. Journal of Neuroscience. 41 (21), 4697-4715 (2021).
- Buschmann, T., et al. Expression of Jun, Fos, and ATF-2 proteins in axotomized explanted and cultured adult rat dorsal root ganglia. Neuroscience. 84 (1), 163-176 (1998).
- Lee, Y. J., Zachrisson, O., Tonge, D. A., McNaughton, P. A. Upregulation of bradykinin B2 receptor expression by neurotrophic factors and nerve injury in mouse sensory neurons. Molecular and Cellular Neuroscience. 19 (2), 186-200 (2002).
- Hari, A., Djohar, B., Skutella, T., Montazeri, S. Neurotrophins and extracellular matrix molecules modulate sensory axon outgrowth. International Journal of Developmental Neuroscience. 22 (2), 113-117 (2004).
- Skoff, A. M., Resta, C., Swamydas, M., Adler, J. E. Nerve Growth Factor (NGF) and Glial Cell Line-Derived Neurotrophic Factor (GDNF) regulate substance P release in adult spinal sensory neurons. Neurochemical Research. 28 (6), 847-854 (2003).
- Lindsay, R. M., Harmar, A. J. Nerve growth factor regulates expression of neuropeptide genes in adult sensory neurons. Nature. 337 (6205), 362-364 (1989).
- Edvinsson, J. C. A., et al. Differences in pituitary adenylate cyclase-activating peptide and calcitonin gene-related peptide release in the trigeminovascular system. Cephalalgia: An International Journal of Headache. 40 (12), 1296-1309 (2020).
Reprints and Permissions
Request permission to reuse the text or figures of this JoVE article
Request PermissionThis article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. All rights reserved