
Method Article
Ex Vivo Rilascio di peptide correlato al gene della calcitonina dal sistema trigeminovascolare nei roditori
In questo articolo
Riepilogo
Il presente protocollo descrive il modello di rilascio ex vivo del peptide correlato al gene della calcitonina (CGRP) e la strategia per quantificare l'effetto degli agenti farmacologici sulla quantità di CGRP rilasciata dal sistema trigeminovascolare nei roditori.
Abstract
Il peptide correlato al gene della calcitonina (CGRP) è stato scoperto per la prima volta nel 1980 come variante di giunzione dal gene della calcitonina. Fin dalla sua scoperta, il suo ruolo nella fisiopatologia dell'emicrania è stato ben stabilito, prima dalle sue potenti proprietà vasodilatatorie e successivamente dalla sua presenza e funzione come neurotrasmettitore nel sistema trigeminovascolare sensoriale. La capacità di provocare l'emicrania di CGRP ha dato supporto all'industria farmaceutica per sviluppare anticorpi monoclonali e antagonisti che inibiscono l'effetto di CGRP. Un nuovo paradigma di trattamento si è dimostrato efficace nel trattamento profilattico dell'emicrania. Uno degli strumenti utili per comprendere ulteriormente i meccanismi dell'emicrania è il modello ex vivo di rilascio di CGRP dal sistema trigeminovascolare. È un metodo relativamente semplice che può essere utilizzato con vari strumenti farmacologici per ottenere know-how per sviluppare ulteriormente nuovi trattamenti efficaci per l'emicrania. Il presente protocollo descrive un modello di rilascio di CGRP e la tecnica per quantificare l'effetto degli agenti farmacologici sulla quantità di CGRP rilasciata dal sistema trigeminovascolare nei roditori. Viene fornita una procedura che descrive l'approccio sperimentale dall'eutanasia alla misurazione dei livelli proteici. L'isolamento essenziale del ganglio trigemino e del nucleo caudale trigemino sia dai topi che dai ratti e la preparazione della dura madre del ratto sono descritti in dettaglio. Inoltre, vengono presentati risultati rappresentativi di entrambe le specie (ratti e topi). La tecnica è uno strumento chiave per studiare i meccanismi molecolari coinvolti nella fisiopatologia dell'emicrania utilizzando vari composti farmacologici e animali geneticamente modificati.
Introduzione
L'emicrania è un disturbo neurologico che, secondo l'OMS, si stima colpisca più di 1 miliardo di persone ed è una delle principali cause di disabilità in tutto il mondo1. Pertanto, l'emicrania ha un impatto significativo sia sui pazienti che sulla società. Nonostante il recente successo clinico dei farmaci antagonisti CGRP, una grande percentuale di pazienti ha bisogno di migliori opzioni di trattamento 2,3,4,5. È necessario chiarire la fisiopatologia dell'emicrania che porta a nuovi trattamenti efficaci. La segnalazione all'interno del sistema trigeminovascolare costituito dalle meningi, dai gangli del trigemino (TG) e dal nucleo caudale del trigemino (TNC) è centrale per la fisiopatologia dell'emicrania 6,7.
Il peptide 37 aminoacido neuropeptide calcitonina genico correlato (CGRP) è stato scoperto per la prima volta nei primi anni 1980 quando Amara e collaboratori hanno dimostrato che la trascrizione primaria dell'RNA del gene della calcitonina potrebbe essere elaborata per dare la codifica dell'mRNA per CGRP oltre alla calcitonina 8,9. La ricerca successiva ha suggerito un collegamento con la fisiopatologia dell'emicrania10. CGRP è un neurotrasmettitore con potenti proprietà vasodilatatorie 11,12,13,14,15,16,1 7, ed è ampiamente distribuito nel sistema nervoso centrale e periferico 13,14,18,19,20,21,22 . Il coinvolgimento di CGRP nell'emicrania è stato sottolineato con la scoperta di un aumento dei livelli di CGRP nella circolazione extracerebrale durante gli attacchi di emicrania negli esseri umani23 e che l'infusione di CGRP provoca dolore simile all'emicrania nei pazienti24. Due anni dopo, il primo studio proof-of-concept sull'efficacia dell'antagonista CGRP olcegepant nel trattamento dell'emicrania è stato pubblicato25.
La CGRP è abbondante nel sistema trigeminovascolare come dimostrato nel TG21,26, nelle fibre nervose sensoriali che innervano la dura madre27,28,29 e nella TNC30. Nel sistema trigeminovascolare, CGRP si trova nei neuroni di piccole e medie dimensioni del TG, nelle fibre C non mielinizzate, ed è espresso in quasi il 50% della popolazione neuronale del TG. Il recettore CGRP è espresso principalmente nei neuroni più grandi e si trova nelle fibre Aδ mieliniche31,32. CGRP viene rilasciato dai neuroni dopo stimolazione chimica o elettrica33,34. Gli studi sui percorsi che portano al rilascio di CGRP e la posizione di questa attivazione sono cruciali per comprendere la fisiopatologia dell'emicrania. Nel corso degli ultimi 5 decenni, gli studi preclinici hanno contribuito ad acquisire ampie conoscenze sulla segnalazione correlata all'emicrania e hanno contribuito allo sviluppo di nuovi trattamenti35. Molti metodi che considerano il coinvolgimento vascolare e neurogeno sono stati modificati e applicati nella ricerca sull'emicrania. Si possono citare modelli in vivo e in vitro di risposte arteriose a composti biologici o trattamenti farmacologici17,36,37 e stimolazione nervosa elettrica 38,39. Inoltre, i neuroni attivati nella TNC possono essere rilevati dall'espressione di c-Fos 40,41,42 e dalle registrazioni elettrofisiologiche in quest'area 43,44. Entrambi i metodi misurano i segnali nocicettivi trasmessi al cervello dalla testa, ad esempio la dura madre. L'uso di un solo modello preclinico non presenta il quadro completo della fisiopatologia dell'emicrania. Pertanto, è importante combinare diversi modelli che coprano il maggior numero possibile di aspetti della fisiopatologia dell'emicrania. Il continuo sviluppo di nuovi modelli coprirà vari aspetti dei meccanismi dell'emicrania e nel tempo verrà scoperto il mistero della fisiopatologia dell'emicrania.
Qui, viene presentato un protocollo dettagliato del metodo di rilascio CGRP, eseguito ex vivo in TG e TNC isolati da topi dopo stimolazione chimica. Il rilascio di CGRP può anche essere studiato nella dura madre dai ratti. Pertanto, nel protocollo sperimentale per i ratti, la dura madre è descritta insieme a TG e TNC. La base per il metodo di rilascio CGRP è stata descritta per la prima volta nel 1999, quando Ebersberger e collaboratori hanno condotto ricerche pionieristiche e hanno scoperto che CGRP è stato rilasciato dalla dura madre dopo stimolazione chimica ed elettrica delle afferenze durali nei ratti45. Successivamente, questo approccio è stato esteso al rilascio CGRP dal TG46 e dal TNC47. Successivamente, il metodo è stato modificato per essere applicato a TG e TNC nei topi. Finora, il rilascio di CGRP dalla dura madre è stato impegnativo nei topi.
Protocollo
Tutte le procedure sperimentali e di cura degli animali sono state eseguite in conformità alla guida della Comunità Europea per la cura e l'uso degli animali (2010/63/UE). Topi maschi C57BL/6JBomTac, di 10 settimane, e ratti maschi Sprague Dawley, di 10 settimane, sono stati utilizzati per dimostrare questo protocollo.
1. Preparazione del liquido interstiziale sintetico
- Preparare il liquido interstiziale sintetico (SIF) secondo la seguente ricetta: 108 mM NaCl, 3,48 mM KCl, 3,50 mM MgSO 4, 26 mM NaHCO3, 11,70 mM NaH 2 PO4, 1,50 mM CaCl2, 9,60 mM Na-gluconato, 5,50 mM glucosio e 7,60 mM saccarosio (vedi tabella dei materiali).
NOTA: La SIF può essere variata a seconda del target di stimolazione, ad esempio, una soluzione priva di calcio può essere utilizzata quando si studiano i canali del calcio. - Regolare il pH a 7,4 e stabilizzare il pH mediante gassificazione del carbogeno (5% CO 2 e 95% O2)46.
2. Eutanasia
- Anestetizzare topi e ratti adulti con una miscela di 70% CO 2 e 30% O2. Decapitare topi usando un paio di forbici e ratti usando una ghigliottina (vedi Tabella dei materiali).
NOTA: Utilizzare un ceppo e un'età adatti allo scopo della ricerca. Sia i maschi che le femmine possono essere utilizzati in questo modello. - Separare la testa dal corpo a livello C3-C4 del midollo spinale.
NOTA: L'eutanasia può essere eseguita anche con un'iniezione intraperitoneale di pentobarbital (100-150 mg/kg).
3. Dissezione
- Preparare il tessuto di ratto seguendo i passaggi seguenti.
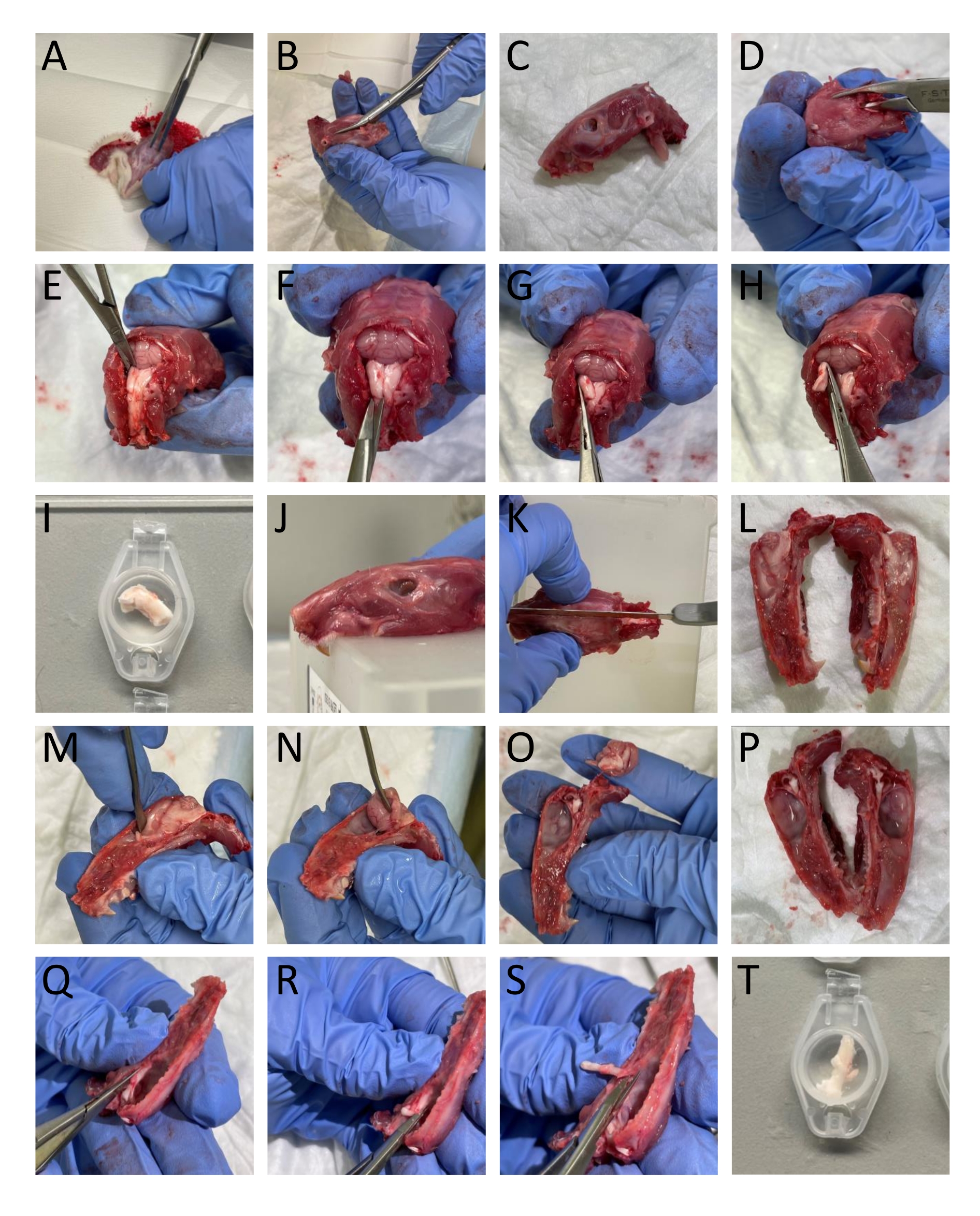
- Rimuovere la pelle e il muscolo intorno alla testa e al collo usando un paio di forbici (Figura 1A).
- Utilizzare un tagliaossa e forbici per separare le mascelle inferiori dalla testa (Figura 1B-C).
- Aprire il midollo spinale inserendo un trimmer osseo caudalmente nella parte dorsale delle vertebre e rimuovere la parte dorsale delle vertebre per esporre il midollo spinale e il tronco cerebrale (Figura 1D).
- Tagliare la parte caudale del cranio dai bordi delle ossa occipitale e interparietale per rimuovere queste strutture ossee che espongono il cervelletto (Figura 1E).
NOTA: È importante non danneggiare il tronco cerebrale e il midollo spinale durante il taglio e la rimozione delle vertebre. - Isolare la TNC (Sp5C) che scorre caudalmente a circa 13-16 mm dal bregma su ciascun lato tagliando la parte dorsolaterale del tronco encefalico con forbici a molla. Immergere il TNC sinistro e destro in SIF (Figura 1E-I).
NOTA: La descrizione corrisponde a ratti adulti. - Tagliare la testa a metà del conflitto per dividere il cranio in due usando una sega (Figura 1J-L).
- Rimuovere con cautela il cervello senza toccare la dura madre attaccata al cranio usando una spatola e tagliando il nervo trigemino dove entra nel tronco cerebrale (Figura 1M-P).
- Per isolare il TG, taglialo, compresi i suoi rami attorno ai confini visivi. Tagliare il ramo mandibolare dove entra nel forame ovale. Tagliare i rami oftalmici e mascellari entrando nel cranio in quanto non sono divisi macroscopicamente. Durante la dissezione del TG, rimuovere la dura madre che copre il TG (Figura 1Q-T).
- Immergere le metà del cranio e i TG in SIF.
- Preparare il tessuto del topo seguendo i passaggi seguenti.
- Rimuovere la pelle e i muscoli intorno alla testa e al collo usando piccole forbici (Figura 2A-B).
- Aprire il midollo spinale inserendo un paio di piccole forbici caudalmente nella parte dorsale delle vertebre e rimuovere la parte dorsale delle vertebre per esporre il midollo spinale e il tronco cerebrale (Figura 2C).
NOTA: È importante non danneggiare il midollo spinale durante il taglio e la rimozione delle vertebre. - Tagliare il cranio dai bordi delle ossa occipitale e interparietale per rimuovere queste strutture ossee che espongono il cervelletto (Figura 2D-F).
- Quindi, tagliare l'osso parietale a metà del sagitto e rimuovere l'osso per esporre il cervello (Figura 2G-I).
- Rimuovere con attenzione il cervelletto con una spatola per esporre il tronco cerebrale (Figura 2J).
- Isolare la parte del tronco cerebrale contenente TNC usando forbici a molla (Figura 2K-N). Immergere il tronco encefalico con TNC in SIF (Figura 2O).
- Rimuovere il cervello e tagliare il nervo trigemino dove entra nel tronco cerebrale (Figura 2P-Q).
- Per isolare il TG, taglialo, compresi i suoi rami attorno ai confini visivi. Tagliare il ramo mandibolare dove entra nel forame ovale. Tagliare i rami oftalmici e mascellari entrando nel cranio in quanto non sono divisi macroscopicamente. Durante la dissezione del TG, rimuovere la dura madre che copre il TG (Figura 2R-S).
- Immergere i TG nella SIF (Figura 2T).
4. Lavaggio
- Lavare le metà del cranio, le TNC e i TG in SIF per 30 minuti sostituendo il SIF ogni 5 minuti a temperatura ambiente.
NOTA: Le fasi di lavaggio possono essere eseguite mantenendo il tessuto in contenitori di plastica con un coperchio in tulle per consentire una facile sostituzione del SIF (Figura 3A-B). - Preparare il tessuto di ratto seguendo i passaggi seguenti.
- Trasferire le metà TNC in tappi di tubi per microcentrifuga separati con 350 μL di SIF (Figura 3C).
- Trasferire i TG ai tappi dei tubi della microcentrifuga - un TG per tappo del tubo della microcentrifuga con 350 μL di SIF (Figura 3C).
- Posizionare le metà del cranio su piattaforme di argilla o una piastra di coltura a 6 pozzetti e riempire il cranio con 400 μL di SIF (Figura 3C).
- Preparare il tessuto del topo seguendo i passaggi seguenti.
- Trasferire il tronco encefalico con TNC in un tappo a tubo per microcentrifuga con 250 μL di SIF (Figura 3D).
- Trasferire i due TG su un tappo a tubo per microcentrifuga con 250 μL di SIF (Figura 3D).
NOTA: Posizionare due TG per tappo del tubo della microcentrifuga quando si utilizza tessuto di topi.
- Collocare teschi di ratto e tappi di tubo di microcentrifuga con tessuto di ratto e topo in un'incubatrice umidificata a 37 °C. Sostituire SIF utilizzando una pipetta ogni 5 minuti per 20 minuti.
NOTA: è importante non toccare il tessuto quando si aggiunge e si rimuove SIF.
5. Test antidroga
- Determinare i livelli di rilascio basale di CGRP.
- Dopo l'ultimo lavaggio, aggiungere 250 μL di SIF al mouse TG e TNC. Aggiungere 350 μL a TG e TNC di ratto e 400 μL a ciascun cranio di ratto.
- Dopo 10 minuti di incubazione, raccogliere 200 μL del campione in una provetta da microcentrifuga e aggiungere 50 μL di tampone EIA 10x (fornito con il kit di dosaggio immunoenzimatico CGRP, vedere Tabella dei materiali) per consentire la misurazione del rilascio basale di CGRP (fase 6). Eliminare il liquido rimanente.
NOTA: Il tempo di incubazione deve essere lo stesso per tutti i campioni. - Conservare immediatamente i campioni a -20 °C.
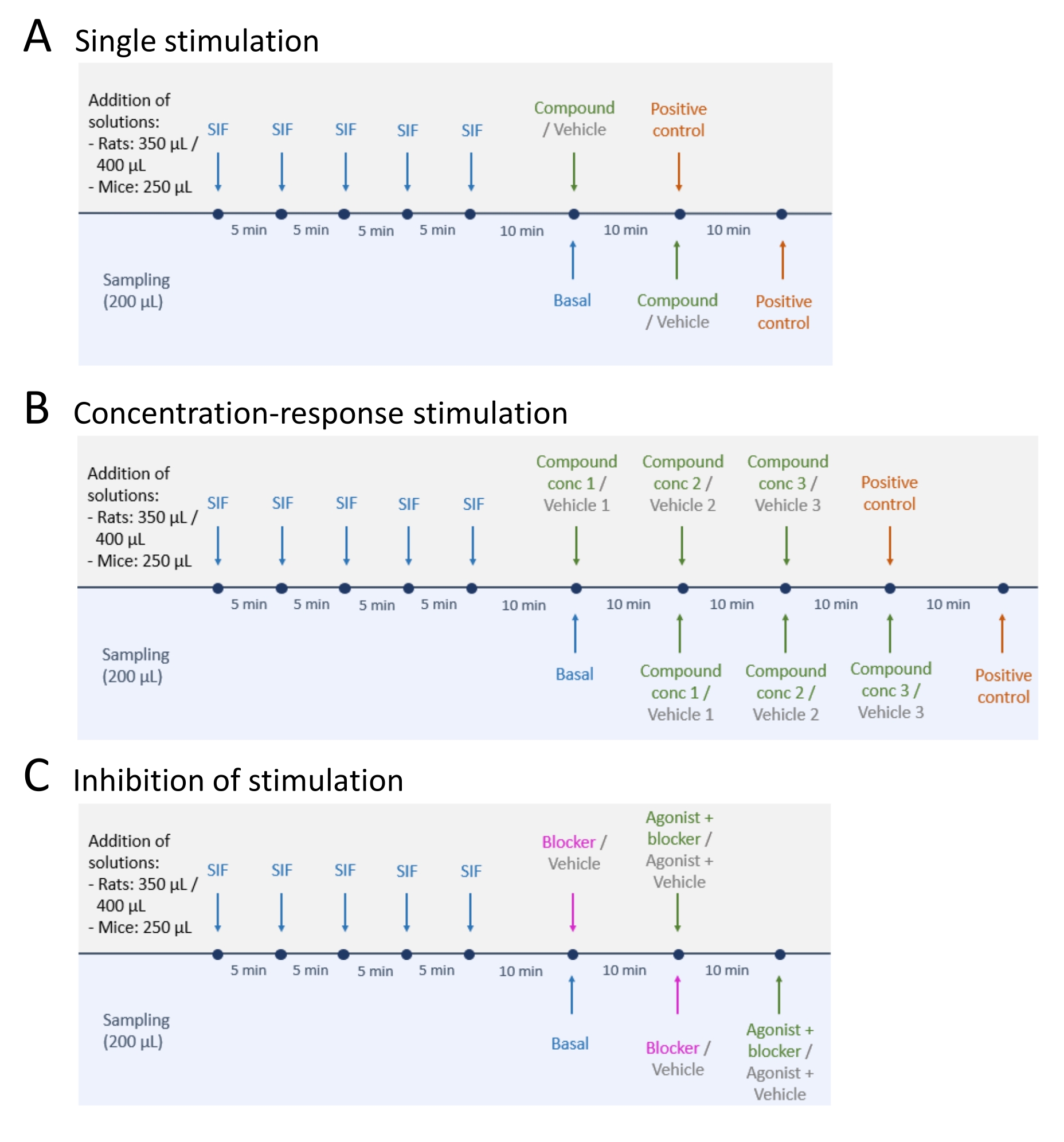
- Dopo il campionamento dei livelli di rilascio basale di CGRP, seguire uno dei tre metodi: (A) Stimolazione singola (fase 5.2); (B) Stimolazione concentrazione-risposta (fase 5.3); e (C) Inibizione della stimolazione (fase 5.4) (Figura 4).
NOTA: Il composto in esame e le concentrazioni utilizzate dipendono dallo scopo dello studio.
- Eseguire una singola stimolazione seguendo i passaggi seguenti.
- Aggiungere il composto in esame o il veicolo al tessuto e lasciarlo per 10 minuti (volume: 250 μL per il topo TG e TNC, 350 μL per il ratto TG e TNC e 400 μL per ciascun cranio di ratto).
- Dopo 10 minuti di incubazione, raccogliere 200 μL del campione in una provetta da microcentrifuga con 50 μL di tampone EIA 10x. Eliminare il liquido rimanente e conservare immediatamente i campioni a -20 °C.
NOTA: Il tempo di incubazione deve essere lo stesso per tutti i campioni.
- Eseguire la stimolazione concentrazione-risposta seguendo i passaggi seguenti.
- Diluire il composto in esame o il veicolo alle concentrazioni desiderate. Aggiungere il composto in esame in concentrazioni crescenti a partire dalla concentrazione più bassa.
NOTA: Il composto in esame e le concentrazioni utilizzate dipendono dallo scopo dello studio. Ad esempio, per il presente studio sono stati utilizzati 1 μM, 10 μM e 100 μM di supercinnamaldeide. - Aggiungere la concentrazione più bassa (1 μM per il presente studio) del composto in esame e del veicolo corrispondente a due preparati tissutali identici e incubare per 10 minuti (volume: 250 μL per il tessuto di topo, 350 μL per il ratto TG e TNC e 400 μL per ciascun cranio di ratto).
- Dopo 10 minuti di incubazione, raccogliere 200 μL del campione in una provetta da microcentrifuga con 50 μL di tampone EIA 10x.
- Eliminare il liquido rimanente e aggiungere la seconda concentrazione più bassa (10 μM per il presente studio) al tessuto.
- Conservare immediatamente i campioni a -20 °C.
- Ripetere questa procedura con le concentrazioni rimanenti (100 μM per il presente studio).
- Diluire il composto in esame o il veicolo alle concentrazioni desiderate. Aggiungere il composto in esame in concentrazioni crescenti a partire dalla concentrazione più bassa.
- Eseguire l'inibizione della stimolazione seguendo i passaggi seguenti.
- Aggiungere il bloccante o il veicolo al tessuto e incubare per 10 minuti (volume: 250 μL per il tessuto di topo, 350 μL per il ratto TG e TNC e 400 μL per ogni cranio di ratto).
NOTA: Il bloccante e la concentrazione utilizzati dipendono dallo scopo dello studio. Ad esempio, è stata utilizzata glibenclamide da 3 μM per il risultato rappresentativo nella Figura 5. - Dopo 10 minuti di incubazione, prelevare un campione da 200 μL in una provetta da microcentrifuga con 50 μL di tampone EIA 10x. Eliminare il liquido rimanente e conservare immediatamente i campioni a -20 °C.
- Aggiungere l'agonista o l'agonista + bloccante (vedi Tabella dei materiali) al tessuto e incubare per 10 minuti.
NOTA: L'agonista, il bloccante e le concentrazioni utilizzati dipendono dallo scopo dello studio. Nel presente studio sono stati utilizzati 3 μM di glibenclamide e 1 μM di capsaicina, Figura 5. - Dopo 10 minuti di incubazione, prelevare un campione da 200 μL in una provetta da microcentrifuga con 50 μL di tampone EIA 10x. Eliminare il liquido rimanente e conservare immediatamente i campioni a -20 °C.
- Aggiungere il bloccante o il veicolo al tessuto e incubare per 10 minuti (volume: 250 μL per il tessuto di topo, 350 μL per il ratto TG e TNC e 400 μL per ogni cranio di ratto).
- Eseguire un controllo positivo per l'esperimento.
- Quando appropriato, aggiungere un controllo positivo (ad esempio, 1-10 μM di capsaicina, vedere Tabella dei materiali) al tessuto alla fine del protocollo e raccogliere un campione di 200 μL in una provetta da microcentrifuga con 50 μL di tampone EIA 10x dopo un periodo di incubazione di 10 minuti.
NOTA: è vantaggioso includere un controllo positivo per garantire che la configurazione e i tessuti funzionino. La capsaicina 48,49,50 o lo stimolo depolarizzante del potassio (40-60 mM di KCl)46,47,49 sono abitualmente utilizzati per causare il rilascio di CGRP dal sistema trigeminovascolare. 40-60 mM di KCl SIF vengono preparati come SIF, tranne che NaCl viene scambiato con KCl su base equimolare.
- Quando appropriato, aggiungere un controllo positivo (ad esempio, 1-10 μM di capsaicina, vedere Tabella dei materiali) al tessuto alla fine del protocollo e raccogliere un campione di 200 μL in una provetta da microcentrifuga con 50 μL di tampone EIA 10x dopo un periodo di incubazione di 10 minuti.
6. Analisi delle concentrazioni di CGRP
- Misurare la quantità di CGRP rilasciata utilizzando un kit di dosaggio immunoenzimatico (EIA) seguendo il protocollo del produttore (vedere la tabella dei materiali).
- Misurare la densità ottica a 410 nm utilizzando un fotometro a piastre. Se si utilizza un kit CGRP EIA diverso, misurare la densità ottica alla lunghezza d'onda fornita nel protocollo del produttore.
NOTA: i campioni devono essere diluiti in modo che corrispondano alla curva standard. - Eseguire l'analisi dei dati.
- Presentare i dati come concentrazioni assolute o normalizzare il rilascio basale di CGRP dal tessuto specifico.
Risultati
Questa tecnica è uno strumento per studiare i meccanismi molecolari correlati al CGRP coinvolti nell'emicrania. Ha il vantaggio di valutare il rilascio di CGRP da diversi livelli del sistema trigeminovascolare e può essere applicato sia su topi e ratti wild-type che transgenici in combinazione con vari composti farmacologici. Qui vengono presentati esperimenti concentrazione-risposta e blocco da ratti e risultati concentrazione-risposta da topi wild-type e transgenici.
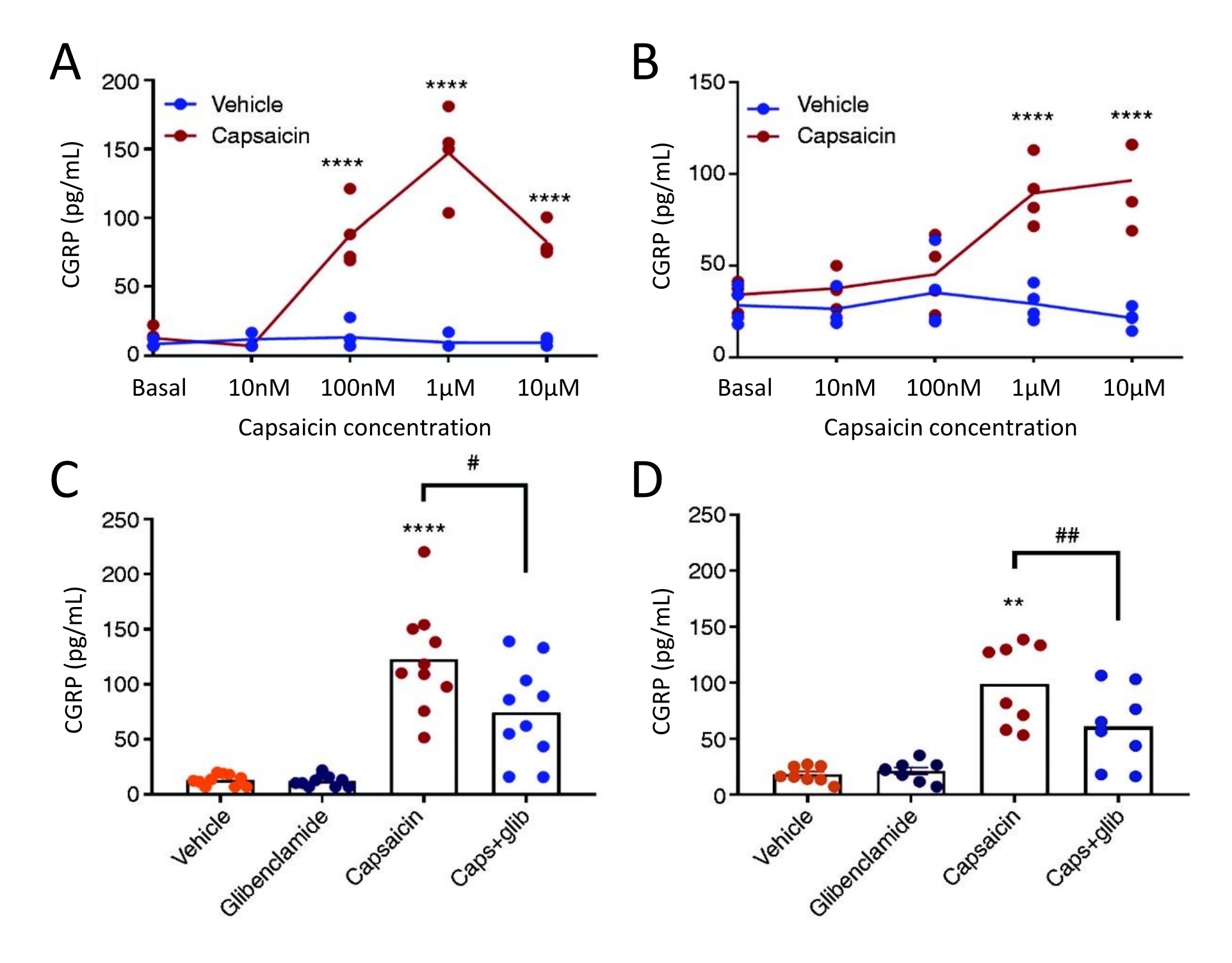
Il metodo di rilascio di CGRP è stato utilizzato per studiare l'effetto dell'inibitore del canale KATP glibenclamide sul rilascio di CGRP da TG e dura madre in ratti allodinici trigeminali spontanei (STA) femmine. In primo luogo, la concentrazione ottimale di capsaicina è stata trovata utilizzando un disegno di studio concentrazione-risposta. L'esposizione alla capsaicina ha indotto un significativo rilascio di CGRP dalla dura madre e dal TG rispetto al veicolo (Figura 5). Nella dura madre, il rilascio massimo di CGRP è stato trovato a 1 μM di capsaicina, e in TG, il rilascio massimo di CGRP è stato trovato a 10 μM di capsaicina (Figura 5A-B). Sulla base degli esperimenti concentrazione-risposta, sono stati utilizzati 1 μM di capsaicina e 3 μM di glibenclamide per esperimenti di blocco. La glibenclamide non ha mostrato alcun effetto sul rilascio basale di CGRP da dura madre (P = 0,441) e TG (P = 0,881) quando analizzata con ANOVA51 unidirezionale. La glibenclamide ha ridotto significativamente il rilascio di CGRP indotto da capsaicina nella dura madre del 40% (P = 0,031) e il TG del 39% (P = 0,003) rispetto alla capsaicina nel veicolo quando analizzato con un ANOVA unidirezionale (Figura 5C-D)51.
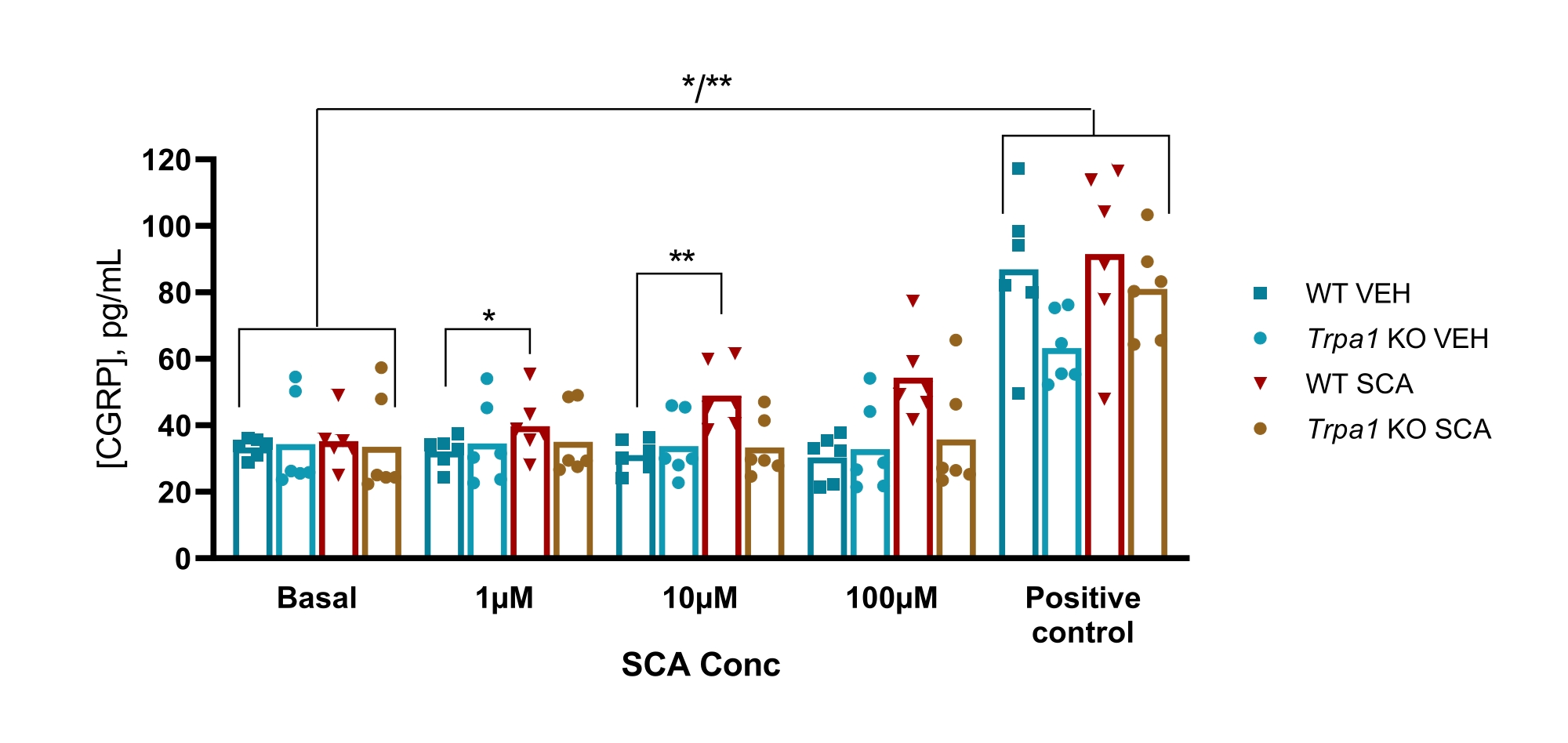
Nei topi, il protocollo è stato utilizzato per esaminare il coinvolgimento del canale ionico del potenziale recettore transitorio anchirina 1 (TRPA1) in un modello murino GTN di emicrania, in cui l'ipersensibilità indotta da GTN era completamente dipendente dai canali TRPA1. È stato riscontrato che l'agonista TRPA1 supercinnamaldeide (SCA) rilascia CGRP in modo dose-dipendente dal TG con 1 μM, 10 μM e 100 μM di SCA, risultando in 9% (P = 0,23), 51% (P = 0,011) e 69% (P = 0,0097) aumento del rilascio di CGRP rispetto al veicolo, rispettivamente quando analizzato con ANOVA a due vie. Questo rilascio era assente in TG da topi nulli Trpa1 in cui l'esposizione a 1 μM, 10 μM e 100 μM di SCA ha determinato una variazione percentuale dell'11% (P > 0,99), -13% (P > 0,99) e del 9% (P = 0,97) nel rilascio di CGRP rispetto al veicolo, rispettivamente quando analizzata con ANOVA bidirezionale. La successiva stimolazione con 10 μM di capsaicina (controllo positivo) mostra che tutti i campioni di tessuto potrebbero rilasciare CGRP (Figura 6)50.

Figura 1: Dissezione passo-passo del tessuto dai ratti. I dettagli (A-T) sono forniti nella sezione protocollo (passaggio 3.1). Fare clic qui per visualizzare una versione ingrandita di questa figura.
{kind=link}

Figura 2: Dissezione passo-passo del tessuto dai topi. I dettagli (A-T) sono forniti nella sezione protocollo (passaggio 3.2). Fare clic qui per visualizzare una versione ingrandita di questa figura.
{kind=link}

Figura 3: Lavaggio e incubazione del tessuto dei roditori. (A-B) Tessuto appena isolato in contenitori di plastica con SIF. I contenitori sono rivestiti in tulle per consentire un facile cambio di SIF. (C) Tessuto di ratto - Due metà del cranio con dura madre che coprono i rivestimenti interni del cranio poste su una piastra di coltura a 6 pozzetti. Il nucleo caudale del trigemino destro e sinistro in coperchi di tubi di microcentrifuga separati (fila superiore di coperchi). I due gangli del trigemino nei singoli coperchi dei tubi della microcentrifuga (fila inferiore di coperchi). (D) Tessuto di topo - La parte del tronco cerebrale contenente il nucleo caudale del trigemino in un coperchio separato del tubo di microcentrifuga (in alto). Due gangli del trigemino di topo sono in un coperchio del tubo della microcentrifuga (in basso). Fare clic qui per visualizzare una versione ingrandita di questa figura.
{kind=link}

Figura 4: Progetti di studio per il rilascio di CGRP. Tre diversi protocolli per l'esecuzione di esperimenti di rilascio CGRP. I farmaci devono essere diluiti nel liquido interstiziale sintetico (SIF). (A) Stimolazione singola. (B) Stimolazione concentrazione-risposta. (C) Inibizione di una stimolazione. Fare clic qui per visualizzare una versione ingrandita di questa figura.
{kind=link}

Figura 5: La glibenclamide inibisce il rilascio di CGRP indotto dalla capsaicina dalla dura madre e dal ganglio trigemino del ratto. I livelli di CGRP dal ganglio trigemino e dalla dura madre isolati da ratti allodinici trigeminali spontanei (STA) femmina (215-318 g) originariamente provenienti dalla Thomas Jefferson University52 sono stati misurati con kit CGRP EIA umani commerciali. (A-B) Rilascio di CGRP da (A) dura madre e (B) ganglio trigemino dopo aumento delle concentrazioni di capsaicina (10 nM, 100 nM, 1 μM e 10 μM) (n = 4). I dati sono presentati come singoli punti e sono stati analizzati con ANOVA bidirezionale. p < 0,0001 (C-D) Livelli di CGRP rilasciati da (C) dura madre e (D) ganglio trigemino dopo 10 minuti di esposizione al veicolo, 3 μM glibenclamide (glib), 1 μM capsaicina e 1 μM capsaicina + 3 μM glib (n = 6-11). I dati sono presentati come singoli punti e valori medi e sono stati analizzati con ANOVA unidirezionale. *rispetto al veicolo. #capsaicina rispetto a capsaicina + glib. #P < 0,05, ** e ##P < 0,01, ****P < 0,0001. Le analisi sono state seguite con il test di confronto multiplo di Bonferroni. Per tutti i test è stato utilizzato un livello significativo di α = 0,05. Questa cifra è stata modificata da Christensen et al. 51. Fare clic qui per visualizzare una versione ingrandita di questa figura.
{kind=link}

Figura 6: L'esposizione alla SCA provoca il rilascio di CGRP dipendente da TRPA1 dal ganglio trigemino del topo. I livelli di CGRP rilasciati dal ganglio trigemino isolato da WT e Trpa1 null (Trpa1tm1/Dpc)53 topi maschi (8-10 settimane) sono stati misurati con kit CGRP EIA di ratto dopo esposizione a supercinnamaldeide (SCA) a 1 μM, 10 μM e 100 μM e capsaicina a 10 μM come controllo positivo (n = 6). I dati di ciascun mouse sono presentati come singoli punti e le barre indicano i valori medi. Statistiche: Il confronto tra SCA e veicolo ad ogni concentrazione e tra controllo basale e positivo è stato eseguito con un ANOVA ripetuto a due vie. Un livello significativo di α = 0,05. *P < 0,05, **P < 0,01. Questa cifra è stata modificata da Christensen et al. 50. Fare clic qui per visualizzare una versione ingrandita di questa figura.
{kind=link}
Discussione
Il metodo descritto è stato sviluppato a seguito di studi che dimostrano l'importanza della CGRP nella fisiopatologia dell'emicrania. È adatto per studiare i meccanismi coinvolti nel rilascio di CGRP dal sistema trigeminovascolare, che è cruciale per la segnalazione del dolore nella regione della testa. La quantità di CGRP ottenuta in questo modello misura direttamente il rilascio di CGRP dai nervi trigemino che innervano la dura madre, TG e TNC. La quantità di rilascio di CGRP è quantitativamente maggiore 45,54 rispetto al rilascio misurato nel plasma dopo termocoagulazione nell'uomo, stimolazione del trigemino nel gatto 39,55,56 e durante gli attacchi di emicrania 23. Una spiegazione potrebbe essere che CGRP è diluito e degradato nel sangue54. Tuttavia, va notato che la stimolazione diretta con sostanze chimiche potrebbe essere superiore all'attivazione fisiopatologica. Ulteriori vantaggi sono che è possibile localizzare il rilascio da tre diversi siti all'interno del sistema trigeminovascolare e che può essere utilizzato insieme alla manipolazione farmacologica e nei tessuti di roditori geneticamente modificati.
Ultimamente, molti modelli preclinici di roditori si concentrano sulla somministrazione sistemica di sostanze e sulle successive letture correlate al dolore o all'emicrania utilizzando il test di von Frey57,58, una smorfia59,60,61 o un'avversione alla luce 62,63,64. Questi metodi sono utili per comprendere le proprietà antidolorifiche e antidolorifiche di diverse sostanze. Tuttavia, questi approcci non forniscono informazioni su specifici tessuti bersaglio coinvolti. Nel metodo attuale, il sistema trigeminovascolare è diviso in tre strutture: la dura madre, TG e TNC. Ciò consente l'esposizione locale di ciascuna struttura e la valutazione del luogo di azione di una sostanza specifica. Questo è stato utilizzato in uno studio del 2011, in cui è stato esplorato il ruolo dei canali del calcio voltaggio-dipendenti nei ratti e l'inibizione di questi canali è risultata diversa nelle tre strutture della via trigeminovascolare47. Quando si sezionano le strutture del sistema nervoso, l'assotomia è inevitabile. È stato dimostrato che l'assotomia altera la trascrizione di vari geni65. Questi cambiamenti trascrizionali sono troppo lenti per influenzare i risultati di questo metodo, ma i cambiamenti nella fosforilazione non possono essere esclusi rispetto alle situazioni in vivo 46. Sebbene i neuropeptidi come CGRP si formino nel soma cellulare, il rilascio e l'azione dei neuropeptidi sono solitamente alle terminazioni nervose centrali o periferiche. Pertanto, gli studi sui neuroni intatti, compresi i terminali, sono interessanti quando si studia il rilascio di neuropeptidi. Pertanto, sono stati stabiliti metodi di studio di colture di neuroni da gangli isolati per servire da modello dei terminali. Tuttavia, le colture cellulari neuronali sono soggette a diversi problemi poiché la dissociazione meccanica può distruggere i neuroni in una coltura46. Il periodo di tempo più lungo associato alla coltura delle cellule rende questo metodo sensibile ai cambiamenti trascrittomici dovuti all'assotomia e alle condizioni di coltura65. Inoltre, l'aggiunta di fattori di crescita e la coltura su rivestimenti superficiali hanno alterato le proprietà neuronali come trasmettitore e espressione recettoriale66,67,68,69. Questi problemi vengono evitati quando si studiano gangli intatti appena isolati invece di colture cellulari neuronali.
Una sfida con il metodo di rilascio ex vivo CGRP è la dissezione precisa del tessuto necessaria per risultati riproducibili. Una dissezione particolarmente accurata del TNC è impegnativa in quanto si tratta di una struttura all'interno del tronco cerebrale senza confini visibili. Inoltre, la dura madre è fragile e la rimozione del cervello deve essere eseguita con attenzione per garantire una struttura intatta. Questi ostacoli possono comportare una variazione delle dimensioni del tessuto e, quindi, variare i livelli basali e di CGRP indotti dalla stimolazione. Tuttavia, questa variazione può essere spiegata dalla normalizzazione al rilascio basale di CGRP. Va anche notato che quando si isola il TNC dai topi, l'intera parte inferiore del tronco cerebrale è isolata e non la parte più specifica contenente TNC come fatto nei ratti. In generale, può essere un vantaggio utilizzare il tessuto del ratto, in quanto ciò consente la misurazione del rilascio di CGRP dalla dura madre e una dissezione più precisa del TNC. Inoltre, la dimensione del tessuto consente anche l'uso di un ratto come controllo del veicolo, poiché un ratto provoca due metà del cranio, due TG e due TNC in cui un pezzo del tessuto viene utilizzato per la stimolazione della sostanza e l'altro per il veicolo. Quando si usano i topi, sono necessari due animali per un esperimento poiché entrambi i TG sono raggruppati in un campione e le TNC sono sezionate come un unico tronco cerebrale. Pertanto, due TG e un tronco cerebrale vengono utilizzati per la stimolazione delle sostanze e due TG e un tronco cerebrale di un altro topo vengono utilizzati per il controllo del veicolo. Ciò si traduce nell'utilizzo del doppio dei topi rispetto ai ratti per ottenere lo stesso numero di repliche. Per ridurre il numero di topi utilizzati, è stato suggerito un metodo per misurare il rilascio di CGRP dalle fette del tronco cerebrale49. È un vantaggio che il metodo sia stato modificato per consentire l'uso di topi. Ciò consente l'uso di molti ceppi di topi transgenici già disponibili, uno strumento utile per studiare, ad esempio, le vie di segnalazione. Un controllo positivo alla fine di un esperimento dovrebbe essere incluso per garantire che il tessuto utilizzato nell'esperimento possa rilasciare CGRP. Il controllo positivo potrebbe essere la capsaicina agonista TRPV1 o lo stimolo depolarizzante potassio (KCl), che sono stati trovati per rilasciare CGRP dal sistema trigeminovascolare sia nei topi che nei ratti 46,47,48,49,50. Inoltre, il metodo è stato anche adattato per misurare il rilascio di altri peptidi rilevanti come il peptide attivante l'adenilato ciclasi ipofisario (PACAP) - un altro peptide di grande interesse nell'ambito della ricerca sull'emicrania70.
Il metodo fornisce uno strumento utile per studiare il rilascio di CGRP da specifici tessuti bersaglio in ratti e topi. È un metodo relativamente veloce che evita i problemi associati alla coltura dei neuroni. Il protocollo del metodo può essere facilmente modificato per studiare la relazione concentrazione-risposta o l'inibizione di una risposta da parte di vari composti farmacologici. Il metodo di rilascio ex vivo di CGRP è uno dei numerosi metodi preclinici utili per studiare il ruolo del CGRP e altri meccanismi correlati al rilascio di CGRP nella fisiopatologia dell'emicrania.
Divulgazioni
Gli autori non hanno nulla da rivelare.
Riconoscimenti
Questo lavoro è stato finanziato dalla Candys Foundation.
Materiali
| Name | Company | Catalog Number | Comments |
| 6-well culture plate | NUNC | 140675 | |
| Calcium chloride dihydrate | Merck | 1.02382.1000 | For SIF buffer |
| Caps for plastic containers | ThermoFisher Scientific | 536617 | |
| Capsaicin | Merck | M2028 | |
| CGRP kits | AH Diagnostics | A05482.96 | |
| CO2 | Strandmøllen | 4.6 | For carbogen gassing of SIF |
| Delicate Bone Trimmer | Fine Science Tools | 16109-14 | |
| Glibenclamide | Tocris | 911 | |
| Glucose | Merck | G7021 | For SIF buffer |
| Guillotine for rats | Scandidact | NS-802 | |
| Magnesium sulfate heptahydrate | Merck | M5921 | For SIF buffer |
| Microcenrifuge tubes + lids/caps | VWR | 700-5239 | |
| Mini Hacksaw | BAHCO | 208 | |
| O2 | Strandmøllen | 4.5 | For carbogen gassing of SIF |
| Pentobarbital | Glostrup pharmacy | NA | Magistral formula |
| Plastic containers | ThermoFisher Scientific | 536455 | |
| Plate photometer - Infinite M200 | Tecan | NA | Infinite M200 is discontinued. A Infinite 200 PRO is available at Tecan. Software: SW Magellan v.6.3 |
| Potassium chloride | Merck | P9333 | For SIF buffer |
| Scissor | Allgaier Instruments | 307-156-170 | |
| Small scissor | Allgaier Instruments | 04-520-115 | |
| Sodium bicarbonate | Merck | S6014 | For SIF buffer |
| Sodium chloride | Merck | S9888 | For SIF buffer |
| Sodium dihydrogen phosphate monohydrate | Merck | 1.06346.1000 | For SIF buffer |
| Sodium gluconate | Merck | S2054 | For SIF buffer |
| Spatula | Bochem Lab Supply | 3018 | |
| Spring scissor | Fine Science Tools | 15024-10 | |
| Sucrose | Merck | 84097 | For SIF buffer |
| Supercinnamaldehyde | Merck | S3322 | |
| Tulle (fabrics) | NA | NA | Bought in the local fabrics store |
Riferimenti
- GBD 2016 Collaborators. Global, regional, and national incidence, prevalence, and years lived with disability for 328 diseases and injuries for 195 countries, 1990-2016: A systematic analysis for the Global Burden of Disease Study. The Lancet. 390 (10100), 1211-1259 (2017).
- Dodick, D. W. CGRP ligand and receptor monoclonal antibodies for migraine prevention: Evidence review and clinical implications. Cephalalgia: An International Journal of Headache. 39 (3), 445-458 (2019).
- Sacco, S., et al. European headache federation guideline on the use of monoclonal antibodies acting on the calcitonin gene related peptide or its receptor for migraine prevention. The Journal of Headache and Pain. 20 (1), 6 (2019).
- Moreno-Ajona, D., Pérez-Rodríguez, A., Goadsby, P. J. Gepants, calcitonin-gene-related peptide receptor antagonists: what could be their role in migraine treatment. Current Opinion in Neurology. 33 (3), 309-315 (2020).
- Khan, S., Olesen, A., Ashina, M. CGRP, a target for preventive therapy in migraine and cluster headache: Systematic review of clinical data. Cephalalgia: An International Journal of Headache. 39 (3), 374-389 (2019).
- Edvinsson, L., Haanes, K. A., Warfvinge, K., Krause, D. N. CGRP as the target of new migraine therapies - successful translation from bench to clinic. Nature Reviews Neurology. 14 (6), 338 (2018).
- Goadsby, P. J., et al. Pathophysiology of migraine: A disorder of sensory processing. Physiological Reviews. 97 (2), 553-622 (2017).
- Amara, S. G., Jonas, V., Rosenfeld, M. G., Ong, E. S., Evans, R. M. Alternative RNA processing in calcitonin gene expression generates mRNAs encoding different polypeptide products. Nature. 298 (5871), 240-244 (1982).
- Rosenfeld, M. G., et al. Production of a novel neuropeptide encoded by the calcitonin gene via tissue-specific RNA processing. Nature. 304 (5922), 129-135 (1983).
- Edvinsson, L., Goadsby, P. J. Discovery of CGRP in relation to migraine. Cephalalgia: An International Journal of Headache. 39 (3), 331-332 (2019).
- Brain, S. D., Williams, T. J., Tippins, J. R., Morris, H. R., MacIntyre, I. Calcitonin gene-related peptide is a potent vasodilator. Nature. 313 (5997), 54-56 (1985).
- Fisher, L. A., et al. Stimulation of noradrenergic sympathetic outflow by calcitonin gene-related peptide. Nature. 305 (5934), 534-536 (1983).
- Hanko, J., Hardebo, J. E., Kåhrström, J., Owman, C., Sundler, F. Calcitonin gene-related peptide is present in mammalian cerebrovascular nerve fibres and dilates pial and peripheral arteries. Neuroscience Letters. 57 (1), 91-95 (1985).
- Uddman, R., Edvinsson, L., Ekblad, E., Håkanson, R., Sundler, F. Calcitonin gene-related peptide (CGRP): perivascular distribution and vasodilatory effects. Regulatory Peptides. 15 (1), 1-23 (1986).
- Edvinsson, L. Functional role of perivascular peptides in the control of cerebral circulation. Trends in Neurosciences. 8, 126-131 (1985).
- Edvinsson, L., Ekman, R., Jansen, I., Ottosson, A., Uddman, R. Peptide-containing nerve fibers in human cerebral arteries: Immunocytochemistry, radioimmunoassay and in vitro pharmacology. Annals of Neurology. 21 (5), 431-437 (1987).
- Edvinsson, L., Fredholm, B. B., Hamel, E., Jansen, I., Verrecchia, C. Perivascular peptides relax cerebral arteries concomitant with stimulation of cyclic adenosine monophosphate accumulation or release of an endothelium-derived relaxing factor in the cat. Neuroscience Letters. 58 (2), 213-217 (1985).
- Brain, S. D., Grant, A. D. Vascular actions of calcitonin gene-related peptide and adrenomedullin. Physiological Reviews. 84 (3), 903-934 (2004).
- Skofitsch, G., Jacobowitz, D. M. Calcitonin gene-related peptide: Detailed immunohistochemical distribution in the central nervous system. Peptides. 6 (4), 721-745 (1985).
- Suzuki, N., Hardebo, J. E., Owman, C. Origins and pathways of cerebrovascular nerves storing substance P and calcitonin gene-related peptide in rat. Neuroscience. 31 (2), 427-438 (1989).
- Uddman, R., Edvinsson, L., Ekman, R., Kingman, T., McCulloch, J. Innervation of the feline cerebral vasculature by nerve fibers containing calcitonin gene-related peptide: trigeminal origin and co-existence with substance P. Neuroscience Letters. 62 (1), 131-136 (1985).
- Warfvinge, K., Edvinsson, L. Distribution of CGRP and CGRP receptor components in the rat brain. Cephalalgia: An International Journal of Headache. 39 (3), 342-353 (2019).
- Goadsby, P. J., Edvinsson, L., Ekman, R. Vasoactive peptide release in the extracerebral circulation of humans during migraine headache. Annals of Neurology. 28 (2), 183-187 (1990).
- Lassen, L. H., et al. CGRP may play a causative role in migraine. Cephalalgia: An International Journal of Headache. 22 (1), 54-61 (2002).
- Olesen, J., et al. Calcitonin gene-related peptide receptor antagonist BIBN 4096 BS for the acute treatment of migraine. New England Journal of Medicine. 350 (11), 1104-1110 (2004).
- Eftekhari, S., et al. Localization of CGRP, CGRP receptor, PACAP and glutamate in trigeminal ganglion. Relation to the blood-brain barrier. Brain Research. 1600, 93-109 (2015).
- Keller, J. T., Marfurt, C. F. Peptidergic and serotoninergic innervation of the rat dura mater. Journal of Comparative Neurology. 309 (4), 515-534 (1991).
- Edvinsson, L., et al. Innervation of the human middle meningeal artery: immunohistochemistry, ultrastructure, and role of endothelium for vasomotility. Peptides. 19 (7), 1213-1225 (1998).
- Lennerz, J. K., et al. Calcitonin receptor-like receptor (CLR), receptor activity-modifying protein 1 (RAMP1), and calcitonin gene-related peptide (CGRP) immunoreactivity in the rat trigeminovascular system: Differences between peripheral and central CGRP receptor distribution. Journal of Comparative Neurology. 507 (3), 1277-1299 (2008).
- Eftekhari, S., Edvinsson, L. Calcitonin gene-related peptide (CGRP) and its receptor components in human and rat spinal trigeminal nucleus and spinal cord at C1-level. BMC Neuroscience. 12, 112 (2011).
- Eftekhari, S., et al. Differential distribution of calcitonin gene-related peptide and its receptor components in the human trigeminal ganglion. Neuroscience. 169 (2), 683-696 (2010).
- Eftekhari, S., Warfvinge, K., Blixt, F. W., Edvinsson, L. Differentiation of nerve fibers storing CGRP and CGRP receptors in the peripheral trigeminovascular system. The Journal of Pain. 14 (11), 1289-1303 (2013).
- Spitzer, M. J. S., Reeh, P. W., Sauer, S. K. Mechanisms of potassium- and capsaicin-induced axonal calcitonin gene-related peptide release: Involvement of L- and T-type calcium channels and TRPV1 but not sodium channels. Neuroscience. 151 (3), 836-842 (2008).
- Evans, A. R., Nicol, G. D., Vasko, M. R. Differential regulation of evoked peptide release by voltage-sensitive calcium channels in rat sensory neurons. Brain Research. 712 (2), 265-273 (1996).
- Gupta, S., Villalón, C. M. The relevance of preclinical research models for the development of antimigraine drugs: Focus on 5-HT 1B/1D and CGRP receptors. Pharmocology & Therapeutics. 128 (1), 170-190 (2010).
- Williamson, D. J., Hargreaves, R. J., Hill, R. G., Shepheard, S. L. Intravital microscope studies on the effects of neurokinin agonists and calcitonin gene-related peptide on dural vessel diameter in the anaesthetized rat. Cephalalgia: An International Journal of Headache. 17 (4), 518-524 (1997).
- Gupta, S., Bhatt, D. K., Boni, L. J., Olesen, J. Improvement of the closed cranial window model in rats by intracarotid infusion of signalling molecules implicated in migraine. Cephalalgia: An International Journal of Headache. 30 (1), 27-36 (2010).
- Knight, Y. E., Edvinsson, L., Goadsby, P. J. Blockade of calcitonin gene-related peptide release after superior sagittal sinus stimulation in cat: a comparison of avitriptan and CP122, 288. Neuropeptides. 33 (1), 41-46 (1999).
- Goadsby, P. J., Edvinsson, L., Ekman, R. Release of vasoactive peptides in the extracerebral circulation of humans and the cat during activation of the trigeminovascular system. Annals of Neurology. 23 (2), 193-196 (1988).
- Tassorelli, C., Joseph, S. A. Systemic nitroglycerin induces Fos immunoreactivity in brainstem and forebrain structures of the rat. Brain Research. 682 (1-2), 167-181 (1995).
- Ramachandran, R., et al. A naturalistic glyceryl trinitrate infusion migraine model in the rat. Cephalalgia: An International Journal of Headache. 32 (1), 73-84 (2012).
- Hoskin, K. L., Zagami, A. S., Goadsby, P. J. Stimulation of the middle meningeal artery leads to Fos expression in the trigeminocervical nucleus: a comparative study of monkey and cat. Journal of Anatomy. 194, 579-588 (1999).
- Charbit, A. R., Akerman, S., Goadsby, P. J. Comparison of the Effects of Central and Peripheral Dopamine Receptor Activation on Evoked Firing in the Trigeminocervical Complex. Journal of Pharmacology and Experimental Therapeutics. 331 (2), 752-763 (2009).
- Koulchitsky, S., Fischer, M., Messlinger, K. Calcitonin gene-related peptide receptor inhibition reduces neuronal activity induced by prolonged increase in nitric oxide in the rat spinal trigeminal nucleus. Cephalalgia: An International Journal of Headache. 29 (4), 408-417 (2009).
- Ebersberger, A., Averbeck, B., Messlinger, K., Reeh, P. W. Release of substance P, calcitonin gene-related peptide and prostaglandin E2 from rat dura mater encephali following electrical and chemical stimulation in vitro. Neuroscience. 89 (3), 901-907 (1999).
- Eberhardt, M., et al. Calcitonin gene-related peptide release from intact isolated dorsal root and trigeminal ganglia. Neuropeptides. 42 (3), 311-317 (2008).
- Amrutkar, D. V., Ploug, K. B., Olesen, J., Jansen-Olesen, I. Role for voltage gated calcium channels in calcitonin gene-related peptide release in the rat trigeminovascular system. Neuroscience. 172, 510-517 (2011).
- Gupta, S., et al. Evidence for CGRP re-uptake in rat dura mater encephali. British Journal of Pharmacology. 161 (8), 1885-1898 (2010).
- Kageneck, C., Nixdorf-Bergweiler, B. E., Messlinger, K., Fischer, M. J. M. Release of CGRP from mouse brainstem slices indicates central inhibitory effect of triptans and kynurenate. Journal of Headache and Pain. 15 (1), 1-9 (2014).
- Christensen, S. L., et al. CGRP-dependent signalling pathways involved in mouse models of GTN- cilostazol- and levcromakalim-induced migraine. Cephalalgia: An International Journal of Headache. 41 (14), 1413-1426 (2021).
- Christensen, S. L., et al. ATP sensitive potassium (KATP) channel inhibition: A promising new drug target for migraine. Cephalalgia: An International Journal of Headache. 40 (7), 650-664 (2020).
- Oshinsky, M. L., et al. Spontaneous trigeminal allodynia in rats: A model of primary headache. Headache. 52 (9), 1336 (2012).
- Kwan, K. Y., et al. TRPA1 contributes to cold, mechanical, and chemical nociception but is not essential for hair-cell transduction. Neuron. 50 (2), 277-289 (2006).
- Eltorp, C., Jansen-Olesen, I., Hansen, A. J. Release of calcitonin gene-related peptide (CGRP) from guinea pig dura mater in vitro is inhibited by sumatriptan but unaffected by nitric oxide. Cephalalgia: An International Journal of Headache. 20 (9), 838-844 (2000).
- Zagami, A. S., Goadsby, P. J., Edvinsson, L. Stimulation of the superior sagittal sinus in the cat causes release of vasoactive peptides. Neuropeptides. 16 (2), 69-75 (1990).
- Goadsby, P. J., Edvinsson, L. The trigeminovascular system and migraine: studies characterizing cerebrovascular and neuropeptide changes seen in humans and cats. Annals of Neurology. 33 (1), 48-56 (1993).
- Bates, E. A., et al. Sumatriptan alleviates nitroglycerin-induced mechanical and thermal allodynia in mice. Cephalalgia: An International Journal of Headache. 30 (2), 170-178 (2010).
- Pradhan, A. A., et al. Characterization of a novel model of chronic migraine. Pain. 155 (2), 269-274 (2014).
- Mogil, J. S., Pang, D. S. J., Silva Dutra, G. G., Chambers, C. T. The development and use of facial grimace scales for pain measurement in animals. Neuroscience and Biobehavioral Reviews. 116, 480-493 (2020).
- Sotocinal, S. G., et al. The rat grimace scale: A partially automated method for quantifying pain in the laboratory rat via facial expressions. Molecular Pain. 7, 55 (2011).
- Langford, D. J., et al. Coding of facial expressions of pain in the laboratory mouse. Nature Methods. 7 (6), 447-449 (2010).
- Mahmoudi, J., et al. Cerebrolysin attenuates hyperalgesia, photophobia, and neuroinflammation in a nitroglycerin-induced migraine model in rats. Brain Research Bulletin. 140, 197-204 (2018).
- Farajdokht, F., Babri, S., Karimi, P., Mohaddes, G. Ghrelin attenuates hyperalgesia and light aversion-induced by nitroglycerin in male rats. Neuroscience Letters. 630, 30-37 (2016).
- Kuburas, A., et al. PACAP Induces Light Aversion in Mice by an Inheritable Mechanism Independent of CGRP. Journal of Neuroscience. 41 (21), 4697-4715 (2021).
- Buschmann, T., et al. Expression of Jun, Fos, and ATF-2 proteins in axotomized explanted and cultured adult rat dorsal root ganglia. Neuroscience. 84 (1), 163-176 (1998).
- Lee, Y. J., Zachrisson, O., Tonge, D. A., McNaughton, P. A. Upregulation of bradykinin B2 receptor expression by neurotrophic factors and nerve injury in mouse sensory neurons. Molecular and Cellular Neuroscience. 19 (2), 186-200 (2002).
- Hari, A., Djohar, B., Skutella, T., Montazeri, S. Neurotrophins and extracellular matrix molecules modulate sensory axon outgrowth. International Journal of Developmental Neuroscience. 22 (2), 113-117 (2004).
- Skoff, A. M., Resta, C., Swamydas, M., Adler, J. E. Nerve Growth Factor (NGF) and Glial Cell Line-Derived Neurotrophic Factor (GDNF) regulate substance P release in adult spinal sensory neurons. Neurochemical Research. 28 (6), 847-854 (2003).
- Lindsay, R. M., Harmar, A. J. Nerve growth factor regulates expression of neuropeptide genes in adult sensory neurons. Nature. 337 (6205), 362-364 (1989).
- Edvinsson, J. C. A., et al. Differences in pituitary adenylate cyclase-activating peptide and calcitonin gene-related peptide release in the trigeminovascular system. Cephalalgia: An International Journal of Headache. 40 (12), 1296-1309 (2020).
Ristampe e Autorizzazioni
Richiedi autorizzazione per utilizzare il testo o le figure di questo articolo JoVE
Richiedi AutorizzazioneEsplora altri articoli
This article has been published
Video Coming Soon
Personale delle biblioteche
Copyright © 2025 MyJoVE Corporation. Tutti i diritti riservati