Method Article
Generazione di Dispersi Presomitic mesoderma colture cellulari, per Immagini del Zebrafish segmentazione Orologio in celle singole
In questo articolo
Riepilogo
Somitogenesis è un processo di sviluppo ritmico che spazialmente modelli l'asse del corpo di embrioni dei vertebrati. In precedenza, abbiamo sviluppato linee di zebrafish transgenico che usano i giornalisti fluorescenti ad osservare i geni ciclici che guidano questo processo. Qui, la cultura dispersi cellule da queste linee e l'immagine dei loro oscillazioni nel corso del tempo in vitro.
Abstract
La segmentazione è un processo morfogenetico periodica e sequenziale nei vertebrati. Questa formazione ritmica di blocchi di tessuto chiamato somiti lungo l'asse del corpo è la prova di un oscillatore genetica patterning embrioni. In zebrafish, il intracellulare orologio segmentazione di guida è composto da membri del suo / Hes fattore di trascrizione della famiglia organizzata in cicli di feedback negativo. Abbiamo recentemente generata transgenici linee reporter fluorescente per la HER1 gene ciclico che ricapitolare il modello spazio-temporale delle oscillazioni nel mesoderma presomitic (PSM). Utilizzando queste linee, abbiamo sviluppato un sistema di coltura in vitro che permette l'analisi in tempo reale di segmentazione oscillazioni orologio entro singolo, isolato cellule PSM. Rimuovendo il tessuto PSM da embrioni transgenici e quindi disperdere le cellule da oscillante regioni su piatti con fondo di vetro, abbiamo generato culture adatti per time-lapse imaging del segnale di fluorescenza dacelle di clock individuali. Questo approccio fornisce un quadro sperimentale e concettuale per la manipolazione diretta del clock di segmentazione con risoluzione unicellulare senza precedenti, consentendo sue proprietà cellule autonome ea livello di tessuto distinguere e sezionati.
Introduzione
La formazione periodica dei segmenti lungo l'asse del corpo vertebrati, o somitogenesis, è la prova di un oscillatore spaziale e temporale negli embrioni. Il meccanismo favorito controllo somitogenesis è concettualmente descritto da un modello "orologio e fronte d'onda" 1, in cui il "clock" costituito da oscillatori cellulari, ora pensato per essere guidato intracellulare dall'espressione ritmica di una serie di geni ciclici 2, scandisce la formazione dei somiti dal mesoderma presomitic (PSM). Come l'embrione si sviluppa, una maturazione "fronte d'onda" nel PSM muove di concerto con il tessuto regressione verso la parte posteriore, rallentando e arrestando oscillatori cellulari che passa 3. Insieme, questo sistema spatiotemporally dinamico è definito l'orologio segmentazione. Gli attuali approcci per studiare l'arco dell'orologio segmentazione tre livelli crescenti di organizzazione del oscillatore genetica in cellule singole a th accoppiamento localia verifica tra le cellule e, infine, alla regolamentazione globale delle informazioni di posizione nel collettivo tessuto PSM 4.
Precedenti studi suggeriscono la segmentazione oscillatore cellula-autonoma in zebrafish consiste di geni e prodotti proteici dalla trascrizione suoi / hes factorfamily, che si pensa a formare un anello di retroazione negativa attraverso la repressione trascrizionale 5-7. La via di segnalazione Delta / Notch sincronizza le oscillazioni tra cellule vicine e regola il periodo collettiva della popolazione 8-10. FGF segnalazione molecole prodotte in tailbud sembrano costruire un gradiente di tutti i zebrafish PSM, e sono quindi ipotizzato di contribuire a rallentare e arrestare le cellule oscillanti nel anteriore 11. Fino ad ora, i ruoli funzionali di ciascuna di tali molecole in somitogenesis sono stati studiati da mutazione genetica, iniezione morfolino, heat-shock sovraespressione e antagonista farmaco Treatimento di componenti e di segnalazione tra le cellule 5,7,10,12 di clock. Utilizzando queste perturbazioni, funzione orologio segmentazione è stato dedotto dalle descrizioni a livello di tessuto di difetti somite e la perdita di oscillazioni uniformi nell'espressione dei geni ciclici come HER1, her7, e delta C. Tuttavia, quasi tutti questi dati sono da embrioni fisse e non riescono a cogliere con precisione i cambiamenti che sono inerenti alla funzione dinamica del clock segmentazione. Più di recente, multiple-embrione time-lapse imaging ha rivelato i primi mutanti con periodo oscillatore alterato, ma queste osservazioni sono state anche fatta al 7,13 livello tissutale. Pertanto, non è stato osservato il comportamento dell'oscillatore cellula-autonoma ipotizzato durante somitogenesis.
Istantanee statiche di somitogenesis danno un quadro incompleto perché, intrinsecamente, il processo è guidato da un sistema oscillatorio. In precedenza il lavoro in cellule di topo e pulcino ha mostrato che i livelli di trascrizionee l'aumento delle proteine e la caduta, ma il campionamento un'oscillazione di circa 2 ore ogni 30 o 45 min limita necessariamente i dati raccolti e quindi le conclusioni che si possono trarre 14,15. Studio di altri oscillatori biologici, in particolare, orologi circadiani, si è allontanato dalle misure messe in scena di espressione genica e proteica di monitoraggio in tempo reale utilizzando fluorescenti e reporter bioluminescenti 16,17. Questi strumenti sono essenziali per dimostrare le proprietà di clock di singole cellule 18. Un reporter bioluminescenti del gene ciclico Hes1 è stato sviluppato e brevemente caratterizzato singole cellule topo PSM 19. Il periodo e la varianza media sono stati calcolati per un piccolo numero di cellule, mostrando che le oscillazioni persistono per più cicli in vitro. Tuttavia, questi studi non hanno quantitativamente affrontare la stabilità e robustezza della frequenza dell'oscillatore e ampiezza, se le cellule possono entrare spontaneamente o oscillazioni di uscita,e come le cellule mantengono le loro relazioni di fase. Inoltre, gli effetti di molecole di segnalazione presenti nell'embrione sull'orologio cellula-autonoma non sono stati testati direttamente. Di conseguenza, queste proprietà fondamentali dell'oscillatore singola cellula rimangono completamente sconosciuta.
Abbiamo recentemente sviluppato linee di pesci transgenici con BAC recombineering 20 a guidare Venere (YFP) reporter di fluorescenza di espressione HER1 30. Tali linee sfruttano gli spunti normativi del locus cromosomico integro in cui sono incorporati e ricapitolano le dinamiche temporali e modello spaziale di HER1. Questa innovazione consente il monitoraggio in tempo reale di espressione genica nell'embrione zebrafish sviluppo in vivo. Per studiare le proprietà fondamentali delle oscillazioni cellule autonome e come tale espressione è regolata nel corso del tempo, abbiamo recentemente sviluppato un metodo affidabile per isolare e registrare da cellule PSM in vitro. Questo protocol descrive come abbiamo utilizzato le nostre linee giornalista transgeniche per produrre colture di cellule disperse, da cui possiamo caratterizzare le oscillazioni del clock segmentazione zebrafish in celle singole. Possiamo quindi affrontare domande in sospeso nel campo non accessibili con analisi statica o livello tissutale, nonché manipolare direttamente l'orologio segmentazione a livello di singola cellula con molecole di segnalazione e inibitori.
Protocollo
1. Prima di dissezione
- Il giorno prima della dissezione, di ottenere embrioni da un incross di coppie di zebrafish eterozigoti per l'allele transgenico.
- Sollevare embrioni in E3 mezzo senza il blu di metilene a 28 ° C fino stadio scudo (6 ore dopo la fecondazione).
- Trasferimento degli embrioni in E3 mezzo senza il blu di metilene a 20 o CO / N. A 20 ° C, gli embrioni si formeranno 1 somite per ora una volta raggiunta fase tailbud. Dopo 17-20 hr O / N a 20 o C, embrioni dovrebbero essere 5 a 8 somite scena la mattina seguente all'inizio del protocollo dissezione.
- Montare strumenti e reagenti necessari per la dissezione.
- Pipetta di vetro del fuoco lucido per il trasferimento di embrioni e tessuti
- Coppia di pinza sottile per la rimozione di chorions da embrioni
- Sylgard rivestite piatto da 35 mm per dissezione - Versare Sylgard polimero in un piatto a 35 mm e curare O / N a 37 ° C. Effettuare la dissezione bene usandouna punta dell'ago per rimuovere una piccola quantità di polimero polimerizzato. Il piatto può essere pulito e successivamente riutilizzato.
- Appuntite, appiattite strumenti filo di tungsteno per la manipolazione dei tessuti PSM
- Micro-bisturi per tagliare pezzi desiderati di PSM alla cultura
- 35-mm piatto di plastica per tripsina incubazione
- L15 terreno contenente siero bovino fetale al 10%
- 0,25% di soluzione di tripsina / EDTA
- Sigmacoted punte gel-carico per la dispersione
- Capsule di Petri in plastica contenenti E3 mezzo senza il blu di metilene.
- Coprire piatto di imaging con fondo di vetro con Fibronectin1 substrato (10 mg / ml in PBS). Lascia piatto sul banco a cappotto durante la dissezione.
- Utilizzare stereoscopio con opportuni filtri di fluorescenza per identificare ed embrioni transgenici liste positive (fase 5-8 somite).
- Identificare gli embrioni transgenici esaminando il mesoderma presomitic (PSM) per l'espressione YFP nell'ambito del canale di fluorescenza (Figura 1A). Fluorescenza shoULD essere visibile nella regione dall'ultimo somite formata per il tailbud. Selezionare embrioni con brillanti del segnale; Il 25% della prole dovrebbe essere embrioni omozigoti che trasportano 2 copie del transgene. Il numero di embrioni necessari individuati varia in base dell'esperimento. Un pezzo tailbud sezionato produrrà cellule 1.000, in media. Tipicamente, alcuni embrioni supplementari positivi sono utili nel caso si commettono errori durante la dissezione.
- Utilizzare luce trasmessa come riferimento posizionale di distinguere un autofluorescenza, soprattutto nella cella tuorlo, dal segnale.
- Trasferire gli embrioni positive per un piatto di plastica Petri separato contenente E3 mezzo senza il blu di metilene.
2. PSM Dissection e Dispersione
- Preparare gli embrioni per la dissezione.
- Sotto un microscopio dissezione, utilizzando una pinza sottile, rimuovere con cautela il corion da ogni embrione nel mezzo E3. Fare attenzione a non danneggiare l'embrione o interrompere la cella tuorlo.
- Riempire Sylgard rivestite dissezione piatto con L15 media con siero. Utilizzando pinze o altro strumento piatta, eliminare eventuali bolle d'aria dalla superficie del Sylgard.
- Trasferire gli embrioni dechorionated utilizzando il vetro pipetta incendio lucidato al piatto dissezione.
- Utilizzando gli strumenti di filo, spostare tutti gli embrioni ad un lato del piatto dissezione.
- Sezionare PSM da un singolo embrione.
- Orient un singolo embrione su un fianco laterale nel piccolo ben fatta nello strato Sylgard nel piatto dissezione (Figura 1B).
- Utilizzando il micro-bisturi, fetta attraverso l'embrione e cellule tuorlo solo anteriore al hindbrain e attraverso il polo ventrale dell'embrione (Figura 1B, linea tratteggiata rossa).
- Rimuovere il pezzo anteriore dell'embrione dal pozzo, allontanandola al lato del piatto, e quindi utilizzare gli strumenti di filo per raschiare rimanenti granuli delle tuorlo dalla sezione posteriore compresa la PSM.
- Una volta che il tuorlo è stata raschiata via, appiattire e orientare il PSM con l'estremità anteriore che punta lontano dalla sperimentatore e la parte posteriore rivolta verso la sperimentatore (Figura 1C).
- Se il sottile strato di ectoderma non ha tirato fuori da questo punto, utilizzare gli strumenti di filo per sbucciare lontano dalla parte superiore del tessuto PSM.
- Utilizzando il micro-bisturi tagliare il tailbud, la punta più posteriore del PSM oltre la fine della notocorda, lontano dal resto del tessuto (Figura 1C, linea tratteggiata rossa). NOTA: Altri pezzi di tessuto dal PSM, per esempio anterior PSM più vicino al ultimo somite formata, possono anche essere presi in coltura, a seconda della domanda sperimentale.
- Spostare il pezzo tailbud ad un angolo del piatto lontano dalla zona dissezione. Utilizzare gli strumenti di filo per eliminare eventuali residui e del tessuto embrionale indesiderato dal campo dissezione.
- Ripetere con la prossima embrione.
- Pezzi Pool tailbud da aerei embryos, a seconda di quanti cellule saranno necessari per l'esperimento. In media, un unico pezzo di tessuto tailbud produce cellule 1.000.
- Riempire vuoto piatto di plastica da 35 mm con un piccolo volume di tripsina-EDTA.
- Utilizzando la pipetta di vetro fuoco lucido, trasferire brani tailbud da dissezione piatto in piatto contenente tripsina / EDTA. Incubare pezzi tailbud in tripsina / EDTA per 20 minuti a temperatura ambiente.
- Mentre il tessuto viene incubata in tripsina, rimuovere la soluzione Fibronectin1 dal piatto di imaging fondo trasparente.
- Lavare la soluzione di vetro 3 volte con acqua MilliQ. Utilizzare aspirazione per rimuovere ogni lavaggio e garantire il piatto è completamente asciutto.
- Disperdere pezzi tailbud in mezzo per l'imaging.
- Aggiungere 100 ml di L15 mezzo con siero nel piatto imaging.
- Utilizzando una punta gel rivestito, rimuovere i pezzi tailbud dalla tripsina / EDTA in piccolo volume possibile.
- Pipettare i pezzi tailbud nel terreno nellapiatto di imaging. Pipettare i pezzi su e giù più volte per rompere loro parte e sospendere le cellule nel mezzo. Fare attenzione a non introdurre bolle d'aria. Verificare la presenza di grumi di cellule sotto il microscopio e disperdere più come necessario.
- Consentono alle cellule disperse in sospensione di depositarsi sul vetro Fibronectin1 rivestite per 20 minuti a RT.
- Aggiungere una piccola quantità di ulteriore mezzo L15 con siero alle cellule prima di iniziare imaging. Fare attenzione a non distruggere le cellule stanziali.
- Data la domanda sperimentale, aggiungere qualsiasi supplementare o trattamenti farmacologici per la cultura.
3. Imaging di cellule disperso PSM
- Istituito piatto di imaging in camera a temperatura sul microscopio time-lapse imaging. Impostare Warner camera alla temperatura desiderata per l'esperimento. Lasciare piatto per stabilizzare almeno 30 min prima di iniziare l'acquisizione delle immagini. Temperatura crosscheck con sonda di temperatura esterna posta nel piatto, se necessario. NOTA: a causail rapporto tra temperatura e tasso somitogenesis 21 Controllo temperatura stabile è essenziale per misure accurate di periodo in singole cellule PSM.
- Acquisire immagini test del canale di fluorescenza per verificare che il tempo di esposizione e guadagno forniscono una vasta gamma dinamica di intensità senza saturazione per assicurare un buon segnale a livelli di rumore. Una tipica immagine di fluorescenza acquisita da cellule disperse generate dalle nostre linee di usare questo protocollo richiede 400 msec e 40 msec per un'immagine luce trasmessa con una macchina fotografica EMCCD opera a un guadagno EM di 85 anni. Inoltre, l'aumento di pre-amp e la velocità di lettura dal fotocamera sono anche essenziali per massimizzare il segnale su rumore.
- Utilizzando il canale di luce trasmessa, scegliere i campi di cellule per l'acquisizione time-lapse.
- Eseguire protocollo di acquisizione time-lapse per la lunghezza di tempo desiderato.
- Acquisire una sola immagine di fluorescenza per campo luce trasmessa e. NOTA: Impostare un intervallo tra ro di acquisizioneONDI che catturerà dinamiche temporali senza fotometabolismo over di imaging avanzata o tossicità indurre nelle cellule. Questo protocollo utilizza un intervallo di 2 min.
- Controllare time-lapse set-up di tanto in tanto durante la registrazione per garantire che le cellule rimangano a fuoco, nessun software o hardware errori, ecc
4. Elaborazione delle immagini acquisite di Time-lapse Movies
- Apri file film per un campo acquisita in un software di elaborazione delle immagini. NOTA: Fiji è stato utilizzato per tutte le elaborazioni in questo protocollo.
- Split luce trasmessa e fluorescenza fotogrammi da un campo per creare 2 pile di immagini.
- Traccia una singola cella nel canale luce trasmessa.
- Posizionare un ROI circolare (regione di interesse) intorno alla cella selezionata nel primo fotogramma nel canale luce trasmessa. NOTE:.. Solo le cellule che 1 sono sane alla fine della registrazione, 2 non si muovono al di fuori del campo, e 3 non entrano in contatto con altri ce.LLS sono monitorati.
- Salvare un ROI ogni pochi fotogrammi al gestore ROI. Traccia cella fino all'ultimo fotogramma.
- Misurare l'intensità utilizzando i ROI salvate sul canale di fluorescenza.
- Selezionare la pila fluorescenza e con il ROI salvato, utilizzare il cerchio personalizzato interpolatore plug-in e macro per misurare l'intensità della cella monitorati nel tempo.
- Controllare la traccia uscita dalla macro. Se la traccia cattura qualitativamente le caratteristiche di fluorescenza time-lapse, esportare i valori da un foglio di lavoro di Excel.
- Salvare la lista ROI per la cella.
- Ripetere inseguimento in canale trasmesso e misurare in canale di fluorescenza di altre cellule del settore.
- Ripetere tutti i passaggi per ulteriori campi dall'esperimento.
Risultati
Questo protocollo produce culture, dispersi, singole cellule PSM vitali per time-lapse imaging del segnale di fluorescenza (Figura 2). Il nostro transgene genera un reporter la cui ciclo di produzione e degradazione avviene con dinamiche simili al gene endogeno e proteine nell'embrione, dell'ordine di mezz'ora. Grazie alla sua fatturato veloce, il segnale YFP in singole cellule dovrebbe essere rilevata rapidamente per minimizzare sbianca e con un'alta risoluzione temporale per acquisire le caratteristiche di ogni ciclo oscillatorio. Inoltre, data la relativa oscurità del segnale, condizioni di coltura e di acquisizione sono stati messi a punto per garantire risultati sensibili e robusti. Abbiamo trovato i seguenti fattori per essere importanti nella generazione di colture cellulari PSM ottimali per l'imaging:. 1 un substrato ECM utilizzato per rivestire i piatti della cultura fondo di vetro. 2. Aggiunta di siero per la dissezione e mezzo di imaging. 3. Dissezione del PSM da embrioni identificati adottate dopo l'accoppiamento hetecoppie rozygous, la cui prole potenzialmente portare 2 copie del transgene. 4. Acquisizione dell'immagine entro un intervallo ottimale di ingrandimento, con obiettivo NA superiore per garantire la cattura efficace del segnale di fluorescenza. 5. Illuminazione con una sorgente di luce a stato solido per ridurre al minimo le fluttuazioni di intensità che potrebbero contribuire al rumore di fondo. 6. Rilevamento del segnale con una telecamera EM-CCD altamente sensibile per massimizzare il segnale di lettura.
Culture sub-ottimali conterranno cellule che non rimangono arrotondate e sano per tutta la registrazione. Abbiamo ipotizzato che il substrato ECM, un frammento di fibronectin1 zebrafish mantiene le cellule in uno stato PSM simile indifferenziata, in confronto ad altri substrati comunemente usati come poli-lisina o laminina. Altri substrati abbiamo testato causati cellule per appiattire sul vetro e la perdita del segnale fluorescente oscillante nel corso della registrazione. Abbiamo anche trovato che l'aggiunta di siero al mezzo durante la dissezione, la dispersione,e la registrazione è stata importante non solo per placare la tripsina utilizzato per la dissociazione, ma anche aumentato l'intensità di fluorescenza su sfondo, probabilmente a causa di una maggiore vitalità cellulare. Per garantire la cattura ottimale del segnale da singole cellule abbiamo usato un obiettivo 40x specificamente progettato per l'imaging di fluorescenza (Zeiss Plan serie NeoFluor) con un elevato NA. Abbiamo anche trovato che una sorgente di luce a stato solido disponibile un'illuminazione più stabile rispetto alle lampade al mercurio tradizionali, che è fondamentale per ridurre al minimo le fluttuazioni di fondo che contribuiscono alle immagini rumorose. Queste modifiche sono importanti per garantire risultati affidabili.
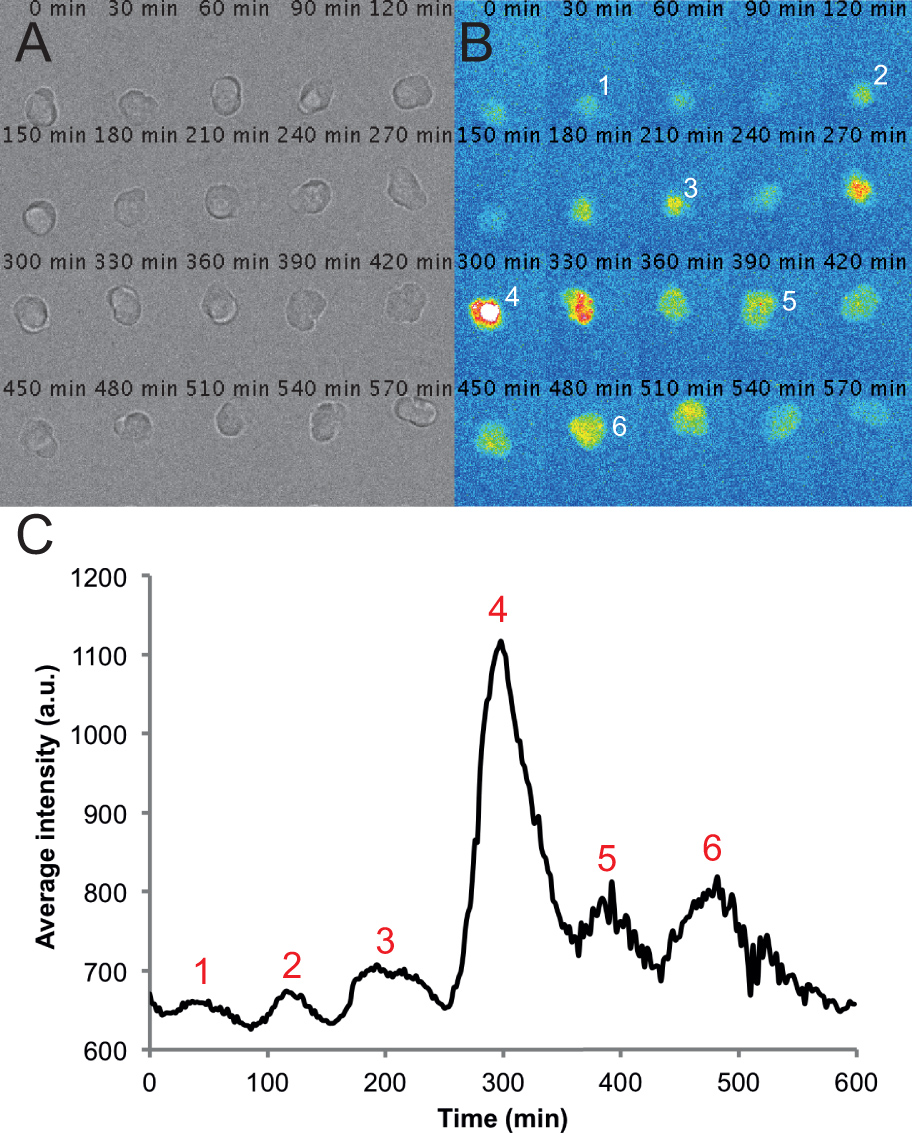
Usando questo protocollo prevediamo culture in cui la maggior parte delle cellule in un dato campo sono fluorescenti ad un certo punto durante la registrazione. Troviamo che le cellule fluorescenti in genere rimangono arrotondate e si sono volte piuttosto mobili durante le registrazioni. Alcune cellule, comprese le cellule fluorescenti, possono diventare apoptotica durante la registrazione. Queste cellule sono esclusi da qualsiasi unaANALISI. Abbiamo anche escludere le cellule che entrano in contatto con altre cellule o spostare al di fuori del campo di vista. In media, si vede una riduzione del 12% del numero di cellule per campo dalla fine della registrazione 10 hr misurata contando il numero di cellule sane nel canale luce trasmessa. Tale perdita include sia la morte cellulare e le cellule che hanno spostato fuori dal campo. Alla luce di questi avvertimenti che possiamo, in media, pista 5 cellule fluorescenti per campo che rimangono vitale e visibile, e in genere registrare 6 campi per condizione. Ad esempio, un esperimento con 4 condizioni avrà 24 campi in totale acquisiti e in genere di circa 30 cellule monitorati per condizione. In condizioni di imaging standard con L15 terreno contenente siero fetale bovino al 10%, troviamo che le cellule PSM (n = 101 celle da 4 repliche sperimentali) in grado di produrre tra i 2 ei 7 vette, con la media e la deviazione standard di 3 ± 1 picchi (2 cicli -3). Il numero di picco mediano, così come il percentile 25%, è 2 picchi e il 75% percentile is 3 cime. Utilizzando il nostro monitoraggio e l'analisi semi-automatico, siamo in grado di generare rapidamente intensità di fluorescenza su tracce di tempo per le singole celle PSM, con una traccia di cellule rappresentante mostrato in Figura 2C. Queste tracce prime possono poi essere utilizzati per effettuare misurazioni quantitative di proprietà delle cellule PSM oscillanti, come frequenza, ampiezza, numero di cicli, e la tempistica dei picchi.

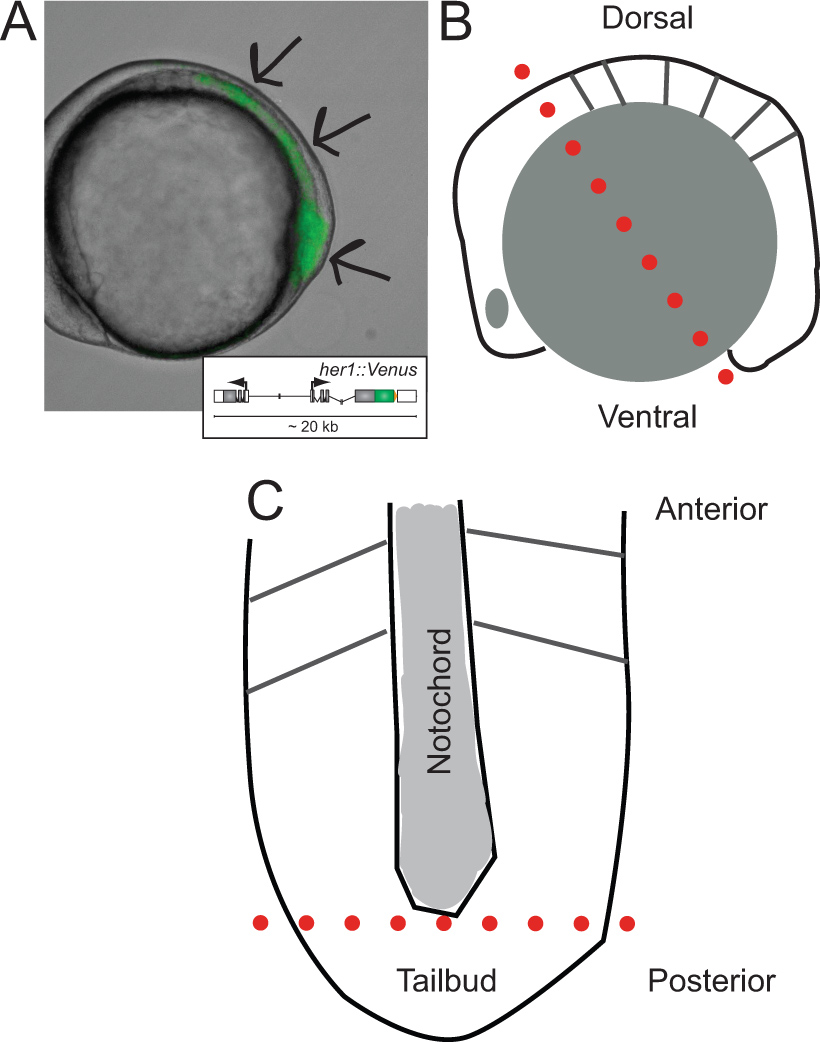
Figura 1. Identificazione di embrioni transgenici e gli schemi della loro dissezione. (A) Vista laterale di un embrione di zebrafish transgenico che esprime YFP fluorescenza a ~ fase 5-somite. L'immagine è una sovrapposizione di luce trasmessa e il canale di fluorescenza. Le frecce indicano le aree di segnale Partendo dalla punta del tailbud, in tutto il PSM, verso l ast somite formato. Inserto mostra uno schema del transgene reporter (per maggiori informazioni sul suo sviluppo e il comportamento in vivo riferimento a Soroldoni et al. 30. (B) Schema di vista laterale dell'embrione prima di tagliare per primo durante la dissezione. Linea rossa tratteggiata indica il primo taglio realizzato attraverso il hindbrain, anche se la cella tuorlo appena dietro il tailbud. (C) Rappresentazione schematica di appiattita PSM dopo la rimozione di granuli di tuorlo e lo strato epidermico, orientato lungo l'asse anteriore-posteriore. linea rossa tratteggiata indica il secondo taglio per rimuovere la punta del tailbud per la dispersione e la cultura. cliccate qui per vedere una versione più grande di questa figura.
{kind=link}
07fig2.jpg "/>
Figura 2. Immagini rappresentative e le tracce di una singola cella in una cultura PSM disperso. (A) Montaggio di trasmettere immagini chiare di una singola cella PSM in un 6-ore di registrazione time-lapse. Si noti che la cella rimane arrotondate e vitalità nel corso della registrazione. (B) corrispondente montaggio di immagini di fluorescenza da una singola cellula PSM. Picchi di intensità della cella sono numerati nel corso della registrazione. (C) intensità media nel tempo misurato da un ROI posto su questa cella. I valori sono presi da ogni fotogramma da un filmato con un frame rate di uno per 2 min utilizzando il plug-in cerchio interpolatore e macro personalizzato scritto in Fiji. Picchi di intensità sono di nuovo numerate in tutta la traccia. Cliccate qui per vedere una versione più grande di questa figura.
{kind=link}
Discussione
Per studiare un processo cellulare che avviene nel corso dello sviluppo embrionale, biologi utilizzano tipicamente un approccio nel contesto dell'intero embrione. Tuttavia, per comprendere appieno come una singola cellula si comporta in un lasso di tempo di sviluppo, un metodo per esaminare e perturbare le singole celle di isolamento è anche estremamente utile. Generando colture cellulari PSM dispersi utilizzando embrioni transgenici zebrafish ora abbiamo uno strumento per studiare direttamente la natura autonoma delle cellule di oscillazioni genetiche orologio segmentazione in modo quantitativo. E 'possibile misurare dinamica delle oscillazioni nel nostro reporter fluorescente in centinaia di cellule in una varietà di condizioni.
Il periodo osservato di singole cellule in coltura è più lungo del periodo somitogenesis nell'embrione intatta. Osserviamo che un espianto PSM intatta nella cultura espone anche oscillazioni più lenti della embrione intatto (dati non mostrati), suggerendo that il periodo più lungo osservato in cellule isolate non è semplicemente a causa di danni da dispersione. Un periodo ed ampiezza variabili sono osservati in più di una serie temporale della singola cellula. La fonte di questa variabilità non è nota, ma dovrebbe rivelare importanti dettagli su circuiti ritmo, rendendo il dell'orologio segmentazione.
Con questo metodo possiamo studiare le componenti genetiche di segmentazione a livello cellulare, e le domande di indirizzo che sono difficili da esaminare l'intero embrione. Ad esempio, per aggiunta controllata di molecole conosciute segnalazione trovati negli embrioni alle nostre culture, possiamo esaminare i loro effetti sul singolo oscillatore cellulare PSM in un saggio robusto e riproducibile. Il nostro sistema di coltura PSM apre le porte ad una rigorosa valutazione di quali fattori, da soli o in combinazione promuovere oscillazioni in queste cellule, quali fattori inibiscono queste oscillazioni, e per testare l'interazione tra tali molecole. Con questi strumenti in mano, ci proponiamo divalutare i modelli esistenti di somitogenesis che si basano su dati a livello di tessuto pubblicati, così come l'uso sono risultati in singole cellule di generare previsioni che possono essere testati in tutto embrione.
A nostra conoscenza, questo è il primo protocollo di coltura cellulare primaria zebrafish per acuti registrazione time-lapse della fluorescenza in singole cellule; ulteriore sviluppo e perfezionamento di questo protocollo è senza dubbio possibile. Altri protocolli spesso utilizzano embrioni di zebrafish per generare linee cellulari stabili che possono essere trasfettate con i reporter ed utilizzate per l'imaging a lungo termine 27-29. Mentre le linee stabili sono utili per l'imaging di un processo che non è legata a tempi di sviluppo, come l'orologio circadiano, la questione della segmentazione embrionale richiede dell'immagine immediata mentre le cellule sono ancora nella loro oscillatorio, stato progenitore. Una volta che le cellule smettono di oscillare, assumono un destino cellula differenziata, e quando nel tessuto, dovrebbe essere incorporata in un somite. E 'possible che con il fattore di destra o fattori presenti in vitro da poter generare PSM-come le linee di zebrafish coltura cellulare che rimarrebbero oscillatorio, consentendo significativamente osservazioni a lungo termine, più perturbazioni sequenziali, o screening ad alto rendimento.
Ci aspettiamo che questo metodo di preparazione di colture primarie di cellule disperse per l'imaging time-lapse è particolarmente adatto per lo studio di ogni processo cellulare che si verifica entro un lasso di tempo di sviluppo che non è accessibile utilizzando linee cellulari di zebrafish stabili. L'isolamento di diversi tipi di cellule a stadi di sviluppo distinti da adeguate linee giornalista transgeniche, sia attraverso la dissezione o tramite FACS dopo dissociazione embrionale, potrebbe fornire le cellule di partenza. Alcuni ottimizzazione delle condizioni di coltura, guidati dalla origine embrionale delle cellule, può essere richiesto. Combinando questo protocollo flessibile e sensibile con la rapida crescita collezione di zebrafish transgenicolinee, speriamo di facilitare un approccio evolutivo vitro in biologia che è complementare ai metodi genetici e embriologici classici.
Divulgazioni
Contributi Autore:
ABW sviluppato e affinato la dispersione, la cultura, l'imaging, e protocolli di monitoraggio delle cellule. DS ha generato le linee transgeniche e ha curato la progettazione del microscopio a fluorescenza time-lapse utilizzato in questo protocollo. AO pioniere esperimenti iniziali proof-of-principio di dissociarsi e le cellule immagine PSM in coltura. JS ha scritto lo strumento plug-in cerchio interpolatore in Fiji utilizzato per misurare l'intensità di fluorescenza da filmati time-lapse. ABW e ACO ha scritto il manoscritto.
Riconoscimenti
Questo lavoro è stato supportato da un EMBO lungo termine fellowship (ABW), un National Science Foundation Internazionale Postdoctoral Research Fellowship (ABW), la Max Planck Gesellschaft (ABW, DS, JS, ACO), una borsa di studio scava-BB (AO), e il Consiglio europeo della ricerca nell'ambito delle Comunità europee del Settimo programma quadro STG-207634 (DS, ACO). Ringraziamo Ravi Desai per gli utili commenti sul manoscritto. Vorremmo anche ringraziare la funzione di espressione della proteina MPI-CBG per la produzione del frammento zebrafish Fibronectin1, il personale della struttura pesce MPI-CBG per la cura e la manutenzione delle nostre linee di pesce e la funzione di microscopia ottica MPI-CBG per il supporto di immagini.
Materiali
| Name | Company | Catalog Number | Comments |
| Name of Reagent/Material | Company | Catalog Number | Comments |
| Epifluorescence microscope | Olympus | Model: SZX16 | |
| Epifluorescence microscope | X-cite Illumination | Series: 120Q | |
| Dissection microscope | Olympus | Model: SZX12 | |
| Fine forceps no. 55 | Fine Science Tools | 11295-51 | |
| Glass transfer pipettes | Assistent | 567/2 | |
| 35 mm plastic petri dishes | Greiner | 627102 | |
| 60 mm plastic petri dishes | Greiner | 628102 | |
| Sylgard polymer | SASCO | 266727 | |
| Manipulation tools | Made in-house | For description of manipulation tools Ref. 22 | |
| Microsurgical knife | World Precision Instruments | 500249 | |
| L15 medium | Invitrogen | 57322 | |
| Penicillin/streptomyocin | PAA | P11-010 | |
| Fetal bovine serum | Invitrogen | 257322 | |
| 0.05% trypsin / 0.02% EDTA | PAA | L11-004 | |
| Gel loading tips | Fisher Scientific | 253188 | |
| Sigmacote | Sigma Aldrich | 254589 | |
| Micropipette set | Gilson International | F167300 | |
| Zebrafish Fibronectin1 70 kD fragment | MPI-CBG protein facility | Generated in-house from construct based on previously published work Ref. 23-25 | |
| Glass bottom imaging dishes | Mattek (single well) | P35G-1.5-14-C | |
| Glass bottom imaging dishes | Greiner (CellView -multi-well) | 262502 | |
| E3 medium without methylene blue | Made in-house | From The Zebrafish Book, 5th Ed. Ref. 26 | |
| Plastic transfer pipettes | Ratiolab | 260011 | |
| Warner heating/cooling chamber | Warner Instruments | TC-324B/344B | |
| EM-CCD camera | Andor | Model: iXOn 888 | |
| Wide-field fluorescence microscope with Venus filter set | Zeiss | Model: Axiovert 200M | |
| Wide-field fluorescence microscope with Venus filter set | NeoFluor 40x, NA 0.75 | ||
| Wide-field fluorescence microscope with Venus filter set | Lumencor Light Engine | Model: Spectra X | |
| Wide-field fluorescence microscope with Venus filter set | |||
| Wide-field fluorescence microscope with Venus filter set | BrightLine HC 575/15 | F39-575 |
Riferimenti
- Cooke, J., Zeeman, E. C. A clock and wavefront model for control of the number of repeated structures during animal morphogenesis. J Theor Biol. 58, 455-476 (1976).
- Oates, A. C., Morelli, L. G., Ares, S. Patterning embryos with oscillations: structure, function and dynamics of the vertebrate segmentation clock. Development. 139, 625-639 (2012).
- Dequeant, M. L., Pourquie, O. Segmental patterning of the vertebrate embryonic axis. Nat Rev Genet. 9, 370-382 (2008).
- Oates, A. C., Gorfinkiel, N., Gonzalez-Gaitan, M., Heisenberg, C. P. Quantitative approaches in developmental biology. Nat Rev Genet. 10, 517-530 (2009).
- Oates, A. C., Ho, R. K. Hairy/E(spl)-related (Her) genes are central components of the segmentation oscillator and display redundancy with the Delta/Notch signaling pathway in the formation of anterior segmental boundaries in the zebrafish. Development. 129, 2929-2946 (2002).
- Lewis, J. Autoinhibition with transcriptional delay: a simple mechanism for the zebrafish somitogenesis oscillator. Curr Biol. 13, 1398-1408 (2003).
- Schroter, C., Oates, A. C. Segment Number and Axial Identity in a Segmentation Clock Period Mutant. Curr Biol. 20, 1254-1258 (2010).
- Riedel-Kruse, I. H., Muller, C., Oates, A. C. Synchrony dynamics during initiation, failure, and rescue of the segmentation clock. Science. 317, 1911-1915 (2007).
- Ozbudak, E. M., Lewis, J. Notch signalling synchronizes the zebrafish segmentation clock but is not needed to create somite boundaries. PLoS Genet. 4, e15 (2008).
- Herrgen, L., et al. Intercellular Coupling Regulates the Period of the Segmentation Clock. Curr Biol. 20, 1244-1253 (2010).
- Sawada, A., et al. Fgf/MAPK signalling is a crucial positional cue in somite boundary formation. Development. 128, 4873-4880 (2001).
- Giudicelli, F., Ozbudak, E. M., Wright, G. J., Lewis, J. Setting the tempo in development: an investigation of the zebrafish somite clock mechanism. PLoS Biol. 5, e150 (2007).
- Herrgen, L., Schroter, C., Bajard, L., Oates, A. C. Multiple embryo time-lapse imaging of zebrafish development. Methods Mol Biol. 546, 243-254 (2009).
- Hirata, H., et al. Oscillatory expression of the bHLH factor Hes1 regulated by a negative feedback loop. Science. 298, 840-843 (2002).
- Maroto, M., Dale, J. K., Dequeant, M. L., Petit, A. C., Pourquie, O. Synchronised cycling gene oscillations in presomitic mesoderm cells require cell-cell contact. Int J Dev Biol. 49, 309-315 (2005).
- Reppert, S. M., Weaver, D. R. Coordination of circadian timing in mammals. Nature. 418, 935-941 (2002).
- Welsh, D. K., Imaizumi, T., Kay, S. A. Real-time reporting of circadian-regulated gene expression by luciferase imaging in plants and mammalian cells. Methods Enzymol. 393, 269-288 (2005).
- Soroldoni, D., Oates, A. C. Live transgenic reporters of the vertebrate embryo's Segmentation Clock. Curr Opin Genet Dev. 21, 600-605 (2011).
- Masamizu, Y., et al. Real-time imaging of the somite segmentation clock: revelation of unstable oscillators in the individual presomitic mesoderm cells. Proc Natl Acad Sci USA. 103, 1313-1318 (2006).
- Soroldoni, D., Hogan, B. M., Oates, A. C. Simple and efficient transgenesis with meganuclease constructs in zebrafish. Methods in molecular biology. 546, 117-130 (2009).
- Schroter, C., et al. Dynamics of zebrafish somitogenesis. Dev Dyn. 237, 545-553 (2008).
- Picker, A., Roellig, D., Pourquie, O., Oates, A. C., Brand, M. Tissue micromanipulation in zebrafish embryos. Methods in molecular biology. 546, 153-172 (2009).
- Mould, A. P., et al. Identification of multiple integrin beta1 homologs in zebrafish (Danio rerio). BMC Cell Biol. 7, 24 (2006).
- Mould, A. P., Koper, E. J., Byron, A., Zahn, G., Humphries, M. J. Mapping the ligand-binding pocket of integrin alpha5beta1 using a gain-of-function approach. Biochem J. 424, 179-189 (2009).
- Zhao, Q., Liu, X., Collodi, P. Identification and characterization of a novel fibronectin in zebrafish. Exp Cell Res. 268, 211-219 (2001).
- Westerfield, M. In The zebrafish book : a guide for the laboratory use of zebrafish (Brachydanio rerio). , (1993).
- Vallone, D., Santoriello, C., Gondi, S. B., Foulkes, N. S. Basic protocols for zebrafish cell lines: maintenance and transfection. Methods Mol Biol. 362, 429-441 (2007).
- Carr, A. J., Whitmore, D. Imaging of single light-responsive clock cells reveals fluctuating free-running periods. Nat Cell Biol. 7, 319-321 (2005).
- Whitmore, D., Foulkes, N. S., Sassone-Corsi, P. Light acts directly on organs and cells in culture to set the vertebrate circadian clock. Nature. 404, 87-91 (2000).
- Soroldoni, D., et al. A Doppler effect in embryonic pattern formation. Science. 345, 222-225 (1126).
Ristampe e Autorizzazioni
Richiedi autorizzazione per utilizzare il testo o le figure di questo articolo JoVE
Richiedi AutorizzazioneThis article has been published
Video Coming Soon
Personale delle biblioteche
Copyright © 2025 MyJoVE Corporation. Tutti i diritti riservati