
Method Article
Geração de Disperso Presomitic mesoderme culturas de células for Imaging da Segmentação Relógio Zebrafish em células individuais
Neste Artigo
Resumo
Somitogenesis é um processo de desenvolvimento rítmico que espacialmente padrões do eixo do corpo de embriões vertebrados. Anteriormente, desenvolvemos linhas de zebrafish transgênicos que usam repórteres fluorescentes para observar os genes cíclicos que impulsionam esse processo. Aqui, a cultura dispersas células destas linhas e imagem suas oscilações ao longo do tempo in vitro.
Resumo
A segmentação é um processo morfogenético periódica e sequencial em vertebrados. Esta formação rítmica de blocos de tecido chamados somitos ao longo do eixo do corpo é a evidência de um oscilador genético padronização do embrião em desenvolvimento. No peixe-zebra, o relógio intracelular segmentação de condução é composta por membros da família de fator de transcrição Ela / Ele é organizado em ciclos de feedback negativo. Geramos linhas recentemente repórter fluorescentes transgénicos para o gene de HER1 cíclico que recapitular o padrão espacio-temporal de oscilações na mesoderme presomitic (PSM). Utilizando estas linhas, foi desenvolvido um sistema de cultura in vitro que permite a análise em tempo real do relógio oscilações de segmentação, dentro de células de PSM individuais isolados. Ao remover o tecido PSM de embriões transgênicos e dispersar as células de oscilação regiões em placas com fundo de vidro, geramos culturas apropriadas para lapso de tempo de imagem de sinal de fluorescência decélulas-relógio individual. Esta abordagem fornece uma estrutura experimental e conceitual para a manipulação direta do relógio da segmentação, com resolução de uma única célula sem precedentes, permitindo que suas propriedades de células-autônoma e em nível de tecido a ser distinguido e dissecados.
Introdução
A formação contínua de segmentos ao longo do eixo do corpo de vertebrados, ou somitogénese, é uma evidência de um oscilador espacial e temporal no embrião em desenvolvimento. O mecanismo favorecido controlando somitogenesis é conceitualmente descrito por um "relógio e frente de onda" modelo 1, em que o "relógio", que consiste de osciladores celulares, agora pensado para ser intracelularmente impulsionada pela expressão rítmica de um conjunto de genes cíclicos 2, carrapatos fora da formação de somitos a partir da mesoderme presomitic (PSM). À medida que o embrião se desenvolve, um amadurecimento "de frente de onda" na PSM se move em conjunto com o tecido regredir para a parte posterior, retardando e prendendo osciladores celulares que passa 3. Em conjunto, este sistema espaçotemporalmente dinâmico é denominado o relógio da segmentação. As abordagens atuais para estudar o relógio período segmentação três níveis crescentes de organização do oscilador genético em células individuais a dia acoplamento locaisa ocorrer entre células e, finalmente, a regulação global da informação posicional no tecido PSM coletiva 4.
Estudos anteriores sugerem que a célula-autônoma segmentação oscilador no peixe-zebra é composto de genes e produtos de proteína do her / hes transcrição factorfamily, que são pensados para formar um loop de feedback negativo através da repressão transcricional 5-7. A via de sinalização Delta / Notch sincroniza oscilações entre células vizinhas e regula o período colectiva da população 8-10. Moléculas fgf sinalização produzidas na tailbud aparecem para construir um gradiente entre os PSM do peixe-zebra, e são, assim, a hipótese de contribuir para abrandar e prendendo as células que oscilam na face anterior 11. Até agora, os papéis funcionais de cada uma destas moléculas em somitogénese foram investigados por mutação genética, injecção morfolino, choque térmico sobre-expressão, e antagonista trea drogatamento de componentes de relógio e de sinalização entre as células 5,7,10,12. Usando estas perturbações, função relógio da segmentação foi inferida a partir de descrições de nível de tecido de defeitos somito ea perda de oscilações uniformes na expressão de genes cíclicos como HER1, her7 e delta C. No entanto, quase todos estes dados são a partir de embriões fixas e não conseguem captar com precisão as mudanças que são inerentes à função dinâmica do relógio da segmentação. Mais recentemente, de vários embriões de lapso de tempo de imagem revelou os primeiros mutantes com período oscilador alterada, mas essas observações também foram feitas no nível do tecido 7,13. Assim, não foi observado o comportamento do oscilador de células-autônoma hipótese durante somitogenesis.
Instantâneos estáticos de somitogenesis dar uma imagem incompleta, porque, por natureza, o processo é conduzido por um sistema oscilatório. Trabalhos anteriores em células de camundongo e de bico mostrou que os níveis de transcriçãoe aumento de proteína e cair, mas a amostragem uma oscilação de aproximadamente 2 horas a cada 30 ou 45 min necessariamente restringe os dados coletados e, portanto, as conclusões que podem ser extraídas 14,15. Estudo de outros osciladores biológicos, principalmente, relógios circadianos, afastou-se das medições encenadas de expressão gênica e protéica de monitoramento em tempo real usando fluorescente e repórteres bioluminescentes 16,17. Estas ferramentas são essenciais para demonstrar propriedades de clock de 18 células individuais. Um repórter bioluminescente do gene cíclico HES1 foi desenvolvido e brevemente caracterizado nas células do rato PSM individuais 19. O período médio e variação foram calculados para um pequeno número de células, que mostra que as oscilações persistir durante vários ciclos in vitro. No entanto, esses estudos não quantitativamente abordar a estabilidade e robustez da freqüência de oscilação e amplitude, se as células podem entrar espontaneamente ou oscilações de saída,e como as células mantêm as suas relações de fase. Além disso, os efeitos de moléculas de sinalização encontrados no embrião no relógio célula-autónoma não foram directamente testados. Por conseguinte, estas propriedades fundamentais do oscilador de célula única permanecem completamente desconhecido.
Recentemente, desenvolveu linhas de peixes transgênicos utilizando BAC recombineering 20 para dirigir Venus (YFP) repórteres de fluorescência de expressão HER1 30. Tais linhas explorar as pistas de regulação do locus cromossômico intacto em que estão inseridas e recapitular a dinâmica temporal e padrão espacial de HER1. Esta descoberta permite o monitoramento em tempo real da expressão do gene no embrião de peixe-zebra desenvolvimento in vivo. Para estudar as propriedades fundamentais das oscilações celulares-autônoma e como tal expressão é regulada ao longo do tempo, que recentemente desenvolveu um método confiável para isolar e gravar a partir de células PSM in vitro. Este protocol descreve como nós utilizamos nossas linhas repórter transgênicos para gerar culturas de células dispersas, a partir do qual podemos caracterizar as oscilações do relógio da segmentação zebrafish em células individuais. Podemos, assim, resolver questões pendentes no campo que não eram acessíveis com a análise de nível de tecido ou estática, bem como manipular diretamente o relógio da segmentação no nível de uma única célula com moléculas de sinalização e inibidores.
Protocolo
1. Antes Dissecção
- No dia antes da dissecação, obter embriões a partir de um dos pares de peixe-zebra incross heterozigóticas para o alelo transgénico.
- Levante embriões em meio E3 sem azul de metileno a 28 o C até o estágio de blindagem (6 horas pós-fertilização).
- Transferência de embriões em meio E3 sem azul de metileno a 20 o CO / N. Aos 20 o C, os embriões formarão um somite por hora, uma vez que eles atinjam fase tailbud. Após 17-20 h de O / N a 20 ° C, os embriões devem ser em 5 a 8 somito fase na manhã seguinte ao início do protocolo de dissecção.
- Montar ferramentas e reagentes necessários para dissecção.
- Pipeta de vidro polido-Fire para a transferência de embriões e tecido
- Par de uma pinça fina para remoção de córions de embriões
- Sylgard-revestido de 35 mm de prato de dissecação - Pour Sylgard polímero num prato de 35 mm e curar S / N a 37 ° C. Faça a dissecção bem usandouma ponta de agulha para remover uma pequena quantidade de polímero curada. O prato pode ser limpa e subsequentemente reutilizado.
- Afiadas, ferramentas de fio de tungstênio achatadas para manipulação de tecidos PSM
- Micro-bisturi para cortar pedaços desejados de PSM para a cultura
- 35 mm prato de plástico para tripsina incubação
- Meio L15 contendo 10% de soro fetal bovino
- Solução de tripsina / EDTA a 0,25%
- Sigmacoted dicas gel de carregamento para a dispersão
- Placas de Petri de plástico contendo meio E3 sem azul de metileno.
- Cubra a assadeira de imagens com fundo de vidro com substrato Fibronectin1 (10 mg / ml em PBS). Deixe prato no banco para cobrir durante a dissecção.
- Use estereoscópio com filtros de fluorescência apropriadas para identificar e embriões transgênicos tipo positivos (fase 5-8 somite).
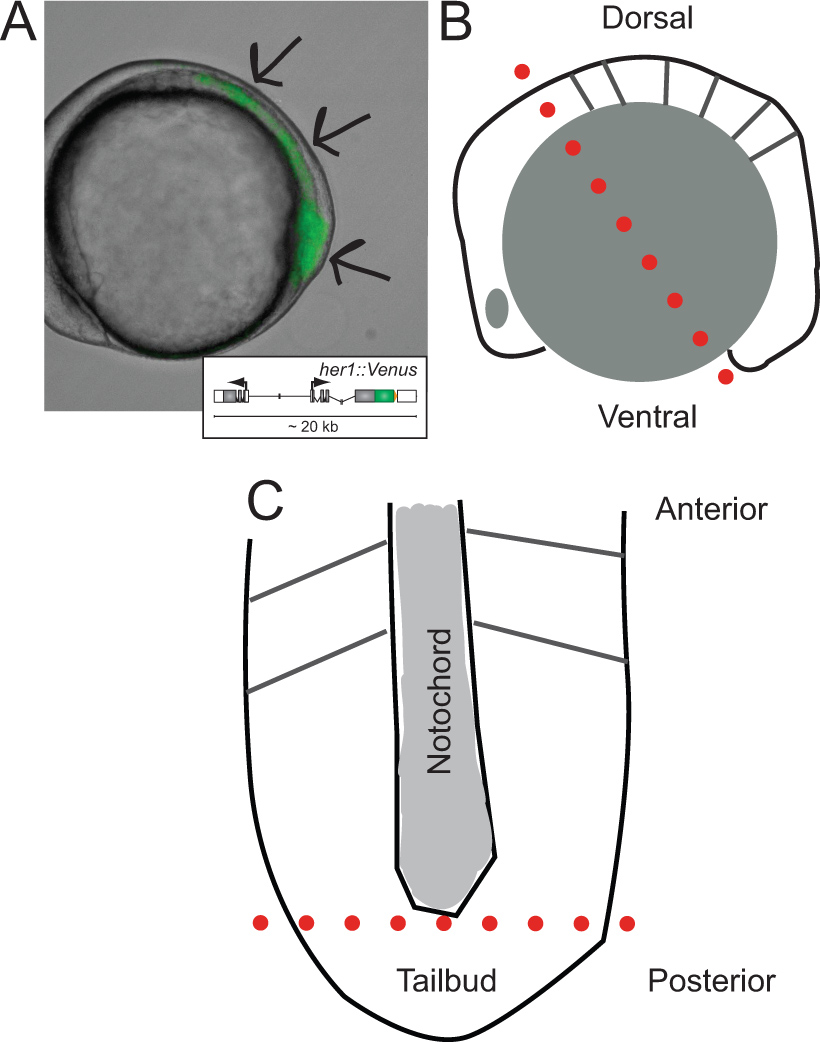
- Identificar os embriões transgénicos por análise de mesoderme presomitic (PSM) para expressão YFP sob canal de fluorescência (Figura 1A). Fluorescência shoULD ser visível na região desde o último somite formada para o tailbud. Selecione embriões com sinal mais brilhante; 25% de descendência devem ser embriões portadores homozigóticos 2 cópias do transgene. O número de embriões identificados necessário varia de acordo com a experiência. Um pedaço tailbud dissecados produzirá 1.000 células, em média. Normalmente, alguns embriões positivos adicionais são úteis no caso são feitos erros durante a dissecção.
- Use luz transmitida como uma referência de posicionamento para distinguir qualquer autofluorescência, particularmente na gema de célula, a partir do sinal.
- Transferência de embriões positivos para uma placa de Petri contendo meio de plástico separado E3 sem azul de metileno.
2. PSM Dissecção e Dispersão
- Prepare embriões para dissecção.
- Sob um microscópio de dissecação, usando uma pinça fina, retire cuidadosamente o córion de cada embrião no meio E3. Cuidado para não danificar o embrião ou interromper o celular gema.
- Encha Sylgard revestido dissecando prato com meio L15 com soro. Utilizando uma pinça ou outra ferramenta plana, remova as bolhas de ar da superfície do Sylgard.
- Transferência de embriões dechorionated utilizar o vidro pipeta-fogo polido para o prato dissecção.
- Usando as ferramentas de arame, mover todos os embriões para um lado do prato de dissecação.
- Dissecar PSM a partir de um único embrião.
- Orientar um único embrião no seu lado lateral, na pequena bem feito na camada de Sylgard dentro do prato de dissecação (Figura 1B).
- Usando o micro-bisturi, fatia através do embrião e das células gema apenas anterior ao cérebro posterior e pelo pólo ventral do embrião (Figura 1B, pontilhada linha vermelha).
- Retire a peça anterior do embrião a partir do bem, transferindo-o para o lado do prato, e, em seguida, usar as ferramentas de arame para raspar restantes grânulos celulares gema da seção posterior, incluindo o PSM.
- Uma vez que a gema tem sido raspado, achatar e orientar o PSM com a extremidade anterior que aponta para longe do experimentador e posterior que aponta para o experimentador (Figura 1C).
- Se a fina camada de ectoderma não tem puxado para fora por este ponto, use as ferramentas de arame para descascá-lo longe do topo do tecido PSM.
- Usando o micro-corte do bisturi tailbud, a ponta mais posterior da PSM para além da extremidade da notocorda, afastado do resto do tecido (Figura 1C, linha pontilhada vermelha). NOTA: outros pedaços de tecido a partir do PSM, para o PSM exemplo anterior mais perto do último somito formado, pode também ser feita em cultura, dependendo da questão experimental.
- Mova a peça tailbud para um canto do prato longe da área de dissecação. Use as ferramentas de arame para limpar todos os detritos e tecido embrião indesejado do campo de dissecação.
- Repita com a próxima embrião.
- Peças Piscina tailbud de múltiplas embryos, dependendo do número de células vai ser necessária para o ensaio. Em média, um único pedaço de tecido tailbud rende 1.000 células.
- Encha vazio de 35 mm de prato plástico com um pequeno volume de tripsina-EDTA.
- Utilizar o vidro pipeta-fogo polido, transferir peças tailbud de dissecar prato em prato contendo tripsina / EDTA. Incubar peças tailbud em tripsina / EDTA durante 20 minutos à temperatura ambiente.
- Enquanto o tecido está a ser incubado em tripsina, remover a solução Fibronectin1 da imagiologia prato de fundo de vidro.
- Lava-se a solução de vidro 3 vezes com água MilliQ. Use sucção para remover cada lavagem e garantir o prato esteja completamente seco.
- Dispersar peças tailbud em meio para a imagem latente.
- Pipetar 100 mL de meio L15 com soro em imagens prato.
- Utilizando uma ponta de gel revestida, remover os pedaços tailbud de tripsina / EDTA em que um volume pequeno quanto possível.
- Pipetar as peças tailbud para o meio noprato de imagem. Pipetar as peças para cima e para baixo várias vezes para separá-las e suspender as células no meio. Tenha cuidado para não introduzir bolhas de ar. Verifique a existência de aglomerados de células ao microscópio e dispersar mais, conforme necessário.
- Permitir que as células dispersas na suspensão assente em cima do vidro Fibronectin1 revestido durante 20 minutos à temperatura ambiente.
- Adicionar um volume pequeno de meio L15 adicional com soro para as células antes do início de imagem. Tenha cuidado para não perturbar as células resolvido.
- Dada a questão experimental, adicionar qualquer suplemento ou tratamentos com drogas para a cultura.
3. Imagem latente de células dispersas PSM
- Configure imagem prato em câmara de temperatura na imagem microscópio time-lapse. Definir Warner câmara até à temperatura desejada para a experiência. Permitir prato a equilibrar durante pelo menos 30 min antes de iniciar a aquisição de imagem. Temperatura Crosscheck com sonda de temperatura externa colocada no prato, se necessário. NOTA: Devido aa relação entre a temperatura ea taxa de somitogenesis 21 controle de temperatura estável é essencial para medições precisas de período em células PSM individuais.
- Adquirir imagens de teste do canal de fluorescência para verificar que o tempo de exposição e ganho de fornecer uma grande gama dinâmica das intensidades, sem saturação para assegurar um bom sinal de níveis de ruído. A imagem típica fluorescência adquiridos a partir de células dispersas gerados a partir de nossas linhas usando este protocolo requer 400 ms e 40 ms para uma imagem de luz transmitida com uma câmera EMCCD operando com um ganho EM de 85. Além disso, o ganho de pré-amplificador e velocidade de leitura do câmera também são essenciais para maximizar o sinal sobre o ruído.
- Usando o canal de luz transmitida, escolher campos de células para a aquisição de lapso de tempo.
- Executar protocolo de aquisição de lapso de tempo para período de tempo desejado.
- Adquirir uma imagem de fluorescência por um campo de luz transmitida e. NOTA: Defina um intervalo entre ro de aquisiçãounds que irá capturar dinâmica temporal sem foto-branqueamento sobre imagem ampliada ou toxicidade induzindo nas células. Este protocolo utiliza um intervalo de 2 min.
- Verifique lapso de tempo set-up de vez em quando durante a gravação para garantir que as células permanecem em foco, nenhum software ou hardware erros, etc
4. Processamento de Imagens de Adquiridas Time-lapse Filmes
- Abrir arquivo de filme para um campo adquirida em um software de processamento de imagem. NOTA: Fiji foi usado para todo o processamento neste protocolo.
- Dividir transmitida quadros de luz e de fluorescência de um campo para criar 2 pilhas de imagens.
- Acompanhe uma única célula no canal de luz transmitida.
- Coloque um ROI circular (região de interesse) em torno da célula selecionada no primeiro quadro no canal de luz transmitida. NOTA:.. Apenas as células que são saudáveis 1 no final da gravação, 2 não se mover para fora do campo, e 3 não entram em contacto com a outra ce.lls são rastreadas.
- Salvar um ROI cada poucos quadros para o gerente de ROI. Acompanhe célula até que o último quadro.
- Medir a intensidade usando os ROIs salvos no canal de fluorescência.
- Selecione a pilha de fluorescência e com o ROIs salvo, use o círculo personalizado interpolador plug-in e macro para medir a intensidade da célula monitorado ao longo do tempo.
- Confira o traço saída da macro. Se o traço qualitativamente captura características da fluorescência de lapso de tempo, exportar os valores para uma planilha do Excel.
- Salvar a lista de ROI para a célula.
- Repita rastreamento no canal transmitido e medir no canal de fluorescência para outras células no campo.
- Repita todas as etapas para campos adicionais do experimento.
Resultados
Este protocolo produz culturas de células, dispersos, PSM únicos viáveis para lapso de tempo de imagem de sinal de fluorescência (Figura 2). Nossa transgene gera um repórter cujo ciclo de produção e degradação ocorre com dinâmica semelhante ao do gene endógeno de proteína e no embrião, no fim de meia hora. Devido ao seu volume de negócios rápido, o sinal YFP em células individuais devem ser detectados rapidamente para minimizar branqueamento e com uma resolução temporal alta para capturar as características de cada ciclo oscilatório. Além disso, dada a imprecisão relativa do sinal, as condições de cultura e de aquisição ter sido cuidadosamente ajustada para assegurar resultados sensíveis e robustos. Nós encontramos os seguintes fatores a ser importantes na geração de culturas de células ótimas PSM para a imagem: 1. Um substrato ECM usado para o revestimento das placas de cultura com fundo de vidro. 2. Adição de soro para a dissecção e meio de imagem. 3. Dissecção do PSM de embriões identificados tomadas após o acasalamento hetepares rozygous, cuja descendência potencialmente levar 2 cópias do transgene. 4. Aquisição de imagens dentro de uma gama de ampliação óptima, utilizando uma objectiva de maior NA para assegurar a captura eficiente do sinal de fluorescência. 5. Iluminação com uma fonte de luz de estado sólido para minimizar flutuações de intensidade que possam contribuir para o ruído de fundo. 6. Detecção de sinal com uma câmara de alta sensibilidade EM-CCD para maximizar o sinal de leitura.
Culturas sub-óptimas irão conter células que não permanecem arredondada e saudável durante a gravação. Colocámos a hipótese de que o substrato de ECM, um fragmento de fibronectin1 zebrafish mantém as células num estado indiferenciado PSM semelhante, em comparação com outros substratos normalmente utilizados como poli-lisina ou laminina. Outros substratos testadas causada células para achatar no vidro e a perda de sinal fluorescente oscilante ao longo da gravação. Nós também descobrimos que a adição de soro para o meio durante a dissecção, dispersãoea gravação foi importante não só para saciar a tripsina usada para dissociação, mas também aumentou a intensidade de fluorescência ao longo do fundo, provavelmente devido à melhoria da viabilidade celular. Para garantir a captação do sinal ideal de células individuais foi utilizada uma objetiva de 40x projetada especificamente para imagens de fluorescência (Zeiss Plano série NeoFluor) com uma NA alto. Descobrimos também que uma fonte de luz de estado sólido, desde a iluminação mais estável do que as lâmpadas de mercúrio tradicionais, o que é fundamental para minimizar as flutuações de fundo que contribuem para imagens ruidosas. Essas modificações são importantes para garantir resultados robustos.
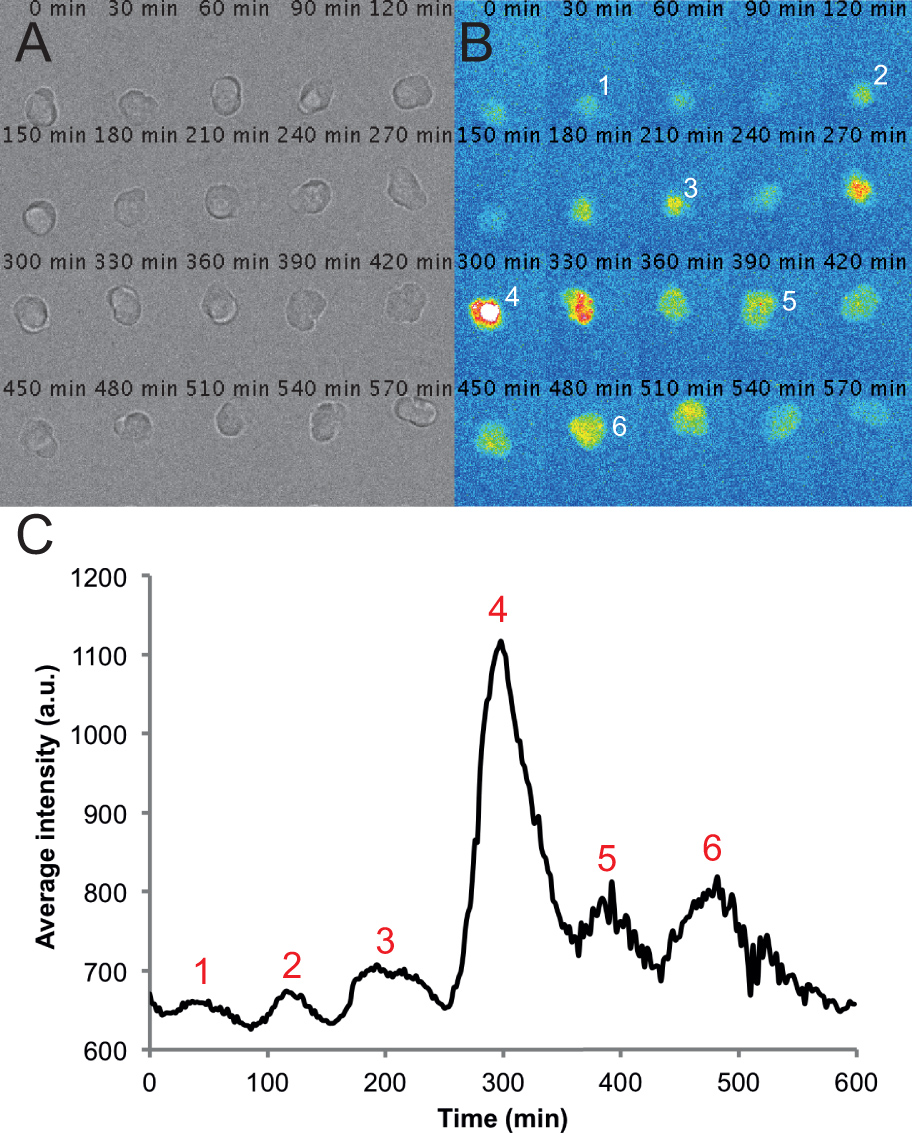
Usando este protocolo, esperamos culturas em que a maioria das células de um determinado campo são fluorescentes, em algum momento durante a gravação. Descobrimos que as células fluorescentes normalmente permanecem arredondado e às vezes são bastante móveis durante as gravações. Algumas células, incluindo as células fluorescentes, podem tornar-se apoptótico durante a gravação. Estas células são excluídos de qualquer umnálise. Também excluir as células que entram em contato com outras células ou se mover para fora do campo de visão. Em média, vemos uma redução de 12% no número de células por campo pelo fim da gravação 10 horas tal como medido por contagem do número de células saudáveis no canal de luz transmitida. Esta perda inclui tanto a morte das células e células que foram movidos para fora do campo. Dadas essas ressalvas, podemos, em média, faixa 5 células fluorescentes por campo que permanecem viável e visível, e tipicamente gravar seis campos por condição. Por exemplo, um experimento com 4 condições terá 24 campos de total adquirido e, normalmente, cerca de 30 células rastreadas por condição. Sob condições normais de imagem com meio L15 contendo 10% de soro fetal bovino, descobrimos que as células PSM (n = 101 células de quatro experiências em replicado) pode produzir entre 2 e 7 picos, com a média e o desvio padrão de 3 ± 1 (2 picos ciclos -3). O número de pico média, bem como o percentil 25%, é de 2 picos e o percentil 75% de is três picos. Usando o nosso acompanhamento e análise semi-automatizada, podemos gerar rapidamente intensidade de fluorescência sobre traços de tempo para as células PSM individuais, com um traço representativo célula mostrada na Figura 2C. Estes vestígios matérias pode ser usado para fazer medições quantitativas de propriedades das células PSM oscilatórios, tais como a frequência, a amplitude, o número de ciclos e tempo de picos.

Figura 1. Identificação de embriões transgênicos e esquemas de sua dissecção. (A) Vista lateral de um embrião de peixe-zebra transgênico expressando YFP fluorescência em fase ~ 5 somite. Imagem é uma sobreposição de luz transmitida e canal de fluorescência. As setas indicam áreas de sinal a partir da ponta da tailbud, durante todo o PSM, em relação a l ast formado somite. Inset mostra um esquema do transgene repórter (para mais informações sobre o seu desenvolvimento e comportamento in vivo referem-se a 30 Soroldoni et al.. (B) Esquema de vista lateral do embrião antes do primeiro corte durante a dissecção. Pontilhada linha vermelha indica o primeiro corte feitos em toda a parte posterior do cérebro, embora a célula gema apenas atrás do tailbud. (C) Representação esquemática de achatada PSM após a remoção de grânulos de vitelo e camada epidérmica, orientada ao longo do eixo ântero-posterior. pontilhada linha vermelha indica o segundo corte para remover a ponta do tailbud para a dispersão e cultura. Clique aqui para ver uma versão maior desta figura.
{kind=link}
07fig2.jpg "/>
Figura 2. Imagens representativas e os traços de uma única célula em uma cultura PSM dispersa. (A) Montagem de imagens de luz transmitida de uma única célula PSM em uma 6-h de gravação de lapso de tempo. Note-se que a célula permanece arredondada e saudáveis ao longo do curso do registo. (B) correspondente montagem de imagens de fluorescência a partir de uma única célula de PSM. Picos na intensidade da célula são numerados no decurso da gravação. (C) intensidade média ao longo do tempo medido a partir de um ROI colocado nesta célula. Os valores são retirados de cada quadro de um filme com uma taxa de quadros de um por 2 min usando o círculo interpolador plug-in personalizado e macro escrito em Fiji. Picos na intensidade são novamente contados em todo o traçado. Clique aqui para ver uma versão maior desta figura.
{kind=link}
Discussão
Para estudar um processo celular que ocorre ao longo do desenvolvimento embrionário, os biólogos normalmente utilizam uma abordagem no contexto de todo o embrião. No entanto, para entender completamente como uma única célula se comporta dentro de um período de desenvolvimento, um método para examinar e perturbar as células individuais em isolamento também é altamente benéfico. Ao gerar culturas de células dispersas PSM usando embriões de peixes-zebra transgénicos agora temos uma ferramenta para estudar diretamente a natureza autônoma da célula de oscilações genéticas no relógio da segmentação de uma forma quantitativa. É possível medir a dinâmica das oscilações no nosso repórter fluorescente em centenas de células sob uma variedade de condições.
O período observado de células isoladas em cultura é mais longo do que o período somitogenesis no embrião intacto. Observa-se que um explante PSM intactas em cultura também apresenta oscilações mais lentas do que o embrião intacta (dados não apresentados), sugerindo that o período de tempo observado em células isoladas não é simplesmente devido a danos causados por dispersão. Um período variável e amplitude é observado na maior parte de nossa série de tempo de uma única célula. A fonte dessa variabilidade é desconhecida, mas deve revelar detalhes importantes sobre circuitos de ritmo fazendo do relógio da segmentação.
Com esse método, podemos estudar os componentes genéticos de segmentação no nível celular e dirigir perguntas que são um desafio para examinar em todo o embrião. Por exemplo, por adição controlada de moléculas sinalizadoras conhecidas encontradas no embrião em desenvolvimento para as culturas, pode-se examinar os efeitos sobre o único oscilador celular PSM em um ensaio robusto e reprodutível. Nosso sistema de cultura PSM abre a porta a uma rigorosa avaliação de quais os fatores que, isoladamente ou em combinação promover oscilações nessas células, quais os fatores que inibem essas oscilações, e para testar a interação entre essas moléculas. Com essas ferramentas em mãos, pretendemosavaliar os modelos existentes das somitogénese que são baseadas em dados de nível de tecido publicados, assim como os resultados de utilização em células individuais para gerar previsões que podem ser testadas em todo o embrião.
Para o nosso conhecimento, este é o primeiro protocolo de cultura de células primárias de peixe-zebra para a gravação de lapso de tempo aguda de fluorescência em células individuais; maior desenvolvimento e refinamento deste protocolo é sem dúvida possível. Outros protocolos usam frequentemente embriões de peixes-zebra para gerar linhas celulares estáveis, que podem ser transfectadas com os repórteres e utilizados para imagiologia de longo prazo 27-29. Enquanto linhas estáveis são úteis para a imagem de um processo que não está vinculado ao tempo de desenvolvimento, como o relógio circadiano, a questão da segmentação embrionária requer imagiologia imediato, enquanto as células ainda estão em sua oscilatório, estado progenitor. Uma vez que as células param de oscilação, que assumem um destino celular diferenciada, e quando no tecido, poderá ser incorporado em um somito. É possible que com o fator ou fatores presentes in vitro direito poderíamos gerar PSM-como linhas de peixe-zebra de cultura de células que permanecem oscilatório, permitindo significativamente observações de longo prazo, vários perturbações seqüenciais, ou high-throughput screening.
Espera-se que este método de preparação de culturas primárias de células dispersas para imagiologia de lapso de tempo é bem apropriado para o estudo de qualquer processo celular que ocorre dentro de um período de desenvolvimento que não é acessível através de linhas celulares estáveis de peixe-zebra. O isolamento de células-tipos distintos em estágios de desenvolvimento diferentes das linhas repórter transgênicos apropriadas, quer através de dissecação ou através FACS após dissociação embrionário, poderiam fornecer as células iniciais. Alguns otimização das condições de cultivo, guiados pela origem embrionária das células, pode ser necessária. Ao combinar este protocolo flexível e sensível, com o rápido crescimento da coleção de peixes-zebra transgénicoslinhas, esperamos facilitar uma abordagem de desenvolvimento in vitro biologia que é complementar aos métodos genéticos e embriológicas clássicas.
Divulgações
Autor Contribuições:
ABW desenvolveu e refinou a dispersão, cultura, imagem e protocolos de rastreamento de celular. DS gerou as linhagens transgênicas e supervisionou o projeto do microscópio de fluorescência de lapso de tempo utilizado neste protocolo. AO pioneira iniciais experimentos de prova de princípio para dissociar e células imagem PSM em cultura. JS escreveu a ferramenta plugin do círculo interpolador em Fiji usado para medir a intensidade de fluorescência de filmes de lapso de tempo. ABW e ACO escreveu o manuscrito.
Agradecimentos
Este trabalho foi apoiado por uma EMBO companheirismo de longo prazo (ABW), a Science Foundation Internacional de Pós-Doutorado Bolsa de Investigação Nacional (ABW), a Max Planck Gesellschaft (ABW, DS, JS, ACO), uma bolsa CAVE-BB (AO), eo Conselho Europeu de Investigação de acordo com as Comunidades Europeias Sétimo Programa-Quadro STG-207634 (DS, ACO). Agradecemos Ravi Desai pelos comentários úteis sobre o manuscrito. Gostaríamos também de agradecer a facilidade de expressão de proteínas MPI-CBG para a produção do fragmento de peixe-zebra Fibronectin1, o MPI-CBG pessoal instalação de peixe para o cuidado e manutenção das nossas linhas de peixe e a facilidade de microscopia de luz MPI-CBG para suporte de imagem.
Materiais
| Name | Company | Catalog Number | Comments |
| Epifluorescence microscope | Olympus | Model: SZX16 | |
| Epifluorescence microscope | X-cite Illumination | Series: 120Q | |
| Dissection microscope | Olympus | Model: SZX12 | |
| Fine forceps no. 55 | Fine Science Tools | 11295-51 | |
| Glass transfer pipettes | Assistent | 567/2 | |
| 35 mm plastic petri dishes | Greiner | 627102 | |
| 60 mm plastic petri dishes | Greiner | 628102 | |
| Sylgard polymer | SASCO | 266727 | |
| Manipulation tools | Made in-house | For description of manipulation tools Ref. 22 | |
| Microsurgical knife | World Precision Instruments | 500249 | |
| L15 medium | Invitrogen | 57322 | |
| Penicillin/streptomyocin | PAA | P11-010 | |
| Fetal bovine serum | Invitrogen | 257322 | |
| 0.05% trypsin / 0.02% EDTA | PAA | L11-004 | |
| Gel loading tips | Fisher Scientific | 253188 | |
| Sigmacote | Sigma Aldrich | 254589 | |
| Micropipette set | Gilson International | F167300 | |
| Zebrafish Fibronectin1 70 kD fragment | MPI-CBG protein facility | Generated in-house from construct based on previously published work Ref. 23-25 | |
| Glass bottom imaging dishes | Mattek (single well) | P35G-1.5-14-C | |
| Glass bottom imaging dishes | Greiner (CellView -multi-well) | 262502 | |
| E3 medium without methylene blue | Made in-house | From The Zebrafish Book, 5th Ed. Ref. 26 | |
| Plastic transfer pipettes | Ratiolab | 260011 | |
| Warner heating/cooling chamber | Warner Instruments | TC-324B/344B | |
| EM-CCD camera | Andor | Model: iXOn 888 | |
| Wide-field fluorescence microscope with Venus filter set | Zeiss | Model: Axiovert 200M | |
| Wide-field fluorescence microscope with Venus filter set | NeoFluor 40x, NA 0.75 | ||
| Wide-field fluorescence microscope with Venus filter set | Lumencor Light Engine | Model: Spectra X | |
| Wide-field fluorescence microscope with Venus filter set | BrightLine HC 575/15 | F39-575 |
Referências
- Cooke, J., Zeeman, E. C. A clock and wavefront model for control of the number of repeated structures during animal morphogenesis. J Theor Biol. 58, 455-476 (1976).
- Oates, A. C., Morelli, L. G., Ares, S. Patterning embryos with oscillations: structure, function and dynamics of the vertebrate segmentation clock. Development. 139, 625-639 (2012).
- Dequeant, M. L., Pourquie, O. Segmental patterning of the vertebrate embryonic axis. Nat Rev Genet. 9, 370-382 (2008).
- Oates, A. C., Gorfinkiel, N., Gonzalez-Gaitan, M., Heisenberg, C. P. Quantitative approaches in developmental biology. Nat Rev Genet. 10, 517-530 (2009).
- Oates, A. C., Ho, R. K. Hairy/E(spl)-related (Her) genes are central components of the segmentation oscillator and display redundancy with the Delta/Notch signaling pathway in the formation of anterior segmental boundaries in the zebrafish. Development. 129, 2929-2946 (2002).
- Lewis, J. Autoinhibition with transcriptional delay: a simple mechanism for the zebrafish somitogenesis oscillator. Curr Biol. 13, 1398-1408 (2003).
- Schroter, C., Oates, A. C. Segment Number and Axial Identity in a Segmentation Clock Period Mutant. Curr Biol. 20, 1254-1258 (2010).
- Riedel-Kruse, I. H., Muller, C., Oates, A. C. Synchrony dynamics during initiation, failure, and rescue of the segmentation clock. Science. 317, 1911-1915 (2007).
- Ozbudak, E. M., Lewis, J. Notch signalling synchronizes the zebrafish segmentation clock but is not needed to create somite boundaries. PLoS Genet. 4, e15 (2008).
- Herrgen, L., et al. Intercellular Coupling Regulates the Period of the Segmentation Clock. Curr Biol. 20, 1244-1253 (2010).
- Sawada, A., et al. Fgf/MAPK signalling is a crucial positional cue in somite boundary formation. Development. 128, 4873-4880 (2001).
- Giudicelli, F., Ozbudak, E. M., Wright, G. J., Lewis, J. Setting the tempo in development: an investigation of the zebrafish somite clock mechanism. PLoS Biol. 5, e150 (2007).
- Herrgen, L., Schroter, C., Bajard, L., Oates, A. C. Multiple embryo time-lapse imaging of zebrafish development. Methods Mol Biol. 546, 243-254 (2009).
- Hirata, H., et al. Oscillatory expression of the bHLH factor Hes1 regulated by a negative feedback loop. Science. 298, 840-843 (2002).
- Maroto, M., Dale, J. K., Dequeant, M. L., Petit, A. C., Pourquie, O. Synchronised cycling gene oscillations in presomitic mesoderm cells require cell-cell contact. Int J Dev Biol. 49, 309-315 (2005).
- Reppert, S. M., Weaver, D. R. Coordination of circadian timing in mammals. Nature. 418, 935-941 (2002).
- Welsh, D. K., Imaizumi, T., Kay, S. A. Real-time reporting of circadian-regulated gene expression by luciferase imaging in plants and mammalian cells. Methods Enzymol. 393, 269-288 (2005).
- Soroldoni, D., Oates, A. C. Live transgenic reporters of the vertebrate embryo's Segmentation Clock. Curr Opin Genet Dev. 21, 600-605 (2011).
- Masamizu, Y., et al. Real-time imaging of the somite segmentation clock: revelation of unstable oscillators in the individual presomitic mesoderm cells. Proc Natl Acad Sci USA. 103, 1313-1318 (2006).
- Soroldoni, D., Hogan, B. M., Oates, A. C. Simple and efficient transgenesis with meganuclease constructs in zebrafish. Methods in molecular biology. 546, 117-130 (2009).
- Schroter, C., et al. Dynamics of zebrafish somitogenesis. Dev Dyn. 237, 545-553 (2008).
- Picker, A., Roellig, D., Pourquie, O., Oates, A. C., Brand, M. Tissue micromanipulation in zebrafish embryos. Methods in molecular biology. 546, 153-172 (2009).
- Mould, A. P., et al. Identification of multiple integrin beta1 homologs in zebrafish (Danio rerio). BMC Cell Biol. 7, 24 (2006).
- Mould, A. P., Koper, E. J., Byron, A., Zahn, G., Humphries, M. J. Mapping the ligand-binding pocket of integrin alpha5beta1 using a gain-of-function approach. Biochem J. 424, 179-189 (2009).
- Zhao, Q., Liu, X., Collodi, P. Identification and characterization of a novel fibronectin in zebrafish. Exp Cell Res. 268, 211-219 (2001).
- Westerfield, M. In The zebrafish book : a guide for the laboratory use of zebrafish (Brachydanio rerio). , (1993).
- Vallone, D., Santoriello, C., Gondi, S. B., Foulkes, N. S. Basic protocols for zebrafish cell lines: maintenance and transfection. Methods Mol Biol. 362, 429-441 (2007).
- Carr, A. J., Whitmore, D. Imaging of single light-responsive clock cells reveals fluctuating free-running periods. Nat Cell Biol. 7, 319-321 (2005).
- Whitmore, D., Foulkes, N. S., Sassone-Corsi, P. Light acts directly on organs and cells in culture to set the vertebrate circadian clock. Nature. 404, 87-91 (2000).
- Soroldoni, D., et al. A Doppler effect in embryonic pattern formation. Science. 345, 222-225 (1126).
Reimpressões e Permissões
Solicitar permissão para reutilizar o texto ou figuras deste artigo JoVE
Solicitar PermissãoThis article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. Todos os direitos reservados