
È necessario avere un abbonamento a JoVE per visualizzare questo. Accedi o inizia la tua prova gratuita.
Method Article
L'utilizzo del Ramsay Assay misurare la secrezione di fluidi e Ion Flux Prezzi in
In questo articolo
Riepilogo
Questo protocollo descrive l'uso del test Ramsay di misurare i tassi di secrezione di fluido dal isolate Malpighian (renale) tubuli da Drosophila melanogaster. Inoltre, l'uso di elettrodi ione-specifico per misurare le concentrazioni di sodio e potassio nel fluido secreto, consentendo il calcolo del flusso transepiteliale di ioni, è descritto.
Abstract
Modulazione di trasporto renale di ioni epiteliale permette organismi di mantenere l'omeostasi ionica e osmotico di fronte alle diverse condizioni esterne. La Drosophila melanogaster Malpighi (renale) tubulo offre un'opportunità senza precedenti per studiare i meccanismi molecolari del trasporto di ioni epiteliale, a causa delle potenti genetica di questo organismo e l'accessibilità dei suoi tubuli renali allo studio fisiologico. Qui, descriviamo l'uso del test Ramsay di misurare i tassi di secrezione di fluido dal isolato mosca tubuli renali, con l'uso di elettrodi ione-specifico per misurare le concentrazioni di sodio e potassio nel fluido secreto. Questo test consente di studio fluidi transepiteliale di ioni e flussi di ~ 20 tubuli alla volta, senza la necessità di trasferire il fluido secreto di apparecchiatura separata per misurare concentrazioni di ioni. Geneticamente tubuli distinti possono essere analizzati per valutare il ruolo di specifici geni nei processi di trasporto. Inoltre, il bsalina athing può essere modificato per esaminare gli effetti delle sue caratteristiche chimiche, o farmaci o ormoni aggiunti. In sintesi, questa tecnica permette la caratterizzazione molecolare dei meccanismi di base di trasporto ionico epiteliale nel tubulo Drosophila, nonché regolazione di questi meccanismi di trasporto.
Introduzione
Renale trasporto di ioni epiteliale sottende organismal iono- e osmoregolazione. La Drosophila melanogaster Malpighi (renale) tubulo offre un'opportunità senza precedenti per studiare i meccanismi molecolari del trasporto di ioni epiteliale. Ciò è dovuto alla combinazione delle potenti genetica di Drosophila, accoppiati con l'accessibilità dei suoi tubuli renali allo studio fisiologico. Il saggio Ramsay, dal nome del ricercatore che ha aperto la strada alla tecnica 1, misura i tassi di secrezione di fluidi da isolate tubuli malpighiani, ed è stato fondato nel Drosophila nel 1994 da Dow e colleghi 2. Questo ha spianato la strada a ulteriori studi utilizzando Drosophila strumenti genetici, come il sistema GAL4-UAS 3,4, per definire vie di segnalazione specifici delle cellule che regolano la secrezione di fluidi. Un esempio include segnalazione di calcio in risposta ad un ormone peptidico 5, tra molti altri 6,7.
ve_content "> una combinazione di tecniche genetiche e studio fisiologico classica ha dimostrato che la produzione di urina nella mosca avviene attraverso la secrezione di un potassio ricco cloruro fluido dal segmento principale del tubulo. Ciò avviene attraverso la secrezione transepiteliale parallelo di cationi, soprattutto K + ma anche Na +, attraverso la cella principale e Cl -. secrezione attraverso la cella stellate 8-12 La capacità di misurare separatamente transepiteliali K + e Na + flussi consente una caratterizzazione più dettagliata dei meccanismi di trasporto rispetto alla misurazione della secrezione di fluido alone. Ad esempio, in unstimulated tubuli Drosophila, Na + / K + -ATPasi inibitore ouabain non ha alcun effetto sulla secrezione di fluido 2, anche quando il suo assorbimento nelle cellule principali è inibito dal trasportatore anionico organico inibitore taurocolato 13. Tuttavia, Linton e O'Donnell ha mostrato che ouabain depolarizzail potenziale di membrana basolaterale, e aumenta Na + flusso 9. Come mostrato nelle Rappresentante dei risultati, abbiamo replicato questi risultati, e ha mostrato che K + flusso è contemporaneamente diminuito 14; l'aumento del flusso di Na + e diminuita K + flusso avere effetti opposti sulla secrezione di liquidi, con conseguente nessun cambiamento netto della secrezione. Così, ci sono due soluzioni a "paradosso ouabaina", cioè, l'osservazione iniziale che uabaina non ha alcun effetto sulla secrezione di fluido nel tubulo Drosophila:. Primo, in tubuli stimolato, l'effetto di ouabain sulla secrezione di fluido non è evidente a causa il suo assorbimento da parte del trasportatore anionico organico 13; e in secondo luogo, nei tubuli unstimulated, ouabain ha opposti effetti sulla transepiteliale Na + e K + flusso, con conseguente nessun variazione netta secrezione di fluidi (vedi Rappresentante dei risultati e rif. 9). Pertanto, il ruolo primario della Na + / K + -ATPase in tubuli non stimolati è quello di ridurre la concentrazione intracellulare di Na + per generare un gradiente di concentrazione favorevole per Na + -coupled processi di trasporto attraverso la membrana basolaterale. Infatti, misurando separatamente Na + e K + flussi, abbiamo dimostrato che tubuli manca la cotrasportatore fly sodio-potassio-2-cloruro (NKCC) sono diminuiti K transepiteliale + flusso, senza ulteriore calo dopo ouabaina aggiunta, e nessun cambiamento in transepiteliale Na + Flusso 14. Questi risultati supportati nostra conclusione che Na + di entrare nella cellula attraverso la NKCC viene riciclato attraverso il Na + / K + ATPasi. In un altro esempio, Ianowski et al. Ha osservato che l'abbassamento bagno K + concentrazione da 10 a 6 mm è diminuito transepiteliale K + flusso e aumento transepiteliale Na + flusso nel tubuli da Rhodnius prolixus, senza variazione netta secrezione di fluidi 15. Diversi effetti sulla Na + K + flusso e flusso attraverso tubuli larvali sono stati osservati anche nei tubuli Drosophila in risposta alle diverse diete sale 16 e in due specie di zanzare in risposta ad allevamento salinità 17.La sfida più grande nella misurazione del flusso transepiteliale di ioni nella preparazione dosaggio Ramsay è la determinazione delle concentrazioni di ioni all'interno del fluido secreto. Questa sfida è stata accolta con soluzioni diverse, tra cui la fiamma photometery 18, l'uso di ioni radioattivi 19, ed elettrone sonda lunghezza d'onda spettroscopia dispersiva 20. Queste tecniche richiedono trasferimento della goccia liquido secreto a uno strumento di misura della concentrazione di ioni. Poiché il volume di fluido secreto dalla non stimolato Drosophila tubulo è piccolo, tipicamente ~ 0,5 Nl / min, questo pone una sfida tecnica e introduce anche errore se parte del fluido è secretoperso al momento del trasferimento. Al contrario, l'uso di elettrodi ione-specifico permette la misura dell'attività ionica (da cui la concentrazione di ioni può essere calcolato) in situ. Il protocollo attuale è stato adattato da quella utilizzata da Maddrell e colleghi per misurare transepiteliale K + flusso attraverso il tubulo Rhodnius utilizzando valinomicina come il K + ionoforo 21, e descrive anche l'uso di un -butylcalix 4- ter [4] acido arene-tetraacetico tetraetile estere basato Na + -specifica elettrodi specifici ioni di litio caratterizzata da Messerli et. al. 22. Elettrodi Ion-specifici sono stati utilizzati anche per misurare le concentrazioni di ioni nel fluido secreto dalle tubuli malpighiani nel test Ramsay negli adulti 9,23 e larvale 16 Drosophila melanogaster, la Nuova Zelanda Alpine Weta (Hemideina maori) 24 e 17 zanzare.
Qui, descriviamo in dettaglio l'uso del Ramsaydire di misurare i tassi di fluido secrezione nei tubuli malpighiani da Drosophila melanogaster, nonché l'uso di elettrodi ione-specifico per determinare le concentrazioni di K + e Na + nel liquido secreto e quindi il calcolo dei flussi transepiteliale di ioni. Una panoramica del test è fornito in Figura 1.

Figura 1. Schema del Malpighi tubuli e la Ramsay test con l'uso di ioni a specifici elettrodi per misurare concentrazioni di ioni. Questa figura illustra la configurazione per il saggio Ramsay. (A) Ogni mosca ha quattro tubuli, un paio di tubuli anteriori e una coppia di tubuli posteriori, che galleggiano nella cavità addominale circondato da emolinfa. In ogni coppia, i due si uniscono a tubuli l'uretere, che poi svuota l'urina all'incrocio tra midgut e hindgut. I tubuli sono ciechi a tempo indeterminato. L'urina viene generato attraverso il segmento principale del fluido-secernente (mostrato in rosso), e scorre verso l'uretere e fuori nell'intestino. Dopo la dissezione, la coppia tubulo è dissociata dall'intestino da sezionare l'uretere. (B) La coppia di tubuli viene poi trasferita in una goccia di soluzione salina in un bagno ben del piatto dosaggio. Uno dei due tubuli, qui definito il "ancoraggio tubulo," è avvolto attorno ad un perno di metallo ed è inerte. L'altro è il tubulo tubulo secernente. Il segmento iniziale (che non secernono liquido) e il segmento principale del tubulo secernono rimangono entro la goccia di soluzione salina balneazione. Ioni e spostare l'acqua dalla salina di balneazione e nel lume del tubulo del segmento principale, e poi si muovono verso l'uretere, come accadrebbe in vivo. Il segmento inferiore (blu) è fuori dalla salina di balneazione e quindi inerte. Poiché l'uretere è tagliato, il liquido secreto emerge come una gocciolina dall'estremità tagliata dell'uretere. Tegli secreto fluido gocciolina allarga nel tempo la secrezione continua, e il suo diametro viene misurato con un micrometro oculare. Uno strato di olio minerale impedisce l'evaporazione del fluido secreto. Gli elettrodi di riferimento specifici e ioni misurano la concentrazione di ioni di liquido secreto. Cliccate qui per vedere una versione più grande di questa figura.
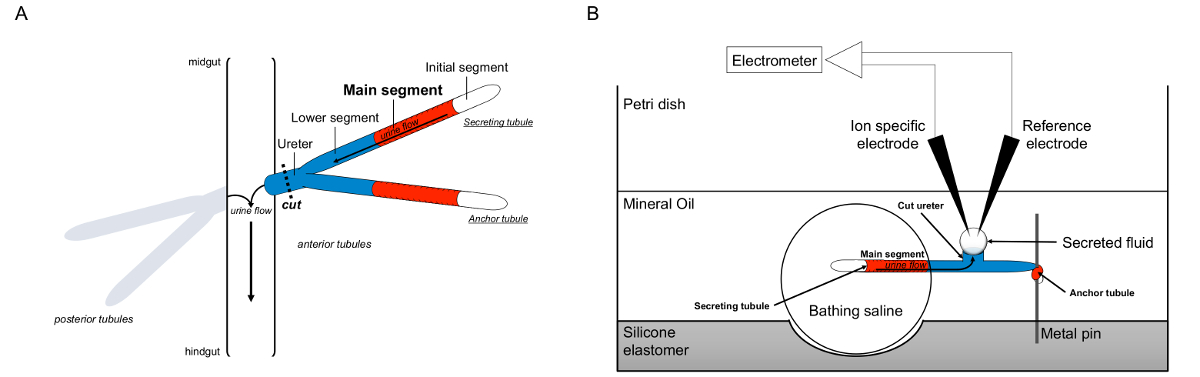{kind=link}
Protocollo
1. Preparazione la dissezione, taratura e Assay Piatti
Nota: In questo passaggio, tre scatole Petri in plastica rivestiti con elastomero siliconico vengono preparati: uno per la dissezione, uno per l'esecuzione del saggio Ramsay ("piatto assay"), ed uno per eseguire la calibrazione. Questi piatti sono riutilizzati da esperimento a esperimento, e, quindi, ciò deve essere ripetuta se si rompe un piatto. Un'immagine del piatto test è mostrato nella Figura 2.

Figura 2. Il piatto saggio. Il piatto utilizzato per il dosaggio Ramsay è mostrato qui. Si tratta di una capsula di Petri 10 cm che è rivestito con elastomero siliconico. Tra 20 e 25 pozzi sono scavate l'elastomero. Un perno di metallo Minutien, tagliate a metà, è posto a destra di ciascun pozzetto (oa sinistra, se lo sperimentatore è mancino).TPS: //www.jove.com/files/ftp_upload/53144/53144fig2large.jpg "target =" _ blank "> Clicca qui per vedere una versione più grande di questa figura.
- Utilizzando guanti, versare ~ 80 g elastomero siliconico di base in un bicchiere di vetro. Aggiungi 1/10 peso (8 g) elastomero siliconico cura. Mescolare con un agitatore di metallo. Posto su un agitatore orbitale con una cima piatta ad una velocità delicato, per esempio 100 rpm, per alcune ore fino a rimuovere tutte le bolle.
- Versare l'elastomero siliconico in capsule di Petri di plastica pulito: 100 mm x 15 mm piatti per i piatti di dissezione e di analisi, e 35 x 10 mm per il piatto di calibrazione. Lo spessore dello strato elastomerico deve essere ~ 6 - 7 mm per i piatti 100 mm, e ~ 5 mm per il piatto 35 mm.
- Porre le capsule in panchina a temperatura ambiente (RT) per curare (indurimento), ~ 24-48 ore.
- Una volta che il piatto test è guarito, preparare un gruppo più piccolo di elastomero siliconico come al punto 1.1, ad es., 10 g di base di silicone elastomero con 1 g di cura. Agitare su orbitaleshaker come al punto 1.1 fino a quando le bolle d'aria non sono più presenti. Questo elastomero verrà utilizzato nel passo 1.5.4 e non deve essere lasciato indurire prima di tale passo.
- Usando un bisturi chirurgico e precauzioni taglienti standard, fare pozzi in uno dei elastomero siliconico rivestite piastre di Petri da 100 mm. Questo sarà il piatto dosaggio.
- Rendere pozzi ~ 1 cm di distanza e con un diametro di ~ 3 - 4 mm. 25 pozzi possono facilmente adattarsi in un piatto di test a 100 mm. Wells dovrebbe essere di almeno 6 mm rimossi dalle pareti del piatto. Figura 2 illustra la spaziatura dei pozzi. Ogni pozzetto conterrà un tubulo secernente fluido durante l'esperimento. Così, un piatto contenente 25 pozzetti permetterà 25 tubuli da analizzare.
- Utilizzare un pennarello indelebile per segnare la posizione dei pozzi, se necessario.
- Effettuare le pareti del pozzo più agevole possibile posizionando il bisturi in elastomero, e quindi utilizzando la mano opposta per ruotare il piatto 360 °.
- Quindi, utilizzando standard di sha RPS precauzioni, immergere un ago G 30 in elastomero non indurito dal passaggio 1.4 e mettere una piccola goccia in fondo a ciascun pozzetto. Questo leviga il fondo del pozzo. Lasciare per curare (indurimento) x 24-48 ore.
- Preparare i perni Minutien.
- Mettere 0,15 mm anodizzato nero Minutien perni in una riga su un pezzo di laboratorio nastro standard di etichettatura 1 pollice. Asse lungo i perni "dovrebbe essere ortogonale all'asse lungo del nastro. Tagliare il nastro lungo la sua lunghezza per recidere ogni pin in due metà approssimativamente uguali (Figura 3). Utilizzare un mezzo-pin per ogni bene.

Figura 3. Taglio Pins Minutien. I perni sono allineati su un pezzo di nastro di etichettatura in parallelo. Poi, forbici sono usati per tagliare i perni a metà.ge.jpg "target =" _ blank "> Clicca qui per vedere una versione più grande di questa figura.
- Inserire ogni semiperno in elastomero di silicone di circa 1 mm a destra di ogni bene (se di mano destra, se mancini, inserire il perno a fianco di ogni bene). Questo è fatto più facilmente quando la visualizzazione dei pozzetti a bassa potenza allo stereomicroscopio dissezione ed è aiutata da uso di una pinza smussato. Figura 2 illustra il posizionamento dei perni.
2. Preparazione Belle Glass Rods
Nota: In questo passaggio, una bacchetta di vetro viene preparato che verrà utilizzato per trasferire i tubuli dal piatto dissezione nella goccia bagno. La bacchetta di vetro viene riutilizzata da esperimento a esperimento, quindi questo passaggio viene eseguito solo una volta, a meno è necessario le pause dell'asta e uno nuovo.
- Ottenere fogli di 3 mm (1/8 di pollice) vetro nero spesso macchiato da un negozio di hobby vetro adeguato-taglio attrezzature, quali un tagliavetro e pinze. Utilizzare i dispositivi di sicurezza adeguate (guanti spessi, occhiali).
- Tagliare il vetro in strisce lunghe ~ 6 mm di larghezza x 10 cm
- Tenere un striscia di vetro in ogni mano. Ammorbidire il breve termine di ogni striscia sulla fiamma di un becco Bunsen, utilizzando apposite misure di sicurezza. Quindi, spingere le estremità delle due strisce insieme e separare in un movimento fluido per creare una bacchetta di vetro fine con una maniglia.
3. Fisiologia Setup
Nota: In questo passaggio, il microscopio, circuiti elettrometro ed elettrico è realizzato. Altro che periodica-chloriding (punto 3.2) dei fili d'argento e ricalibrazione dell'elettrometro (passo 3.8), questa operazione viene eseguita solo una volta. La Figura 4 illustra il programma di installazione.

Figura 4. Fisiologia Setup. Il setup fisiologia è raffigurata qui. (A) Presentazione di setup. Lo stereomicroscopio è collocato all'interno della gabbia di Faraday con micromanipolatori su entrambi i lati. Una luce a fibre ottiche viene infilata attraverso un foro nel lato della gabbia di Faraday. Elettrometro è posto all'esterno della gabbia di Faraday. (B) al cloruro di rivestire i fili d'argento, il filo viene immerso in candeggina. (C) Primo piano del setup. Il titolare microelettrodo diritta, mostrato in questa immagine sulla sinistra, è avvitato sonda del elettrometro. L'elettrodo specifica di litio verrà infilato il filo d'argento nel supporto elettrodi. A destra, l'elettrodo di riferimento viene infilato il filo d'argento del supporto 45 ° microelettrodo. Il circuito deve quindi essere opportunamente collegato a terra. Il piatto test viene mostrato come sarà posizionato nella misurazione. Favoreclicca qui per vedere una versione più grande di questa figura.
{kind=link}
- Posizionare il stereomicrosope con micrometro oculare all'interno della gabbia di Faraday. Terra all'interno della gabbia di Faraday, che viene poi collegato alla terra del telaio del elettrometro. Posizionare i micromanipolatori su entrambi i lati del microscopio (Figura 4A).
- Chloride due fili d'argento per immergersi in candeggina per almeno 1 ora. Estensione overnight (O / N) se necessario (Figura 4B). Ripetere questo passaggio quando i fili d'argento devono essere ri-clorurata, ad esempio se sono in aspetto grigio anziché nero.
- Discussione uno clorurata filo d'argento in ciascuno dei titolari di microelettrodi.
- Stabilire il circuito elettrico con adeguata messa a terra. Ad esempio, posizionare il ventilato, titolare microelettrodo dritto, che terrà l'elettrodo specifici ioni di litio (ISE), sulla sonda elettrometro, che è fissato sul micromanipolatore (Figura 4C).
- Fissare il ventilato, 45 ° titolare microelettrodi, che terrà l'elettrodo di riferimento, sull'altro micromanipolatore (Figura 4C). Poi massa alla massa del circuito sul elettrometro.
- A terra la "AB fuori" uscita BNC dell'elettrometro alla massa del telaio dell 'elettrometro.
- Posizionare la sorgente di luce a fibre ottiche di fuori della gabbia di Faraday, con il tubo a collo d'oca filettato attraverso un foro nella gabbia di Faraday (Figura 4A).
- Configurare e calibrare l'elettrometro secondo le istruzioni del produttore. Ricalibrare l'elettrometro regolarmente (ogni 1 - 2 settimane). Una volta completata, e tra le misure, lasciare l'elettrometro in modalità "standby" con l'interruttore di posizione impostata su "IN", impostare il metodo di metro "A" e la gamma impostata su "200 mV".
4. Preparare le soluzioni di dissezione e Bath
- Preparare Drosophila salina come dettagliato nella tabella 1. Per l'uso negli esperimenti, versare 40 ml ~ in un tubo conico da 50 ml e mantenerlo a RT. Eliminare se vi è evidenza di crescita batterica o fungina.
- Per preparare il bagno di media standard (SBM), mescolare Drosophila salina 1: 1 con la media di Schneider, e passare attraverso un filtro siringa da 0,22 micron. Preparare in piccole aliquote (~ 10 - 15 ml), conservare a 4 ° C e scartare se vi è evidenza di crescita batterica o fungina. I componenti del mezzo di Schneider sono elencati nella Tabella 2.
5. Fare il dell'elettrodo ionico specifico: silanizzante Pipette
Nota: In questo passaggio, diclorodimetilsilano viene utilizzato per leggerezza "silanizzare" l'elettrodo specifico di litio. Questo aggiunge un cappotto idrofoba all'interno dell'elettrodo che permette di conservare lo ionoforo idrofoba. Silanizzazione eccessivo è evitato per prevenire l'assorbimento di olio minerale quando si effettua measurements in gocce sott'olio. Elettrodi silanizzata sono buoni per diverse settimane. Pertanto, questo passaggio viene eseguito ogni poche settimane.
- Fiamma-polacco le estremità di 5 - 6 tubi capillari unfilamented vetro borosilicato (diametro esterno 1,2 mm, diametro interno 0,69 millimetri, lunghezza 10 cm) su un fuoco lento, utilizzando appropriate misure di sicurezza.
- Posizionare i tubi capillari nel fondo di un bicchiere di vetro 1 L.
- In un cappuccio e con dispositivi di protezione adeguati, versare l'acido nitrico 70% (ATTENZIONE: infiammabile e corrosivo, vedi scheda di sicurezza per lo stoccaggio sicuro e di movimentazione) sui tubi capillari e immergere per 5 minuti.
- Versare l'acido nitrico indietro in una bottiglia di vetro. Riutilizzo di successivo lavaggio acido nitrico.
- Aggiungi ~ 200 ml di deionizzata H 2 O al bicchiere. Rifiuti vuoto in una bottiglia di vetro dedicato per i rifiuti acido nitrico. Ripetere con altri 200 ml di H 2 O. deionizzata Seguire le linee guida istituzionali per the smaltimento sicuro dei rifiuti acidi.
- Fare tre lavaggi supplementari con grandi volumi di acqua deionizzata. Svuotare in lavandino.
- Nella cappa, posizionare tubi capillari su una piastra calda con ceramica set top a 200 ° C e asciugare per un minimo di 20 min, in modo ottimale 1 hr. Può anche essere lasciato per lunghi periodi di tempo.
- Su un estrattore pipetta (Figura 5A), tirare pipette ad un diametro punta di ~ 1-2 micron.
- Posizionare pipette tirati indietro sulla piastra, facendo attenzione a non rompere suggerimenti, per almeno 10 minuti, in modo ottimale 30 minuti, ma può essere lasciato a lungo.
- Aggiungere 20 ml di diclorodimetilsilano (ATTENZIONE: infiammabile, corrosivo, tossicità acuta, vedi scheda di sicurezza per lo stoccaggio sicuro e di movimentazione) in una capsula di Petri in vetro da 15 cm ed invertire piatto sopra le pipette sulla piastra calda (Figura 5B). Lasciare in posa per almeno 20 minuti, in modo ottimale 2 ore. La stessa capsula di Petri può essere riutilizzato in esperimenti successivi.
- Determinare il amou nt di diclorodimetilsilano aggiunto per tentativi ed errori. Dopo ionoforo viene aggiunto (passo 8.4), assicurare che l'interfaccia tra lo ionoforo e la soluzione di riempimento è piatta (Figura 5C). Se l'interfaccia è concava, questo indica over-silanizzazione, e meno silano deve essere utilizzato. Se l'interfaccia è convessa, questo indica sotto-silanizzazione, e più silano deve essere utilizzato.
Nota: Il diclorodimetilsilano tende ad andare "off" nel tempo, vale a dire, la silanizzazione meno efficace si ottiene con lo stesso volume di silano.. A questo punto, sia nuovo silano può essere ordinato, o il valore rettificato per raggiungere silanizzazione equivalente.
- Determinare il amou nt di diclorodimetilsilano aggiunto per tentativi ed errori. Dopo ionoforo viene aggiunto (passo 8.4), assicurare che l'interfaccia tra lo ionoforo e la soluzione di riempimento è piatta (Figura 5C). Se l'interfaccia è concava, questo indica over-silanizzazione, e meno silano deve essere utilizzato. Se l'interfaccia è convessa, questo indica sotto-silanizzazione, e più silano deve essere utilizzato.
- Spegnere piastra calda e lasciare raffreddare. Rimuovere capsula di Petri in vetro e trasferire pipette di barattolo di memorizzazione contenente gel di silice, che mantiene essiccazione. Maneggiare con cura pipette (forcipe utile) per evitare la rottura punta.
5 "src =" / files / ftp_upload / 53144 / 53144fig5.jpg "/>
Figura 5. silanizzante Pipette. (A) Esempio di pipetta estrattore. (B) Immagine delle pipette tirato sulla piastra calda. Il piatto di vetro contenente una goccia di diclorodimetilsilano è stata invertita negli pipette tirato. (C), schematicamente l'interfaccia tra il cocktail ionoforo e la soluzione di riempimento. Un'interfaccia bassi indica silanizzazione ottimale. Clicca qui per vedere una versione più grande di questa figura.
{kind=link}
6. Preparare il dispositivo di aspirazione negativa
Nota: In questo passaggio, un semplice dispositivo di aspirazione negativo viene preparato (Figura 6) che verrà utilizzato per riempire l'elettrodo specifico di litio. Questo passo viene eseguito solo una volta.
- Attaccare una siringa da 3 ml di un rubinetto a 3 vie con luer lock. Al'estremità opposta del rubinetto, che contiene il collare e guardia rotante, avvitare un connettore di bloccaggio luer femmina con fine spinato. Poi, collegare il tubo in silicone, diametro interno 1/16 pollici con diametro esterno di 1/8 di pollice, al fine spinato del connettore. Quindi, inserire tubi in plastica con diametro interno 1/32 pollici e diametro esterno 3/32 pollici

Figura 6. Il dispositivo di aspirazione negativa. Immagine di componenti del dispositivo di aspirazione negativo (3 ml con siringa luer lock, rubinetto a 3 vie con collare girevole e protezione, luer femmina Connettore bloccaggio con estremità sagomata, tubo di silicone, tubi in plastica) e il prodotto finale. Clicca qui per vedere una versione più grande di questa figura.
{kind=link}
7. Mosche Raccogliere per la dissezione < / p>
- Utilizzare tecniche di allevamento mosca standard di 25 per impostare croci mosca e modificare, se necessario. Ad esempio, l'uso aumento della temperatura (ad esempio 28 ° C) in esperimenti in cui si desidera una maggiore attività GAL4.
Nota: E 'importante che le mosche non sono allevati in condizioni eccessivamente affollate; il numero di genitori maschi e femmine devono essere diminuiti in questo caso. Se si utilizzano diversi genotipi, il numero dei genitori maschi e femmine devono essere regolati per ottenere il numero di circa simile della progenie. - Raccogliere le mosche per la dissezione (punto 11), utilizzando tecniche di allevamento mosca livello 25 entro 1 - 2 giorni di eclosion. Tubuli da mosche femminili sono più facilmente sezionati, ma tubuli da linea maschile può anche essere utilizzati, se necessario o desiderato. Inserire vola in un flacone contenente mosca cibo. Posizionare flaconi alla temperatura desiderata per 3 - 5 giorni prima della dissezione.
8. Riempimento dell'elettrodo ionico specifico (ISE)
e_content "> Nota:.. In questa fase, l'ISE è riempito con una soluzione salina e poi ionoforo viene introdotto nella punta La ISE può essere riutilizzato di giorno in giorno fino a quando si sta lavorando bene Pertanto, questo passaggio è eseguito ogni pochi giorni, se necessario.- Per fare un elettrodo -specific K +, backfill una pipetta silanizzata con 0,5 M KCl con una siringa da 1 ml e microfilamenti (riutilizzo microfilamenti da esperimento per esperimento). Assicurarsi che la soluzione di riempimento riempie fino alla punta della pipetta - senza aria in punta. In caso di dubbi, visualizzare al microscopio composto. Rimuovere le bolle d'aria muovendo delicatamente la pipetta.
- Per un elettrodo -specific Na +, recupero informazioni con NaCl 150 mm.
- Inserire l'estremità posteriore della ISE nel tubo di plastica del dispositivo di aspirazione negativo preparato al punto 6. Nella cappa, posizionare l'ISE su una plastica cm Petri invertita 3.5 con un pezzo di plastilina per fissarlo in posizione.
- Genera p negativoressione utilizzando il dispositivo di aspirazione. Arretrare sulla siringa con il "fuori" del manico rubinetto rivolto verso la porta laterale. La quantità di arretramento varierà ma è solitamente nell'intervallo di 0,6 - 0,7 ml. Poi girare la maniglia rubinetto in modo che il "off" è rivolta verso il tubo.
- Nel cofano e con dispositivi di protezione adeguati, immergere un 1 - microlitri pipetta punta nella soluzione ionoforo 10 (ATTENZIONE: tossico Vedi scheda di dati di sicurezza per lo stoccaggio sicuro e di movimentazione.). Espellere una piccola goccia mettendo un dito guantato sulla grande apertura del puntale. Quindi, toccare la goccia di ionoforo alla punta della ISE, senza toccare la punta ISE con la punta della pipetta per evitare di rompere la punta ISE.
- Utilizzare il ionoforo potassio elencato nella tabella dei materiali "come sono". Per preparare il ionoforo sodio portare una soluzione di (in% w / w) 10% 4- ter -butylcalix [4] acido tetraethylester arene-tetraacetico, 890,75% nitrofenile ottil etere, e lo 0,25% di sodio borato tetrafenile (ATTENZIONE:. Tossico Vedi Scheda di dati di sicurezza per lo stoccaggio sicuro e di movimentazione). Conservare in un flaconcino di vetro avvolto in carta stagnola per schermare dalla luce.
- Esaminare il ISE sotto 40X con lo zoom utilizzando un microscopio composto per determinare se il ionoforo era "ripreso" in punta ISE e se l'interfaccia soluzione ionoforo / recupero è piatta (vedi punto 5.10.1 e Figura 5C).
- Se non ionoforo è stato ripreso, aumentare la quantità di pressione negativa generata dal dispositivo di aspirazione. Se questo non è successo, l'elettrodo può essere stato adeguatamente silanizzata. Ripetere il punto 5 usando un quantitativo di diclorodimetilsilano.
- Posizionare la ISE, punta verso il basso, sulla parete di un becher parzialmente riempito con KCl 150 mM. Fissare la ISE posizionando plastilina sul lato del bicchiere. La punta si trova all'interno mM KCl 150. Continuare a utilizzare il ov ISEer più giorni fino a quando si sta lavorando bene (vedi punto 10.6).
- Per un Na + ISE, conservare l'elettrodo in NaCl 150 mM.
9. Preparare l'elettrodo di riferimento
Nota: i passaggi 9,1-9,3 può essere eseguita in anticipo. I passaggi 9,4-9,6 vengono eseguite ogni giorno sperimentale.
- Fiamma-polacco le estremità di 10 tubi capillari filamented vetro borosilicato (diametro esterno 1,2 mm, diametro interno 0,69 millimetri, lunghezza 10 cm) a fuoco lento, con le precauzioni di sicurezza.
- Su un estrattore pipetta, tirare pipette ad un diametro punta di ~ 1-2 micron.
- Conservare pipette in un vaso di archiviazione fino al momento dell'uso. Pipetta Pipette possono essere conservati a tempo indeterminato.
- Il giorno dell'esperimento, utilizzando un microfilamenti e la siringa, riempire la punta e gambo della pipetta con acetato di sodio 1 M. Assicurarsi che ci siano bolle d'aria e che la soluzione va alla punta. Flick delicatamente la pipetta se sono presenti bolle d'aria.
- Utilizzo di un secondomicrofilamenti e la siringa, riempimento la pipetta con 3 M KCl. Ancora una volta, assicurano bolle d'aria non sono presenti.
- Conservare l'elettrodo di riferimento in un bicchiere contenente 150 mM KCl (vedi punto 8.6).
10. Taratura di ISE
Nota: Questa fase viene eseguita tre volte il giorno esperimento: nelle prime ore del giorno per assicurarsi che il ISE è in funzione, e quindi prima e dopo misurazioni dei 20 - 25 gocce liquido secreto (Tabella 3).
- Per tarare il potassio ISE, posizionare due 0.6 ml gocce ciascuno dei seguenti quattro concentrazioni di KCl sul rivestito-elastomero 3,5 centimetri Petri silicone (preparata al punto 1): 15 mm, 75 mm, 150 mm e 200 mm. Strato cautela 2 ml di olio minerale sulle gocce.
- Per sodio ISE, utilizzare gocce di taratura di 15 mm e 150 mM NaCl.
- Porre la capsula di calibrazione sul palco dello stereomicroscopio nella gabbia di Faraday e illuminare.
- Threannuncio l'elettrodo ISE e di riferimento sui fili d'argento e fissare in supporti di microelettrodi.
- Utilizzando i micromanipolatori, avanzare l'ISE e elettrodi di riferimento in calo del 15 mM KCl.
- Passare l'elettrometro a "operare" modalità. Consentire la lettura per risolvere.
- Lettura registrare nel notebook. Ripetere con il 75 mm, mm 150 mm e 200 gocce. Calcola pendenza per determinare se ISE sta funzionando bene (vedi punto 13.1 e Tabella 3). In caso contrario, preparare una nuova ISE.
Nota: I segni che l'ISE non funziona bene: fallimento per ottenere una lettura; lento equilibrare (alcuni secondi o più); lettura instabile; Pendenza <49 mV cambiamento / decile della concentrazione K + o Na +. - Tra la prima taratura del giorno e le misure effettuate nel passaggio 12, cioè durante l'esecuzione del passo 11 (dissezioni tubulari), memorizzare l'ISE e elettrodi di riferimento in 150 mM KCl (come descritto al punto 8.6).
11. Tubulo Dissection
Nota: Questo passo viene eseguita il giorno dell'esperimento.
- Aliquotare una piccola quantità (~ 500 - 600 ml) di mezzo di balneazione standard (SBM), preparato al punto 4.2, per uso nel giorno dell'esperimento e lasciarla riscaldare a RT. Questo dovrebbe essere fatto almeno 30 minuti prima dissezioni, ma può anche essere fatto in precedenza. Inoltre, avere almeno 20 ml di soluzione fisiologica RT Drosophila (passo 4.1) disponibile prima di iniziare dissezioni.
- Immediatamente prima di iniziare dissezioni: visualizzare il piatto test a 10 ingrandimenti allo stereomicroscopio e aggiungere abbastanza SBM per riempire quasi ogni bene nel piatto test, di solito tra 10 e 30 ml. Se i farmaci o peptide stanno per essere aggiunto alla SBM metà esperimento, prendere nota del volume esatto di SBM aggiunti a ciascun pozzetto. Evitare un eccesso di riempire il pozzo come questo può portare alla tubulo galleggiante via durante l'esperimento.
- Strato attentamente ~ 12 - 13 ml di olio minerale sulla parte superiore in modo che i pozzetti unri coperto. Ciò impedirà evaporazione dei secreti gocce di fluido durante l'esperimento.
- Posto vola a essere sezionato su CO 2 pad.
- Prendete una mosca attraverso la sua gamba o ala con una pinza e posto sul dorso (ventrale rivolto verso l'alto) sul piatto dissezione elastomero siliconico rivestite preparata nella fase 1. Al palo il torace con un perno Minutien per fissare il volo a posto.
- Aggiungere una goccia di RT Drosophila salina (passo 4.1) per immergere la mosca in soluzione salina.
- Opzionale: clip fuori le ali e le gambe. In pratica, questo non è solitamente necessario.
- Utilizzare le pinze mano non dominante a "hold" addome della mosca alla giunzione toraco-addominale. Utilizzare le pinze mano dominante a sbucciare la cuticola addominale via, a partire dalla giunzione toraco-addominale e si muove verso la fine della coda della mosca. L'intestino, con tubuli malpighiani allegati, deve essere esposto con questa manovra.
- Senza toccare i tubuli, sezionare gratuito midgut /hindgut e tubuli collegati. Mantenere l'intestino nelle pinze mano non dominante e utilizzare un ago da 30 G per tagliare l'uretere dall'intestino, staccando i tubuli dall'intestino e libero dalla fly.It è essenziale che né lacrime né rendite essere introdotti nel tubulo, eccettuato dell'uretere.
Nota: La coppia anteriore tubuli è più facilmente sezionato, tuttavia i tubuli posteriori possono essere usati pure. - Usando la bacchetta di vetro fine (fase 2), prendere la coppia tubulo e trasferire in un pozzo del piatto test.
- Subito dopo la coppia tubulo è stato trasferito nel pozzo, prendere l'estremità di uno dei tubuli con la bacchetta di vetro, ritirarsi dalla goccia bagno fino alla fine taglio dell'uretere è a metà strada tra il perno e la goccia bagno e avvolgere Alla fine del tubulo intorno al perno con la bacchetta di vetro. Alla fine di questa manovra, uno tubulo rimane nella goccia salina balneazione e secernono fluido dall'estremità tagliata dell'uretere, come illustrato in figura 1 . L'altra tubulo, etichettato il "ancoraggio tubulo" in Figura 1, è avvolto intorno al perno. E 'ancora il tubulo secernente a posto, è circondato da olio, e non secernono liquido.
- Subito dopo il punto 11.11, scrivere il bene (ad esempio, A, B, C), informazioni tubulo identificazione (ad es., Il genotipo o condizione), e il tempo (questa è l'ora di inizio quando il fluido inizierà ad essere secreto dal tubulo in gocciolina di soluzione salina piattaforma).
- Procedere con il successivo dissezione. Una volta che lo sperimentatore è esperto in questa tecnica, che in genere dura 3 - 4 min di sezionare un paio di tubuli, trasferirli al bagno salino, e avvolgere il tubulo ancoraggio intorno al perno. Pertanto, 20 - 25 tubuli possono essere impostati nel dosaggio Ramsay entro 1,5 ore. L'ora di inizio di ogni tubulo sarà quindi di circa 3-4 minuti dopo l'orario di inizio del tubulo precedente.
12. Esecuzione delle misure
Nota: Questa fase viene eseguita il giorno dell'esperimento.
- Calibrare la ISE (punto 10) circa 20 minuti prima della prima misurazione. Questo permette di tempo per fare un nuovo ISE se necessario.
Nota: Al momento desiderato, per esempio dopo 2 ore della secrezione, la goccia di liquido secreto ogni tubulo è pronto per la misura. - Registrare il tempo di misura. Misurare il diametro della goccia liquido secreto con il micrometro oculare e registrare. Si noti l'ingrandimento, per esempio, 50X.
- Far avanzare il ISE ed elettrodo di riferimento nella goccia di fluido. Passare l'elettrometro a "operare". Attendere che la lettura si stabilizzi. Registrare il valore.
- Ripetere l'operazione per la prossima goccia.
- Alla fine dell'esperimento, ripetere le misurazioni di calibrazione (passo 10).
13. Calcoli
Nota: Questa fase può essere eseguita al termine della giornata esperimento, o in un secondo momento.
- Calcolare la mediapista / cambio decile della concentrazione di potassio. Vedere la Tabella 3 per un esempio.
Nota: Per sodio, i valori saranno per la differenza tra il 15 mM e 150 mM NaCl misurazioni. - Determinare il valore medio delle due misure (prima e dopo) di KCl 200 mM (o 150 mM NaCl).
- Calcolare il volume di ciascuna gocciolina. V = πd 3/6, dove d è il diametro della goccia misurata con il micrometro oculare nel passo 12.2.
- Calcolare il tasso di secrezione = V / ora (nL / min / tubulo), dove V è il volume della goccia determinato passo 13.3, e il tempo è il tempo in cui la liquido secreto tubulo (= tempo di misurazione - tempo di tirare dell'uretere fuori delle gocce di balneazione).
- Calcolare la concentrazione di ioni utilizzando la formula [K] = 10e (DV / S) * 200 o [Na] = 10e (DV / S) * 150, dove Δv = la differenza (in mV) tra il potenziale misurata del fluido secreto goccia, e il potenziale della goccia calibrazione 200 mM (fo di potassio; 150 goccia mM sodio). S = la pendenza determinata al punto 13.1.
- Calcola ion flusso = [ione] x tasso secrezione di fluidi. Per tubuli Drosophila, questo sarà pmol / min / tubulo.
14. Pulizia Up
Nota: Questa fase viene eseguita alla fine della giornata dell'esperimento.
- Pulizia accurata dei pozzetti è essenziale per garantire che i cristalli di sale residui non rimangono nei pozzetti, alterando la concentrazione di ioni e osmolarità in esperimenti futuri.
- Lasciare che l'olio minerale a defluire.
- Lavare i pozzetti del piatto test. Una punta della pipetta 200 microlitri può essere utilizzato per raschiare delicatamente cristallo di sale residuo. Utilizzando tubi di plastica attaccato al rubinetto, stringere il tubo per creare un getto ad alta pressione di acqua calda per sciacquare abbondantemente fuori ogni bene.
- Lasciare asciugare O / N. In alternativa, un asciugacapelli può essere utilizzato per asciugare i pozzetti.
- Risciacquare con pinze deionizzata H 2 O eimmergere in etanolo per 15 minuti a diverse ore.
- Scaricare l'olio fuori di piatto calibrazione e lavare con acqua calda. Usare sapone così finché è accuratamente risciacquato. Eseguire l'ultimo risciacquo con acqua distillata H 2 O.
- Microfilamenti Lavare e siringhe con H 2 O. distillata
Risultati
Le figure 7 e 8 dimostrano che l'uso del saggio Ramsay con elettrodi specifici ioni per misurare concentrazioni K + e Na + possono distinguere geneticamente e farmacologicamente distinto K + e flussi Na +, informazioni che non sono catturati misurando i tassi di secrezione di fluido da solo. Figura 7 mostra che diminuiscono la secrezione di fluidi nei tubuli da mosche trasportano un null mutazione omozigotica nel NKCC è guidato da una dim...
Discussione
L'uso del saggio Ramsay, insieme con elettrodi ione-specifico, permette la misura dei tassi di secrezione di fluido e flussi ionici in isolato insetti malpighiani (renali) tubuli. Venti o più tubuli possono essere dosati in un momento, consentendo un throughput più elevato rispetto al dosaggio dei singoli tubuli in vitro microperfused. Inoltre, gli elettrodi ione-specifico consentono la determinazione delle concentrazioni di ioni all'interno del fluido secreto in situ, limitando errori che po...
Divulgazioni
The authors have nothing to disclose.
Riconoscimenti
The authors wish to thank Drs. Sung-wan An and Mike O’Donnell for practical advice on establishing this assay, Dr. Chih-Jen Cheng for helpful discussions on the use of ion-specific electrodes, and Dr. Chou-Long Huang for his mentorship and support. This work was supported by the National Institutes of Health (K08DK091316 to ARR) and the American Society of Nephrology Gottschalk Award to ARR.
Materiali
| Name | Company | Catalog Number | Comments |
| Sylgard 184 Silicone Elastomer Kit | Ellsworth Adhesives | http://www.ellsworth.com/dow-corning-sylgard-184-silicone-encapsulant-0-5kg-kit-clear/ | May be purchased from multiple distributors |
| Petri dish, polystyrene, 100 mm x 15 mm | Fisher | FB0875712 | Specific brand is not important |
| Petri dish, polystyrene, 35 mm x 10 mm | Corning Life Sciences | Fisher 08-757-100A | Specific brand is not important |
| Scalpel Handle #3 | Fine Science Tools | 10003-12 | Specific brand is not important |
| Scalpel Blades #1 | Fine Science Tools | 10011-00 | Specific brand is not important; use appropriate sharps precautions |
| Needle, 30 G x 1/2 | Becton Dickinson | 305106 | Use appropriate sharps precautions |
| Minutien pins, black anodized, 0.15 mm | Fine Science Tools | 26002-15 | |
| Stereomicroscope with ocular micrometer | Nikon | SMZ800 | Specific brand is not important; this is given as an example |
| Sheet of black stained glass, 3 mm (1/8 inch) thick | Hobby shop | Example includes Spectrum Black Opal by Spectrum Glass (http://www.delphiglass.com/spectrum-glass/opalescent/spectrum-black-opal) | |
| Glass cutting tools (glass cutter, glass cutting pliers) | Hobby shop | Examples include the Studio Pro Lightweight Running Pliers by Diamond Tech (http://www.delphiglass.com/glass-cutters-tools/pliers-nippers/studio-pro-lightweight-running-pliers) and the Studio Pro Brass Glass Cutter by Diamond Tech (http://www.delphiglass.com/glass-cutters-tools/glass-cutters/studio-pro-brass-glass-cutter). Use appropriate safety precautions when cutting glass | |
| Borosilicate glass capillary tube, unfilamented, GC120-10, OD 1.2 mm, ID 0.69 mm, length 10 cm | Warner Instruments | 30-0042 | |
| Borosilicate glass capillary tube, filamented, GC120F-10, OD 1.2 mm, ID 0.69 mm, length 10 cm | Warner Instruments | 30-0044 | |
| Nitric acid, 70% | Sigma | 438073 | CAUTION: see Material Data Safety Sheet for appropriate storage and handling guidelines. Specific brand is not important |
| Cimarec 7 in x 7 in hotplate | Fisher | 11675911Q | Specific brand is not important; caution when heated |
| Selectophore dichlorodimethylsilane | Sigma | 40136-1ML | CAUTION: see Material Data Safety Sheet for appropriate storage and handling guidelines |
| Two-step vertical pipet puller | Narishige | PC-10 | Other pipet pullers can be used; this is given as an example |
| Glass petri dish, 150 mm diameter x 15 mm height | Fisher | 08-748E | Specific brand is not important; only one dish needed |
| World Precision Instruments E210 1 mm micropipette storage jar | Fisher | 50-821-852 | May be available from other distributors. Useful to have two jars. Note that although this jar is specified for 1 mm pipets, and the pipets used here are 1.2 mm, in our experience the 1 mm jar works best for the 1.2 mm pipets. |
| Silica Gel, Tel-Tale Desiccant, indicating, 10-18 mesh | Fisher | S161-500 | Indicating silica useful for determining whether silica gel retains desiccating ability |
| World Precision Instruments MicroFil, 34G | Fisher | 50-821-914 | May be available from other distributors. |
| 1 ml syringe with luer lock | Becton Dickinson | 309659 | May be available from other distributors. |
| 3 ml syringe with luer lock | Becton Dickinson | 309657 | May be available from other distributors. |
| D300 3-way stopcock with female luer lock inlet port, male luer outlet port with rotating collar and guard | Cole-Parmer | UX-30600-02 | Specific brand is not important |
| Female Luer Locking Connector | 4 Medical Solutions | ADC 9873-10 | Specific brand is not important; barbed end is ~4 mm at narrowest point and ~7 mm at widest point. |
| Silicone Tubing I.D. x O.D. x Wall: 1/16 x 1/8 x 1/32 in. (1.59 x 3.18 x 0.79 mm) | Fisher | 14-179-110 | Specific brand is not important |
| E-3603 tubing, I.D. x O.D.: 1/32 x 3/32 in | Fisher | 14171208 | Specific brand is not important |
| Modeling clay | Specific brand is not important | ||
| Selectophore potassium ionophore I, cocktail B | Sigma | 99373 | CAUTION: see Material Data Safety Sheet for appropriate storage and handling guidelines |
| Selectophore sodium ionophore X | Sigma | 71747 | Sodium ionophore X = 4-tert-butylcalix[4]arene-tetraacetic acid tetraethylester |
| Selectophore 2-nitrophenyl octyl ether | Sigma | 73732 | |
| Selectophore sodium tetraphenylborate | Sigma | 72018 | CAUTION: see Material Data Safety Sheet for appropriate storage and handling guidelines |
| Schneider's Drosophila medium | Life Technologies | 21720024 | |
| High impedance electrometer | World Precision Instruments | FD223a | |
| Microelectrode holder 1 mm with 45° body, vented, with handle | Warner Instruments | 64-1051 | |
| Microelectrode holder 1 mm with straight body, vented | Warner Instruments | 64-1007 | |
| Silver wire | Warner Instruments | 64-1318 | |
| Micromanipulators, pair | Leitz | Various brands/models will work; this is an example | |
| Faraday cage | Technical Manufacturing Corporation | 81-334-03 | This is an example; any Faraday cage will work |
| Single gooseneck fiberoptic light | Nikon | Specific brand is not important | |
| mineral oil | Fisher | BP-2629 | Specific brand is not important |
| forceps, Dumont #5 with Biologie tip | Fine Science Tool | 11295-10 | May be available from other distributors. |
Riferimenti
- Ramsay, J. A. Active Transport of Water by the Malpighian Tubules of the Stick Insect, Dixippus-Morosus (Orthoptera, Phasmidae). J Exp Biol. 31, 104-113 (1954).
- Dow, J. A., et al. The malpighian tubules of Drosophila melanogaster: a novel phenotype for studies of fluid secretion and its control. J Exp Biol. 197, 421-428 (1994).
- Brand, A. H., Perrimon, N. Targeted gene expression as a means of altering cell fates and generating dominant phenotypes. Development. 118, 401-415 (1993).
- Sozen, M. A., Armstrong, J. D., Yang, M., Kaiser, K., Dow, J. A. Functional domains are specified to single-cell resolution in a Drosophila epithelium. Proc Natl Acad Sci U S A. 94, 5207-5212 (1997).
- Rosay, P., et al. Cell-type specific calcium signalling in a Drosophila epithelium. J Cell Sci. 110 (15), 1683-1692 (1997).
- Dow, J. T., Davies, S. A. Integrative physiology and functional genomics of epithelial function in a genetic model organism. Physiol Rev. 83, 687-729 (2003).
- Beyenbach, K. W., Skaer, H., Dow, J. A. The developmental, molecular, and transport biology of Malpighian tubules. Annu Rev Entomol. 55, 351-374 (2010).
- Donnell, M. J., et al. Hormonally controlled chloride movement across Drosophila tubules is via ion channels in stellate cells. Am J Physiol. 274, 1039-1049 (1998).
- Linton, S. M., O'Donnell, M. J. Contributions of K+:Cl- cotransport and Na+/K+-ATPase to basolateral ion transport in malpighian tubules of Drosophila melanogaster. J Exp Biol. 202, 1561-1570 (1999).
- Rheault, M. R., O'Donnell, M. J. Analysis of epithelial K(+) transport in Malpighian tubules of Drosophila melanogaster: evidence for spatial and temporal heterogeneity. J Exp Biol. 204, 2289-2299 (2001).
- Donnell, M. J., Dow, J. A., Huesmann, G. R., Tublitz, N. J., Maddrell, S. H. Separate control of anion and cation transport in malpighian tubules of Drosophila Melanogaster. J Exp Biol. 199, 1163-1175 (1996).
- Cabrero, P., et al. Chloride channels in stellate cells are essential for uniquely high secretion rates in neuropeptide-stimulated Drosophila diuresis. Proc Natl Acad Sci U S A. 111, 14301-14306 (2014).
- Torrie, L. S., et al. Resolution of the insect ouabain paradox. Proc Natl Acad Sci U S A. 101, 13689-13693 (2004).
- Rodan, A. R., Baum, M., Huang, C. L. The Drosophila NKCC Ncc69 is required for normal renal tubule function. Am J Physiol Cell Physiol. 303, 883-894 (2012).
- Ianowski, J. P., Christensen, R. J., O'Donnell, M. J. Na+ competes with K+ in bumetanide-sensitive transport by Malpighian tubules of Rhodnius prolixus. J Exp Biol. 207, 3707-3716 (2004).
- Naikkhwah, W., O'Donnell, M. J. Salt stress alters fluid and ion transport by Malpighian tubules of Drosophila melanogaster: evidence for phenotypic plasticity. J Exp Biol. 214, 3443-3454 (2011).
- Donini, A., et al. Secretion of water and ions by malpighian tubules of larval mosquitoes: effects of diuretic factors, second messengers, and salinity. Physiol Biochem Zool. 79, 645-655 (2006).
- Maddrell, S. H. Secretion by Malpighian Tubules of Rhodnius movements of Ions and Water. J Exp Biol. 51, 71-97 (1969).
- Maddrell, S. H., Overton, J. A. Stimulation of sodium transport and fluid secretion by ouabain in an insect malpighian tubule. J Exp Biol. 137, 265-276 (1988).
- Williams, J. C., Beyenbach, K. W. Differential effects of secretagogues on Na and K secretion in the Malpighian tubules of Aedes Aegypti (L). J Comp Physiol. 149, 511-517 (1983).
- Maddrell, S. H., O'Donnell, M. J., Caffrey, R. The regulation of haemolymph potassium activity during initiation and maintenance of diuresis in fed Rhodnius prolixus. J Exp Biol. 177, 273-285 (1993).
- Messerli, M. A., Kurtz, I., Smith, P. J. Characterization of optimized Na+ and Cl- liquid membranes for use with extracellular, self-referencing microelectrodes. Anal Bioanal Chem. 390, 1355-1359 (2008).
- Ianowski, J. P., O'Donnell, M. J. Basolateral ion transport mechanisms during fluid secretion by Drosophila Malpighian tubules: Na+ recycling, Na+:K+:2Cl- cotransport and Cl- conductance. J Exp Biol. 207, 2599-2609 (2004).
- Neufeld, D. S., Leader, J. P. Electrochemical characteristics of ion secretion in malpighian tubules of the New Zealand alpine weta (Hemideina maori). J Insect Physiol. 44, 39-48 (1997).
- Greenspan, R. J. . Fly Pushing: The Theory and Practice of Drosophila Genetics. , (1997).
- Jayakannan, M., Babourina, O., Rengel, Z. Improved measurements of Na+ fluxes in plants using calixarene-based microelectrodes. J Plant Physiol. 168, 1045-1051 (2011).
- Wu, Y., Schellinger, J. N., Huang, C. L., Rodan, A. R. Hypotonicity Stimulates Potassium Flux through the WNK-SPAK/OSR1 Kinase Cascade and the Ncc69 Sodium-Potassium-2-Chloride Cotransporter in the Drosophila Renal Tubule. J Biol Chem. 289, 26131-26142 (2014).
- Blumenthal, E. M. Modulation of tyramine signaling by osmolality in an insect secretory epithelium. Am J Physiol Cell Physiol. 289, 1261-1267 (2005).
- Dow, J. A., Maddrell, S. H., Davies, S. A., Skaer, N. J., Kaiser, K. A novel role for the nitric oxide-cGMP signaling pathway: the control of epithelial function in Drosophila. Am J Physiol. 266, 1716-1719 (1994).
- Dube, K., McDonald, D. G., O'Donnell, M. J. Calcium transport by isolated anterior and posterior Malpighian tubules of Drosophila melanogaster: roles of sequestration and secretion. J Insect Physiol. 46, 1449-1460 (2000).
- Efetova, M., et al. Separate roles of PKA and EPAC in renal function unraveled by the optogenetic control of cAMP levels in vivo. J Cell Sci. 126, 778-788 (2013).
- Rheault, M. R., O'Donnell, M. J. Organic cation transport by Malpighian tubules of Drosophila melanogaster: application of two novel electrophysiological methods. J Exp Biol. 207, 2173-2184 (2004).
- Donnell, M. J. Too much of a good thing: how insects cope with excess ions or toxins in the diet. J Exp Biol. 212, 363-372 (2009).
- Cheng, C. J., Truong, T., Baum, M., Huang, C. L. Kidney-specific WNK1 inhibits sodium reabsorption in the cortical thick ascending limb. Am J Physiol Renal Physiol. 303, 667-673 (2012).
- Cheng, C. J., Yoon, J., Baum, M., Huang, C. L. STE20/SPS1-related Proline/alanine-rich Kinase (SPAK) is Critical for Sodium Reabsorption in Isolated Perfused Thick Ascending Limb. Am J Physiol Renal Physiol. , (2014).
Ristampe e Autorizzazioni
Richiedi autorizzazione per utilizzare il testo o le figure di questo articolo JoVE
Richiedi AutorizzazioneThis article has been published
Video Coming Soon
Personale delle biblioteche
Copyright © 2025 MyJoVE Corporation. Tutti i diritti riservati