
Method Article
Adattamento degli organoidi gastrointestinali per l'infezione da agenti patogeni e il sequenziamento di singole cellule in condizioni di livello di biosicurezza 3 (BSL-3)
In questo articolo
Riepilogo
Questo protocollo descrive come infettare gli organoidi intestinali umani dal loro lato apicale o basolaterale per caratterizzare le interazioni ospite/patogeno a livello di singola cellula utilizzando la tecnologia di sequenziamento dell'RNA a singola cellula (scRNAseq).
Abstract
Gli organoidi intestinali umani costituiscono il miglior modello cellulare per studiare le infezioni patogene del tratto gastrointestinale. Questi organoidi possono essere derivati da tutte le sezioni del tratto gastrointestinale (gastrico, digiuno, duodeno, ileo, colon, retto) e, al momento della differenziazione, contengono la maggior parte dei tipi di cellule che si trovano naturalmente in ogni singola sezione. Ad esempio, gli organoidi intestinali contengono enterociti che assorbono i nutrienti, cellule secretorie (calice, Paneth e enteroendocrino), cellule staminali e tutti gli intermedi di differenziazione specifici del lignaggio (ad esempio, tipi di cellule precoci o immature). Il più grande vantaggio nell'utilizzo di organoidi derivati dal tratto gastrointestinale per studiare le malattie infettive è la possibilità di identificare con precisione quale tipo di cellula è bersaglio del patogeno enterico e di affrontare se le diverse sezioni del tratto gastrointestinale e i loro specifici tipi di cellule rispondono in modo simile alle sfide dei patogeni. Negli ultimi anni, modelli gastrointestinali, così come organoidi di altri tessuti, sono stati impiegati per studiare il tropismo virale e i meccanismi di patogenesi. Tuttavia, l'utilizzo di tutti i vantaggi dell'utilizzo di organoidi quando si impiegano virus ad alta patogenicità rappresenta una sfida tecnica e richiede rigorose considerazioni di biosicurezza. Inoltre, poiché gli organoidi sono spesso coltivati in tre dimensioni, il lato basolaterale delle cellule è rivolto verso l'esterno dell'organoide mentre il loro lato apicale è rivolto verso l'interno (lume) degli organoidi. Questa organizzazione rappresenta una sfida per i patogeni enterici poiché molte infezioni enteriche iniziano dal lato apicale / luminale delle cellule dopo l'ingestione. Il seguente manoscritto fornirà un protocollo completo per preparare gli organoidi intestinali umani per l'infezione da patogeni enterici considerando il lato dell'infezione (apicale vs basolaterale) per eseguire il sequenziamento dell'RNA a singola cellula per caratterizzare le interazioni ospite / patogeno specifiche del tipo cellulare. Questo metodo descrive in dettaglio la preparazione degli organoidi e le considerazioni necessarie per eseguire questo lavoro in condizioni di contenimento del livello di biosicurezza 3 (BSL-3).
Introduzione
Studiare il tropismo specifico del tipo cellulare e la risposta immunitaria specifica del tipo cellulare ai virus enterici umani è stato storicamente impegnativo a causa della mancanza di modelli cellulari umani primari. Questa limitazione è stata ora parzialmente sradicata con lo sviluppo di organoidi1. Nel caso del tratto gastrointestinale, sono stati sviluppati modelli organoidi gastrici e intestinali per l'uomo e diverse altre specie (adesempio, murine, bovine, feline, pipistrelli)2,3,4,5,6. Gli organoidi intestinali riproducono l'architettura strutturale dell'epitelio intestinale umano e contengono strutture simili a cripte e villi, lignaggi intestinali funzionali e sono stati persino utilizzati per identificare lignaggi cellulari precedentemente sconosciuti. Due diversi approcci possono essere utilizzati per coltivare organoidi intestinali. In primo luogo, le cellule staminali intestinali contenenti cripte vengono isolate da resezioni tissutali o biopsie e coltivate in condizioni di coltura specifiche (ad esempio, Wnt3A, R-spondin, Noggin ed EGF) per espandersi, e quindi differenziare le cellule staminali dalla maggior parte delle linee cellulari intestinali (adesempio, enterociti, cellule paneth, cellule calici, cellule enteroendocrine)7. Questo metodo consente l'isolamento degli organoidi da tutte le sezioni del tratto gastrointestinale (adesempio,stomaco, duodeno, digiuno, ileo e colon). Il secondo metodo si basa su cellule staminali pluripotenti o embrionali indotte dall'uomo, che vengono poi differenziate in un processo graduale in cellule epiteliali intestinali8. Questi organoidi indotti a base di cellule staminali sono spesso descritti come di natura più embrionale rispetto agli organoidi derivati dal paziente. Mentre tutti questi modelli organoidi sono stati fondamentali per svelare i segnali di sviluppo necessari per formare il tratto intestinale, il loro uso nella ricerca sulle malattie infettive è ancora agli inizi.
Virus enterico è un termine ampio che copre tutti i virus, che infettano attraverso il tratto gastrointestinale, come picornavirus (adesempio,EV-71), reovirus (ad esempio, rotavirus) e calicivirus (ad esempio, norovirus)9. I virus enterici iniziano il loro ciclo di vita infettivo attraverso l'ingestione di cibo e acqua contaminati, che lascia le persone nei paesi in via di sviluppo ad alto rischio a causa dello scarico di rifiuti non trattati nell'ambiente e della mancanza di cure mediche dopo l'insorgenza dell'infezione10. A seconda del tipo di agente patogeno, l'infezione può portare a gastroenterite, vomito e / o diarrea acquosa a causa della perdita del rivestimento intestinale. I norovirus umani sono un patogeno enterico altamente diffuso e altamente infettivo, che porta a oltre 600 milioni di infezioni e 15 milioni di ricoveri in tutto il mondo11. Gli organoidi sono stati fondamentali per la ricerca sul norovirus in quanto supportano l'infezione e la replicazione del norovirus umano, che in precedenza non era in grado di essere coltivato in modelli di coltura cellulare standard12.
Negli ultimi due decenni, i coronavirus sono emersi come patogeni umani chiave13. Questa famiglia comprende i MERS ad alta patogenicità, SARS-CoV-1 e SARS-CoV-2, che richiedono rigorosi livelli di sicurezza quando si eseguono ricerche su questi virus. È interessante notare che, mentre tutti e tre questi agenti patogeni sono per lo più riconosciuti per i loro sintomi respiratori indotti e distress, è ora evidente che questi virus non infettano solo le vie respiratorie ma anche altri organi. Un'importante patologia indotta nei pazienti infetti da SARS-CoV-2 oltre al distress respiratorio è la presenza di sintomi gastrointestinali14. Una frazione dei pazienti infetti da SARS-CoV-2 mostra tali sintomi, che vanno da diarrea molto lieve a grave. Inoltre, i genomi SARS-CoV-2 possono essere rilevati nelle biopsie delle feci e del tratto gastrointestinale di pazienti infetti15. È importante sottolineare che la presenza di sintomi gastrointestinali non è limitata a SARS-CoV-2 come sono stati osservati anche in pazienti con infezione da MERS e SARS-CoV-1. Per capire come SARS-CoV-2 induce disturbi gastrointestinali e identificare con precisione il tropismo di SARS-CoV-2 nel tratto gastrointestinale, gli organoidi intestinali umani sono stati uno strumento chiave e sono ora sfruttati per svelare le risposte specifiche del tipo cellulare a questo patogeno16,17.
Il profilo trascrizionale di una popolazione cellulare (sequenziamento dell'RNA di massa) è stata una pratica standard quando si valutano le infezioni patogene sia di linee cellulari immortalizzate che con organoidi. Mentre questo ci consente di determinare i cambiamenti globali in risposta ai patogeni (ad esempio, l'upregulation delle citochine), l'RNAseq di massa non ci consente di determinare perché cellule specifiche in una popolazione sono più inclini all'infezione rispetto ad altre. Il sequenziamento dell'RNA a singola cellula (scRNAseq) è diventato un potente strumento per svelare i programmi trascrizionali specifici del lignaggio cellulare e può essere utilizzato per determinare come questi programmi supportano o reprimono l'infezione virale18,19. La prima descrizione di scRNAseq risale al 2009 ed è stata utilizzata per valutare i profili di trascrizione delle diverse cellule trovate in un blastomero di topo20. Queste tecnologie sono state ora ampliate e possono essere implementate attraverso diverse piattaforme. Le prime versioni di questa tecnologia applicavano il selezionatore di cellule attivato dalla fluorescenza (FACS) per separare le singole cellule per il sequenziamento, che era spesso limitato a piastre a 96 o 384 pozzetti, dando così 300 singole cellule da analizzare per campione21. Questi metodi sono stati ora avanzati dalle piattaforme di sequenziamento a singola cellula, che utilizzano un dispositivo microfluidico per incapsulare singole cellule in singole goccioline con codice a barre contenente perline. Questa tecnologia consente di acquisire fino a 10.000 celle per condizione del campione.
La combinazione della tecnologia degli organoidi con scRNAseq ci consente di studiare come i patogeni enterici influenzano il tratto gastrointestinale in modo specifico per tipo di cellula. Tuttavia, è necessario prendere diverse considerazioni tecniche e di biosicurezza. Innanzitutto, i metodi classici di coltura degli organoidi (organoidi tridimensionali (3D), incorporati in una matrice extracellulare (ECM)) espongono il lato basolaterale delle cellule epiteliali all'esterno dell'organoide. Poiché i patogeni enterici iniziano la loro infezione attraverso l'ingestione di cibo / acqua contaminati, l'infezione inizia più spesso dal lato apicale delle cellule, che non è accessibile in questi organoidi intestinali 3D. Pertanto, gli organoidi devono essere preparati per rendere il lato apicale accessibile all'infezione da agenti patogeni attraverso la semina 2D, esponendo così direttamente il lato apicale delle cellule, o attraverso la microiniezione22,23. In secondo luogo, per eseguire scRNAseq di campioni biologici infetti, è importante considerare la loro natura infettiva. Mentre sono stati proposti metodi per fissare le cellule e inattivare i patogeni prima dell'isolamento unicellulare per il successivo RNAseq, questi metodi spesso portano ad una diminuzione della qualità del sequenziamento18. Il protocollo seguente descriverà diversi approcci per infettare gli organoidi intestinali con virus enterici considerando il lato dell'infezione (infezione apicale vs basolaterale) (Figura 1). Inoltre, il protocollo includerà un flusso di lavoro per dissociare e isolare singole cellule da organoidi infettati da virus ad alta patogenicità per scRNAseq. Il protocollo evidenzierà i passaggi chiave che devono essere implementati quando si lavora in condizioni di contenimento di livello di biosicurezza 3 (BSL-3) per evitare la generazione di aerosol e la potenziale contaminazione.
Protocollo
Il tessuto umano è stato ricevuto dalla resezione del colon o dalle biopsie dell'ileo dall'Ospedale Universitario di Heidelberg per il seguente protocollo. Questo studio è stato condotto sotto le raccomandazioni dell'Ospedale Universitario di Heidelberg con il consenso scritto informato di tutti i soggetti in conformità con la Dichiarazione di Helsinki. Tutti i campioni sono stati ricevuti e mantenuti in modo anonimo. Il protocollo è stato approvato dalla commissione etica dell'ospedale universitario di Heidelberg ai sensi del protocollo S-443/2017.
1. Mantenimento e passaggio degli organoidi intestinali e del colon
ATTENZIONE: Gli organoidi intestinali umani sono derivati da tessuto umano o da cellule staminali pluripotenti/embrionali indotte e, come tali, è richiesta l'approvazione etica. È necessario seguire le normative specifiche per paese. Il materiale umano non è generalmente testato ed è quindi spesso considerato materiale BSL-2. Adeguate condizioni di contenimento devono essere confermate nel paese in cui si svolge l'esperimento.
- Preparare organoidi intestinali e del colon da tessuti isolati e/o biopsie utilizzando i metodi precedentemente descritti2. Inoltre, maggiori dettagli tecnici su come preparare gli organoidi da materiale derivato dal paziente o da iPSC possono essere trovati in Lees et al. e Mahe et al.24,25.
- Una volta stabilite le colture organoidi, seguire la routine di scissione descritta di seguito per preparare gli organoidi a eseguire infezioni con patogeni enterici.
- Seminare 20-100 organoidi in 50 μL di soluzione ECM al 100% in piastre trattate con colture non tissutali a 24 pozzetti. Aggiungere 500 μL di substrati di crescita (Tabella 1) per pozzo.
- Cambiare il supporto ogni 2 giorni rimuovendo 250 μL del vecchio supporto e aggiungendo 250 μL del mezzo di crescita fresco a ciascun pozzetto. Riscaldare il supporto a temperatura ambiente prima di cambiare il supporto.
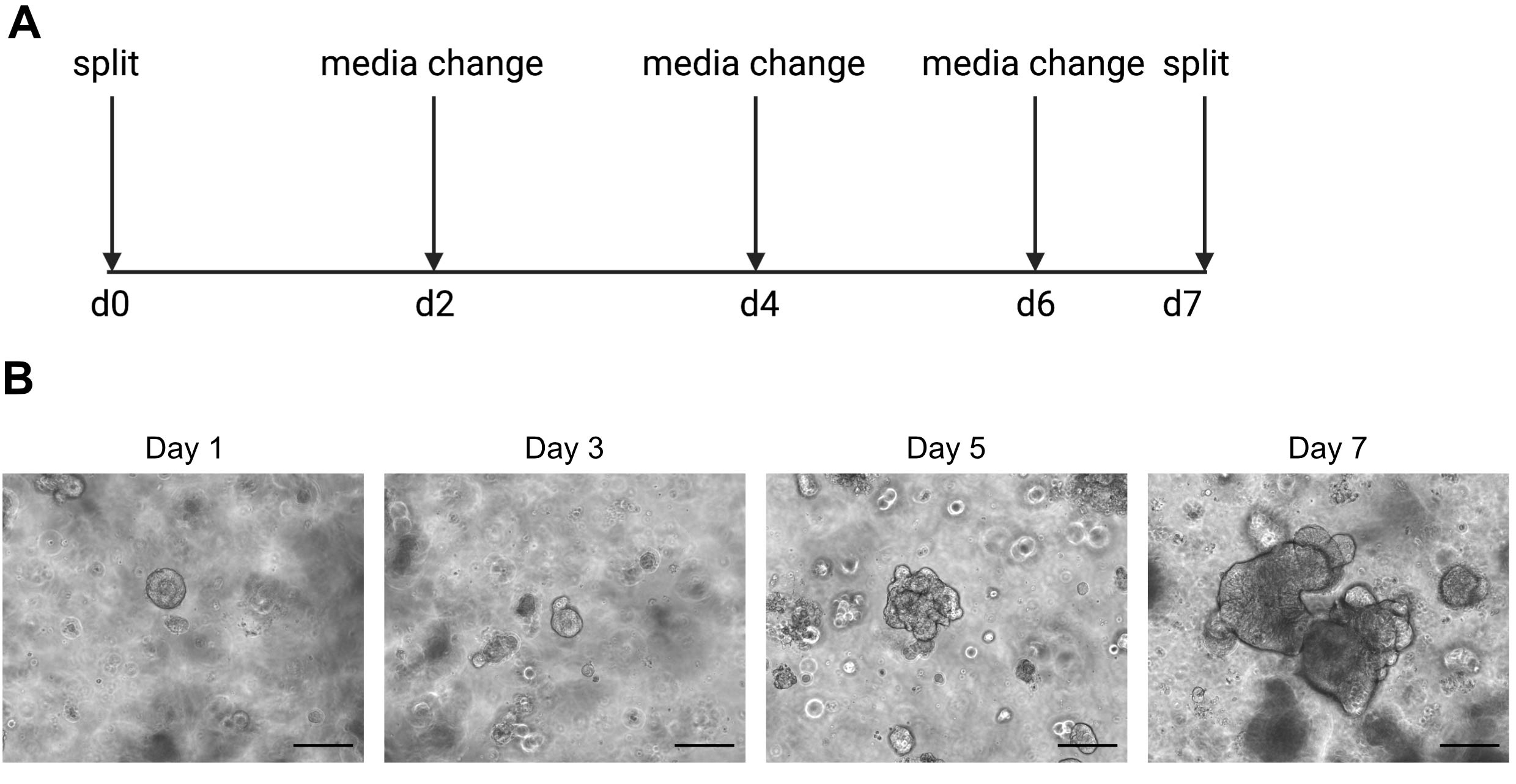
ATTENZIONE: I fluidi freddi liquefanno la soluzione di ECM contenente organoidi. - Passare gli organoidi una volta alla settimana quando l'interno inizia a diventare buio a causa dell'accumulo di cellule morte. Un esempio di questo può essere trovato nella Figura 2.
- Il giorno della scissione, rimuovere la soluzione di ECM da -20 °C e scongelare sul ghiaccio.
- Posizionare la nuova piastra di coltura cellulare vuota che verrà utilizzata per seminare gli organoidi dopo la scissione a 37 °C (questo richiede un minimo di 1 ora per essere caldo e potrebbe essere riscaldato durante la notte). Riscaldare i terreni di coltura a temperatura ambiente.
- Rimuovere il mezzo di crescita con una pipetta P1000 e aggiungere 500 μL di soluzione salina fredda tamponata con 1x fosfato (PBS) a ciascun pozzetto per 3 minuti per liquefare parzialmente la soluzione ECM e dissociarsi dalla piastra.
- Per garantire la completa interruzione della soluzione ECM, utilizzare una pipetta P1000 (set-up a 450 μL). Pipettare su e giù 10 volte per risospese il PBS, la soluzione ECM e gli organoidi, trasferire gli organoidi risospesi in un tubo conico da 15 ml e metterli sul ghiaccio.
- Raccogliere più pozzetti degli stessi organoidi nello stesso tubo conico.
- Se più organoidi diversi (donatori diversi, sezioni diverse, pre-trattamento diversi, ecc.) vengono divisi contemporaneamente, raccoglierli in tubi conici diversi. Durante la raccolta, mantenere i tubi conici sul ghiaccio.
- Girare i campioni a 500 x g per 5 minuti a 4 °C. Rimuovere con attenzione il PBS con una pipetta per mantenere il pellet organoide sul fondo.
- Evitare l'utilizzo di un sistema di scarico sottovuoto con una pipetta montata poiché il pellet di organoidi è molto sciolto e può essere facilmente aspirato quando si rimuove il PBS troppo rapidamente.
- Aggiungere 1 mL di tripsina allo 0,05% al tubo conico e risospesciare gli organoidi pipettando su e giù 10 volte con una pipetta P1000. Incubare il tubo conico contenente organoidi a 37 °C per 3 min.
- Aggiungere 2 ml di dmEM/F12 media contenenti il 10% di siero bovino fetale (FBS) e l'1% di penicillina/streptomicina per fermare la digestione e risospesire per interrompere gli organoidi tubando su e giù 10 volte con una pipetta P1000.
- Girare i campioni a 500 x g per 5 minuti a 4 °C. Rimuovere i tubi conici dalla centrifuga e conservarli sul ghiaccio.
- Rimuovere con attenzione il fluido / tripsina con una pipetta monouso da 5 ml per mantenere il pellet organoide sul fondo. Lasciare circa 500 μL di materiale e rimuoverlo con una pipetta P1000.
- Una volta che il fluido/tripsina è stato rimosso da tutti i tubi conici, rimuovere la piastra trattata con 24 pozzetti non in coltura cellulare dall'incubatore a 37 °C e posizionarla sotto il cofano della coltura cellulare.
- Aggiungere la soluzione di ECM al 100% (mantenuta sul ghiaccio) al tubo conico con gli organoidi divisi in rapporto da 1:3 a 1:5 a seconda delle abitudini di crescita dei singoli organoidi donatori. (Ad esempio, se un pozzo contenente 20-100 organoidi in una goccia di 50 μL di soluzione di ECM è stato passato, risospesere il pellet organoide in 150-250 μL di soluzione ECM ghiacciata.)
- Seme 50 μL di soluzione ECM/miscela organoide per pozzetto della piastra trattata con coltura non cellulare a 24 pozzetti preriscaldata a 37 °C.
NOTA: la soluzione ECM polimerizzerà molto rapidamente una volta riscaldata. Mantenere la soluzione ECM fredda in ogni momento (conservata su ghiaccio) durante l'uso. Quando gli organoidi sono stati risospesi, seminare immediatamente su un nuovo piatto. Per i principianti, si consiglia di conservare una scatola di punte per pipette a -20 °C per consentire un tempo extra per la semina. - Incubare la piastra a 24 pozzetti a 37 °C per 10-15 minuti per consentire alla soluzione ECM di polimerizzare.
- Dopo la polimerizzazione, aggiungere 500 μL di terreno di crescita (Tabella 1)26 a ciascun pozzetto e incubare gli organoidi a 37 °C. Controllare gli organoidi quotidianamente al microscopio. Cambiare il supporto ogni 2 giorni come nel passaggio 1.4.
2. Preparazione di organoidi in due dimensioni (2D) per l'infezione apicale
NOTA: Il seguente protocollo descriverà come seminare gli organoidi intestinali come un monostrato di cellule in una piastra di coltura cellulare per infettare le cellule dell'epitelio intestinale dal loro lato apicale. Utilizzare la piastra a 48 pozzetti per gli esperimenti di sequenziamento e il vetrino a camera di vetro a 8 pozzetti per controllare l'infezione utilizzando approcci di immunofluorescenza.
- Coltivare e mantenere gli organoidi come descritto nel paragrafo 1.
- Prima di seminare gli organoidi in 2D, rivestire le piastre a 48 pozzetti e lo scivolo della camera di vetro a 8 pozzetti con 200 μL di collagene umano al 2,5% in acqua per pozzo per 1 ora a 37 °C.
NOTA: gli organoidi intestinali sono meglio seminati in piastre a 48 pozzetti e in vetrino a 8 pozzetti. L'esperienza ha dimostrato che non infettano bene in piastre da 96 pozzetti. Gli inserti Transwell possono anche essere utilizzati per consentire l'infezione sia apicale che basolaterale in 2D. Se si utilizzano pozzi trans, monitorare la resistenza elettrica transepiteliale (TEER) prima dell'infezione per confermare un monostrato confluente. Normalmente, gli organoidi intestinali hanno una barriera stretta con un TEER di 450-600 Ohm/cm2. - Semina 100-150 organoidi in ogni pozzetto di una piastra a 48 pozzetti o per un pozzetto di una slitta a camera di vetro a 8 pozzetti.
- Per stimare il numero di organoidi, contare il numero di organoidi presenti nella goccia di soluzione di 50 μL di ECM della piastra a 24 pozzetti preparata nella sezione 1. In media, sono necessari 1-2 pozzi, che daranno circa 15.000-30.000 cellule.
- Per interrompere la soluzione ECM e tripsinizzare gli organoidi, seguire i passaggi 1.8-1.17.
- Rimuovere il collagene umano dalle piastre a 48 pozzetti e dalla slitta della camera con fondo di vetro a 8 pozzetti.
- Sospendere il pellet organoide nel tubo conico in 250 μL di terreno di crescita/pozzetto e aggiungere la miscela ai pozzetti rivestiti di collagene. Posizionare la piastra in un'incubatrice a 37 °C.
NOTA: quando si eseguono gli esperimenti, vengono confrontate più condizioni (finto vs. infetto +/- trattamento di interesse). Per ridurre al minimo la variabilità tra ogni pozzetto di organoidi seminati 2D, raccogliere il numero totale di organoidi necessari nello stesso tubo conico. La raccolta di organoidi da un massimo di 12 pozzetti di una piastra a 24 pozzetti può utilizzare 1 mL di tripsina e 2 ml di mezzi neutralizzanti. Quando si utilizzano 12-24 pozzetti, aumentare questo a 2 ml di tripsina e 4 ml di mezzi neutralizzanti. - Dopo 48 ore, rimuovere la piastra dall'incubatore e posizionarla nella cappa di coltura cellulare. Rimuovere il mezzo di crescita e sostituirlo con 250 μL per pozzetto di mezzi di differenziazione (Tabella 1). Ripetere questa modifica del supporto 48 ore dopo.
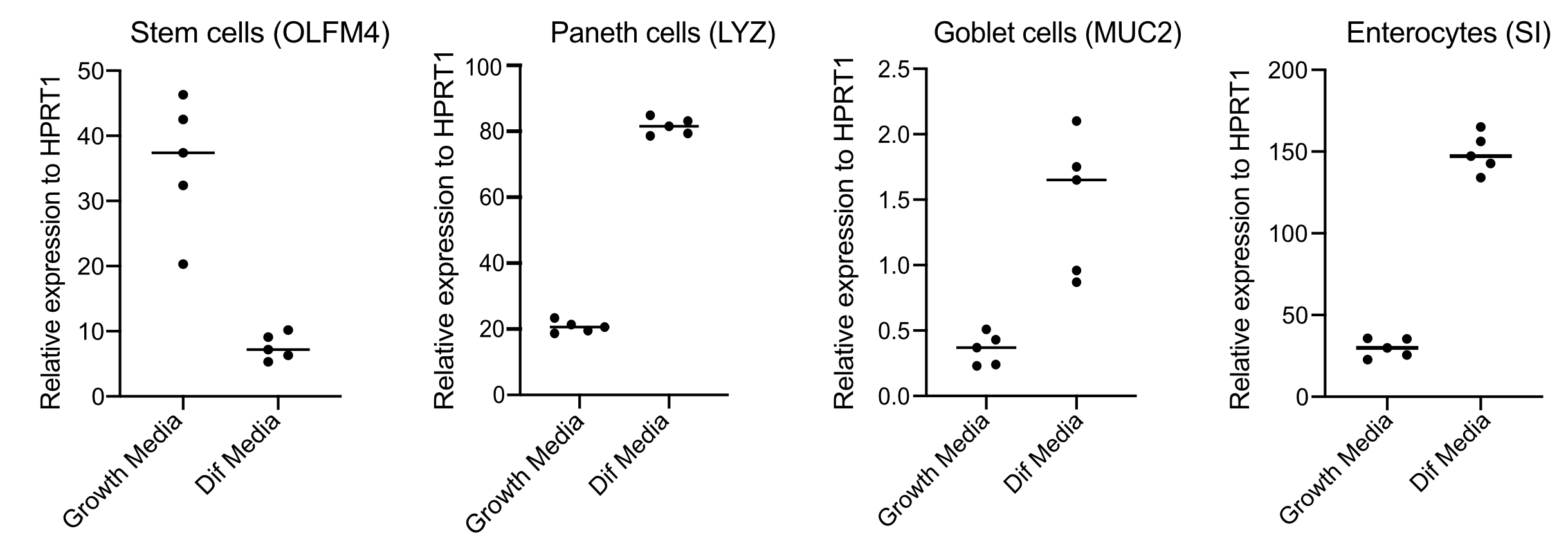
- Dopo 48 ore, confermare la differenziazione (quattro giorni dopo il passaggio ai mezzi di differenziazione) estraendo RNA, producendo cDNA con 250 ng di RNA ed eseguendo qPCR a base verde SYBR utilizzando primer per cellule staminali (OLFM4 e / o SMOC2), cellule calici (MUC2), cellule enteroendocrine (CHGA) e enterociti (SI e / o CYP3A4) (Figura 3).
NOTA: Al passaggio dai mezzi di crescita ai mezzi di differenziazione, gli organoidi diminuiranno la loro espressione di marcatori di cellule staminali (OLFM4 e/o SMOC2) e aumenteranno l'espressione dei marcatori delle cellule caliciformi (MUC2), delle cellule enteroendocrine (CHGA) e degli enterociti (SI e/o CYP3A4) mediante qPCR. Preparare un pozzo extra e mantenere con i mezzi di crescita per confrontare il livello di espressione di ciascun marcatore specifico del tipo di cellula in crescita rispetto ai mezzi di differenziazione. - Alla conferma della differenziazione, gli organoidi sono pronti per l'infezione apicale. Spostare gli organoidi al livello di contenimento della biosicurezza richiesto per l'agente patogeno di scelta.
ATTENZIONE: Fare riferimento alle normative locali dell'istituto e alle procedure operative standard quando si maneggiano agenti patogeni sotto il contenimento BSL-2 o BSL-3. - Per infettare gli organoidi intestinali coltivati in 2D dal loro lato apicale, rimuovere i mezzi con una pipetta P1000 e aggiungere l'agente patogeno diluito alla molteplicità di infezione richiesta per l'esperimento nei mezzi di differenziazione (il volume minimo di miscela patogeno/media è di 50 μL per pozzetto e il volume massimo è di 250 μL per pozzetto).
- Lasciare che l'infezione proceda per 2 ore a 37 °C con un tavolo a bilanciere situato nell'incubatore di colture cellulari o oscillando manualmente la piastra ogni 15-20 minuti. Ottimizza questo tempo in base all'agente patogeno scelto.
- Dopo 2 ore, rimuovere il supporto con una pipetta P1000 e sostituirlo con 250 μL per pozzetto di mezzi di differenziazione freschi. Incubare le cellule a 37 °C fino al punto temporale del sequenziamento a singola cellula.
- Per preparare le cellule per il sequenziamento a singola cellula, procedere alla sezione 4.
3. Preparazione di organoidi in tre dimensioni (3D) per l'infezione apicale e basolaterale
- Coltivare organoidi come descritto nel paragrafo 1 in una piastra trattata con 24 pozzetti non cellulari.
- Due giorni dopo il passaggio, rimuovere la piastra dall'incubatrice e metterla in una cappa di coltura cellulare.
- Rimuovere il mezzo di crescita con una pipetta P1000, sostituirlo con 500 μL/pozzetto di mezzi di differenziazione preriscaldati a temperatura ambiente e posizionare la piastra nell'incubatore a 37 °C.
- Dopo 48 ore, sostituire con mezzi di differenziazione freschi (500 μL/pozzetto).
NOTA: Gli organoidi sono mantenuti in mezzi di differenziazione per un totale di 4 giorni prima dell'infezione. - Confermare la differenziazione come descritto nel passaggio 2.9.
- Alla conferma che si è verificata la differenziazione, gli organoidi sono pronti per l'infezione.
- Scongelare ECM sul ghiaccio. Riscaldare il mezzo di differenziazione a temperatura ambiente e riscaldare una piastra a 24 pozzetti a 37 °C.
- Per eseguire infezioni, rimuovere gli organoidi dalla soluzione ECM.
NOTA: rimuovere quanta più soluzione ECM possibile in quanto i virus preferiscono attenersi alla soluzione ECM piuttosto che alle cellule. Se c'è troppa soluzione di ECM rimanente intorno agli organoidi, l'infettività sarà gravemente influenzata. - Interrompere l'ECM aggiungendo 500 μL di PBS freddo 1x e incubando per 3 minuti. Utilizzare un P1000 per pipettare su e giù 10x. Unire gli organoidi in un tubo e centrifugare a 500 x g per 5 min. Rimuovere PBS con un P1000.
NOTA: Per ridurre al minimo la variabilità tra ciascuna condizione di infezione, combinare gli organoidi provenienti da diversi pozzetti di una piastra a 24 pozzetti nello stesso tubo conico. Ad esempio, se sono necessarie otto condizioni per l'infezione, otto pozzetti di una piastra a 24 pozzetti (contenente ~ 100 organoidi ciascuno) saranno combinati e successivamente divisi in otto pozzetti di una nuova piastra a 24 pozzetti dopo l'interruzione dell'ago (vedere il passaggio 3.12). - Risospese gli organoidi in 1 mL di mezzi di differenziazione. Per l'infezione apicale e basolaterale seguire il passo 3.10.1 e per l'infezione basolaterale seguire solo il passo 3.10.2.
- Attaccare un ago da 27 G a una siringa da 1 mL e risospesciare il pellet sei volte per consentire l'interruzione degli organoidi e per l'infezione sia sul lato apicale che basolaterale. Procedere al passaggio 3.11.
NOTA: Gli organoidi dovrebbero essere i classici organoidi dall'aspetto a cisti e dal polpo con un centro scuro se osservati prima della rottura dell'ago. Dopo l'interruzione, questi organoidi appariranno più piccoli con un interno meno o nessun scuro. Quando si esegue l'infezione, ci saranno sempre un minimo di due condizioni (simulazione e infezione), un minimo di due pozzetti di una piastra a 24 pozzetti sarà inizialmente combinato in un tubo conico da 15 ml per l'interruzione basata sull'ago (spiegando perché un minimo di 1 mL viene utilizzato per la risospensione). Quando si eseguono più di due condizioni, combinare fino a 10 pozzetti di una piastra a 24 pozzetti in un singolo tubo conico da 15 ml e risospesiare in 1 mL di mezzo di differenziazione per la rottura dell'ago. Dopo l'interruzione, aggiungere 500 μL di mezzi di differenziazione per n + 1 (ad esempio, se si utilizzano 10 pozzetti, quindi ricaricare con 4,5 ml per avere 5,5 ml totali). Se sono necessarie più di 10 condizioni, utilizzare diversi tubi conici. Si raccomanda una siringa da 1 mL poiché gli organoidi vengono interrotti meglio con questo volume della siringa. Un'alternativa a una siringa consiste nell'impiegare una punta P1000 ben appiattita. - Sospendere gli organoidi in 500 μL di mezzi di differenziazione per pozzetto. Fare attenzione a non disturbare gli organoidi e assicurarsi che il lato apicale non sia accessibile. Procedere al passaggio 3.11.
- Attaccare un ago da 27 G a una siringa da 1 mL e risospesciare il pellet sei volte per consentire l'interruzione degli organoidi e per l'infezione sia sul lato apicale che basolaterale. Procedere al passaggio 3.11.
- Trasferire 500 μL di sospensioni organoidi per pozzetto in una piastra di coltura cellulare a 24 pozzetti utilizzando una pipetta P1000 e spostare la piastra al livello di contenimento di biosicurezza richiesto per l'agente patogeno di scelta.
ATTENZIONE: Fare riferimento alle normative locali e alle procedure operative standard dell'istituto quando si maneggiano agenti patogeni sotto il contenimento BSL-2 o BSL-3. Eseguire sempre la rottura dell'ago degli organoidi al di fuori del BSL-3 per evitare potenziali incidenti con l'ago. Le normative locali possono anche impedire l'uso di aghi nel BSL-3. - Aggiungere l'agente patogeno diluito in mezzi di differenziazione per raggiungere la molteplicità di infezione richiesta per l'esperimento. Non superare un volume totale di 500 μL (per avere 1 mL totale nella piastra a 24 pozzetti).
- Lasciare che l'infezione proceda per 2 ore (questa volta dovrà essere adattata all'agente patogeno di scelta) a 37 °C con un tavolo a bilanciere situato nell'incubatore di colture cellulari o dondolando manualmente la piastra ogni 15-20 minuti.
- Dopo l'incubazione di 2 ore, raccogliere gli organoidi in un tubo da 1,5 ml per condizione e ruotare a 500 x g per 5 minuti a 4 °C.
- Rimuovere il mezzo contenente l'agente patogeno con una pipetta P1000 e conservarlo sul ghiaccio. Lavare gli organoidi pellet una volta con PBS.
- Sospendere gli organoidi in 50 μL di soluzione ECM al 100% (che in precedenza era scongelata su ghiaccio), placcare in una piastra trattata con colture non cellulari a 24 pozzetti preriscaldata a 37 °C e incubare la piastra a 24 pozzetti a 37 °C per 10-15 minuti per consentire alla soluzione ECM di polimerizzare.
- Dopo la polimerizzazione, aggiungere 500 μL di mezzi di differenziazione a temperatura ambiente in ciascun pozzetto e incubare a 37 °C fino alla raccolta per il sequenziamento monocellulare.
NOTA: se la raccolta deve avvenire più di 48 ore dopo la semina, cambiare il supporto ogni 2 giorni rimuovendo il vecchio supporto e sostituendolo con 500 μL di mezzi di differenziazione freschi. Tuttavia, l'esperienza ha dimostrato che, poiché gli organoidi sono coltivati sotto mezzi di differenziazione, non sopravviveranno molto più a lungo di 48 ore.
4. Preparazione della soluzione monocellulare e preparazione delle perle di gel in emulsione (OGM) in condizioni di livello di biosicurezza 3 (BSL-3)
NOTA: Il completamento delle seguenti fasi richiede che l'apparecchiatura di sequenziamento a cella singola (Tabella dei materiali) e una macchina PCR in grado di gestire reazioni da 100 μL siano presenti all'interno di un impianto BSL-3. Gli organoidi sono coltivati come descritto nel paragrafo 1 e infettati come descritto nei paragrafi 2-3 a seconda dell'agente patogeno e della via di ingresso. In un momento predeterminato dopo l'infezione, gli organoidi vengono raccolti. Di seguito è descritto il metodo utilizzato per raccogliere gli organoidi intestinali.
- Prima della raccolta degli organoidi infetti, rimuovere le perle di gel unicellulare(Tabella dei materiali)da -80 °C e riscaldarle a temperatura ambiente (almeno 30 minuti prima).
- Inoltre, equilibrare il reagente RT, l'agente riducente B e l'enzima RT C (tutti conservati a -20 °C) a temperatura ambiente. Sostituire l'oligo dell'interruttore del modello come descritto nelle istruzioni del produttore.
NOTA: Tutti i seguenti passaggi devono essere eseguiti in relazione alle normative locali sulla biosicurezza seguendo la procedura operativa standard stabilita per l'agente patogeno considerato. Come regola generale per il lavoro BSL-3, è necessario assicurarsi che l'esterno di tutti i vasi (piastre, tubi, ecc.) che escono da una cappa di coltura cellulare siano adeguatamente disinfettati. Lo stesso vale per le mani/guanti della persona che esegue gli esperimenti. - Per eseguire infezioni su organoidi seminati in 2D (sezione 2) procedere al passaggio 4.4. Per eseguire infezioni su organoidi 3D (paragrafo 3) procedere al paragrafo 4.5.
- Per le infezioni di organoidi 2D, portare la piastra di coltura cellulare nella cappa e rimuovere il mezzo di differenziazione con una pipetta P1000.
- Aggiungere 250 μL di 1x PBS a temperatura ambiente a ciascun pozzetto.
- Rimuovere il PBS con una pipetta P1000 e aggiungere 250 μL di enzima di dissociazione a temperatura ambiente (ad esempio, TrypLE Express) a ciascun pozzetto di una piastra a 48 pozzetti. Cambiare i guanti, pulire la piastra e posizionare la piastra con l'enzima di dissociazione nell'incubatore a 37 °C.
- Osservare la dissociazione cellulare al microscopio ogni 5 minuti.
NOTA: Approssimativamente, ci vogliono 15 minuti per dissociare gli organoidi intestinali coltivati in 2D in singole cellule. Questo tempo dovrà essere regolato in quanto dipenderà da quanti organoidi sono stati inizialmente seminati e infettati, nonché dall'agente patogeno poiché alcuni agenti patogeni sono più citopatici e causano la dissociazione degli organoidi molto più velocemente di altri. - Dopo la conferma di un'apparente sospensione monocellulare, riportare la piastra nella cappa di coltura cellulare e interrompere la digestione aggiungendo 250 μL di mezzo DMEM/F12 contenente il 10% di FBS per pozzetto di una piastra a 48 pozzetti.
- Raccogliere le celle in un tubo conico da 15 ml utilizzando una pipetta P1000.
- Cambiare i guanti, pulire il tubo e far girare i campioni a 500 x g per 5 minuti a 4 °C.
- Rimuovere con attenzione l'enzima media/dissociazione con un P1000 per mantenere il pellet cellulare sul fondo. Risospesere singole celle in un volume minimo di PBS contenente lo 0,1% di BSA. Per gli organoidi intestinali, sospendere in 250 μL per ogni pozzetto della piastra a 48 pozzetti.
- Passare le sospensioni cellulari in un tubo FACS con un filtro per rimuovere eventuali grumi di grandi dimensioni e posizionare il tubo FACS contenente cellule sul ghiaccio.
- Procedere al passaggio 4.6 per continuare con il conteggio delle cellule e la preparazione del mix principale.
- Per le infezioni di organoidi 3D, rimuovere la piastra dall'incubatore e posizionarla nella cappa di coltura cellulare.
- Rimuovere il mezzo di differenziazione da ciascun pozzetto della piastra a 24 pozzetti con una pipetta P1000. Aggiungere 500 μL di PBS 1x ghiacciato a ciascun pozzetto e incubare per 3 minuti sul ghiaccio.
- Per garantire la completa interruzione della soluzione ECM, utilizzare una pipetta P1000 (impostata su 450 μL). Pipetta su e giù 10 volte per risospese la soluzione PBS, ECM e gli organoidi; trasferire gli organoidi risospesi in un tubo conico da 15 ml e metterli su ghiaccio. Raccogliere ogni condizione di infezione nel proprio tubo conico da 15 ml.
NOTA: l'uso di un tubo conico da 15 mL fornisce un pellet di cella migliore e più distinto rispetto a un tubo conico da 50 mL o un tubo da 1,5 ml. - Cambiare i guanti e rimuovere il tubo dal cappuccio di coltura cellulare. Pulire l'esterno del tubo.
- Girare i campioni a 500 x g per 5 minuti a 4 °C.
- Spostare nuovamente il tubo nella cappa di coltura cellulare e rimuovere PBS con una pipetta P1000 per evitare di riutilizzare il pellet di organoidi dal fondo del tubo.
- Risospendare il pellet in 1 mL di enzima di dissociazione (ad esempio, TrypLE Express). Cambiare i guanti, pulire il tubo e incubare i campioni a 37 °C.
- Ogni 10 minuti per 30 minuti, spostare il tubo nella cappa di coltura cellulare e risospesciare gli organoidi pipettando su e giù con una pipetta P1000 10 volte.
NOTA: Normalmente, gli organoidi intestinali richiedono circa 30 minuti per formare una sospensione monocellulare quando infettati in 3D. - Per determinare quando gli organoidi sono dissociati a singole cellule, prelevare 10 μL della sospensione organoide usando una pipetta p10.
- Posizionare la sospensione in un vetrino monouso per contatori di celle in plastica. Sigillare la porta di ingresso del campione con nastro trasparente.
- Cambiare i guanti e pulire l'esterno del contatore cellulare. Utilizzare un microscopio a campo luminoso per determinare se viene prodotta una sospensione a singola cellula.
- Dopo la conferma di una sospensione a cella singola, interrompere la digestione aggiungendo 1 mL di supporto DMEM/F12 contenente il 10% di FBS e pipetta su e giù 10 volte con una pipetta P1000.
- Cambiare i guanti, pulire il tubo e far girare i campioni a 500 x g per 5 minuti a 4 °C.
- Rimuovere con attenzione l'enzima media/dissociazione con una pipetta P1000 per evitare di risospendere il pellet cellulare dal fondo del tubo.
- Risospesciare le singole celle in un volume minimo di PBS contenente lo 0,1% di BSA. Per gli organoidi intestinali si sospende in 250 μL.
- Passare le sospensioni cellulari in un tubo FACS con un filtro per rimuovere eventuali grumi di grandi dimensioni e posizionare il tubo FACS contenente cellule sul ghiaccio.
- Procedere al passaggio 4.6 per continuare con il conteggio delle cellule e la preparazione del mix principale.
- Determinare il numero di cellule per μL aggiungendo 10 μL della sospensione cellulare a una camera di conteggio delle cellule di plastica usa e getta.
- Sigillare la porta di ingresso del campione con nastro trasparente prima di rimuovere il campione dalla cappa di coltura cellulare, poiché in questa fase la sospensione cellulare è ancora infettiva.
- Cambia i guanti, pulisci la camera di conteggio delle cellule e conta il numero di cellule usando un microscopio a campo luminoso.
- All'interno della cappa di coltura cellulare, preparare un mix principale di reagente RT, Oligo Interruttore modello, Agente riducente B ed enzima RT C in un tubo da 1,5 ml secondo le istruzioni del produttore a seconda del numero di campioni nell'esperimento. Per ogni campione, aliquota 33,4 μL di miscela master in un tubo PCR e conservare su ghiaccio.
- Aggiungere le celle e l'acqua alla miscela principale in base al numero di cella di destinazione come descritto nelle istruzioni del produttore.
NOTA: il 50%-60% delle celle viene normalmente recuperato (ad esempio, quando si caricano 10.000 celle sul chip, vengono utilizzate 5.000-6.000 celle per l'analisi). Pertanto, caricare sempre 10.000 celle. Se la densità cellulare non lo consente, centrifugare le celle e risospenderle in un volume inferiore. Fare attenzione a non sovraccaricare il chip in quanto ciò comporterà l'intasamento del chip. Inoltre, se le cellule non sono adeguatamente dissociate, ci sarà un aumentato rischio di ottenere più cellule per perlina, che dovranno essere rimosse nell'elaborazione bioinformatica a valle. - Cambiare i guanti, spostare il controller a cella singola nella cappa di coltura cellulare e preparare il chip poiché il chip è coperto solo da una guarnizione non sigillata.
NOTA: la macchina deve essere all'interno del cofano per evitare l'esposizione a sospensioni cellulari infette e potenziali aerosol. - Aggiungere il chip a cella singola al portachip e riempire le corsie inutilizzate con il 50% di glicerolo.
- Aggiungere il mix principale, le perline e l'olio di partizionamento alle corsie utilizzate per i campioni secondo le istruzioni del produttore.
- Coprire il chip con la guarnizione, caricare il chip nel controller e avviare il programma.
NOTA: si consiglia di caricare solo sei delle otto corsie. I problemi con emulsioni improprie si verificano spesso nelle corsie uno e otto. La società afferma che tutte le corsie sono indipendenti e questo non dovrebbe accadere; tuttavia, se possibile, evitare queste due corsie ed eseguire due chip se sono necessari otto campioni. - Al termine del programma, rimuovere il chip e la guarnizione.
- Utilizzare una pipetta multicanale e trasferire 100 μL delle emulsioni in un tubo PCR pulito. Assicurarsi che ogni pozzo abbia un colore bianco uniforme che indichi che si è verificata un'emulsione completa.
- Cambiare i guanti, pulire i tubi e trasferire il tubo PCR a una macchina PCR in grado di supportare reazioni da 100 μL. Eseguire il programma: 53 °C per 45 min; 85 °C per 5 min.
- Al termine, conservare i campioni a 4 °C. A questo punto, elaborare la reazione secondo le istruzioni del produttore o conservare a 4 °C per 3 giorni o -20 °C per 1 settimana.
- Dopo 5 minuti a 85 °C, la maggior parte dei virus avvolti verrà inattivata, rimuoverà il cDNA dal BSL-3 secondo la normale procedura operativa ed eseguirà i preparativi della libreria in un laboratorio BSL-1.
NOTA: Questo esperimento deve essere eseguito come tre repliche biologiche (ad esempio, in tre giorni diversi) perché l'entità della differenziazione di ciascun tipo di cellula può variare leggermente tra ogni esperimento. Le librerie di sequenziamento risultanti potrebbero essere indicizzate e sequenziate in modo diverso in un'unica esecuzione di sequenziamento.
Risultati
Preparazione di organoidi per il sequenziamento monocellulare
I risultati del sequenziamento a singola cellula dipendono fortemente dall'utilizzo di cellule di buona qualità. Per garantire che gli organoidi siano di buona qualità, dovrebbero essere adeguatamente mantenuti e osservati quotidianamente per determinare quando sono pronti per essere suddivisi (Figura 2). La tempistica della scissione degli organoidi dipende dal donatore; alcuni donatori crescono più rapidamente e devono essere divisi ogni 5 giorni, mentre altri sono più lenti e devono essere divisi ogni 10 giorni. In media, gli organoidi vengono divisi una volta alla settimana quando i centri diventano scuri (Figura 2B). Se gli organoidi sono lasciati diventare troppo grandi e accumulare troppe cellule morte al centro, l'organoide morirà.
Gli organoidi sono mantenuti in un mezzo che contiene elevate quantità di Wnt3A. Questo supporta la nicchia delle cellule staminali e promuove gli organoidi a continuare a crescere e proliferare. In queste condizioni di crescita, gli organoidi contengono elevate quantità di cellule staminali e cellule che amplificano il transito e una minore quantità di popolazioni cellulari differenziate come enterociti maturi, cellule a calice e cellule enteroendocrine. Tuttavia, per imitare la complessità cellulare che si trova all'interno dell'intestino umano, è importante spingere la differenziazione cellulare e produrre più di queste cellule. Ciò si ottiene modificando le condizioni del supporto e rimuovendo Wnt3A e riducendo R-Spondin e Noggin (Tabella 1). Normalmente, la differenziazione cellulare verso enterociti, cellule caliciformi e cellule enteroendocrine richiede 4 giorni di mezzi di differenziazione (Figura 3). È fondamentale ottenere una buona differenziazione; altrimenti, la valutazione del tropismo patogeno e delle risposte specifiche del tipo cellulare diventerà difficile.
Conferma dell'inattivazione del patogeno BSL-3
La piena inattivazione deve essere confermata con l'agente patogeno di scelta e convalidata che è sicuro rimuovere il cDNA dal BSL-3. Per SARS-CoV-2, la completa inattivazione del virus è stata convalidata prendendo 100 μL di SARS-CoV-2 e incubandolo in una macchina PCR per 5 minuti a 85 °C. Il virus è stato quindi aggiunto di nuovo alle cellule vero naïve e l'infezione virale è stata confrontata con il virus non trattato termicamente mediante test di immunofluorescenza e placca a 24 ore, 48 ore e 72 ore dopo l'infezione per garantire che tutte le particelle non fossero più infettive. Questi risultati sono stati inviati all'agenzia di regolamentazione locale e, dopo la loro approvazione, è stato eseguito l'esperimento a cella singola e l'elaborazione del cDNA.
Risultati del sequenziamento a singola cellula
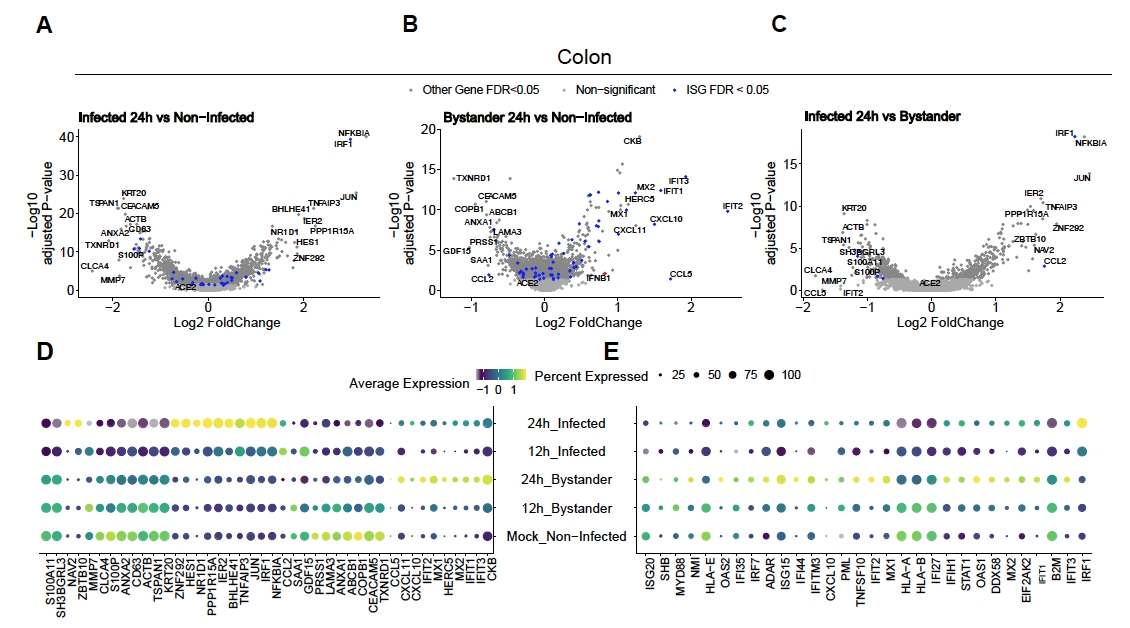
Per valutare come SARS-CoV-2 infetta il colon umano e gli organoidi dell'ileo, è stato eseguito il sequenziamento a singola cellula. Gli organoidi sono stati preparati come descritto sopra e infettati in un formato 2D per consentire l'infezione apicale da SARS-CoV-2. Le cellule infette sono state raccolte a 12 ore e 24 ore dopo l'infezione e sono state elaborate per il sequenziamento a singola cellula come descritto sopra. L'analisi dei dati di sequenziamento a singola cellula ha permesso di determinare che solo una sottopopolazione (enterocita 2 immaturo) di cellule epiteliali intestinali umane supportava l'infezione da SARS-CoV-2 (Figura 4). Inoltre, poiché non tutte le cellule di una popolazione sono state infettate, sono state analizzate sia le cellule infette che le cellule astanti non infette (Figura 5). Questi risultati hanno mostrato che SARS-CoV-2 ha indotto una cascata di segnali pro-infiammatori nelle cellule infette mentre le cellule astanti non infette hanno mostrato una risposta immunitaria mediata dall'interferone. Inoltre, scRNA-Seq ha mostrato che le cellule infette non erano in grado di rilevare gli interferoni a causa del blocco mediato dal virus della via (Figura 5). Non è stato possibile ottenere queste informazioni quando si utilizza il sequenziamento dell'RNA di massa.

Figura 1: Schema che illustra i tre diversi metodi per preparare gli organoidi intestinali umani all'infezione da patogeni enterici. L'infezione apicale può essere ottenuta attraverso la semina di organoidi intestinali in 2D. Un'infezione apicale e basolaterale può essere eseguita interrompendo l'organoide 3D. Infine, un'infezione solo basolaterale può essere eseguita infettando organoidi intestinali 3D intatti. Ognuno di questi metodi può essere utilizzato per generare campioni per il sequenziamento di singole cellule. Fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}

Figura 2: Schema di mantenimento degli organoidi e immagini rappresentative dei campi luminosi. (A) Schema per il mantenimento e il passaggio di organoidi intestinali umani. (B). Immagini rappresentative in campo luminoso dei giorni 1, 3, 5 e 7 post-scissione. Entro il giorno 7 gli organoidi diventano grandi e scuri a causa dell'accumulo di cellule morte e sono pronti per essere divisi. La barra della scala indica 25 μm. Fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}

Figura 3: QPCR rappresentativa degli organoidi intestinali umani 4 giorni dopo il passaggio ai mezzi di differenziazione. Gli organoidi intestinali sono stati mantenuti nei mezzi di crescita o sono passati ai mezzi di differenziazione per 4 giorni. L'RNA è stato raccolto e la qPCR è stata eseguita per i marcatori di cellule staminali (OLFM4), cellule paneth (LYZ), cellule caliciformi (MUC2) e enterociti (SI). N = 5. Fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}

Figura 4: Identificazione della popolazione cellulare infettata da SARS-CoV-2. Gli organoidi derivati dal colon umano e dall'ileo sono stati infettati da SARS-CoV-2. Dopo 12 e 24 ore le cellule post-infezione sono state raccolte e sottoposte al sequenziamento dell'RNA unicellulare per identificare quali popolazioni cellulari supportavano l'infezione da SARS-CoV-2. È stato riscontrato che l'infezione da virus aumenta nel tempo e infetta principalmente l'enterocita immaturo 2. Questa cifra è stata modificata da Triana et al.19. Fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}

Figura 5: Determinazione della risposta immunitaria intrinseca innata. Gli organoidi derivati dal colon umano sono stati infettati da SARS-CoV-2. Dopo 12 e 24 ore le cellule post-infezione sono state raccolte e sottoposte al sequenziamento dell'RNA a singola cellula per determinare la risposta immunitaria innata intrinseca sia nelle cellule astanti viralmente infette che in quelle non infette. Le cellule infette da SARS-CoV-2 hanno mostrato una forte risposta pro-infiammatoria mentre le cellule astanti non infette hanno mostrato una risposta mediata dall'interferone. Figura modificata da Triana et al.19. Fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}
| Mezzi di crescita | |
| Composto | Concentrazione finale |
| Annuncio DMEM/F12 | GlutaMAX (1X) |
| +GlutaMAX | HEPES 1 mM |
| +HEPES | Penna 10 U/mL |
| +P/S | Streptococco 10 μg/mL |
| L-WRN | 50% in volume |
| B27 · | 01:50 |
| N-acetil-cisteina | 1 mM |
| FEG | 50 ng/ml |
| A83-01 | 500 nM |
| IGF-1 · | 100 ng/ml |
| FGF di base | 50 ng/ml |
| Gastrina | 10 mM |
| Mezzi di differenziazione | |
| Composto | Concentrazione finale |
| Annuncio DMEM/F12 | GlutaMAX (1x) |
| +GlutaMAX | HEPES 1 mM |
| +HEPES | Penna 10 U/mL |
| +P/S | Streptococco 10 μg/mL |
| B27 · | 01:50 |
| N-acetil-cisteina | 1 mM |
| R-spondin | 5% in volume |
| Zucca | 50 ng/ml |
| FEG | 50 ng/ml |
| Gastrina | 10 mM |
| A83-01 | 500 nM |
Tabella 1: Composizione dei media per i mezzi di crescita e differenziazione.
Discussione
I patogeni enterici iniziano più spesso il loro ciclo di vita infettando le cellule epiteliali intestinali dal loro lato apicale rivolto verso il lume dell'intestino. Mentre gli organoidi sono ben riconosciuti come un buon modello per riprodurre la complessità cellulare e l'organizzazione dell'epitelio intestinale, la loro organizzazione come strutture tridimensionali e chiuse rende la loro membrana apicale inaccessibile al patogeno. Questo protocollo descriveva metodi per infettare gli organoidi intestinali dal loro lato apicale, dal loro lato basolaterale o da entrambi con patogeni BLS-3. Questi protocolli possono essere facilmente adattati per studiare qualsiasi agente patogeno enterico sotto il contenimento BSL-2 o BLS-3 o qualsiasi altro modello organoide seguendo alcuni passaggi critici evidenziati di seguito. Il metodo sopra descritto è per l'isolamento e la preparazione di goccioline a cellula singola in conformità con le normative in Germania. Come dichiarazione di non responsabilità, questo protocollo non descrive le misure di gestione della biosicurezza (procedure operative standard) che devono essere adottate mentre si lavora in condizioni BSL-3. È anche importante insistere sul fatto che le normative possono variare in altri paesi e che le autorità locali devono essere contattate per assicurarsi che tutte le normative locali siano rispettate.
Uno dei passaggi critici nella semina di organoidi in due dimensioni per l'infezione apicale è il controllo che le cellule si differenzino in modo simile rispetto a quando vengono coltivate come organoidi tridimensionali classici. A seconda del patogeno enterico, il tropismo potrebbe essere limitato a cellule molto rare o a cellule che devono essere altamente differenziate. In questo caso, l'utilizzo di un organoide bidimensionale che non si differenzia completamente potrebbe comportare l'errata conclusione che questo patogeno enterico non possa infettare gli organoidi intestinali. Si suggerisce, se possibile, di eseguire infezioni utilizzando le tre configurazioni di questo protocollo: organoide 2D solo per l'infezione apicale (sezione 2), organoidi 3D aperti crackati per infezione apicale e basolaterale (sezione 3) e organoidi 3D completi solo per l'infezione basolaterale (sezione 3). Questo approccio aiuterà a discernere la via di ingresso del patogeno (apicale vs basolaterale) e controllerà anche che sia stato raggiunto un livello simile di differenziazione. Un'alternativa per l'infezione apicale 2D è la microiniezione, che utilizzerà un organoide 3D ma consegnerà l'agente patogeno direttamente nel lato apicale (vedi Bartfeld et al.27 per i dettagli). Questo metodo richiede un iniettore esperto per garantire che l'agente patogeno sia posizionato correttamente e che gli organoidi rimangano intatti. La microiniezione è comunemente usata nel contenimento di BSL-2 e potrebbe non essere adatta per il contenimento di BSL-3.
Un'ulteriore considerazione importante quando si eseguono esperimenti di infezione in organoidi seminati 2D è la densità cellulare finale. Come accennato nel passaggio 2.3, 100-150 organoidi saranno seminati in un pozzetto di una piastra a 48 pozzetti o in un pozzo di una slitta a camera di vetro a 8 pozzetti. A seconda della linea organoide e della persona che maneggia gli organoidi, la dimensione di questi organoidi può essere significativamente diversa. Ciò potrebbe comportare densità cellulari molto diverse nella piastra a 48 pozzetti o nella slitta della camera con fondo di vetro a 8 pozzetti. A seconda del virus enterico, alcuni virus preferiscono cellule più sparse, mentre altri saranno anche in grado di infettare le cellule confluenti. L'origine molecolare di tali differenze di infettività per diverse confluenze cellulari non è chiara; pertanto, gli esperimenti pilota volti a trovare la migliore densità cellulare per il patogeno enterico di scelta dovrebbero essere eseguiti prima di eseguire un'ulteriore caratterizzazione a valle.
Spesso lo smistamento FACS viene eseguito prima di eseguire l'emulsione di goccioline a cellula singola. Questo passaggio viene spesso utilizzato per separare le cellule morte da quelle vive e le singole cellule dai doppietti. Quando si lavora con agenti patogeni BSL-3, è necessario che la struttura sia dotata di un selezionatore FACS appropriato, il che non accade spesso. Inoltre, non tutte le cellule di un organoide hanno le stesse dimensioni ed è spesso difficile discriminare tra un doppietto o una cellula più grande, causando così il rischio di selezionare negativamente un tipo di cellula specifico. Inoltre, c'è ancora discussione sul campo se il tempo necessario per l'ordinamento tra 5.000-10.000 per ciascun campione possa comportare una modifica significativa del profilo di trascrizione delle singole celle. Mentre sono stati descritti metodi di fissazione cellulare compatibili con il sequenziamento a singola cellula (ad esempio, metanolo e RNAassist), è stato osservato che ciò porta ad una diminuzione della qualità del sequenziamento18. Infine, si sospetta che lo smistamento delle cellule utilizzando marcatori di morte cellulare possa anche portare a un pregiudizio. Data la proliferazione direzionale e la differenziazione delle cellule attraverso l'asse cripto-villi, le cellule più differenziate, che stanno per essere liberate e rilasciate, si trovano sulla punta dei villi. Queste cellule sono spesso positive per diversi marcatori delle vie di morte cellulare (adesempio, apoptosi, necrosi e necroptosi); tuttavia, quando si osserva l'infezione da rotavirus dell'intestino del topo, la punta dei villi è l'area più infetta28. Pertanto, filtrare le cellule che possono sembrare positive per i marcatori di morte comporterebbe una selezione negativa delle cellule infette che possono rappresentare l'infezione fisiologica. Attualmente, non esiste una buona soluzione per lo smistamento e il fissaggio degli organoidi prima del sequenziamento a singola cellula. Si raccomanda l'uso di cellule vive e non selezionate in quanto sono necessari ulteriori studi per trovare protocolli alternativi adatti.
Il sequenziamento a singola cellula ha rivoluzionato il modo in cui le risposte cellulari possono essere valutate. Questa tecnica consente l'identificazione di risposte specifiche del lignaggio cellulare sia in condizioni basali che sotto infezioni patogene. Questo metodo ha aperto porte in molti campi che in precedenza erano limitati da letture di massa. Mentre questo metodo è molto potente, ha i suoi limiti. Una limitazione chiave è l'ampia analisi bioinformatica richiesta a valle del sequenziamento. Ciò è particolarmente importante quando si analizzano i tessuti e si assegnano tipi di cellule in cui attualmente non vi è alcuna annotazione. Avere un bioinformatico esperto è necessario per supportare tutti gli studi su singole cellule.
Questo protocollo descrive come seminare e gestire gli organoidi intestinali umani, infettarli con agenti patogeni enterici ed eseguire scRNAseq. Adattare questo approccio ad altri organi è ora possibile, poiché per la maggior parte degli organi sono stati sviluppati sistemi di modelli organoidi. Gli organoidi polmonari ed epatici sono organizzati in modo simile rispetto agli organoidi intestinali e, come tali, l'uso di un approccio analogo potrebbe essere trasposto a questi organoidi. Il controllo critico sarà quello di convalidare che quando vengono coltivati in due dimensioni o aperti, questi organoidi raggiungono una differenziazione simile a quella delle loro controparti organoidi 3D. Le caratteristiche specifiche e i geni che definiscono uno stato differenziato sono specifici per ciascun modello di organo. Altri modelli organoidi come gli organoidi renali e vascolari, grandi strutture dense, avranno bisogno di metodi aggiuntivi per dissociare serialmente queste strutture in singole cellule.
Divulgazioni
Gli autori non dichiarano interessi finanziari concorrenti.
Riconoscimenti
Megan Stanifer e Steeve Boulant sono state sostenute da borse di ricerca della Deutsche Forschungsgemeinschaft (DFG): (numero di progetto 240245660, 278001972, 415089553 e 272983813 a Steeve Boulant e 416072091 a Megan Stanifer), allo stato del Baden-Wuerttemberg e al Bundesministerium für Bildung und Forschung BMBF 01KI20239B a MS e 01KI20198A e (NUM-COVID 19, Organo-Strat 01KX2021) a SB. Gli schemi sono stati creati in BioRender.
Materiali
| Name | Company | Catalog Number | Comments |
| Recombinant mouse noggin | Peprotech | Cat#250-38 | |
| [Leu15]-Gastrin I | Sigma-Aldrich | Cat# G9145 | |
| 0.05% Trypsin-EDTA | Thermo Fischer Scientific | Cat#25300054 | |
| 24-well non-cell culture treated plate | Corning | Cat#3738 | |
| 8-well glass bottom chamber slide | iBIDI | Cart#80827 | |
| A83-01 | Tocris | Cat#2939 | |
| Advanced DMEM/F12 | Thermo Fischer Scientific | Cat# 12634010 | |
| B27 | Thermo Fischer Scientific | Cat#17504-044 | |
| Chromium Controller & Next GEM Accessory Kit | 10X Genomics | Cat#1000202 | Used in the preparation of single cell solution and preparation of Gel beads-in-emulsion (GEM) |
| Chromium Next GEM Chip G Single Cell Kit | 10X Genomics | Cat #1000127 | Used in the preparation of single cell solution and preparation of Gel beads-in-emulsion (GEM) |
| Chromium Next GEM Single Cell 3′ Kit v3.1 | 10X Genomics | Cat#1000268 | Used in the preparation of single cell solution and preparation of Gel beads-in-emulsion (GEM) |
| Collagen from human placenta | Sigma Aldrich | Cat#C5533-5MG | |
| CYP34A forward | Eurofins | GATGGCTCTCATCCCAGACTT | Primers used to check for differentiation |
| CYP3A4 reverse | Eurofins | AGTCCATGTGAATGGGTTCC | Primers used to check for differentiation |
| DMEM/F12 | Thermo Fischer Scientific | Cat#11320074 | |
| EDTA | Sigma Aldrich | Car#E9884 | |
| Fast Read 102 counting slides | Biosigma | Cat# BVS100 | |
| Fetal Bovein Serum (FBS) | Capricorn | Cat#FBS-12A | |
| GlutaMAX | Thermo Fischer Scientific | Cat# 35050061 | |
| HEPES | Thermo Fischer Scientific | Cat3 15630080 | |
| L-WRN cells | ATCC | CRL-3276 | This cell line is used to make the conditioned media containg Wnt 3A, R-Spondin and Noggin. The protocol for the production of the conditioned media can be found on the manufacterures site. |
| MatriGel. GFR, LDEV free | Corning | Cat#354230 | |
| MUC-2 forward | Eurofins | TGTAGGCATCGCTCTTCTCA | Primers used to check for differentiation |
| MUC-2 reverse | Eurofins | GACACCATCTACCTCACCCG | Primers used to check for differentiation |
| N-acetyl-cysteine | Sigma Aldrich | Cat# A9165 | |
| OLMF4 forward | Eurofins | ACCTTTCCCGTGGACAGAGT | Primers used to check for differentiation |
| OLMF4 reverse | Eurofins | TGGACATATTCCCTCACTTTGGA | Primers used to check for differentiation |
| Penicillin/Streptomycin | Thermo Fischer Scientific | Cat#15140122 | |
| Recombinant human FGF basic | Peprotech | Cat#100-18B | |
| Recombinant human IGF-1 | BioLegend | Cat#590904 | |
| Recombinant mouse EGF | Thermo Fischer Scientific | Cat# PMG8043 | |
| SI forward | Eurofins | AATCCTTTTGGCATCCAGATT | Primers used to check for differentiation |
| SI reverse | Eurofins | GCAGCCAAGAATCCCAAT | Primers used to check for differentiation |
| TrypLE Express | Thermo Fischer Scientific | Cat#12605036 | |
| Y-27632 | Caymann Chemicals | Cat#10005583 |
Riferimenti
- Kretzschmar, K., Clevers, H. Organoids: Modeling development and the stem cell niche in a dish. Developmental Cell. 38, 590-600 (2016).
- Sato, T., et al. Long-term expansion of epithelial organoids from human colon, adenoma, adenocarcinoma, and Barrett's epithelium. Gastroenterology. 141, 1762-1772 (2011).
- Sato, T., et al. Single Lgr5 stem cells build crypt-villus structures in vitro without a mesenchymal niche. Nature. 459, 262-265 (2009).
- Tekes, G., Ehmann, R., Boulant, S., Stanifer, M. L. Development of feline ileum- and colon-derived organoids and their potential use to support feline coronavirus infection. Cells. 9, (2020).
- Derricott, H., et al. Developing a 3D intestinal epithelium model for livestock species. Cell and Tissue Research. 375, 409-424 (2019).
- Zhou, J., et al. Infection of bat and human intestinal organoids by SARS-CoV-2. Nature Medicine. 26, 1077-1083 (2020).
- Sato, T., Clevers, H. Growing self-organizing mini-guts from a single intestinal stem cell: mechanism and applications. Science. 340, 1190-1194 (2013).
- Spence, J. R., et al. Directed differentiation of human pluripotent stem cells into intestinal tissue in vitro. Nature. 470, 105-109 (2011).
- de Graaf, M., van Beek, J., Koopmans, M. P. Human norovirus transmission and evolution in a changing world. Nature Reviews Microbiology. 14, 421-433 (2016).
- Leshem, E., et al. Real-world effectiveness of pentavalent rotavirus vaccine among bedouin and jewish children in southern Israel. Clinical Infectious Diseases : An Official Publication of the Infectious Diseases Society Of America. 62, 155-160 (2016).
- Karst, S. M. The influence of commensal bacteria on infection with enteric viruses. Nature Reviews Microbiology. 14, 197-204 (2016).
- Ettayebi, K., et al. Replication of human noroviruses in stem cell-derived human enteroids. Science. 353, 1387-1393 (2016).
- de Wit, E., van Doremalen, N., Falzarano, D., Munster, V. J. SARS and MERS: recent insights into emerging coronaviruses. Nature Reviews Microbiology. 14, 523-534 (2016).
- Scaldaferri, F., et al. The thrilling journey of SARS-CoV-2 into the intestine: From pathogenesis to future clinical implications. Inflammatory Bowel Diseases. 26, 1306-1314 (2020).
- Kipkorir, V., Cheruiyot, I., Ngure, B., Misiani, M., Munguti, J. Prolonged SARS-CoV-2 RNA detection in anal/rectal swabs and stool specimens in COVID-19 patients after negative conversion in nasopharyngeal RT-PCR test. Journal of Medical Virology. 92, 2328-2331 (2020).
- Lamers, M. M., et al. SARS-CoV-2 productively infects human gut enterocytes. Science. 369, 50-54 (2020).
- Stanifer, M. L., et al. Critical role of type III interferon in controlling SARS-CoV-2 Infection in human intestinal epithelial cells. Cell Reports. 32, 107863 (2020).
- Triana, S., et al. Single-cell transcriptomics reveals immune response of intestinal cell types to viral infection. bioRxiv. , (2020).
- Triana, S., et al. Single-cell analyses reveal SARS-CoV-2 interference with intrinsic immune response in the human gut. Molecular Systems Biology. 17, 10232 (2021).
- Tang, F., et al. mRNA-Seq whole-transcriptome analysis of a single cell. Nature Methods. 6, 377-382 (2009).
- Hwang, B., Lee, J. H., Bang, D. Single-cell RNA sequencing technologies and bioinformatics pipelines. Experimental & Molecular Medicine. 50, 1-14 (2018).
- García-Rodríguez, I., Sridhar, A., Pajkrt, D., Wolthers, K. C. Put some guts into it: Intestinal organoid models to study viral infection. Viruses. 12, 1288 (2020).
- Stanifer, M. L., et al. Asymmetric distribution of TLR3 leads to a polarized immune response in human intestinal epithelial cells. Nature Microbiology. 5, 181-191 (2020).
- Lees, E. A., et al. Using human induced pluripotent stem cell-derived intestinal organoids to study and modify epithelial cell protection against salmonella and other pathogens. Journal of Visualized Experiments: JoVE. (147), e59478 (2019).
- Mahe, M. M., Sundaram, N., Watson, C. L., Shroyer, N. F., Helmrath, M. A. Establishment of human epithelial enteroids and colonoids from whole tissue and biopsy. Journal of Visualized Experiments: JoVE. (97), e52483 (2015).
- Fujii, M., et al. Human Intestinal organoids maintain self-renewal capacity and cellular diversity in niche-inspired culture condition. Cell Stem Cell. 23 (6), 787-793 (2018).
- Bartfeld, S., Clevers, H. Organoids as model for infectious diseases: Culture of human and murine stomach organoids and microinjection of Helicobacter Pylori. Journal of Visualized Experiments: JoVE. (105), e53359 (2015).
- Hernandez, P. P., et al. Interferon-lambda and interleukin 22 act synergistically for the induction of interferon-stimulated genes and control of rotavirus infection. Nature Immunology. 16, 698-707 (2015).
Ristampe e Autorizzazioni
Richiedi autorizzazione per utilizzare il testo o le figure di questo articolo JoVE
Richiedi AutorizzazioneThis article has been published
Video Coming Soon
Personale delle biblioteche
Copyright © 2025 MyJoVE Corporation. Tutti i diritti riservati