
Method Article
濃縮ヒト記憶B細胞集団からの所望特異性を持つ抗体の選択と回収のための単細胞スクリーニング法
要約
ヒト末梢血単核細胞から個々の抗原特異的抗体を同定し、回収するBSelex法は、フローサイトメトリーと単一細胞PCRおよびクローニングを組み合わせたものです。
要約
ヒト抗体レパートリーは、潜在的な治療抗体および有用なバイオマーカーの大部分が未開拓の供給源を表す。次世代シーケンシング(NGS)などの現在の計算方法は、配列レベルで抗体レパートリーに関する膨大なデータセットを生成する一方で、特定の抗原または一連に関連する配列を特定するために機能的データが必要である。抗原。ここでは、ヒト血液ドナーから末梢血単核細胞(PBMC)から個々の抗原特異的抗体を同定・回収する方法について説明する。この方法は、成熟したB細胞の初期濃縮を利用し、フローサイトメトリーを介してIgGメモリB細胞を単離するために、発眼細胞マーカーと蛍光標識タンパク質の組み合わせを必要とする。重いチェーンと軽鎖の可変領域がクローン化され、再スクリーニングされます。メモリBセルコンパートメントに限定されますが、この方法は、何百万ものB細胞を調知し、発現と特異性の確認の準備が整った形式で単一のセルからペアの重鎖配列と軽鎖配列を返すフローサイトメトリーを利用します。この方法で回収された抗体は、治療の可能性を考慮することができますが、特異性と機能をバイオインフォマティクスアプローチとリンクして、個体内のB細胞レパートリーを評価することもできます。
概要
抗体は、治療分子の成長クラスであり、任意のヒトにおける既存のB細胞レパートリーは、そのような抗体の潜在的な供給源である。ヒトドナーから回収された場合、それらは適応または「ヒト化」を必要としない、他の動物系で生成される抗体に必要なステップである。B細胞活性化および増殖1、EBV形質転換2、3、およびハイブリドーマ細胞株の生成を含むヒト抗体の同定および単離のためのいくつかの方法が存在する。 、5.しかし、これらの方法はすべて、抗原特異的抗体をスクリーニングして回収するために広範な細胞培養を必要とします。ヒト抗体レパートリーに関する情報は、次世代シーケンシング(NGS)技術の開発により大幅に拡大され、ドナーサンプル中に存在する大量の個々の配列を同定することが可能となっています。しかし、NGSは存在するすべての配列の非依存的なビューを生み出すので、特に希少または低周波抗体の場合には、抗原特異的抗体の同定および単離を可能にしません。
「BSelex」法の目的は、ヒトドナー中の末梢血単核細胞の循環から抗原特異的抗体を同定し、さらなる分析のためにこれらの抗体の配列を単離して回収することです。この方法は、フローサイトメトリーと細胞選別を利用して、記憶B細胞の表面に発現するB細胞受容体(BCR)を利用する。より低スループットの分子生物学法が開始される前に、何百万ものB細胞をフローサイトメトリーを介して抗原特異性のスクリーニングが可能です。重鎖と軽鎖の組み合わせは、細胞配列を一括で分析するほとんどのNGS法では不可能です。ここで説明する方法では、細胞を個別に単離し、重鎖配列と軽鎖配列の両方のペア回復が可能であり、完全なIgGの直接クローニングおよび発現を可能にする。
プロトコル
人間のボランティアからのサンプルの使用は、スクリプス研究所機関試験委員会によって承認されたプロトコルに従いました。献血前にドナーからインフォームド・コンセントが得られた。
1. 試薬の調製
-
末梢血単核細胞(PBMC)
- フィコルプラークプラス分離により、正常なヒトドナーから末梢血単核細胞を回収する。90%の胎児牛血清(FBS)および10%ジメチルスルホキシド(DMSO)の5000万細胞/mLで凍結保存する。-80°Cで細胞を凍結し、長期保存のために液体窒素に移します。
-
選別用標的抗原ペプチドの標識
- 末端ビオチンを用い、標的タンパク質に特異的なペプチド(最大70残渣)を生成または得る。
- ストレプトアビジンを用いた個々のバイオチリン化ペプチドの標識4nmolは、別々のチューブ内のPE(R-フィコエリスリン)またはAPC(アロフィコシアニン)に共有的に付着した。9:1ペプチドのモル比をストレプトアビジン(SA)に使用し、SA-PEの最終濃度は約3.2 μM、SA-APCは5.7 μM SA-APCを調製します。マイナスコントロールとしてビオチンテトラマーを準備します。
- 各ペプチドミックスを一晩、暗闇の中で、ゆっくりと混合して4°Cでインキュベートする。
- ポリアクリルアミドビーズを含むマイクロカラムを通して標識ペプチドを通過させることにより、結合のない蛍油を除去します。
- ペプチドテトラマーを4°Cで2ヶ月まで保存します。
2. セルの並べ替え
- CD22+分離の前に少なくとも1時間16時間、ドナーPBMCを解凍し、37°Cで休息することを可能にする。
- 冷凍PBMCのバイアルを冷凍庫から取り出し、直ちに37°Cの水浴に移します。バイアルがほぼ解凍されると、作業領域に転送します。
- 各バイアルの内容物を、予備温め培地を含む50mLチューブに移します(ドナーを別々に保ちます)。50 mLの最終容積に媒体を加える。内容物を優しく混ぜます。
- 室温で6分間370 x gの遠心分離機。上清を廃棄し、RPMI完全培地の15mLで細胞を再懸濁し、細胞数を行う。
- RPMI完全培地で細胞濃度を2.0 x 107セル/mLに調整し、T75フラスコ以上に細胞を移し、37°C以上で37°Cで1時間以上、16時間までインキュベートして、解凍細胞の回収時間を確保します。
- 4°Cで6分間370 x gで細胞(必要に応じてプール)と遠心分離機を収集します。上清を廃棄し、氷冷MACSバッファー(PBS、pH 7.6+0.5%BSAおよび2 mM EDTA)の5mLで細胞を再懸濁し、MACSバッファで50 mLに持ち込み、セル数を実行します。
- 4°Cで7分間400 x gで細胞を遠心分離します。
- CD22マイクロビーズの捕捉による正の選択によってCD22+B細胞を単離する。すべての洗い出しに MACS バッファを使用します。4 °Cで7分間のカウントと遠心分離機400 x g。予想される回復は、PBMC人口全体の5-10%です。
- mL FACS バッファーあたり 4,000 万個の細胞を再中断します (トリス バッファー、pH 8.0 は 0.5% BSA および 2 mM EDTA を含む)、ドナーの細胞染色を個別に、またはプールとして進めます。
- メモリ B セルの並べ替え用ステイン セル
- 細胞計の設定と使用する標識蛍光球のそれぞれに対する補償のための細胞のアリコートを削除します。コントロールとして染色されていないセルを含めます。
- 得られたCD22+細胞数の最終染色量を計算します(100μLあたり200万個の染色)。
- B細胞(CD19-PerCP-Cy5.5)、IgG(IgG-FITC)、およびメモリコンパートメント(CD27-PECy7)の細胞外マーカーをメーカーの希釈推奨に従って追加し、穏やかに混合する。アリコート107細胞を1.5mLチューブに入れ、残りの細胞を15mLチューブに移す。
- PEとAPCにそれぞれ36nMの最終濃度で負のコントロールチューブにデュアル標識されたビオチン-SAテトラマーを追加し、ソートチューブ内のペプチドの数を掛け、FACSバッファー(例えば、10ペプチドx 36 nM=360 nM)で体積を0.5 mLファイナルにします。
- PEおよびAPCのためにそれぞれ36 nMでソートチューブにペプチドテトラマーを追加し、TBSバッファで100 μLあたり2x106に細胞を持って来ます。暗闇の中で4°Cでインキュベートし、30-60分間穏やかな回転でインキュベートします。
- 4 °C、400 x g、7分で2xを洗浄し、最後の洗浄の前にカウントするアリコートを除去します。 5 mLフィルターキャップチューブを使用してセルをフィルタリングします。DAPI(4',6-diamidino-2-フェニリンドール)染色を細胞膜完全性のマーカーとして選別する直前に0.3 μM最終濃度に添加します。
- フローサイトメトリーによる単一細胞ソート
- ソート用の96ウェルPCRプレートの48ウェルを準備します。
- マスターミックスを調製する:サンプルあたり(10x RTバッファーの2μL、RNase阻害剤の0.5 μL、無菌PCRグレードの水の7.5 μL)。アリコート10 μL/ウェル当たり、カバーし、ソートの準備ができるまで4°Cで保存します。
- セルソーターで負のコントロールを実行し、サンプル全体を分析します。
- 単一細胞のソートに適切な細胞集団を分離するために、フローサイトメトリーゲートを設定します。
- FSC領域対SSC領域をプロットし、ゲートR1を設定してリンパ球を単離します。
- FSC の高さと FSC の幅をプロットし、ゲート R3 を設定して FSC ダブルトを除外します。
- SSC の高さと SSC の幅をプロットし、ゲート R4 を設定して SSC ダブルトを除外します。
- SSC の高さと DAPI をプロットし、ゲート R5 を設定してライブ セルを分離します (DAPI-)。
- IgG 対 CD19 をプロットし、ゲート R6 を設定して IgG+ B セルを分離します (CD19+ IgG+ )
- IgG 対 CD27 をプロットし、ゲート R7 を設定してメモリ B セル (CD27高)を選択します。
- 抗原PE(Ag-PE)対抗原APC(Ag-APC)をプロットし、ゲートR8をAg-PE+/Ag-APC+二重正ゲート内のイベントの数が少ない象限として設定します。
- 単一細胞のソートに適切な細胞集団を分離するために、フローサイトメトリーゲートを設定します。
- ノイズに対するシグナルの決定のために、抗原陽性(Ag+)サンプルから同数のメモリ細胞を収集します。
- 表現型CD19+CD27+Ag-PE+Ag-APC+(ゲートR8)を有する単一細胞を調製96ウェルPCRプレートにソートする。
- アルミテープパッド付きカバープレート、400 x gで1分間遠心分離機、-80°Cで保存して将来のクローニングを行います。
- ソート用の96ウェルPCRプレートの48ウェルを準備します。
3. シングルセルクローニング
-
逆転転写反応
- 単一のBセルソートプレートを80°Cから取り外します。氷の上で2分間解凍し、開く前に井戸の底部に内容物をプールするために3,300 x gで2分間プレートを回します。
- 逆プライマーとして10%の非イオン洗剤とオリゴ(dT)を使用したdNTP/バッファマスターミックスを準備します。単一細胞を含む各ウェルに酵素ミックスを追加し、ミックスしないでください。
- 65°Cで5分インキュベートします。100mM DTT、RNase阻害剤、および逆転写酵素を含む酵素マスターミックスを追加し、総体積を20μLにし、50°Cで10分、80°Cで10分をインキュベートします。
- PCR ステップを開始する前に氷に転送する
-
マルチステップネストされたPCR反応
- 重鎖(HC)および軽鎖(LC)増幅のために別々の入れ子になったPCR反応を使用する。
- PCR増幅の最初のラウンド(ステップI)では、フォワードプライマーのプールと単一のリバースプライマーを使用して、HCおよびLC可変領域を別々のPCR反応で増幅します(図2)。設計上、フォワードプライマーのプールは、生殖細胞のリーダー(L)配列の大部分を増幅し、逆プライマーは、軽鎖16のカッパ(κ)およびラムダ(λ)の両方を含む各鎖の下流定数領域に固有である。
- 各PCR(ステップI)反応のテンプレートとして2.5 μLのcDNA製品を使用してください。それぞれ1 μMの最終反応濃度にプライマーを追加します。10xポリメラーゼバッファーを2倍の濃度に倍増し、反応あたり25μLに最終容積をもたらします。[94 °C/4 分;94 °C /15 s,55 °C /20 s, 68 °C/60 s; 68 °C/3 分] を使用して HC PCR 反応を 50 サイクル実行します。[98 °C/4 分;98 °C /15 s,72 °C /20 s, 72 °C/60 s; 72 °C/3 分] を使用して LC PCR 反応を 50 サイクル実行します。
- ステップI製品の2.5 μLを各(ステップII)PCR反応のテンプレートとして使用します。HCおよびLC(κおよびλ)フレームワーク1領域に特異的なフォワードプライマーのプールと、各抗体鎖の接合領域に特異的なリバースプライマーのプールを追加する。10xポリメラーゼバッファーを2倍濃度に倍増し、ステップIと同じパラメータを使用してそれぞれ50サイクルのPCR反応を実行します。
- 1%アガロースゲルで完成したPCR反応を実行し、正の増幅ヒットを可視化します。ペアアンプリコン(同じセルから重鎖と軽鎖の両方)を回収し、ゲル抽出を介して分離します。正確なライゲーションミックス計算のためにOD260を介してDNA断片濃度を決定します。
- 回収された重鎖および軽鎖断片をリンカーフラグメントと発現ベクターバックボーンと組み合わせ、市販キットを用いて4フラグメントライゲーション反応を用いる。
- 市販キットを使用して、ライゲーション反応を化学的に有能な細菌に変換します。抗生物質プレートにめっきしたら、残りの形質転換培養に成長培地(抗生物質を含む)の4mLを加え、250rpmで一晩37°Cをインキュベートする。これが「ライゲーションミックス培養」です。
- 市販キットを用いて一晩ライゲーション混合培養物からミニプレップDNAを調作し、得られたプラスミドDNA濃度を決定する。
4. 抗原特異性の確認のためのELISAスクリーン
-
カチオン性脂質系試薬を用いて10μgのミニプレップDNAを懸濁細胞293細胞の10mLに変換し、125rpmで回転しながら37°C(8%CO2)で3〜4日間インキュベートする。収穫のために、遠心分離機培養10分1,000xgで、明確化された媒体を回収する。
- タンパク質Aとの親和性を介して上清中のIgG濃度を測定する。
- 酵素連動吸着アッセイ(ELISA)を用いて20μg/mLで各IgG(上清中)を、並べ替えに使用する個々のペプチドに対して試験し、ネガコントロールとしてストレプトアビジンプレートまたはアクチン上で捕捉する。ヒトIgG Fabに対するヤギの抗ヒトペルオキシダーゼ二次抗体を使用して、組換えクローンを検出します。背景を減算した後、OD > 0.5 は画面正として定義されます。
- 追加のELISAを行い、IgG上清の希釈を用いて1mL当たり20μgから始まる濃度曲線を生成し、初期スクリーンELISAで反応性を示す各抗原に対してODに対してプロットすることにより、スクリーン陽性ヒットを確認する。
結果
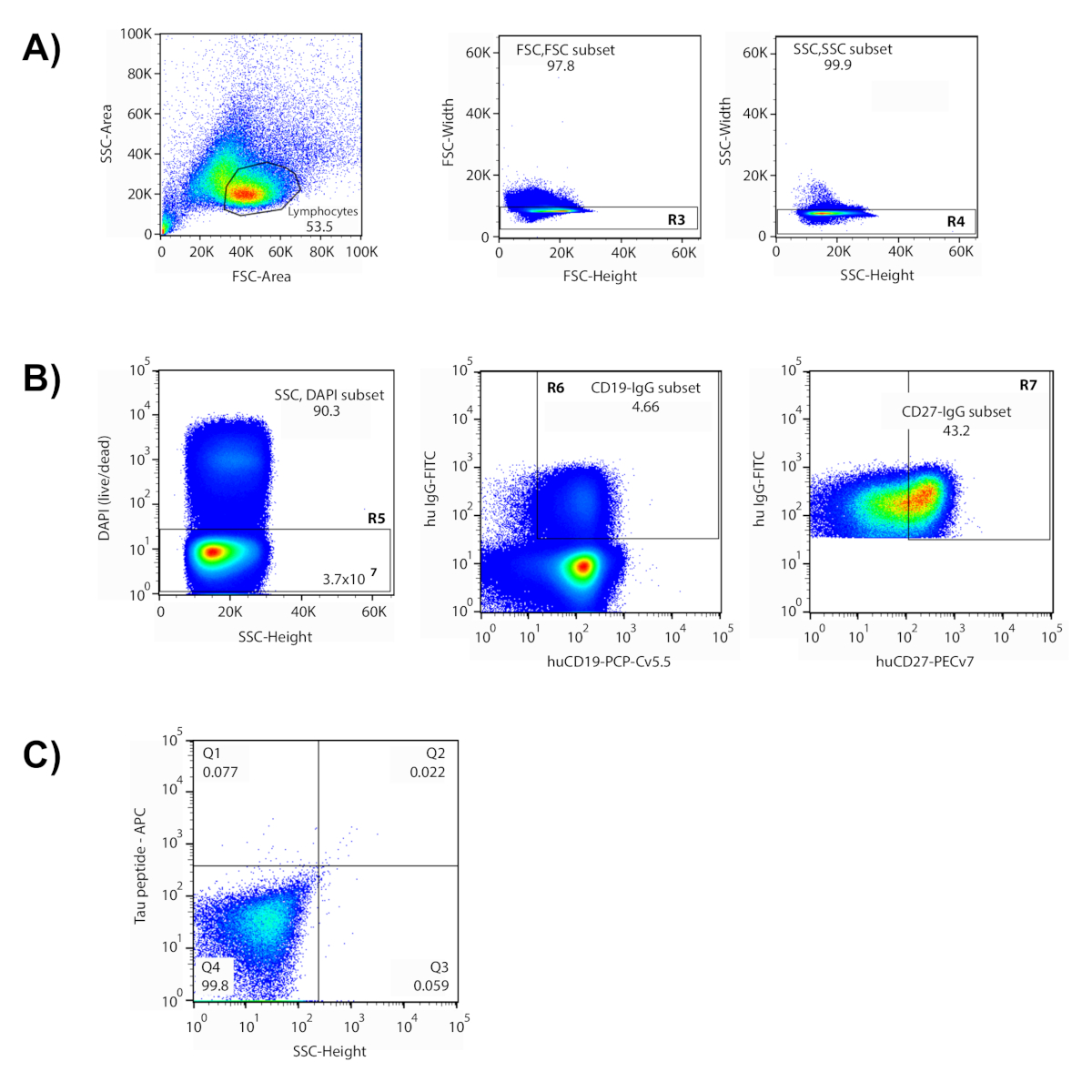
この方法は、ヒトドナーから抗原特異的抗体を単離するための多段階プロセスをカバーする。ここに示す代表的なデータでは、細胞を、基質リン酸化部位を模倣するリン酸化ペプチドを含むタウタンパク質のいくつかの異なるドメインを表す蛍光標識ペプチドのプールでインキュベートした。これらのペプチドは、目的とするタウエピトープで反応性のある細胞を同定するために「餌」として使用された。選別の準備として、蛍光標識型表マーカーのパネルを使用して、濃縮B細胞集団内の異なる細胞集団を同定した。一連のサイトメトリーゲートは、標的メモリB細胞を単離するために考案された(図1)。リンパ球は、流れ細胞メトリー6、7における前方散乱(FSC)および側散乱(SSC)プロットを用いて細胞サイズおよび粒度に基づいて単離された。複数の細胞(「ダブルト」)および死細胞を排除した後、発催マーカーはIgG、CD19(B細胞)およびCD27(メモリ)を介してIgG+メモリB細胞の分離を可能にした。このアプローチでは、CD27 マーカーは 2 つの離散母集団に分散しないため、CD27+発現セルの上位 45% が最終ゲートに含まれます。最後に、APCおよびPE蛍石孔の両方に対して二重陽性の細胞がゲーティンググラフの右上象限に分配され、ペプチドの両方の標識されたバージョンに対する反応性を示す。引き出されたゲート内に落ちた細胞を単離し、96ウェルプレートの個々の井戸に分類した。2つの異なる標識を持つ抗原の使用は、信号対雑音比を増加させ、その後の分子生物学プロセスにおける偽陽性の数を減少させる。ソートされた細胞は、最終ゲート内のメモリB細胞の約0.1%、および開始細胞サンプルの0.001%を表した。
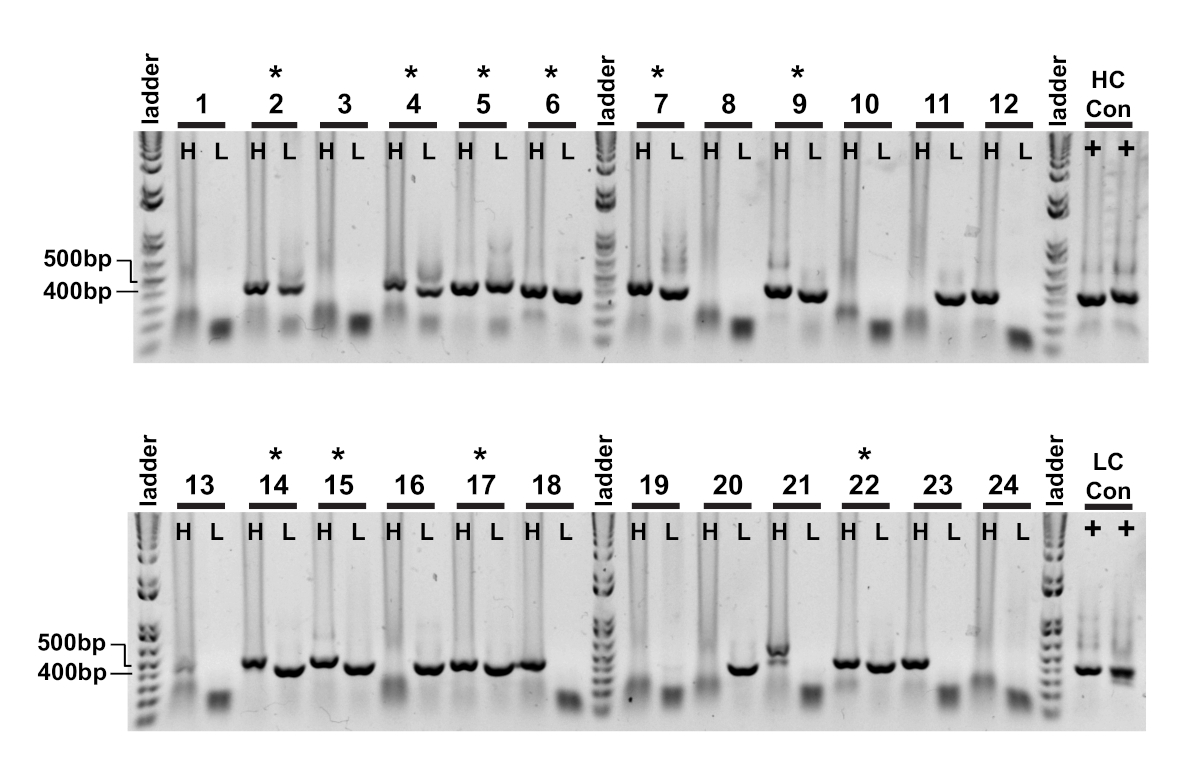
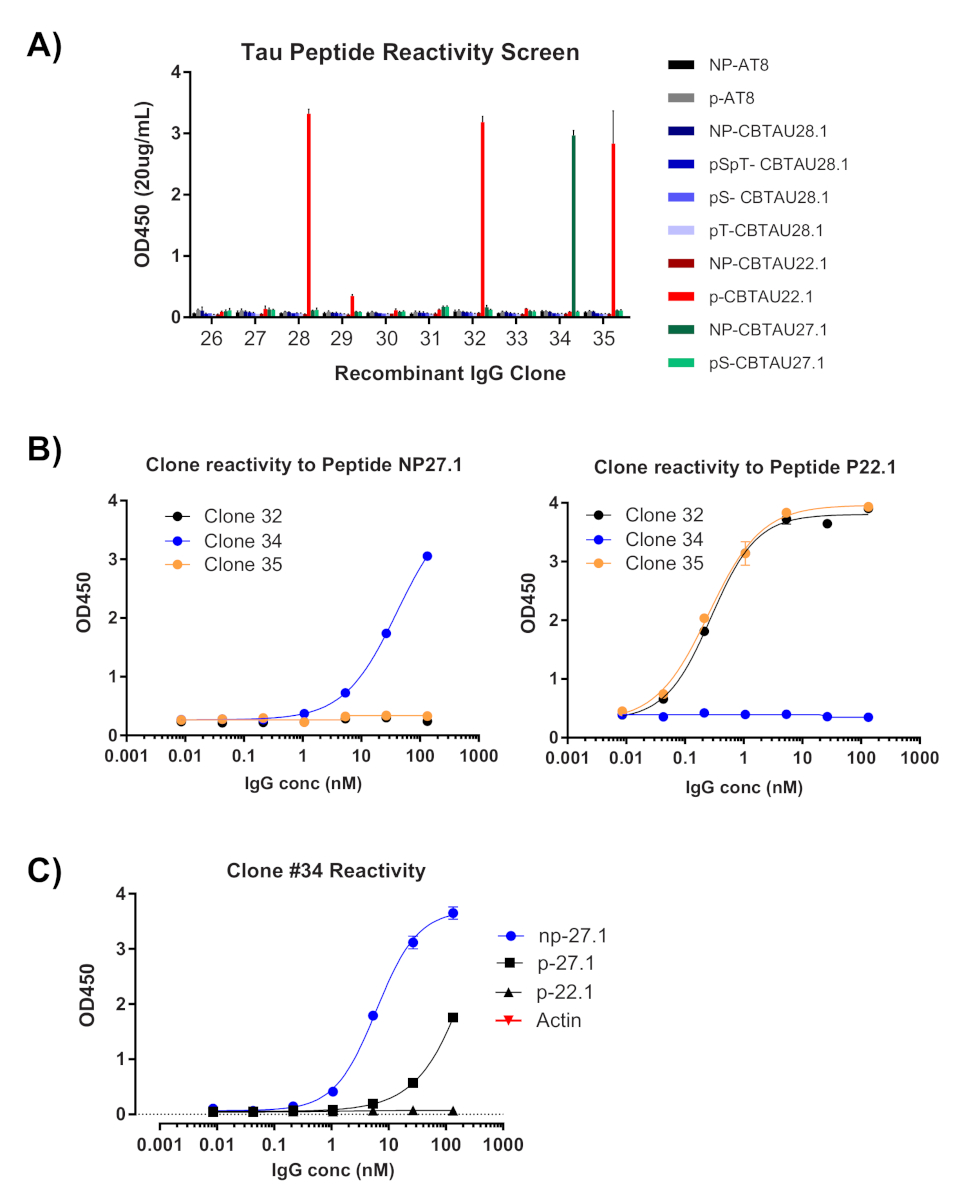
単一細胞クローニングの最初の読み出しは、それぞれの重い可変鎖と軽い可変鎖の増幅の確認である(図2)。両方のアンプリコンの対化回収が望ましいため、PCR反応はアガロースゲル上で並べて評価され、一致した対は抽出されたゲルおよびDNA断片から切除される。増幅の典型的な効率は30-50%の重鎖および50-70%の軽い(カッパ)鎖である。ペアアンプリコンの回収は、通常25〜40%の効率の間である。これらの効率はドナープール間で異なり、代表的なデータ(図3)は、24個の単一細胞からの非常に効率的な増幅の例である(42%対対回収)。IgGクローニングに続いて、IgG発現ベクターはヒト胚性腎臓(HEK-293)懸濁細胞にトランスフェクトされ、発現を最大化するために使用される。無血清培地の使用は、組換え抗体の準備からタンパク質を汚染することを低減し、その後の結合アッセイにおける騒音を最小限に抑えるのに役立ちます。回収された抗体は、タウペプチドの元のパネルに対してスクリーニングされ、ELISA(図4)によって反応性のためにスコア付けされた。バックグラウンド上のOD=0.5の初期閾値は、陽性抗原反応性を示すために使用され、β-アクチンタンパク質は非特異的結合の対照として使用される。フローサイトメトリーおよびソートステップがペプチドの混合プールを使用する場合、スクリーニングELISAは回収されたIgGの特定の反応をデコンボルトする最初のステップである。10 IgGアッセイのうち3つは、リン酸化CBTAU22.1ペプチドに対する反応性を示し、1つは非リン酸化CBTAU27.1(図4A)に反応した。追加の確認は、初期スクリーンで同定されたペプチドに対して同じ組換え抗体試料の濃度曲線を用いて完了し、および陰性対照として追加のペプチドを用いた(図4B)。陽性ヒットごとに、個々のプラスミドクローンを変換プールから分離し、同じELISA法によって再確認した。クローン34のみが提示されたデータを示し、非リン酸化CBTAU27.1に対する反応性が確認された一方で、リン酸化27.1ペプチドとの低い親和性結合も観察された(図4C)。

図 1: フローサイトメトリーによる抗原反応性単一細胞の単一細胞の単一の単一細胞の単離。(A) 図示は、リンパ球集団が引き出されたゲート内に含まれていた代表的なプロットである。(B) 前方および側面散乱に基づくゲート (FSC 高さ v FSC 幅 (R3);SSC-高さvSSC-幅(R4))を使用して二重線を除外した。描画されたゲート内のセルのみが、後続のプロットで評価されました。DAPI+セルは死んでいると見なされ、除外された(R5)。この実験では、3.7 x 10 7「生きている」細胞を尋問した。生細胞の大部分はB細胞(CD19+)、IgG+細胞(4.66%)この集団(R6)から隔離された。CD27+発現メモリセルの上位43.2%が選択ゲート(R7)に含まれていた。(C) クアドラント2(Q2)は、両標識抗原(ペプチド-APC vペプチド-PE)に反応する細胞を含み、これらの細胞を96ウェルプレートに個別にソートして回収した。この図のより大きなバージョンを表示するには、ここをクリックしてください。
{kind=link}

図 2: PCR 増幅は、クローニング用の IgG 重くて軽い可変シーケンスを回復します。(A) 重(VH)と光(VkまたはVl)鎖可変領域の両方が、ネストされたPCR反応を使用して回収されます。ステップ I プライマーは、ネイティブ リーダー シーケンスから前方に増幅し、一定の領域内から反転します。複数の矢印は、可能な生殖細胞の広範なカバレッジを確保するために使用される7-12プライマー間のプールを表します。ステップIIプライマーは、可変開読フレームの極端な端に固有のネストされ、発現ベクトル内の隣接する配列に相同であるアンプリコンの両端に配列を追加する。(B)リンカーは、定数軽鎖(CkまたはCl)から構成され、続いて重鎖プロモーターおよび非ネイティブシグナルペプチド配列が続く。アンプリコン、リンカー、およびプラスミドバックボーンは、ステップII PCR増幅中に生成された重なり合った相同配列を介して同時にライゲーションされ、無傷のプラスミドとして単離される。最終発現ベクターにおける組換え重鎖および軽鎖は、独立した(および同一の)CMVプロモーターによって駆動され、別個のタンパク質として翻訳される。この図のより大きなバージョンを表示するには、ここをクリックしてください。
{kind=link}

図 3:IgG重くて軽い可変鎖アンプリコンは単一細胞から回収される。ネストされたPCR反応(ステップII)を1%アガロースゲルに直接ロードし、可視化した。重鎖(H)とカッパ軽鎖(L)のPCR反応を隣接するウェルに積み込んだため、対アンプリコンが容易に観察された。成功した重鎖および軽鎖のプロダクトはそれぞれおよそ400 bpおよび350 bpである。対になったアンプリコンの正常な回復は、(*)で示されます。この図のより大きなバージョンを表示するには、ここをクリックしてください。
{kind=link}

図 4: 組換えIgGsから抗原特異性が確認される。回収された重鎖および軽鎖配列を含むプラスミドは、発現のためにHEK細胞にトランスフェクトされる。4日後、組換え抗体はELISAによって反応性について評価される。(A)明らかにされた培中のIgG濃度を測定し、20μg/mLに希釈した。選別に使用されるペプチドのプールを、ストレプトアビジンプレートのウェル当たり40pmolを用いて個別に評価した。すべてのIgGは、重複する井戸でテストされました。(B) OD450測定>0.5を表示したクローンを、画面上で同定されたペプチドに対して1:5希釈ステップを用いて再評価した(クローン#32、#34、#35)。(C) ヒットとして確認されると、単一のプラスミドクローンをクローンから単一のプラスミドクローンを#34形質転換ライゲーションから単離し、トランスフェクトし、ELISAによって再確認した。#32と#35のクローンについても同じことが行われました(データは表示されません)。この図のより大きなバージョンを表示するには、ここをクリックしてください。
{kind=link}
ディスカッション
ここで提示される方法は、フローサイトメトリーと単一細胞クローニングを組み合わせたものであり、ここで説明する方法は、Tillerと同僚によって以前に開発された方法に基づいています11.彼らの研究は、個々の細胞のレベルでヒトのB細胞レパートリーを研究するために、モノクローナル抗体の回収とクローニングについて説明する。我々は、多段階増幅プライマー戦略を含むメモリB細胞の集団からの抗原特異的モノクローナル抗体の回収を可能にするために、そのプロセスの主要な構成要素を適応させた。主な改変は、標識抗原「餌」の添加である。クローニングベクターバックボーンを単一の発現プラスミドに変更する(ただしこれらに限定されない)、生殖細胞系レパートリーの追加プライマーカバレッジ(リーダーシーケンスとフレームワーク1の両方)を含む、公開されたプロトコルに対する追加の適応がなされている。より高い発現のための懸濁液HEK293細胞のトランスフェクション、およびPCR増幅時の高忠実度ポリメラーゼの使用。
ここで説明する方法の場合、最も重要なステップは、フローサイトメトリーと単一細胞クローニングの間の遷移の近くにあります。まず、選別された単一細胞をプレートに適切に配置することが不可欠です。ソーターでドロップディレイを正しく設定することは重要なステップです。ターゲットのソートプレートで静的銃を使用しない限り、回収効率が大幅に低下する可能性があるため、湿度の低さなどの環境要因も考慮する必要があります。細胞の配置に続いて、プレートは、細胞がウェルの底部に10 μLのバッファーに接触していることを確認するために遠心分離されます。これらの対策はすべて、分子生物学の部分が成功するために重要です。単一細胞の逆転転写反応に対して条件が最適でない場合、β-アクチンのPCR増幅が困難でありうる。提示された方法の強さは、何百万もの細胞を尋問し、選択した抗原の抗原に対する抗原反応性基準に一致するもののみをソートする能力です。これには、信号対雑音比を可能な限り高くする必要があり、これはソート前に餌の濃度を最適化することによって行われます。二重標識餌は、蛍心の1つが高い背景を持っている場合に発生する可能性があり、偽陽性細胞の回復を減少させるために使用されます。複数の抗原餌を使用すると、シグナル:ノイズの最適化がより困難になりますが、複数のペプチドを同時に調知する能力は、この方法のもう一つの利点です。
回収効率が低い場合は、一連のネストされたβアクチンプライマーを使用して、テンプレートcDNAが存在することを確認します。このアプローチの欠点は、ウェルに細胞が存在しなかったか、逆転転写反応が失敗したのかを判断することが困難であり、トラブルシューティングが困難である。時折、逆が発生し、効率が予想よりも高くなります。重鎖の典型的な回復は30-45%の間であり、軽鎖は40-60%の間で高い。各PCR反応(ステップI&II)は、単一の細胞から増幅するために50サイクルを使用しています。総サイクル数が多いため、この方法は汚染の影響を受けやすくなります。アンプリコンの配列解析は、汚染物質が導入されたか、または異常に高い回収率が合法的に達成されたかどうかを判断するために使用できます。
選択した抗原に対して天然ヒト抗体を回収するこの方法の有用性には、いくつかの重要な制限がある。第1に、標識可能な可溶性タンパク質のみが、フローサイトメトリーの「餌」として使用されてもよい。代表的な結果に提示された種類については、タンパク質全体を使用することは困難であることが判明したので、タウタンパク質から一連の合成重複ペプチドを使用しました。線形ペプチドの使用は、非線形または構造エピトープに対する抗体の同定を制限する可能性があるため、最適ではない可能性が高い。しかし、我々はタウに対するいくつかのユニークな抗体を同定することができ、これらは現在、さらなる評価8,9を受けている。もう 1 つの制限は、IgG の分子生物学の回復とクローニングの低スループットです。最初のソート戦略は、数百万の細胞のスクリーニングを可能にしますが、その後の分子生物学処理は、複数の段階で構成されています。ギブソンアセンブリ10などの新しいテクノロジーを組み込むための継続的な最適化の取り組みは、いくつかのステップを合理化しましたが、ボトルネックは個々の重鎖と軽鎖のペアのクローンを作成したままです。
「BSelex」法は、記憶B細胞の表面に発現したBCRを利用して抗原反応性を示す細胞を同定し、次いでフローサイトメトリー11、12を介してこれらの個々の細胞を回収する。このBCRへの依存により、この方法はメモリB細胞コンパートメントに限定され、プラズマブラストなどの抗体分泌細胞(ASC)を捕捉しません。しかしながら、このアプローチは、ASCと比較した場合、抗原特異的免疫グロビンのより広いレパートリーを回復するのに有利であり得る。インフルエンザワクチン接種研究は、反応性ASCが少数の膨張B細胞クローンによって支配されうるが、抗原特異的記憶B細胞集団がクローン12を有することはめったにないものを示している。T細胞表面のT細胞受容体(TCR)は、多様性を最大化するために遺伝子配置および組み換えにおいてB細胞受容体と類似性を有する。本書に記載されている方法に記載されているものと同様の単一細胞アプローチは、αおよびβ鎖13の回収および単一細胞クローニングを含むT細胞レパートリーを評価するために開発された。しかし、TCR認識はMHC分子によって提示されるペプチドを必要とし、ペプチド特異的T細胞を同定する標識された餌のアプローチに重要な複雑さを加える。B細胞とは異なり、T細胞は可変領域全体の親和性成熟を受けないため、短いCDR3領域といくつかの側面配列の同定は、同定と再構成に必要なすべてです。最後に、この方法は、フローサイトメトリーを用いたIgG分子の同定を記述するが、異なるアイソタイプのB細胞を同定するために代替表現型マーカーを使用することも可能である。
現在、次世代シーケンシングから収集した膨大なデータを抗原特異性の機能解析と結びつける方法が開発されていますが、その方法は未だ洗練されています。その限界にもかかわらず、ここで説明する方法は、感染性および非感染性疾患8、14、15の両方の潜在的な治療価値を有する抗体を同定するために使用され、信頼性の高いB細胞や広範な細胞培養の広範な操作なしに、ヒトから関連する抗原特異的IgGを回復するためのアプローチ。
開示事項
著者は開示するものが何もない
謝辞
著者らは、ルーシー・チャマス、マーサ・コスタ、ジュリー・キム、ナンシー・エレディア、ジェレミー・マセドに、現在のBSelexプラットフォームの多くの方法の変更と改良の広範なテストに感謝したいと思います。
資料
| Name | Company | Catalog Number | Comments |
| MoFlo Astrio EQ, Cell Sorter | MoFlo | cell sorting | |
| Biotinylated peptides | New England Peptide | Cell sorting "bait" | |
| Micro Bio-spin P30 gel column | BioRad | #7326223 | |
| Streptavidin (R-PE) | Thermo Scientific | SA10041 | |
| Streptavidin (APC) | Thermo Scientific | S32362 | |
| CD22 MicroBeads, human | Miltenyi | #130-046-401 | |
| LS columns | Miltenyi | #130-042-401 | |
| Pre separation filters | Miltenyi | #130-041-407 | |
| PerCp-Cy5.5 mouse anti-human CD19 | BD Biosciences | BD#340951 | |
| PE-Cy7 mouse anti-human CD27 | BD Biosciences | #560609 | |
| FITC mouse anti-human IgG | BD Biosciences | #560952 | |
| DAPI | Life Technologies | #D21490 | |
| RNaseOUT | Life Technologies | #10777-019 | |
| Bovine serum albumin, Fraction V | Sigma | #A4503 | |
| RPMI media | Hyclone | #SH30096.01 | |
| HI FBS (Fetal bovine serum) | Invitrogen | #10082147 | |
| Mastercycler Gradient | Eppendorf | Model #6325 | PCR machine |
| PCR 96-well plates | Phenix | MPS-500 | |
| PCR Plate mats | Phenix | SMX-PCR96 | |
| Advantage UltraPure PCR Deoxynucleotide Mix | Clontech | #639125 | |
| Superscript IV First-strand synthesis system | Invitrogen | #18091050 | |
| Phusion High Fidelity Polymerase | Thermo Scientific | F-531-L | |
| Platinum polymerase | Invitrogen | #10966108 | |
| Nuclease-free water | QIAGEN | #129114 | |
| Oligonucleotide primers | IDT | assorted | Primers for all PCR steps |
| Gel Extraction Kit | QIAGEN | #28706 | |
| PCR Purification kit | QIAGEN | #28106 | |
| Miniprep Kit | QIAGEN | #27106 | |
| NEBuilder HiFi DNA Assembly Cloning Kit | New England Biolabs | E5520S | Gibson assembly |
| Expi293 Expression Media | Invitrogen | #A14351-01 | |
| ExpiFectamine 293 Transfection Kit | Invitrogen | #A14525 | |
| Opti-MEM Media | Invitrogen | #31985070 | |
| CO2 incubator (Multitron) | |||
| Octet RED384 System | Pall/ Forte Bio | ||
| Pierce Streptavidin Coated High Binding Capacity Clear 96-well Plates | Thermo Scientific | #15500 | |
| Corning Costar 96-well Polystyrene Plate High Binding Half Area | Corning | #3690 | |
| Goat Anti-human IgG Fab Secondary Antibody | Jackson Labs | #109-036-097 | |
| Goat Anti-mouse HRP Secondary Antibody | Jackson Labs | #115-035-072 | |
| SureBlueTM TMB Microwell Peroxidase Substrate (1-Component) | KPL | #52-00-03 | |
| TMB Stop Solution | KPL | #50-85-06 |
参考文献
- Su, K. Y., Watanabe, A., Yeh, C. H., Kelsoe, G., Kuraoka, M. Efficient Culture of Human Naive and Memory B Cells for Use as APCs. Journal of Immunology. 197 (10), 4163-4176 (2016).
- Corti, D., Lanzavecchia, A. Broadly neutralizing antiviral antibodies. Annual Review of Immunology. 31, 705-742 (2013).
- Steinitz, M., Klein, G., Koskimies, S., Makel, O. EB virus-induced B lymphocyte cell lines producing specific antibody. Nature. 269 (5627), 420-422 (1977).
- Bloom, A. D., Nakamura, F. T. Establishment of a tetraploid, immunoglobulin-producing cell line from the hybridization of two human lymphocyte lines. Proceedings of the National Academy of Sciences of the United States of America. 71 (7), 2689-2692 (1974).
- Olsson, L., Kaplan, H. S. Human-human hybridomas producing monoclonal antibodies of predefined antigenic specificity. Proceedings of the National Academy of Sciences of the United States of America. 77 (9), 5429-5431 (1980).
- Lechner, J., et al. Alterations in Circulating Immune Cells in Neovascular Age-Related Macular Degeneration. Scientific Reports. 5, 16754(2015).
- Loken, M. R., Brosnan, J. M., Bach, B. A., Ault, K. A. Establishing optimal lymphocyte gates for immunophenotyping by flow cytometry. Cytometry. 11 (4), 453-459 (1990).
- Pascual, G., et al. Immunological memory to hyperphosphorylated tau in asymptomatic individuals. Acta Neuropathologica. 133 (5), 767-783 (2017).
- van Ameijde, J., et al. Enhancement of therapeutic potential of a naturally occurring human antibody targeting a phosphorylated Ser(422) containing epitope on pathological tau. Acta Neuropathologica Communications. 6 (1), 59(2018).
- Gibson, D. G., et al. Enzymatic assembly of DNA molecules up to several hundred kilobases. Nature Methods. 6 (5), 343-345 (2009).
- Tiller, T., et al. Efficient generation of monoclonal antibodies from single human B cells by single cell RT-PCR and expression vector cloning. Journal of Immunological Methods. 329 (1-2), 112-124 (2008).
- Wrammert, J., et al. Rapid cloning of high-affinity human monoclonal antibodies against influenza virus. Nature. 453 (7195), 667-671 (2008).
- Han, A., Glanville, J., Hansmann, L., Davis, M. M. Linking T-cell receptor sequence to functional phenotype at the single-cell level. Nature Biotechnology. 32 (7), 684-692 (2014).
- Apetri, A., et al. A common antigenic motif recognized by naturally occurring human VH5-51/VL4-1 anti-tau antibodies with distinct functionalities. Acta Neuropathologica Communications. 6 (1), 43(2018).
- Jones, H. G., et al. Structural basis for recognition of the central conserved region of RSV G by neutralizing human antibodies. PLoS Pathogens. 14 (3), 1006935(2018).
- Janssen Vaccines & Prevention B.B. US patent. , U.S. Patent Application No. 20170210787, Published July 27, 2017 (2017).
転載および許可
このJoVE論文のテキスト又は図を再利用するための許可を申請します
許可を申請This article has been published
Video Coming Soon
Copyright © 2023 MyJoVE Corporation. All rights reserved