
Method Article
Método de triagem de célula única para a seleção e recuperação de anticorpos com especificidades desejadas de populações de células B de memória humana enriquecida
Neste Artigo
Resumo
O método de BSelex para identificar e recuperar anticorpos antígeno-específicos individuais das pilhas mononucleares do sangue periférico humano combina a citometria do fluxo com o único PCR da pilha e a clonagem.
Resumo
O repertório do anticorpo humano representa uma fonte pela maior parte inexplorada de anticorpos terapêuticos potenciais e de Biomarkers úteis. Embora os métodos computacionais atuais, como o sequenciamento da próxima geração (NGS), produzam enormes conjuntos de dados no repertório de anticorpos no nível de sequência, os dados funcionais são necessários para identificar quais sequências são relevantes para um determinado antígeno ou conjunto de Antígenos. Aqui, nós descrevemos um método para identificar e recuperar anticorpos antígeno-específicos individuais das pilhas mononucleares do sangue periférico (PBMCs) de um doador de sangue humano. Este método utiliza um enriquecimento inicial de pilhas B maduras e exige uma combinação de marcadores fenotípicos da pilha e de proteína fluorescently-etiquetada para isolar pilhas da memória B de IgG através da citometria do fluxo. As regiões variáveis de cadeia pesada e leve são então clonadas e reexaminadas. Embora limitado ao compartimento da pilha da memória B, este método aproveita-se da citometria do fluxo para interrogar milhões de pilhas de B e retorna seqüências pesadas e claras emparelhadas da corrente de uma única pilha em um formato pronto para a expressão e a confirmação da especificidade. Os anticorpos recuperados com este método podem ser considerados para o potencial terapêutico, mas podem igualmente lig a especificidade e a função com as aproximações Bioinformatic para avaliar o repertório da pilha de B dentro dos indivíduos.
Introdução
Os anticorpos são uma classe crescente de moléculas terapêuticas, e o repertório existente da pilha de B em todo o ser humano é uma fonte potencial de tais anticorpos. Quando recuperados de um doador humano, eles não necessitam de adaptação ou "humanização", passos que são necessários para os anticorpos gerados em outros sistemas animais. Existem vários métodos para a identificação e isolamento de anticorpos humanos, incluindo a ativação e proliferação de célulasB1, imortalização via EBV transformação2,3e geração de linhas de células de hibridoma4 ,5. Entretanto, todos estes métodos exigem a cultura de pilha extensiva de tela e de recuperar anticorpos antígeno-específicos. A informação sobre o repertório do anticorpo humano foi expandida extremamente com o desenvolvimento da tecnologia de sequenciamento da próxima geração (NGS), permitindo a identificação de quantidades maciças de seqüências individuais atuais nas amostras do doador. No entanto, como a NGS produz uma visão agnóstica de todas as sequências presentes, não permite a identificação e o isolamento de anticorpos antígeno-específicos, especialmente no caso de anticorpos raros ou de baixa frequência.
O objetivo do método "BSelex" é identificar anticorpos antígeno-específicos de células mononucleares do sangue periférico circulante em doadores humanos, e isolar e recuperar as sequências desses anticorpos para posterior análise. Este método utiliza a citometria de fluxo e a classificação da pilha para tirar proveito do receptor da célula B (BCR) expressado na superfície de pilhas da memória B. Os milhões de pilhas de B podem ser selecionados para a antígeno-especificidade através da citometria do fluxo antes que os métodos mais low-throughput da biologia molecular sejam iniciados. A identificação Chain pesada e clara emparelhada não é possível em a maioria de métodos de NGS, que analisam seqüências da pilha no volume. No método que nós descrevemos aqui, as pilhas são isoladas individualmente, e a recuperação emparelhada de seqüências pesadas e claras da corrente é possível, que permite a clonagem direta e a expressão do IgG cheio.
Protocolo
O uso de amostras de voluntários humanos seguiu protocolos aprovados pela Diretoria de revisão institucional do Scripps Research Institute. O consentimento informado foi obtido dos doadores antes da doação de sangue.
1. preparação do reagente
-
Células mononucleares do sangue periférico (PBMCs)
- Recupere pilhas mononucleares do sangue periférico dos doadores humanos normais pelo isolamento de Ficoll-Plaque Plus. Cryopreserve em 50 milhões Cells/ml em 90% soro bovino fetal (FBS) e 10% dimetil sulfóxido (DMSO). Congele as pilhas em-80 ° c e transfira ao nitrogênio líquido para o armazenamento a longo prazo.
-
Rotulando peptídeos de antígeno alvo para classificação
- Gerar ou obter peptídeos específicos para a proteína alvo (até 70 resíduos de comprimento) com uma biotina terminal.
- Etiqueta 4 nmol de cada peptídeo biotinylated individual com estreptavidina covalentemente Unido ao PE (R-phycoerythrin) ou ao APC (allophycocyanin) em tubos separados. Prepare-se usando uma relação molar de 9:1 peptídeo para estreptavidina (SA), com uma concentração final de aproximadamente 3,2 μM para SA-PE e 5,7 μM SA-APC. Prepare os tetrâmeros da biotina como um controle negativo.
- Incubar cada mistura peptídica durante a noite, no escuro, a 4 ° c com mistura lenta.
- Remova o fluoróforo não acoplado passando peptídeos rotulados através de microcolunas contendo grânulos de poliacrilamida.
- Armazene os tetrâmeros do peptide até 2 meses em 4 ° c.
2. classificação de célula
- Pelo menos 1 h até 16 h antes do isolamento CD22 +, descongelar PBMCs doadores e permitir descansar a 37 ° c.
- Retire os frascos de PBMCs congelados do congelador e transfira imediatamente para um banho de água de 37 ° c. Quando os frascos estão quase descongelados, transfira para a área de trabalho.
- Transfira o conteúdo de cada frasco para injetáveis para um tubo de 50 mL contendo um meio pré-aquecido (mantendo os doadores separados). Adicionar meio a um volume final de 50 mL. Misture suavemente o conteúdo.
- Centrifugar a 370 x g durante 6 min à temperatura ambiente. Descarte o sobrenadante, resuma as células em 15 mL de meio completo de RPMI e realize uma contagem de células.
- Ajuste a concentração celular para 2,0 x 107 células/ml com meio RPMI completo, e transfira células para um balão T75 ou maior e incubar a 37 ° c por pelo menos 1 h e até 16 h para permitir o tempo de recuperação para as células descongelados.
- Recolher as células (pooling, se desejado) e centrifugar a 370 x g por 6 min a 4 ° c. Descarte o sobrenadante e refaça as células em 5 mL de buffer de MACS com gelo frio (PBS, pH 7,6 contendo 0,5% de BSA e 2 mM de EDTA), leve para 50 mL com buffer de MACS e realize uma contagem de células.
- Centrifugue as células a 400 x g durante 7 min a 4 ° c.
- Isole as células CD22+ B por seleção positiva via captura em MICROESFERAS CD22. Use o buffer de MACS para todas as lavas. Contagem e centrifugação 400 x g por 7 min a 4 ° c. A recuperação esperada é de 5-10% da população total de PBMC.
- Reressuscitem as células em 40 milhões por mL de tampão FACS (tampão Tris, pH 8,0 contendo 0,5% de BSA e 2 mM de EDTA) e proceder com coloração celular dos doadores individualmente ou como uma piscina.
- Células de mancha para a triagem de célula B de memória
- Retire uma alíquota de células para o citometro configurado e compensação para cada um dos fluoróforos de rotulagem utilizados. Inclua células não manchadas como um controle.
- Calcule o volume de coloração final para o número de células CD22+ obtidas (coloração 2 milhões por 100 μL).
- Adicione marcadores extracelulares para células B (CD19-PerCP-Cy 5.5), IgG (IgG-FITC) e compartimento de memória (CD27-PECy7) de acordo com as recomendações de diluição dos fabricantes e misture suavemente. Aliquot 107 células num tubo de 1,5 ml para o controlo negativo e transfira as restantes células para um tubo de 15 ml.
- Adicione os tetrâmeros de biotina-SA rotulados duplos ao tubo de controle negativo em uma concentração final de 36 nm cada para PE e APC multiplicado pelo número de peptídeos no tubo de ordenação e trazer o volume para 0,5 ml final com tampão FACS (por exemplo, 10 peptídeos x 36 nm = 360 nm).
- Adicione os tetrâmeros do peptide ao tubo da sorte em 36 nanômetro cada para o PE e o APC e traga as pilhas a 2x106 por 100 μl com amortecedor de TBS. Incubar a 4 ° c no escuro e com rotação suave por 30-60 min.
- Lave 2x a 4 ° c, 400 x g, 7 min, removendo uma alíquota para contar antes da última lavagem. Filtre células usando tubos de tampão de filtro de 5 mL. Adicionar DAPI (4 ', 6-diamidino-2-phenylindole) mancha para 0,3 μM concentração final antes de classificar como um marcador de integridade da membrana celular.
- Triagem de célula única via citometria de fluxo
- Prepare 48 poços de placas do PCR 96-well para classificar.
- Prepare uma mistura mestra: (2 μL de tampão de 10x RT, 0,5 μL de inibidor de RNase, 7,5 μL de água estéril da PCR-classe) por a amostra. Aliquot 10 μL por poço, cubra e armazene a 4 ° c até estar pronto para a ordenação.
- Execute o controle negativo no classificador de célula, analisando o exemplo inteiro.
- Definir portões de citometria de fluxo para isolar as populações de células apropriadas para a triagem de célula única.
- Plotar área FSC vs área SSC e definir portão R1 para isolar linfócitos.
- Plotar a altura FSC versus a largura do FSC e configurar o Gate R3 para excluir os doublets do FSC.
- Plotar altura SSC vs largura SSC e definir portão R4 para excluir doublets SSC.
- Plotar a altura SSC vs DAPI e definir o portão R5 para isolar células ao vivo (DAPI-).
- Plotar IgG vs CD19 e definir o portão R6 para isolar as células IgG+ B (CD19+ IgG+).
- Plotar IgG vs CD27 e configurar o portão R7 para selecionar células B de memória (CD27alta).
- Plotar Antigen-PE (AG-PE) vs Antigen-APC (AG-APC) e definir portão R8 como um AG-PE+/AG-APC+ quadrante com um baixo número de eventos no portão duplo positivo.
- Definir portões de citometria de fluxo para isolar as populações de células apropriadas para a triagem de célula única.
- Colete um número igual de células de memória da amostra antígeno-positiva (AG+) para a determinação do sinal ao ruído.
- Classifique únicas pilhas que têm o phenotype CD19+ CD27+ AG-PE+ AG-APC+ (porta R8) em placas preparadas do PCR de 96-well.
- Cubra as placas com as almofadas de fita de alumínio, centrifugue por 1 minuto em 400 x g, e armazene-o em-80 ° c para a clonagem futura.
- Prepare 48 poços de placas do PCR 96-well para classificar.
3. clonagem de célula única
-
Reações de transcrição reversa
- Remova a placa de classificação de célula B única de 80 ° c. Descongelar no gelo por 2 min. Gire a placa por 2 min em 3.300 x g para o conteúdo da piscina na parte inferior dos poços antes de abrir.
- Prepare um Mix mestre dNTP/buffer com 10% de detergente não iônico e oligo (dT) como primer reverso. Adicione a mistura da enzima a cada poço que contem a única pilha e não misture.
- Incubar 5 min a 65 ° c. Adicione a mistura mestre da enzima contendo 100 mM DTT, inibidor de RNase, e enzima reversa da transcriptase para trazer o volume total a 20 μL. Incubar 10 min a 50 ° c, depois 10 min a 80 ° c.
- Transferência ao gelo antes de começar etapas do PCR
-
Reações de PCR aninhadas em várias etapas
- As reações aninhadas separadas do PCR do uso são usadas para a corrente pesada (HC) e a amplificação da corrente clara (LC).
- Para a primeira rodada de amplificação de PCR (Step I), use um conjunto de primers para a frente e uma única cartilha reversa para amplificar as regiões variáveis HC e LC em reações de PCR separadas (Figura 2). Pelo projeto, o pool de primers para diante amplificará uma grande porcentagem de seqüências do líder do germline (L) e a primeira demão reversa é específica às regiões constantes a jusante de cada corrente, incluindo o Kappa (κ) e o lambda (λ) para correntes claras16.
- Use 2,5 μL do produto do cDNA como o molde para cada reação do PCR (etapa I). Adicionar primers à concentração de reacção final de 1 μM para cada um. Dobre o amortecedor da polimerase 10x à concentração 2x, traga o volume final a 25 μL por a reação. Executar reações de PCR HC usando [94 ° c/4 min; 94 ° c/15 s, 55 ° c/20 s, 68 ° c/60 s; 68 ° c/3 min] para 50 ciclos. Executar reações de PCR LC usando [98 ° c/4 min; 98 ° c/15 s, 72 ° c/20 s, 72 ° c/60 s; 72 ° c/3 min] para 50 ciclos.
- Use 2,5 μL do produto Step I como modelo para cada reação de PCR (passo II). Adicione o pool de primers Forward específicos para a região do quadro 1 de HC e LC (κ e λ) e um conjunto de primers reversos específicos para a região de junção de cada cadeia de anticorpos. Dobre o amortecedor da polimerase 10x à concentração 2x, e execute reações do PCR 50 ciclos cada um usando os mesmos parâmetros que etapa I.
- Execute as reações de PCR concluídas em gel de agarose a 1% para visualizar acertos positivos de amplificação. Recupere amplicões emparelhados (corrente pesada e clara da mesma pilha) e isole através da extração do gel. Determine a concentração do fragmento do ADN através do OD260 para cálculos exatos da mistura da ligadura.
- Combine fragmentos de cadeia pesada e leve recuperados com um fragmento de vinculador e a espinha dorsal do vetor de expressão usando uma reação de ligadura de 4 fragmentos usando um kit comercial.
- Transforme reações de ligadura em bactérias quimicamente competentes usando um kit comercial. Uma vez chapeado em placas antibióticas, adicionar 4 mL de meios de crescimento (com antibiótico) para a cultura de transformação remanescente, e incubar 37 ° c durante a noite em 250 rpm. Esta é a "cultura da mistura de ligadura".
- Prepare o ADN do protocolo das culturas da mistura da ligadura da noite usando um jogo comercial, e determine a concentração resultante do ADN do plasmídeo.
4. tela ELISA para confirmação de antígeno-especificidade
-
Transfect 10 μg de DNA protocolo com reagente à base de lipídios catiônico em 10 ml de células de suspensão 293, e incubar por 3-4 dias a 37 ° c (8% co2) enquanto gira a 125 RPM. Para a colheita, centrifugue a cultura 10 minutos em 1.000 x g e recupere meios esclarecidos.
- Meça a concentração de IgG nos sobrenadantes através da afinidade à proteína A.
- Teste cada IgG (em sobrenadante) a 20 μg/mL por ensaio de adsorvente enzimático (ELISA) contra os peptídeos individuais utilizados para a espécie, capturados em uma placa de estreptavidina ou actina como um controle negativo. Use um anticorpo secundário da peroxidase antihumana da cabra de encontro ao IgG humano Fab para detectar clones recombinantes. Após a subtração do fundo, o OD > 0,5 é definido como o positivo da tela.
- Confirme acertos positivos da tela realizando um ELISA adicional, usando diluições de sobrenadante de IgG para gerar uma curva de concentração a partir de 20 μg por mL, e plotando contra OD para cada antígeno mostrando reatividade na tela inicial ELISA.
Resultados
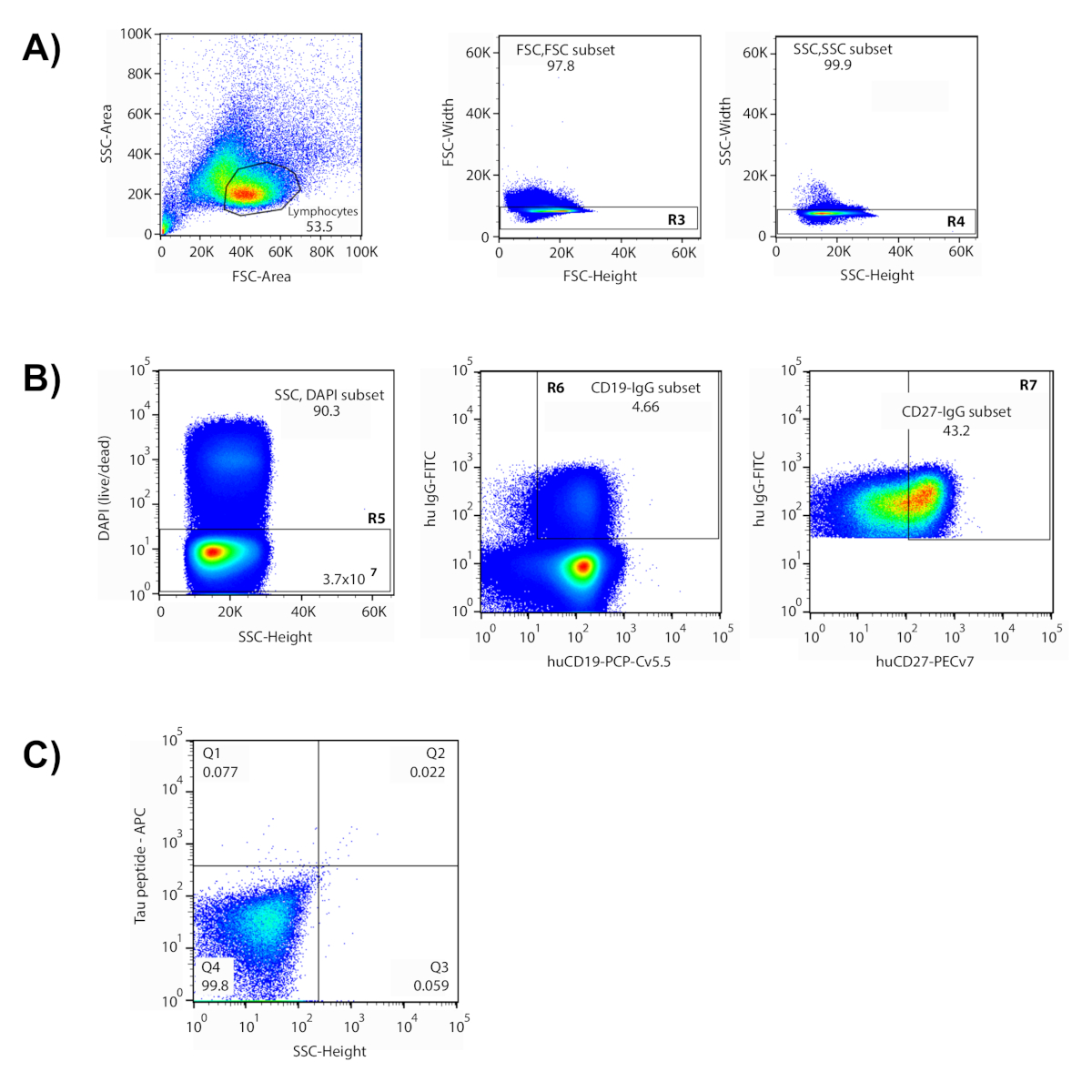
Este método abrange um processo de várias etapas para isolar anticorpos antígeno-específicos de doadores humanos. Nos dados representativos mostrados aqui, as células foram incubadas com um conjunto de peptídeos rotulados com fluorescently representando vários domínios diferentes da proteína Tau, incluindo peptídeos fosforilados para imitar sítios putativos de fosforilação. Esses peptídeos foram utilizados como "isca" para identificar células que são reativas com epítopos de Tau (s) de interesse. Na preparação para a classificação, um painel de marcadores fenotípicos fluorescently-etiquetados foi usado para identificar populações diferentes da pilha dentro da população enriquecida da pilha de B. Uma série de portões de citometria foi concebida para isolar as células B da memória alvo (Figura 1). Os linfócitos foram isolados com base no tamanho e granularidade das células, utilizando-se parcelas de dispersão direta (FSC) e dispersão lateral (SSC) em citometria de fluxo6,7. Depois da exclusão de pilhas múltiplas ("doublets") e de pilhas inoperantes, os marcadores fenotípicos permitiram a segregação de pilhas de IgG+ de memória b através de IgG, de CD19 (pilha de b) e de CD27 (memória). Nesta aproximação, o marcador CD27 não distribui em duas populações discretas, assim que os 45% superiores de CD27+ que expressam pilhas são incluídos para a porta final. Finalmente, as células de duplo-positivo para os fluoróforos APC e PE distribuir no quadrante superior direito do gráfico de gating, indicando reatividade para ambas as versões rotuladas de peptídeos. As pilhas que caíram dentro da porta desenhada foram isoladas e classificadas em poços individuais de uma placa 96-well. O uso de antígeno com duas etiquetas diferentes aumenta a relação sinal-ruído e reduz o número de falsos positivos no processo de biologia molecular subsequente. As células classificadas representaram cerca de 0,1% das células B da memória no portão final e 0, 1% da amostra da célula inicial.
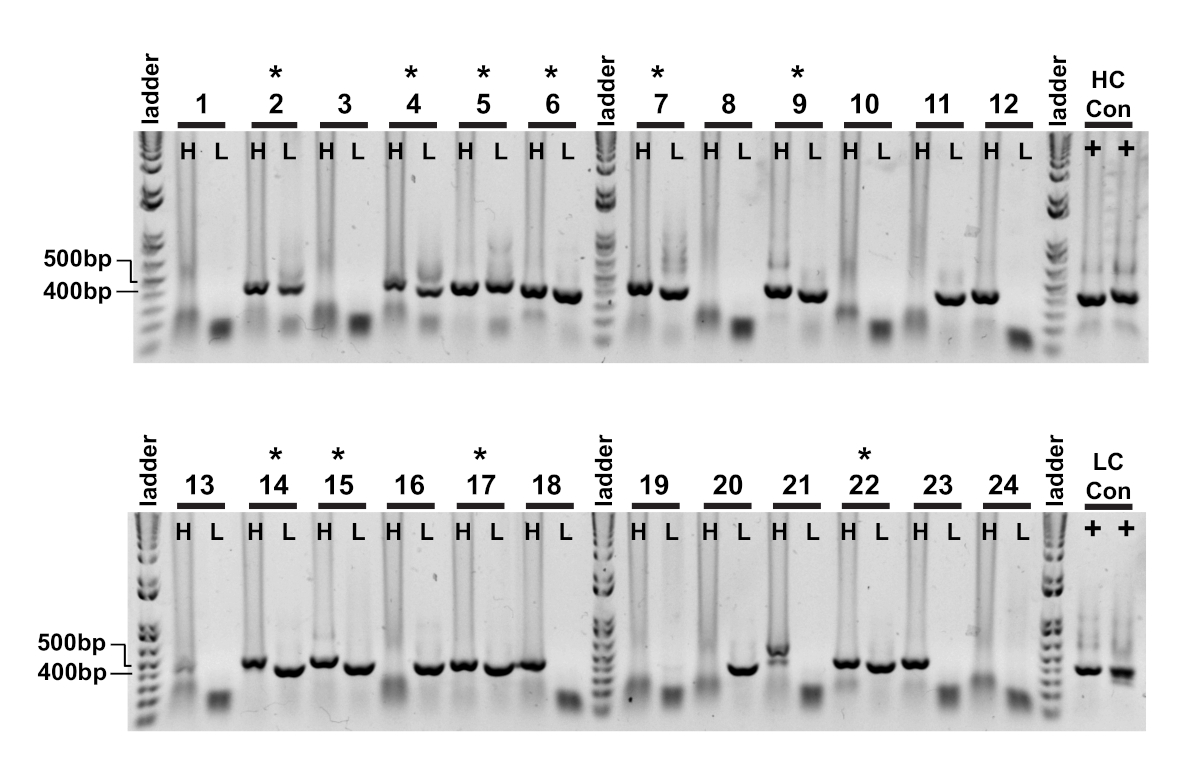
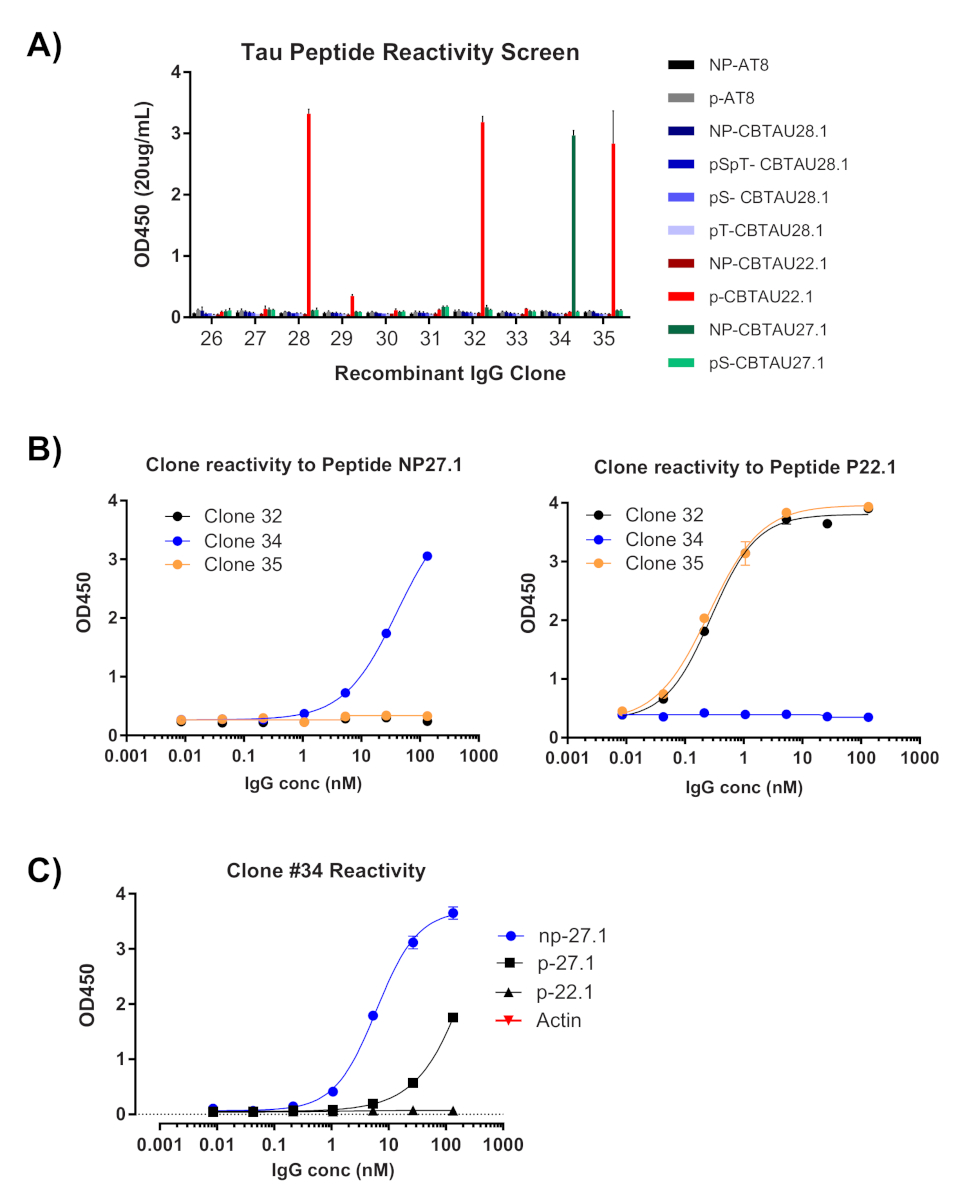
A primeira leitura da clonagem de uma única célula é a confirmação da amplificação das respectivas cadeias variáveis pesadas e leves (Figura 2). Desde que a recuperação emparelhada de ambos os amplicões é desejada, as reações do PCR são avaliadas lado a lado no gel do agarose, e os pares combinados são extirpado dos fragmentos do gel e do ADN extraídos. A eficiência típica da amplificação é a corrente pesada de 30-50% e a corrente da luz 50-70% (Kappa). A recuperação de amplicões emparelhados é geralmente entre 25-40% de eficiência. Essas eficiências variam entre os pools de doadores e os dados representativos (Figura 3) são um exemplo de amplificação muito eficiente de 24 células individuais (42% de recuperação pareada). Após a clonagem de IgG, o vetor de expressão de IgG é transfected em células de suspensão do rim embrionário humano (HEK-293), que são usadas para maximizar a expressão. O uso de meio livre de soro reduz as proteínas contaminantes da preparação do anticorpo recombinante e ajuda a minimizar o ruído em ensaios de ligação subsequentes. Os anticorpos recuperados são rastreados contra o painel original de peptídeos Tau e marcados para reatividade por ELISA (Figura 4). Um limiar inicial de OD = 0,5 acima do fundo é usado para indicar a reactividade positiva do antígeno, e a proteína do β-Actin é usada como um controle para a ligação não específica. Se a citometria de fluxo e as etapas de triagem usarem um conjunto misto de peptídeos, a triagem ELISA é o primeiro passo de reatividades específicas deconvoluting dos IgGs recuperados. Três dos 10 IgGs analisados apresentaram reatividade contra peptídeo de CBTAU 22,1 fosforilado e um foi reativo para CBTAU 27,1 não fosforilado (Figura 4a). A confirmação adicional foi completada por meio de uma curva de concentração das mesmas amostras de anticorpos recombinantes contra os peptídeos identificados na primeira tela, e um peptídeo adicional como controle negativo (Figura 4B). Para cada acerto positivo, um clone de plasmídeo individual foi isolado da piscina transformada e reconfirmado pelo mesmo método ELISA. Somente o clone 34 é mostrado os dados apresentados, e quando a reactividade ao cbtau 27.1 não-fosforilada foi confirmada, uma ligação mais baixa da afinidade foi observada igualmente com o peptide 27,1 fosforilated (Figura 4C).

Figura 1 : Isolação de pilhas únicas antígeno-reactivas através da citometria do fluxo. (A) mostrado é um gráfico representativo onde a população de linfócitos estava contida dentro do portão desenhado. (B) portões com base no espalhamento dianteiro e lateral (FSC-altura v FSC-Width (R3); SSC-altura v SSC-Width (R4)) foram usados para excluir doublets. Somente as células dentro das portas desenhadas foram avaliadas em parcelas subsequentes. As células DAPI+ foram consideradas mortas e excluídas (R5). Neste experimento, 3,7 x 107 células "ao vivo" foram interrogadas. A maioria de pilhas vivos era pilhas de B (CD19+), e as pilhas de IgG+ (4,66%) foram isolados desta população (R6). As principais 43,2% das células de memória expressando CD27+foram incluídas no portão de seleção (R7). (C) quadrante 2 (Q2) continha células reativas para ambos os antígenos rotulados (PEPTÍDEO-APC v PEPTÍDEO-PE), e essas células foram classificadas e recuperadas individualmente em uma placa de 96 poços. Por favor clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 2 : A amplificação do PCR recupera a seqüência variável pesada e clara de IgG para a clonagem. (A) as regiões variáveis de cadeia pesada (VH) e Light (VK ou VL) são recuperadas usando reações de PCR aninhadas. Os iniciadores passo I amplificam a partir da sequência de líder nativa e revertem de dentro da região constante. As setas múltiplas representam um pool de entre 7-12 primers, que são usados para assegurar a cobertura larga de germlines possíveis. Os primers de etapa II são aninhados, específicos às extremidades extremas do quadro de leitura aberto variável, e adicionam a seqüência às extremidades dos amplicões que são homólogo à seqüência adjacente no vetor da expressão. (B) o linker consiste na corrente clara constante (CK ou CL), seguida pelo promotor da corrente pesada e por uma seqüência não-nativa do peptide do sinal. Os amplicons, o linker, e a espinha dorsal do plasmídeo são ligados simultaneamente através da seqüência homous de sobreposição gerada durante a amplificação do PCR da etapa II, e isoladas como um plasmídeo intacto. As cadeias leves e pesadas recombinantes no vetor de expressão final são impulsionadas por promotores de CMV independentes (e idênticos), e são traduzidas como proteínas separadas. Por favor clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 3 : Os amplicões Chain variáveis pesados e claros de IgG são recuperados das únicas pilhas. As reações aninhadas do PCR (etapa II) foram carregadas diretamente no gel do agarose de 1% e visualizadas. A corrente pesada (H) e a corrente clara do Kappa (L) reações do PCR da mesma pilha foram carregadas em poços adjacentes assim que os amplicões emparelhados foram observados facilmente. Os produtos bem sucedidos da corrente pesada e clara são aproximadamente 400 BP e 350 BP respectivamente. A recuperação bem-sucedida de amplicões emparelhados é indicada por um (*). Por favor clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 4 : A especificidade do antígeno é confirmada de IgGs recombinantes. Os Plasmids que contêm as seqüências pesadas e claras recuperadas da corrente são transfected em pilhas de HEK para a expressão. Após quatro dias, os anticorpos recombinantes são avaliados para reatividade por ELISA. (A) a concentração de IgG nos meios esclarecidos foi medida e diluída a 20 μg/ml. O grupo de peptídeos utilizados para triagem foi avaliado individualmente por meio de 40 pmol por poço em placas de estreptavidina. Todos os IgGs foram testados em poços duplicados. (B) clones que EXIBIAM OD450 medida > 0,5 foram reavaliados (clones #32, #34, #35) usando 1:5 etapas de diluição contra os peptídeos identificados na tela. (C) uma vez confirmada como uma batida, um único clone do plasmídeo foi isolado do clone #34 ligações transformadas, transfected, e reconfirmado por Elisa. O mesmo foi feito para clones #32 e #35 (dados não mostrados). Por favor clique aqui para ver uma versão maior desta figura.
{kind=link}
Discussão
O método aqui apresentado combina citometria de fluxo e clonagem de células únicas, e os métodos que descrevemos aqui são baseados em métodos previamente desenvolvidos por Tiller e colegas11. Seu trabalho descreve a recuperação e a clonagem de anticorpos monoclonais a fim estudar o repertório da pilha de B nos seres humanos a nível de pilhas individuais. Nós adaptamos os componentes principais de seu processo para permitir a recuperação de anticorpos monoclonais antígeno-específicos de uma população de pilhas da memória B, incluindo a estratégia da primeira demão da amplificação da multi-etapa. A modificação principal é a adição de "isca" do antígeno rotulado. Adaptações adicionais foram feitas para o protocolo publicado, incluindo (mas não limitado a) modificando o backbone vetorial de clonagem em um plasmídeo de expressão única, cobertura de primer adicional do repertório de germline (ambos seqüência de líderes e quadro 1), transfection de pilhas da suspensão HEK293 para a expressão mais elevada, e o uso de polymerases da fidelidade elevada durante a amplificação do PCR.
Para os métodos descritos aqui, as etapas mais críticas estão perto da transição entre a citometria de fluxo e a clonagem de células únicas. Primeiramente, a colocação apropriada de pilhas únicas classificadas na placa é essencial. Definir o atraso de queda corretamente no classificador é uma etapa-chave. Fatores ambientais, como a baixa umidade, também devem ser levados em conta, como descobrimos que nossas eficiências de recuperação caem significativamente, a menos que uma arma estática seja usada nas placas de classificação alvo. Após a colocação da célula, as placas são centrifugadas para garantir que as células tenham contactado os 10 μL de tampão na parte inferior dos poços. Todas estas medidas são críticas para que a parcela da biologia molecular seja bem sucedida. Se as condições forem subideais para a reação de transcrição reversa de uma única célula, a amplificação de PCR de mesmo β-actina pode ser difícil. A força do método apresentado é a capacidade de interrogar milhões de células e apenas classificar os que correspondem aos critérios de reatividade do antígeno contra o antígeno de escolha. Isso requer que a relação sinal-ruído seja o mais alto possível, o que é feito otimizando as concentrações de isca antes de classificar. A isca Dual-etiquetada é usada para reduzir a recuperação de pilhas falso-positivas, que podem ocorrer se um dos fluoróforos tem o fundo elevado. Usando mais de uma isca de antígeno pode fazer o sinal: otimização de ruído mais difícil, mas a capacidade de interrogar múltiplos peptídeos simultaneamente é outro benefício deste método.
Quando as eficiências de recuperação são baixas, um conjunto de primers de β-actina aninhados é usado para confirmar que o modelo cDNA está presente. O inconveniente a esta aproximação é que é difícil determinar se não havia nenhuma pilha atual no poço ou a reação reversa da transcrição falhou, fazendo a Troubleshooting difícil. Ocasionalmente, o oposto ocorrerá e as eficiências são mais elevadas do que o esperado. A recuperação típica para a corrente pesada é entre 30-45% e as correntes claras são mais elevadas, entre 40-60%. Cada reação de PCR (passo I & II) usa 50 ciclos para amplificar a partir de uma única célula. O número elevado de ciclos totais igualmente faz este método suscetível à contaminação. A análise da sequência de amplicões pode ser usada para determinar se um contaminante foi introduzido, ou uma taxa de recuperação excepcionalmente elevada foi alcançada legitimamente.
A utilidade deste método para recuperar anticorpos humanos nativos de encontro a um antígeno da escolha tem diversas limitações significativas. Primeiro, apenas proteínas solúveis que podem ser rotuladas podem ser usadas como "isca" para a citometria de fluxo. Para os tipos apresentados nos resultados representativos, utilizou-se uma série de peptídeos sobrepostos sintetizados da proteína Tau, uma vez que o uso de toda a proteína se mostrou difícil. O uso de peptídeos lineares provavelmente não é ideal, uma vez que pode limitar a identificação de anticorpos contra Epitopes não-lineares ou estruturais. No entanto, pudemos identificar vários anticorpos únicos contra a Tau e estes estão actualmente a ser submetidos a uma nova avaliação8,9. Outra limitação é a baixa taxa de transferência da recuperação da biologia molecular e a clonagem de IgGs. A estratégia de triagem inicial permite a triagem de milhões de células, mas o processamento subsequente de biologia molecular consiste em vários estágios. Os esforços de otimização contínuos para incorporar novas tecnologias, como o Gibson assembly10 , simplificaram várias etapas, mas o gargalo continua a clonar pares de cadeias pesadas e leves individuais.
O método "bselex" utiliza o BCR expresso na superfície das células B da memória para identificar as células que exibem a reatividade do antígeno e, em seguida, recuperar essas células individuais via citometria de fluxo11,12. Devido a esta confiança no BCR, o método é restrito ao compartimento da pilha da memória B, e não captura as pilhas secretoras do anticorpo (ASCs) tais como plasmablasts. Entretanto, esta aproximação pode ser vantajosa a recuperar um repertório mais largo de imunuglobinas antígeno-específicos quando comparado aos ASCs. Os estudos da vacinação da gripe demonstram que quando os ASCs reactivos podem ser dominados por um número pequeno de clones expandidos da pilha de B, a população antígeno-específica da pilha da memória B é raramente clonal12. O receptor de célula T (TCR) na superfície das células T possui semelhanças com o receptor de células B no arranjo genético e recombinação para maximizar a diversidade. Abordagens de células únicas semelhantes às descritas no método aqui apresentado foram desenvolvidas para avaliar o repertório de células T, incluindo a recuperação e a clonagem de células alfa e beta13. No entanto, o reconhecimento do TCR exige que os peptídeos sejam apresentados por moléculas de MHC, acrescentando uma complexidade significativa à abordagem de isca rotulada para identificar células T peptídicas específicas. Ao contrário das células B, as células T não sofrem maturação de afinidade de toda a região variável, portanto, a identificação da região CDR3 curta e alguma seqüência de flanquear é tudo o que é necessário para a identificação e reconstituição. Finalmente, este método descreve a identificação de moléculas de IgG usando citometria de fluxo, mas também é possível usar marcadores fenotípicos alternativos para identificar células B de diferentes isotipos.
Atualmente, existem métodos que estão sendo desenvolvidos para conectar a enorme quantidade de dados coletados do sequenciamento da próxima geração com análise funcional da especificidade do antígeno, mas esses métodos ainda estão sendo refinados. Apesar de suas limitações, o método descrito aqui tem sido usado para identificar anticorpos com potencial valor terapêutico para doenças infecciosas e não infecciosas8,14,15, e representa um confiável aproximação para recuperar IgGs antígeno-específicos relevantes dos seres humanos, sem manipulação extensiva de pilhas de B ou de cultura extensiva da pilha.
Divulgações
Os autores não têm nada a divulgar
Agradecimentos
Os autores gostariam de agradecer a Lucy Chammas, Martha costa, Julie Kim, Nancy Heredia e Jeremy Macedo pelo extenso teste de muitas modificações de método e refinamento da atual plataforma BSelex.
Materiais
| Name | Company | Catalog Number | Comments |
| MoFlo Astrio EQ, Cell Sorter | MoFlo | cell sorting | |
| Biotinylated peptides | New England Peptide | Cell sorting "bait" | |
| Micro Bio-spin P30 gel column | BioRad | #7326223 | |
| Streptavidin (R-PE) | Thermo Scientific | SA10041 | |
| Streptavidin (APC) | Thermo Scientific | S32362 | |
| CD22 MicroBeads, human | Miltenyi | #130-046-401 | |
| LS columns | Miltenyi | #130-042-401 | |
| Pre separation filters | Miltenyi | #130-041-407 | |
| PerCp-Cy5.5 mouse anti-human CD19 | BD Biosciences | BD#340951 | |
| PE-Cy7 mouse anti-human CD27 | BD Biosciences | #560609 | |
| FITC mouse anti-human IgG | BD Biosciences | #560952 | |
| DAPI | Life Technologies | #D21490 | |
| RNaseOUT | Life Technologies | #10777-019 | |
| Bovine serum albumin, Fraction V | Sigma | #A4503 | |
| RPMI media | Hyclone | #SH30096.01 | |
| HI FBS (Fetal bovine serum) | Invitrogen | #10082147 | |
| Mastercycler Gradient | Eppendorf | Model #6325 | PCR machine |
| PCR 96-well plates | Phenix | MPS-500 | |
| PCR Plate mats | Phenix | SMX-PCR96 | |
| Advantage UltraPure PCR Deoxynucleotide Mix | Clontech | #639125 | |
| Superscript IV First-strand synthesis system | Invitrogen | #18091050 | |
| Phusion High Fidelity Polymerase | Thermo Scientific | F-531-L | |
| Platinum polymerase | Invitrogen | #10966108 | |
| Nuclease-free water | QIAGEN | #129114 | |
| Oligonucleotide primers | IDT | assorted | Primers for all PCR steps |
| Gel Extraction Kit | QIAGEN | #28706 | |
| PCR Purification kit | QIAGEN | #28106 | |
| Miniprep Kit | QIAGEN | #27106 | |
| NEBuilder HiFi DNA Assembly Cloning Kit | New England Biolabs | E5520S | Gibson assembly |
| Expi293 Expression Media | Invitrogen | #A14351-01 | |
| ExpiFectamine 293 Transfection Kit | Invitrogen | #A14525 | |
| Opti-MEM Media | Invitrogen | #31985070 | |
| CO2 incubator (Multitron) | |||
| Octet RED384 System | Pall/ Forte Bio | ||
| Pierce Streptavidin Coated High Binding Capacity Clear 96-well Plates | Thermo Scientific | #15500 | |
| Corning Costar 96-well Polystyrene Plate High Binding Half Area | Corning | #3690 | |
| Goat Anti-human IgG Fab Secondary Antibody | Jackson Labs | #109-036-097 | |
| Goat Anti-mouse HRP Secondary Antibody | Jackson Labs | #115-035-072 | |
| SureBlueTM TMB Microwell Peroxidase Substrate (1-Component) | KPL | #52-00-03 | |
| TMB Stop Solution | KPL | #50-85-06 |
Referências
- Su, K. Y., Watanabe, A., Yeh, C. H., Kelsoe, G., Kuraoka, M. Efficient Culture of Human Naive and Memory B Cells for Use as APCs. Journal of Immunology. 197 (10), 4163-4176 (2016).
- Corti, D., Lanzavecchia, A. Broadly neutralizing antiviral antibodies. Annual Review of Immunology. 31, 705-742 (2013).
- Steinitz, M., Klein, G., Koskimies, S., Makel, O. EB virus-induced B lymphocyte cell lines producing specific antibody. Nature. 269 (5627), 420-422 (1977).
- Bloom, A. D., Nakamura, F. T. Establishment of a tetraploid, immunoglobulin-producing cell line from the hybridization of two human lymphocyte lines. Proceedings of the National Academy of Sciences of the United States of America. 71 (7), 2689-2692 (1974).
- Olsson, L., Kaplan, H. S. Human-human hybridomas producing monoclonal antibodies of predefined antigenic specificity. Proceedings of the National Academy of Sciences of the United States of America. 77 (9), 5429-5431 (1980).
- Lechner, J., et al. Alterations in Circulating Immune Cells in Neovascular Age-Related Macular Degeneration. Scientific Reports. 5, 16754(2015).
- Loken, M. R., Brosnan, J. M., Bach, B. A., Ault, K. A. Establishing optimal lymphocyte gates for immunophenotyping by flow cytometry. Cytometry. 11 (4), 453-459 (1990).
- Pascual, G., et al. Immunological memory to hyperphosphorylated tau in asymptomatic individuals. Acta Neuropathologica. 133 (5), 767-783 (2017).
- van Ameijde, J., et al. Enhancement of therapeutic potential of a naturally occurring human antibody targeting a phosphorylated Ser(422) containing epitope on pathological tau. Acta Neuropathologica Communications. 6 (1), 59(2018).
- Gibson, D. G., et al. Enzymatic assembly of DNA molecules up to several hundred kilobases. Nature Methods. 6 (5), 343-345 (2009).
- Tiller, T., et al. Efficient generation of monoclonal antibodies from single human B cells by single cell RT-PCR and expression vector cloning. Journal of Immunological Methods. 329 (1-2), 112-124 (2008).
- Wrammert, J., et al. Rapid cloning of high-affinity human monoclonal antibodies against influenza virus. Nature. 453 (7195), 667-671 (2008).
- Han, A., Glanville, J., Hansmann, L., Davis, M. M. Linking T-cell receptor sequence to functional phenotype at the single-cell level. Nature Biotechnology. 32 (7), 684-692 (2014).
- Apetri, A., et al. A common antigenic motif recognized by naturally occurring human VH5-51/VL4-1 anti-tau antibodies with distinct functionalities. Acta Neuropathologica Communications. 6 (1), 43(2018).
- Jones, H. G., et al. Structural basis for recognition of the central conserved region of RSV G by neutralizing human antibodies. PLoS Pathogens. 14 (3), 1006935(2018).
- Janssen Vaccines & Prevention B.B. US patent. , U.S. Patent Application No. 20170210787, Published July 27, 2017 (2017).
Reimpressões e Permissões
Solicitar permissão para reutilizar o texto ou figuras deste artigo JoVE
Solicitar PermissãoThis article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. Todos os direitos reservados