
Method Article
Metodo di screening a singola cellula per la selezione e il recupero di anticorpi con specifiche desiderate da popolazioni di cellule di memoria umana b arricchite
In questo articolo
Riepilogo
Il metodo BSelex per identificare e recuperare singoli anticorpi specifici dell'antigene dalle cellule mononucleari del sangue periferico umano combina citometria di flusso con PCR a cella singola e clonazione.
Abstract
Il repertorio degli anticorpi umani rappresenta una fonte in gran parte non sfruttata di potenziali anticorpi terapeutici e biomarcatori utili. Mentre gli attuali metodi computazionali, come il sequenziamento di nuova generazione (NGS), producono enormi insiemi di dati sul repertorio degli anticorpi a livello di sequenza, sono necessari dati funzionali per identificare quali sequenze sono rilevanti per un particolare antigene o insieme di Antigeni. Qui, descriviamo un metodo per identificare e recuperare singoli anticorpi specifici dell'antigene dalle cellule mononucleari del sangue periferico (PBMC) da un donatore di sangue umano. Questo metodo utilizza un arricchimento iniziale di cellule B mature e richiede una combinazione di marcatori cellulari fenotipici e proteine fluorescenti per isolare le cellule di memoria IgG B tramite la citometria di flusso. Le regioni variabili della catena leggera e pesante vengono quindi clonate e ri-schermate. Anche se limitato al raggruppamento delle celle B di memoria B, questo metodo sfrutta la citometria di flusso per interrogare milioni di celle B e restituisce sequenze a catena pesante e leggera accoppiate da una singola cella in un formato pronto per l'espressione e la conferma della specificità. Gli anticorpi recuperati con questo metodo possono essere considerati per potenziale terapeutico, ma possono anche collegare specificità e funzione con approcci bioinformatici per valutare il repertorio delle cellule B all'interno degli individui.
Introduzione
Gli anticorpi sono una classe crescente di molecole terapeutiche, e il repertorio delle cellule B esistente in qualsiasi essere umano è una potenziale fonte di tali anticorpi. Una volta recuperati da un donatore umano, non richiedono alcun adattamento o "umanizzazione", passi necessari per gli anticorpi generati in altri sistemi animali. Esistono diversi metodi per l'identificazione e l'isolamento di anticorpi umani, tra cui l'attivazione e la proliferazione delle cellule B1, l'immortalizzazione tramite trasformazione EBV2,3e la generazione di linee cellulari ibrideoma4 ,5. Tuttavia, tutti questi metodi richiedono un'estesa coltura cellulare per lo screening e il recupero di anticorpi specifici dell'antigene. Le informazioni sul repertorio degli anticorpi umani sono state notevolmente ampliate con lo sviluppo della tecnologia di sequenziamento (NGS) di nuova generazione, consentendo l'identificazione di enormi quantità di singole sequenze presenti nei campioni di donatori. Tuttavia, poiché NGS fornisce una visione agnostica di tutte le sequenze presenti, non consente l'identificazione e l'isolamento di anticorpi specifici dell'antigene, in particolare nel caso di anticorpi rari o a bassa frequenza.
Lo scopo del metodo "BSelex" è identificare gli anticorpi specifici dell'antigene provenienti dalle cellule mononucleari del sangue periferico nei donatori umani e isolare e recuperare le sequenze di questi anticorpi per ulteriori analisi. Questo metodo utilizza la citometria di flusso e lo smistamento delle cellule per sfruttare il recettore delle cellule B (BCR) espresso sulla superficie delle cellule B di memoria. Milioni di cellule B possono essere sottoposte a screening per la specificità dell'antigene tramite la citometria di flusso prima che vengano avviati i metodi di biologia molecolare più a basso consumo. L'identificazione a catena pesante e leggera accoppiata non è possibile nella maggior parte dei metodi NGS, che analizzano le sequenze cellulari alla rinfusa. Nel metodo qui descritto, le cellule sono isolate singolarmente e il recupero accoppiato di sequenze a catena sia pesanti che leggere è possibile, che consente la clonazione diretta e l'espressione dell'IgG completo.
Protocollo
L'uso di campioni di volontari umani ha seguito protocolli approvati dal Scripps Research Institute Institutional Review Board. Il consenso informato è stato ottenuto dai donatori prima della donazione di sangue.
1. Preparazione del reagente
-
Cellule mononucleari del sangue periferiche (PBMC)
- Recuperare le cellule mononucleari del sangue periferico dai normali donatori umani mediante l'isolamento di Ficoll-Plaque Plus. Criopreserve a 50 milioni di cellule / mL nel 90% siero bovino fetale (FBS) e 10% di zolfo dimetilo (DMSO). Congelare le cellule a -80 gradi centigradi e trasferirle in azoto liquido per una conservazione a lungo termine.
-
Etichettatura dei peptidi dell'antigene bersaglio per lo smistamento
- Generare o ottenere peptidi specifici per la proteina bersaglio (fino a 70 residui lunghi) con una biotina terminale.
- Etichetta 4 nmol di ogni singolo peptide biotinylato con streptavidina covalentmente attaccato al PE (R-phycoerythrin) o APC (allophycocyanin) in tubi separati. Preparare utilizzando un rapporto molare di 9:1 peptide a streptavidin (SA), con una concentrazione finale di circa 3,2 M per SA-PE e 5,7 SA-APC. Preparare i tetrameri di biotina come controllo negativo.
- Incubare ogni miscela di peptidi durante la notte, al buio, a 4 gradi centigradi con una miscelalenta.
- Rimuovere il fluoroforo non legato passando peptidi etichettati attraverso microcolonne contenenti perline di poliacrilammide.
- Conservare i tetrameri peptidi fino a 2 mesi a 4 gradi centigradi.
2. Ordinamento delle celle
- Almeno 1 h fino a 16 h prima dell'isolamento del CD22, scongelare i PBMC dei donatori e lasciare riposare a 37 gradi centigradi.
- Togliere le fiale dei PBC congelati dal congelatore e trasferirle immediatamente a un bagno d'acqua a 37 gradi centigradi. Quando le fiale sono quasi scongelate, trasferire nell'area di lavoro.
- Trasferire il contenuto di ciascuna fiala su un tubo da 50 mL contenente un mezzo preriscaldato (mantenendo separati i donatori). Aggiungere un volume medio a un volume finale di 50 mL. Mescolare delicatamente il contenuto.
- Centrifuga a 370 x g per 6 min a temperatura ambiente. Eliminare il supernatante, riutilizzare le cellule in 15 mL di RPMI mezzo completo ed eseguire un conteggio delle cellule.
- Regolare la concentrazione cellulare a 2,0 x 107 cellule / mL con il mezzo completo RPMI, e trasferire le cellule a un pallone T75 o più grande e incubare a 37 c per almeno 1 h e fino a 16 h per consentire il tempo di recupero per le cellule scongelate.
- Raccogliere le celle (pooling se lo si desidera) e centrifugare a 370 x g per 6 min a 4 gradi centigradi. Scartare supernatante e risospendere nuovamente le cellule in 5 mL di buffer MACS freddo ghiaccio (PBS, pH 7.6 contenente 0,5% BSA e 2 mM EDTA), portare a 50 mL con buffer MACS ed eseguire un conteggio delle celle.
- Centrifugare le cellule a 400 x g per 7 min a 4 gradi centigradi.
- Isolare le cellule CD22e B mediante selezione positiva tramite acquisizione su microsfere CD22. Utilizzare il buffer MACS per tutti gli washe. Contare e centrifugare 400 x g per 7 min a 4 gradi centigradi. Il recupero previsto è del 5-10% della popolazione totale di PBMC.
- Risospendere le celle a 40 milioni per buffer FACS mL (Tampolo di Tris, pH 8.0 contenente 0,5% BSA e 2 mM EDTA) e procedere con la colorazione cellulare dei donatori singolarmente o come pool.
- Celle di macchie per l'ordinamento delle celle B della memoria
- Rimuovere un'aliquota di cellule per l'impostazione del citometro e la compensazione per ciascuno dei fluorofori di etichettatura utilizzati. Includere celle non colorate come controllo.
- Calcolare il volume di colorazione finale per il numero di cd22- cellule ottenute (colorazione 2 milioni per 100 l).
- Aggiungere marcatori extracellulari per le cellule B (CD19-PerCP-Cy5.5), IgG (IgG-FITC) e vano di memoria (CD27-PECy7) secondo le raccomandazioni di diluizione dei produttori, e mescolare delicatamente. Aliquota 107 cellule in un tubo da 1,5 mL per il controllo negativo e trasferire le cellule rimanenti in un tubo da 15 mL.
- Aggiungere i tetrameri biotina-SA con etichetta doppia al tubo di controllo negativo a una concentrazione finale di 36 nM ciascuno per PE e APC moltiplicato per il numero di peptidi nel tubo di ordinamento e portare il volume a 0,5 mL finale con tampone FACS (ad esempio, 10 peptides x 36 nM
- Aggiungere i tetrameri peptidi al tubo di ordinamento a 36 nM ciascuno per PE e APC e portare le celle a 2x106 per 100 - L con buffer TBS. Incubare a 4 gradi centigradi al buio e con una leggera rotazione per 30-60 min.
- Lavare 2x a 4 gradi centigradi, 400 x g,7 min, rimuovendo un'aliquota da contare prima dell'ultimo lavaggio. Filtrare le celle utilizzando tubi di tappo filtro 5 mL. Aggiungere la macchia DAPI (4',6-diamidino-2-phenylindole) a 0,3 m di concentrazione finale appena prima dello smistamento come marcatore dell'integrità della membrana cellulare.
- Ordinamento di una singola cella tramite citometria di flusso
- Preparare 48 pozze di piastre PCR 96-bene per la selezione.
- Preparare una miscela master: (2 , L di 10x tampone RT, 0,5 l di inibitore di RNase, 7,5 L di acqua sterile di grado PCR) per ogni campione. Aliquota 10 l per pozzo, coprire, e conservare a 4 gradi centigradi fino a quando non è pronto per l'ordinamento.
- Eseguire il controllo negativo sull'ordinatore di celle, analizzando l'intero campione.
- Impostare i cancelli di citometria di flusso per isolare le popolazioni di cellule appropriate per lo smistamento di singole cellule.
- Area di plotto FSC vs area SSC e impostare Gate R1 per isolare i linfociti.
- Traccial' FSC vs larghezza FSC e imposta Gate R3 per escludere i doppietti FSC.
- Traccia l'altezza SSC rispetto alla larghezza SSC e imposta Gate R4 per escludere i doppietti SSC.
- Tracciare l'altezza SSC rispetto a DAPI e impostare Gate R5 per isolare le celle vive (DAPI-).
- Stampa IgG vs CD19 e impostare Gate R6 per isolare le celle IgGe B (CD19- IgG) .
- Stampa IgG vs CD27 e impostare Gate R7 per selezionare le celle B di memoria (CD27alto).
- Tracciaanti antigeni-PE (Ag-PE) vs antigene-APC (Ag-APC) e impostaGate R8 come un Ag-PE -/Ag-APC- quadrante con un basso numero di eventi nel doppio cancello positivo.
- Impostare i cancelli di citometria di flusso per isolare le popolazioni di cellule appropriate per lo smistamento di singole cellule.
- Raccogliere un numero uguale di celle di memoria dal campione antigene-positivo (Ag)per la determinazione del segnale al rumore.
- Ordinare le singole cellule con il fenotipo CD19, CD27, Ag-PE, Ag-APC(Gate R8) in piastre PCR da 96 pozzetto.
- Copertine con nastri in alluminio, centrifuga per 1 min a 400 x g, e conservare a -80 gradi centigradi per la clonazione futura.
- Preparare 48 pozze di piastre PCR 96-bene per la selezione.
3. Clonazione a cella singola
-
Reazioni di trascrizione inversa
- Rimuovere la singola piastra di ordinamento della cella B da 80 . Scongelare sul ghiaccio per 2 min.
- Preparare una miscela master dNTP/buffer con il 10% di detergente non ionico e oligo(dT) come primer inverso. Aggiungere mix di enzimi ad ogni bene contenente la singola cellula e NON MIX.
- Incubare 5 min a 65 gradi centigradi. Aggiungere la miscela master dell'enzima contenente 100 mM DTT, l'inibitore di RNase e l'enzima della trascrizione inversa per portare il volume totale a 20 . Incubare 10 min a 50 gradi centigradi, quindi 10 min a 80 .
- Trasferimento sul ghiaccio prima di avviare i passaggi PCR
-
Reazioni PCR nidificate in più fasi
- Utilizzare reazioni PCR nidificate separate vengono utilizzate per l'amplificazione a catena pesante (HC) e a catena leggera (LC).
- Per il primo round di amplificazione PCR (passaggio I), utilizzare un pool di primer in avanti e un singolo primer inverso per amplificare le regioni variabili HC e LC in reazioni PCR separate (Figura 2). Per impostazione predefinita, il pool di primer in avanti amplificherà una grande percentuale di sequenze di germinali (L) e il primer inverso è specifico per le regioni costanti a valle di ogni catena, comprese le kappa (z) e lambda (lambda) per le catene luminose16.
- Utilizzare 2,5 ll del prodotto cDNA come modello per ogni reazione PCR (fase I). Aggiungere i primer alla concentrazione di reazione finale di 1 M per ciascuno. Raddoppiare il buffer di polimerasi 10x a 2volte di concentrazione, portare il volume finale a 25 l per reazione. Eseguire le reazioni della PCR HC utilizzando [94 s/4 min; 94 s /C, 55 s /20 s, 68 C/60 s; 68 min c/3 per 50 cicli. Eseguire le reazioni della PcR LC usando [98 min/4 min; 98 s /C, 72 s /20 s, 72 c/60 s; 72 min/C per 50 cicli.
- Utilizzare 2,5 l del prodotto Step I come modello per ogni reazione PCR (fase II). Aggiungere il pool di primer di inoltro specifico per la regione del quadro HC e LC (e -) e un pool di primer inversi specifiche per la regione di giunzione di ogni catena di anticorpi. Raddoppiare il buffer 10x di polimerasi a 2x concentrazione ed eseguire le reazioni PCR 50 cicli ciascuno utilizzando gli stessi parametri del passo I.
- Eseguire le reazioni PCR completate su 1% gel di agarose per visualizzare i risultati positivi di amplificazione. Recuperare gli amplificatori accoppiati (sia pesante che a catena leggera dalla stessa cellula) e isolare tramite estrazione gel. Determinare la concentrazione di frammenti di DNA tramite OD260 per calcoli accurati della miscela di legatura.
- Combina frammenti di catene pesanti e leggeri recuperati con un frammento del linker e la spina dorsale vettoriale dell'espressione usando una reazione di legatura a 4 frammenti utilizzando un kit commerciale.
- Trasforma le reazioni di legatura in batteri chimicamente competenti utilizzando un kit commerciale. Una volta placcati su piastre antibiotiche, aggiungere 4 mL di mezzi di comunicazione di crescita (con antibiotici) alla coltura di trasformazione rimanente e incubare 37 gradi durante la notte a 250 rpm. Questa è la "cultura della miscela di legamento".
- Preparare il DNA di miniprep dalle colture di mix di legatura durante la notte utilizzando un kit commerciale e determinare la concentrazione di DNA plasmide risultante.
4. Schermo ELISA per la conferma della specificità dell'antigene
-
Transfect 10 g di DNA miniprep con reagente a base di lipidi cationici in 10 mL di sospensioni 293cellule, e incuba per 3-4 giorni a 37 gradi centigradi (8% CO 2) mentre ruota a 125 giri/ . Per la raccolta, centrifugare cultura 10 min a 1.000 x g e recuperare i media chiarificati.
- Misurare la concentrazione di IgG nei supernatanti tramite affinità alla proteina A.
- Testare ogni IgG (in supernatante) a 20 g/mL da un assaggio di adsorbenti enzimatico (ELISA) contro i singoli peptidi utilizzati per il tipo, catturati su una piastra di streptavidin o actina come controllo negativo. Utilizzare un anticorpo secondario perossidase capra contro l'uomo IgG Fab per rilevare cloni ricombinanti. Dopo la sottrazione dello sfondo, OD > 0.5 è definito come positivo dello schermo.
- Confermare i risultati positivi dello schermo eseguendo un ELISA aggiuntivo, utilizzando diluizioni del supernatante IgG per generare una curva di concentrazione a partire da 20 g per mL e tracciando contro OD per ogni antigene che mostra la reattività nella schermata iniziale ELISA.
Risultati
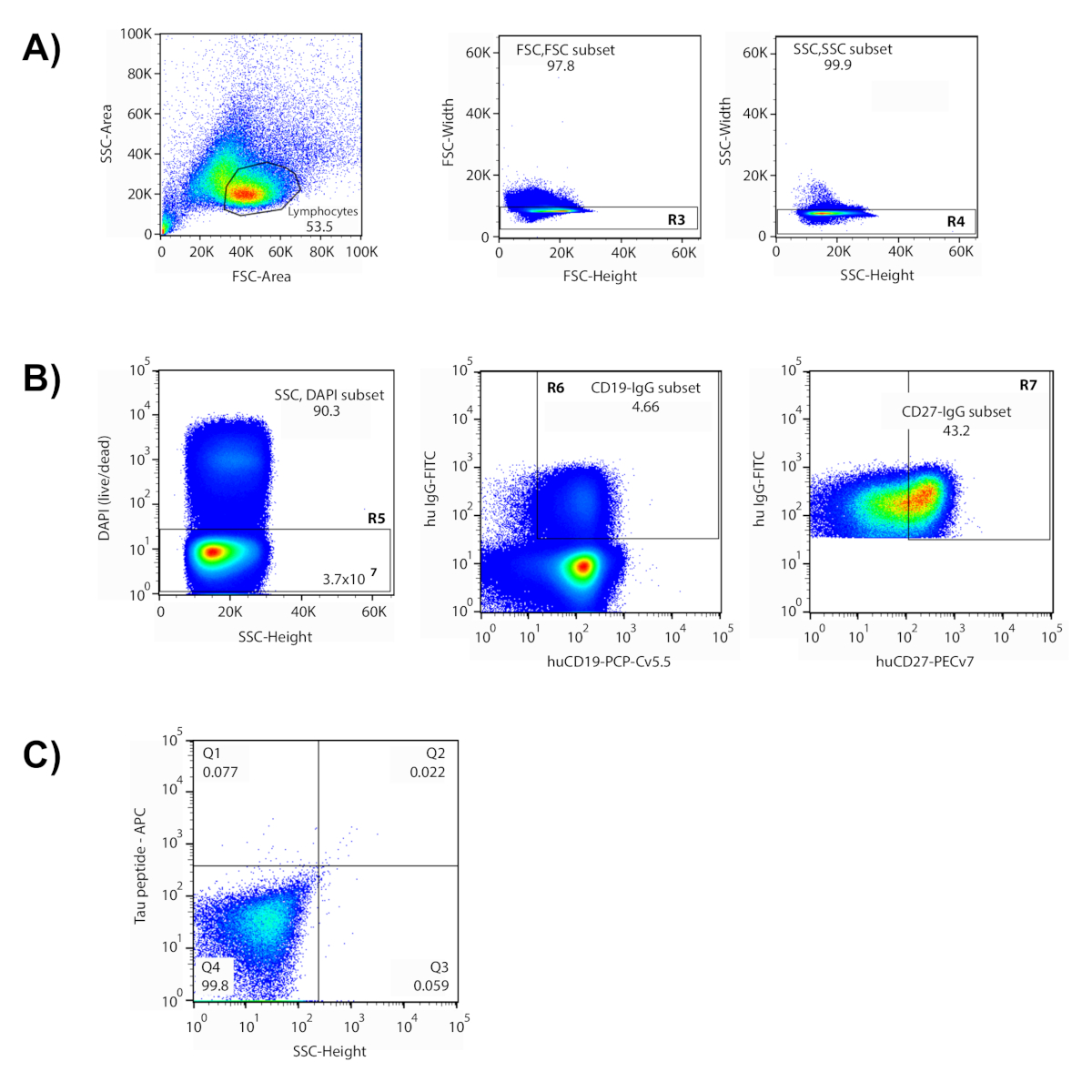
Questo metodo copre un processo a più fasi per isolare gli anticorpi specifici dell'antigene dai donatori umani. Nei dati rappresentativi qui mostrati, le cellule sono state incubate con un pool di peptidi con etichetta fluorescente che rappresenta diversi domini della proteina tau, tra cui peptidi fosforizzati per imitare siti di fosforo putarificati. Questi peptidi sono stati utilizzati come "bait" per identificare le cellule che sono reattive con epitopi tau di interesse. In preparazione per la selezione, è stato utilizzato un gruppo di marcatori fenotipici etichettati fluorescentmente per identificare diverse popolazioni di cellule all'interno della popolazione di cellule B arricchite. Una serie di porte di citometria sono state concepite per isolare le celle B di memoria di destinazione (Figura1). I linfociti sono stati isolati in base alle loro dimensioni e granularità delle cellule utilizzando grafici a dispersione in avanti (FSC) e a dispersione laterale (SSC) nella citometria di flusso6,7. In seguito all'esclusione di più celle ("doppietti") e cellule morte, i marcatori fenotipici hanno permesso la segregazione delle cellule B di igg- tramite IgG, CD19 (cellula B) e CD27 (memoria). In questo approccio, il marcatore CD27 non si distribuisce in due popolazioni discrete, quindi il 45% superiore delle cellule espressive CD27- sono inclusi per il cancello finale. Infine, le cellule raddoppiate positive per i fluorofori APC e PE si distribuiscono nel quadrante superiore destro del grafico di gating, indicando la reattività a entrambe le versioni etichettate dei peptidi. Le celle che cadevano all'interno del cancello disegnato sono state isolate e ordinate in singoli pozzi di una piastra di 96 pozze. L'uso di antigene con due etichette diverse aumenta il rapporto segnale-rumore e riduce il numero di falsi positivi nel successivo processo di biologia molecolare. Le celle ordinate rappresentavano circa lo 0,1% delle celle B della memoria nel cancello finale e lo 0,001% del campione di celle iniziale.
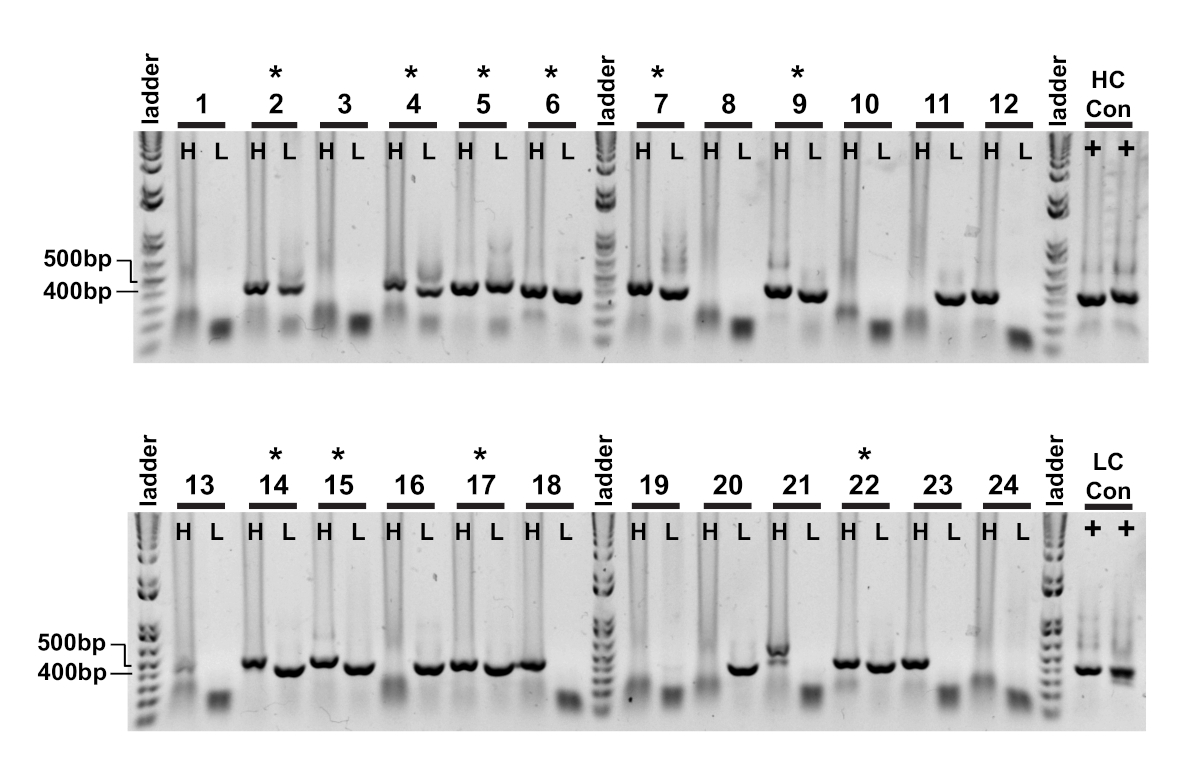
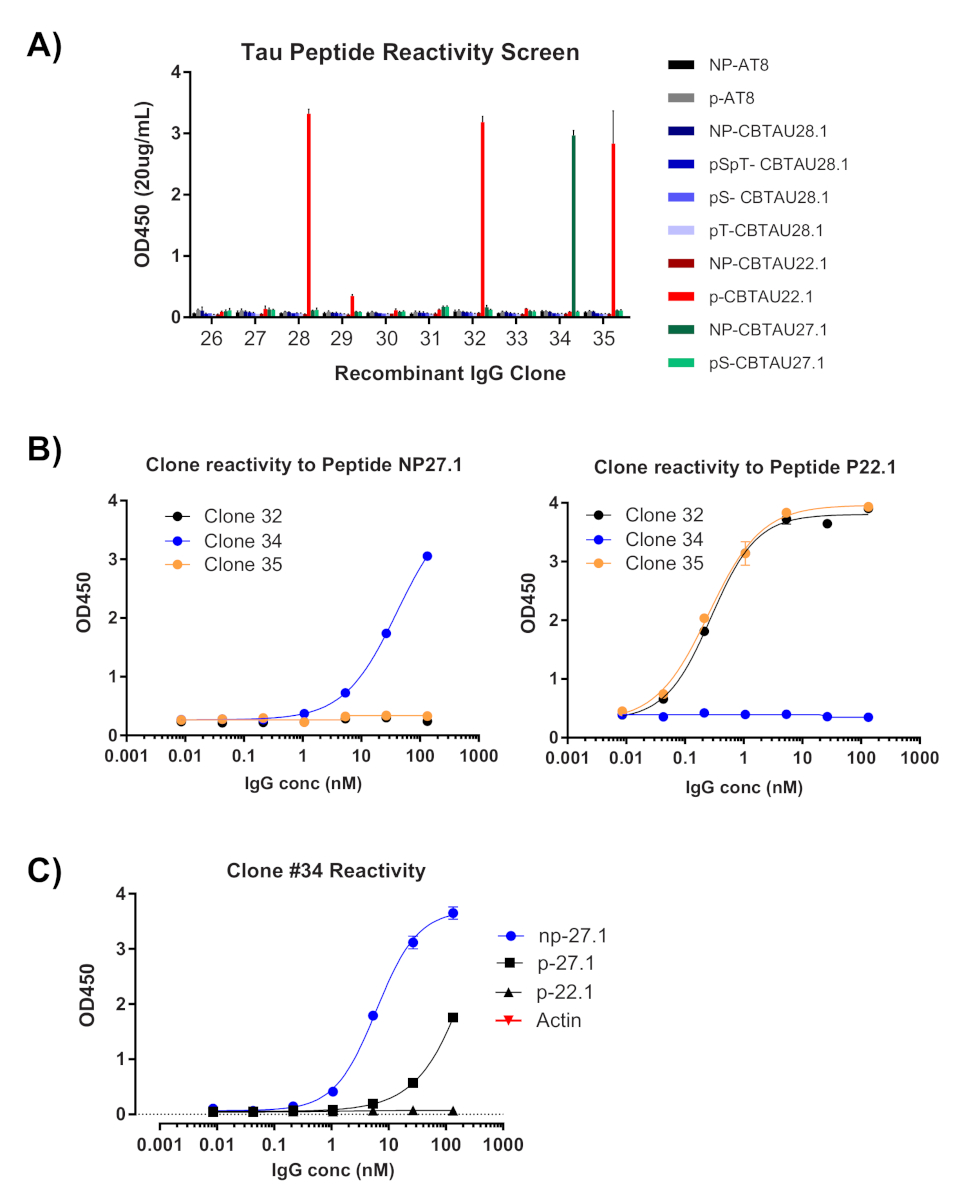
La prima lettura della clonazione a cella singola è la conferma dell'amplificazione delle rispettive catene di variabili pesanti e leggere (Figura 2). Poiché si desidera il recupero accoppiato di entrambi gli amplificatori, le reazioni PCR vengono valutate fianco a fianco sul gel di agarose e le coppie abbinate vengono asciste dal gel e dai frammenti di DNA estratti. L'efficienza tipica dell'amplificazione è del 30-50% a catena pesante e 50-70% della catena leggera (kappa). Il recupero degli amplificatori accoppiati è di solito tra il 25-40% di efficienza. Queste efficienze variano tra i pool di donatori e i dati rappresentativi (Figura 3) sono un esempio di amplificazione molto efficiente da 24 celle singole (recupero accoppiato del 42%). Dopo la clonazione IgG, il vettore di espressione IgG viene trafitto in cellule di sospensione del rene embrionale umano (HEK-293), che vengono utilizzate per massimizzare l'espressione. L'uso di un mezzo privo di siero riduce le proteine contaminanti dalla preparazione dell'anticorpo ricombinante e aiuta a ridurre al minimo il rumore nei successivi saggi di legame. Gli anticorpi recuperati vengono sottoposti a screening rispetto al pannello originale dei peptidi tau e segnati per la reattività da ELISA (Figura 4). Una soglia iniziale di OD - 0,5 sopra lo sfondo viene utilizzata per indicare la reattività positiva dell'antigene, e la proteina di atto in actina viene utilizzata come controllo per il legame non specifico. Se la citometria di flusso e le fasi di smistamento utilizzano un pool misto di peptidi, lo screening ELISA è il primo passo di deconvoluzione reattività specifiche degli IgG recuperati. Tre dei 10 IgG analizzati hanno dimostrato reattività contro il peptide CBTAU22.1 fosforo, e uno era reattivo al CBTAU27.1 (Figura 4A). Ulteriori conferme sono state completate utilizzando una curva di concentrazione degli stessi campioni di anticorpi ricombinanti rispetto ai peptidi identificati nella schermata iniziale e un peptide aggiuntivo come controllo negativo (Figura 4B). Per ogni colpo positivo, un singolo clone di plasmide è stato isolato dal pool trasformato e riconfermato dallo stesso metodo ELISA. Solo Clone 34 è mostrato i dati presentati, e mentre la reattività al CBTAU27 non fosforato è stato confermato, legame di affinità inferiore è stato osservato anche con il peptide fosforelare 27.1 (Figura 4C).

Figura 1 : isolamento delle singole cellule antigene-reattiva tramite citometria di flusso. (A) Il grafico è rappresentativo in cui la popolazione dei linfociti era contenuta all'interno della porta disegnata. (B) Cancelli basati sulla dispersione in avanti e laterale (FSC-height v FSC-width (R3); SSC- altezza v SSC- width(R4)) sono stati utilizzati per escludere i doppietti. Solo le celle all'interno dei cancelli disegnati sono state valutate nei complotti successivi. DAPI- le cellule sono state considerate morte ed escluse (R5). In questo esperimento sono state interrogate 3,7 x 107 cellule "vive". La maggior parte delle cellule vive erano cellule B (CD19)ecellule IgG (4,66%) sono stati isolati da questa popolazione (R6). Nel cancello di selezione (R7) sono stati inclusi il 43,2% delle celle di memoriaCD27che esprime l'espressione. (C) Il quadrante 2 (Q2) conteneva cellule reattive a entrambi gli antigeni etichettati (peptide-APC v peptide-PE), e queste cellule sono state smistate e recuperate singolarmente in una piastra di 96 pozze. Fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}

Figura 2 : l'amplificazione PCR recupera la sequenza variabile pesante e leggera IgG per la clonazione. (A) Entrambe le regioni variabili della catena pesanti (VH) e della luce (Vk o Vl) vengono recuperate utilizzando reazioni PCR nidificate. Passo I primer amplificare in avanti dalla sequenza di associazione nativa e invertire dall'interno dell'area costante. Le frecce multiple rappresentano un pool di 7-12 primer, che vengono utilizzati per garantire un'ampia copertura di possibili linee germinali. I primer del passo II sono nidificati, specifici per le estremità estreme del fotogramma di lettura aperto variabile, e aggiungono sequenza alle estremità degli amplificatori che sono omologi alla sequenza adiacente nel vettore di espressione. (B) Il linker è costituito dalla catena di luce costante (Ck o Cl), seguita dal promotore della catena pesante e da una sequenza di peptidi del segnale non nativo. Gli amplificatori, il linker e la spina dorsale plasmide vengono contemporaneamente ligate tramite la sovrapposizione di sequenza omologa generata durante l'amplificazione PCR del Passo II e isolata come un plasmide intatto. Le catene pesanti e leggere ricombinanti nel vettore di espressione finale sono guidate da promotori CMV indipendenti (e identici) e sono tradotte come proteine separate. Fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}

Figura 3 : gli amplificatori a catena variabile igG pesanti e leggeri vengono recuperati da singole celle. Le reazioni PCR nidificate (fase II) sono state caricate direttamente su gel di agarose dell'1% e visualizzate. Le reazioni PCR a catena pesante (H) e kappa a catena leggera (L) dalla stessa cellula sono state caricate in pozzi adiacenti, quindi gli amplificatori accoppiati sono stati facilmente osservati. I prodotti a catena pesante e leggera di successo sono rispettivamente di circa 400 bp e 350 bp. Il recupero riuscito degli amplificatori accoppiati è indicato da un . Fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}

Figura 4 : La specificità dell'antigene è confermata dagli IGM ricombinanti. Plasmidi contenenti le sequenze a catena pesante e leggera recuperate vengono trascate in cellule HEK per l'espressione. Dopo quattro giorni, gli anticorpi ricombinanti sono valutati per la reattività da ELISA. (A) La concentrazione di IgG nei supporti chiarificati è stata misurata e diluita a 20 g/mL. La riserva di peptidi utilizzata per lo smistamento è stata valutata individualmente utilizzando 40 pmol per pozzo in piastre di streptavidin. Tutti gli Ig Sono stati testati in pozzi duplicati. (B) I cloni che mostravano la misurazione OD450 >0.5 sono stati rivalutati (Cloni #32, #34, #35) utilizzando passaggi di diluizione 1:5 contro i peptidi identificati sullo schermo. (C) Una volta confermato come un colpo, un singolo clone di plasmid è stato isolato dal clone #34 le legature trasformate, trasconfettate e riconfermate da ELISA. Lo stesso è stato fatto per i cloni #32 e #35 (dati non mostrati). Fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}
Discussione
Il metodo qui presentato combina la citometria di flusso e la clonazione a cella singola, e i metodi qui descritti si basano su metodi precedentemente sviluppati da Tiller e colleghi11. Il loro lavoro descrive il recupero e la clonazione di anticorpi monoclonali al fine di studiare il repertorio delle cellule B negli esseri umani a livello di singole cellule. Abbiamo adattato i principali componenti del loro processo per consentire il recupero di anticorpi monoclonali specifici dell'antigene da una popolazione di cellule B di memoria, compresa la strategia di amplificazione a più fasi. La modifica principale è l'aggiunta di antigene etichettato "bait". Ulteriori adattamenti sono stati apportati al protocollo pubblicato, tra cui (ma non solo) la modifica della spina dorsale del vettore di clonazione in un'unica espressione plasmid, ulteriore copertura primer del repertorio germinale (sia sequenza leader che framework 1), trasfezione delle cellule HEK293 di sospensione per un'espressione superiore e l'uso di polimerasi ad alta fedeltà durante l'amplificazione PCR.
Per i metodi descritti di seguito, i passaggi più critici sono vicini alla transizione tra citometria di flusso e clonazione a cella singola. In primo luogo, il corretto posizionamento di singole cellule ordinate nella piastra è essenziale. L'impostazione corretta del ritardo di rilascio sulla selezionatrice è un passaggio chiave. Anche i fattori ambientali, come la bassa umidità, devono essere presi in considerazione, poiché abbiamo scoperto che le nostre efficienze di recupero diminuiscono in modo significativo a meno che non venga utilizzata una pistola statica sulle piastre di ordinamento bersaglio. Dopo il posizionamento delle cellule, le piastre sono centrifughe per garantire che le cellule abbiano contattato il buffer 10 -L nella parte inferiore dei pozzi. Tutte queste misure sono fondamentali per il successo della porzione di biologia molecolare. Se le condizioni sono sub-ottimali per la reazione di trascrizione inversa di una singola cellula, l'amplificazione PCR di anche la z-actin può essere difficile. La forza del metodo presentato è la capacità di interrogare milioni di cellule e ordinare solo quelle che corrispondono ai criteri di reattività dell'antigene contro l'antigene di scelta. Ciò richiede che il rapporto segnale/rumore sia il più alto possibile, il che avviene ottimizzando le concentrazioni di esche prima dello smistamento. L'esca a doppio marchio viene utilizzata per ridurre il recupero delle cellule falsi positive, che possono verificarsi se uno dei fluorofori ha uno sfondo elevato. L'utilizzo simultaneo di più esche antigene può rendere più difficile l'ottimizzazione del segnale:rumore, ma la capacità di interrogare più peptidi contemporaneamente è un altro vantaggio di questo metodo.
Quando l'efficienza di recupero è bassa, viene utilizzato un set di primer nidificati di atto z, per confermare la presenza del modello cDNA. Lo svantaggio di questo approccio è che è difficile determinare se non c'era nessuna cellula presente nel pozzo o la reazione di trascrizione inversa non è riuscita, rendendo difficile la risoluzione dei problemi. Occasionalmente si verificherà il contrario e le efficienze sono superiori al previsto. Il recupero tipico per la catena pesante è tra il 30-45% e le catene leggere sono più alte, tra il 40-60%. Ogni reazione PCR (Passaggio I & II) utilizza 50 cicli per amplificare da una singola cella. L'elevato numero di cicli totali rende anche questo metodo suscettibile di contaminazione. L'analisi della sequenza degli amplificatori può essere utilizzata per determinare se è stato introdotto un contaminante o se un tasso di recupero insolitamente elevato è stato legittimamente raggiunto.
L'utilità di questo metodo per recuperare gli anticorpi umani nativi contro un antigene di scelta ha diversi limiti significativi. In primo luogo, solo le proteine solubili che possono essere etichettate possono essere utilizzate come "bait" per la citometria di flusso. Per i tipi presentati nei risultati rappresentativi, abbiamo usato una serie di peptidi sovrapposti sintetizzati della proteina tau, poiché l'utilizzo dell'intera proteina si è rivelato difficile. L'uso di peptidi lineari probabilmente non è ottimale, poiché può limitare l'identificazione di anticorpi contro epitopi non lineari o strutturali. Tuttavia, siamo stati in grado di identificare diversi anticorpi unici contro il tau e questi sono attualmente in fase di ulteriore valutazione8,9. Un'altra limitazione è la bassa produttività del recupero della biologia molecolare e la clonazione degli IGB. La strategia di selezione iniziale consente lo screening di milioni di cellule, ma il successivo elaborazione della biologia molecolare è costituito da più fasi. Gli sforzi di ottimizzazione in corso per incorporare le tecnologie più recenti, come Gibson Assembly10 hanno semplificato diversi passaggi, ma il collo di bottiglia rimane clonare singole coppie di catene pesanti e leggere.
Il metodo "BSelex" utilizza il BCR espresso sulla superficie delle cellule di memoria B per identificare le cellule che visualizzano la reattività dell'antigene, e quindi recuperare queste singole cellule tramite citometria di flusso11,12. A causa di questa dipendenza dal BCR, il metodo è limitato al compartimento delle cellule B della memoria B e non cattura le cellule di secernerazione degli anticorpi (ASC) come i plasmablasti. Tuttavia, questo approccio può essere vantaggioso per recuperare un repertorio più ampio di immunoglobini specifici dell'antigene rispetto agli ASC. Gli studi sulla vaccinazione influenzale dimostrano che, sebbene gli ASC reattivi possano essere dominati da un piccolo numero di cloni di cellule B espansi, la popolazione di cellule B della memoria specifica dell'antigene è raramente clonale12. Il recettore delle cellule T (TCR) sulla superficie delle cellule T possiede somiglianze con il recettore delle cellule B nella disposizione genica e nella ricombinazione per massimizzare la diversità. Approcci a singola cellula simili a quelli descritti nel metodo qui presentato sono stati sviluppati per valutare il repertorio delle cellule T, compreso il recupero e la clonazione a singola cellula delle catene alfa e beta13. Tuttavia, il riconoscimento TCR richiede che i peptidi siano presentati dalle molecole MHC, aggiungendo una significativa complessità all'approccio dell'esca etichettata per identificare le cellule T specifiche del peptide. A differenza delle cellule B, le cellule T non subiscono la maturazione dell'affinità dell'intera regione variabile, quindi l'identificazione della regione Breve CDR3 e una sequenza di fianco è tutto ciò che è necessario per l'identificazione e la ricostituzione. Infine, questo metodo descrive l'identificazione delle molecole IgG usando la citometria di flusso, ma è anche possibile utilizzare marcatori fenotipici alternativi per identificare cellule B di diversi isotipi.
Attualmente, ci sono metodi in fase di sviluppo per collegare l'enorme quantità di dati raccolti dalla prossima generazione di sequenziamento con l'analisi funzionale della specificità dell'antigene, ma questi metodi sono ancora in fase di perfezionamento. Nonostante i suoi limiti, il metodo qui descritto è stato utilizzato per identificare gli anticorpi con potenziale valore terapeutico sia per le malattie infettive che non infettive8,14,15, e rappresenta un approccio per recuperare i relativi IgG antigeni specifici dagli esseri umani, senza un'ampia manipolazione delle cellule B o un'estesa coltura cellulare.
Divulgazioni
Gli autori non hanno nulla da rivelare
Riconoscimenti
Gli autori desiderano ringraziare Lucy Chammas, Martha Costa, Julie Kim, Nancy Heredia e Jeremy Macedo per l'ampio test di molte modifiche al metodo e il perfezionamento dell'attuale piattaforma BSelex.
Materiali
| Name | Company | Catalog Number | Comments |
| MoFlo Astrio EQ, Cell Sorter | MoFlo | cell sorting | |
| Biotinylated peptides | New England Peptide | Cell sorting "bait" | |
| Micro Bio-spin P30 gel column | BioRad | #7326223 | |
| Streptavidin (R-PE) | Thermo Scientific | SA10041 | |
| Streptavidin (APC) | Thermo Scientific | S32362 | |
| CD22 MicroBeads, human | Miltenyi | #130-046-401 | |
| LS columns | Miltenyi | #130-042-401 | |
| Pre separation filters | Miltenyi | #130-041-407 | |
| PerCp-Cy5.5 mouse anti-human CD19 | BD Biosciences | BD#340951 | |
| PE-Cy7 mouse anti-human CD27 | BD Biosciences | #560609 | |
| FITC mouse anti-human IgG | BD Biosciences | #560952 | |
| DAPI | Life Technologies | #D21490 | |
| RNaseOUT | Life Technologies | #10777-019 | |
| Bovine serum albumin, Fraction V | Sigma | #A4503 | |
| RPMI media | Hyclone | #SH30096.01 | |
| HI FBS (Fetal bovine serum) | Invitrogen | #10082147 | |
| Mastercycler Gradient | Eppendorf | Model #6325 | PCR machine |
| PCR 96-well plates | Phenix | MPS-500 | |
| PCR Plate mats | Phenix | SMX-PCR96 | |
| Advantage UltraPure PCR Deoxynucleotide Mix | Clontech | #639125 | |
| Superscript IV First-strand synthesis system | Invitrogen | #18091050 | |
| Phusion High Fidelity Polymerase | Thermo Scientific | F-531-L | |
| Platinum polymerase | Invitrogen | #10966108 | |
| Nuclease-free water | QIAGEN | #129114 | |
| Oligonucleotide primers | IDT | assorted | Primers for all PCR steps |
| Gel Extraction Kit | QIAGEN | #28706 | |
| PCR Purification kit | QIAGEN | #28106 | |
| Miniprep Kit | QIAGEN | #27106 | |
| NEBuilder HiFi DNA Assembly Cloning Kit | New England Biolabs | E5520S | Gibson assembly |
| Expi293 Expression Media | Invitrogen | #A14351-01 | |
| ExpiFectamine 293 Transfection Kit | Invitrogen | #A14525 | |
| Opti-MEM Media | Invitrogen | #31985070 | |
| CO2 incubator (Multitron) | |||
| Octet RED384 System | Pall/ Forte Bio | ||
| Pierce Streptavidin Coated High Binding Capacity Clear 96-well Plates | Thermo Scientific | #15500 | |
| Corning Costar 96-well Polystyrene Plate High Binding Half Area | Corning | #3690 | |
| Goat Anti-human IgG Fab Secondary Antibody | Jackson Labs | #109-036-097 | |
| Goat Anti-mouse HRP Secondary Antibody | Jackson Labs | #115-035-072 | |
| SureBlueTM TMB Microwell Peroxidase Substrate (1-Component) | KPL | #52-00-03 | |
| TMB Stop Solution | KPL | #50-85-06 |
Riferimenti
- Su, K. Y., Watanabe, A., Yeh, C. H., Kelsoe, G., Kuraoka, M. Efficient Culture of Human Naive and Memory B Cells for Use as APCs. Journal of Immunology. 197 (10), 4163-4176 (2016).
- Corti, D., Lanzavecchia, A. Broadly neutralizing antiviral antibodies. Annual Review of Immunology. 31, 705-742 (2013).
- Steinitz, M., Klein, G., Koskimies, S., Makel, O. EB virus-induced B lymphocyte cell lines producing specific antibody. Nature. 269 (5627), 420-422 (1977).
- Bloom, A. D., Nakamura, F. T. Establishment of a tetraploid, immunoglobulin-producing cell line from the hybridization of two human lymphocyte lines. Proceedings of the National Academy of Sciences of the United States of America. 71 (7), 2689-2692 (1974).
- Olsson, L., Kaplan, H. S. Human-human hybridomas producing monoclonal antibodies of predefined antigenic specificity. Proceedings of the National Academy of Sciences of the United States of America. 77 (9), 5429-5431 (1980).
- Lechner, J., et al. Alterations in Circulating Immune Cells in Neovascular Age-Related Macular Degeneration. Scientific Reports. 5, 16754 (2015).
- Loken, M. R., Brosnan, J. M., Bach, B. A., Ault, K. A. Establishing optimal lymphocyte gates for immunophenotyping by flow cytometry. Cytometry. 11 (4), 453-459 (1990).
- Pascual, G., et al. Immunological memory to hyperphosphorylated tau in asymptomatic individuals. Acta Neuropathologica. 133 (5), 767-783 (2017).
- van Ameijde, J., et al. Enhancement of therapeutic potential of a naturally occurring human antibody targeting a phosphorylated Ser(422) containing epitope on pathological tau. Acta Neuropathologica Communications. 6 (1), 59 (2018).
- Gibson, D. G., et al. Enzymatic assembly of DNA molecules up to several hundred kilobases. Nature Methods. 6 (5), 343-345 (2009).
- Tiller, T., et al. Efficient generation of monoclonal antibodies from single human B cells by single cell RT-PCR and expression vector cloning. Journal of Immunological Methods. 329 (1-2), 112-124 (2008).
- Wrammert, J., et al. Rapid cloning of high-affinity human monoclonal antibodies against influenza virus. Nature. 453 (7195), 667-671 (2008).
- Han, A., Glanville, J., Hansmann, L., Davis, M. M. Linking T-cell receptor sequence to functional phenotype at the single-cell level. Nature Biotechnology. 32 (7), 684-692 (2014).
- Apetri, A., et al. A common antigenic motif recognized by naturally occurring human VH5-51/VL4-1 anti-tau antibodies with distinct functionalities. Acta Neuropathologica Communications. 6 (1), 43 (2018).
- Jones, H. G., et al. Structural basis for recognition of the central conserved region of RSV G by neutralizing human antibodies. PLoS Pathogens. 14 (3), 1006935 (2018).
- . . US patent. , (2017).
Ristampe e Autorizzazioni
Richiedi autorizzazione per utilizzare il testo o le figure di questo articolo JoVE
Richiedi AutorizzazioneThis article has been published
Video Coming Soon
Personale delle biblioteche
Copyright © 2025 MyJoVE Corporation. Tutti i diritti riservati