
Method Article
Одноклеточный метод скрининга для отбора и восстановления антител с желаемой спецификой от обогащенной человеческой памяти B Популяции клеток
В этой статье
Резюме
Метод BSelex для выявления и восстановления отдельных антиген-специфических антител из человеческих периферических клеток крови сочетает в себе цитометрию потока с одноклеточным ПЦР и клонированием.
Аннотация
Репертуар человеческих антител представляет собой в значительной степени неиспользованный источник потенциальных терапевтических антител и полезных биомаркеров. В то время как современные вычислительные методы, такие как секвенирование следующего поколения (NGS), дают огромные наборы данных о репертуаре антител на уровне последовательности, функциональные данные необходимы для определения того, какие последовательности имеют отношение к конкретному антигену или набору Антигенов. Здесь мы описываем метод выявления и извлечения отдельных антиген-специфических антител из периферических моноядерных клеток крови (ПБМК) от донора крови человека. Этот метод использует первоначальное обогащение зрелых В-клеток и требует сочетания фенотипических маркеров клеток и флуоресцентно помеченного белка, чтобы изолировать клетки памяти IgG B через цитометрию потока. Переменные области тяжелой и легкой цепи затем клонируются и повторно проверяются. Хотя этот метод ограничен отсеком ячейки памяти B, он использует цитометрию потока, чтобы допросить миллионы В-клеток и возвращает парные тяжелые и светлые цепные последовательности из одной ячейки в формате, готовом к выражению и подтверждению специфичности. Антитела, извлеченные с помощью этого метода, могут быть рассмотрены для терапевтического потенциала, но также могут связать специфичность и функции с биоинформатическими подходами для оценки репертуара B-клеток в организме человека.
Введение
Антитела являются растущим классом терапевтических молекул, и существующий репертуар В-клеток у любого человека является потенциальным источником таких антител. При извлечении из человека донора, они не требуют адаптации или "гуманизации", шаги, которые необходимы для антител, генерируемых в других системах животных. Существует несколько методов идентификации и изоляции человеческих антител, включая активацию В-клеток и пролиферацию1,увековечение через преобразование EBV2,3,и генерацию линий клеток гибридомы4 ,5. Тем не менее, все эти методы требуют обширной клеточной культуры для проверки и восстановления антигена конкретных антител. Информация о репертуаре человеческих антител была значительно расширена с развитием технологии секвенирования следующего поколения (NGS), позволяющей выявлять огромное количество отдельных последовательностей, присутствующих в образцах доноров. Однако, поскольку NGS дает агностик зрения всех присутствующих последовательностей, он не позволяет для идентификации и изоляции антиген-специфических антител, особенно в случае редких или низкочастотных антител.
Целью метода «BSelex» является выявление антиген-специфических антител из циркулирующих периферических моноядерных клеток крови у доноров человека, а также изоляция и восстановление последовательностей этих антител для дальнейшего анализа. Этот метод использует цитометрию потока и сортировки клеток, чтобы воспользоваться рецептором В-клеток (BCR), выраженным на поверхности клеток памяти В. Миллионы В-клеток могут быть проверены на антиген-специфическость с помощью цитометрии потока, прежде чем более низкой пропускной способностью молекулярной биологии методы инициируются. Парная тяжелая и световая цепная идентификация невозможна в большинстве методов NGS, которые анализируют клеточные последовательности оптом. В методе, который мы описываем здесь, клетки изолированы по отдельности, и парное восстановление как тяжелых, так и световых цепных последовательностей возможно, что позволяет прямое клонирование и выражение полного IgG.
протокол
Использование образцов из человеческих добровольцев следовали протоколам, одобренным Советом по институциональному обзору Научно-исследовательского института Скриппса. Информированное согласие было получено от доноров до сдачи крови.
1. Подготовка реагента
-
Моноядерные клетки периферической крови (ПБМК)
- Восстановление периферических моноядерных клеток крови от обычных доноров человека путем изоляции Ficoll-Plaque Plus. Криоконсервацию на 50 миллионов клеток/мл в 90% сыворотки крупного рогатого скота плода (FBS) и 10% диметилсульфоксида (DMSO). Заморозить клетки при -80 градусов и переложить на жидкий азот для длительного хранения.
-
Маркировка целевых пептидов антигена для сортировки
- Генерировать или получать пептиды, специфичные для белка-мишени (до 70 остатков в длину) с терминальным биотином.
- Этикетка 4 нмоль каждого отдельного биотининатированного пептида с стрептавидином ковалентно прилагается к ПЭ (R-фикоэритрин) или APC (аллофикоцианин) в отдельных трубах. Приготовьтесь с использованием молярного соотношения 9:1 пептида к стрептавидину (SA), с окончательной концентрацией примерно 3,2 мкм для SA-PE и 5,7 мкм SA-APC. Подготовка тетрамеры Biotin в качестве отрицательного контроля.
- Инкубировать каждую пептидную смесь на ночь, в темноте, при 4 градусах Цельсия при медленном смешивании.
- Удалить несвязанный фторофор, проходя помеченные пептиды через микроколонны, содержащие бусы полиакриламид.
- Храните пептидные тетрамы до 2 месяцев при 4 градусах Цельсия.
2. Сортировка клеток
- По крайней мере 1 ч до 16 ч до CD22 "изоляции, оттепель донора PBMCs и позволяют отдохнуть при 37 градусов по Цельсию.
- Извлеките флаконы замороженных ПБМК из морозильной камеры и немедленно перенесите на водяную ванну 37 градусов по Цельсию. Когда флаконы почти разморозятся, перенесите на рабочую зону.
- Перенесите содержимое каждого флакона в трубку 50 мл, содержащую предварительно разогретую среду (сохраняя доноры отдельно). Добавьте средний к окончательному объему 50 мл. Аккуратно перемешайте содержимое.
- Центрифуга при температуре 370 х г в течение 6 мин при комнатной температуре. Откажитесь от супернатанта, resuspend клетки в 15 мл RPMI полной среде и выполнять количество клеток.
- Отрегулируйте концентрацию клеток до 2,0 х 107 клеток/мл с rpMI полной среде, и передачи клеток в колбу T75 или больше и инкубировать при 37 градусов по Цельсию, по крайней мере 1 ч и до 16 ч, чтобы время восстановления для оттаять клетки.
- Соберите клетки (при желании) и центрифугу при 370 х г в течение 6 мин при 4 градусах Цельсия. Отбросить супернатант и повторно гофрел клетки в 5 мл ледяного буфера MACS (PBS, pH 7.6, содержащий 0,5% BSA и 2 мМ EDTA), довести до 50 мл с буфером MACS и выполнять количество клеток.
- Центрифуги клетки на 400 х г в течение 7 мин при 4 кв. C.
- Изолировать CD22и B-клетки путем положительного отбора через захват на CD22 микробусы. Используйте буфер MACS для всех смок. Считайте и центрифуги 400 х г в течение 7 мин при 4 градусах Цельсия. Ожидаемое восстановление составляет 5-10% от общей численности населения ПБМК.
- Resuspend клетки на 40 миллионов мл буфера FACS (Tris буфер, pH 8.0, содержащий 0,5% BSA и 2 мМ EDTA) и приступить к клеточному окрашиванию доноров индивидуально или в бассейн.
- Пятно клетки для сортировки ячейки памяти B
- Удалите аликвот клеток для установки цитометра и компенсации для каждого из используемых флюорофоров маркировки. Включите неокрашенные ячейки в качестве элемента управления.
- Рассчитайте окончательный объем окрашивания для количества полученных клеток CD22 (окрашивание 2 млн на 100 л).
- Добавьте внеклеточные маркеры для В-клеток (CD19-PerCP-Cy5.5), IgG (IgG-FITC) и отсека памяти (CD27-PECy7) в соответствии с рекомендациями производителей по разбавлению и аккуратно перемешайте. Aliquot 107 ячеек в трубку 1,5 мл для отрицательного контроля и передачи оставшихся клеток в трубку 15 мл.
- Добавьте двойной помеченный биотин-SA тетрамеры в отрицательную контрольную трубку при конечной концентрации 36 нм каждый для ПЭ и APC, умноженный на количество пептидов в сортированной трубке, и доведите объем до 0,5 мл финала с буфером FACS (например, 10 пептидов x 36 nM-360 nM).
- Добавьте пептидные тетрамы в сортировку по 36 нм каждый для ПЭ и БТР и доведите клетки до 2x106 на 100 л с tbS Buffer. Инкубировать при 4 градусах Цельсия в темноте и с мягким вращением в течение 30-60 мин.
- Вымойте 2x при 4 кв с, 400 х г,7 мин, удалив аликот, чтобы рассчитывать до последней стирки. Фильтровые ячейки с использованием 5 мл фильтра крышки труб. Добавьте DAPI (4',6-diamidino-2-phenylindole) пятно до 0,3 мкм конечной концентрации непосредственно перед сортировкой в качестве маркера целостности клеточной мембраны.
- Одноклеточная сортировка с помощью цитометрии потока
- Подготовьте 48 скважин из 96-колодных плит ПЦР для сортировки.
- Подготовьте мастер-микс: (2 зл 10-х буфера РТ, 0,5 л ингибитора RNase, 7,5 л стерильной воды ПЦР- класса) на образец. Aliquot 10 л на колодец, накройте крышкой и храните при 4 градусах Цельсия до готовности к сортировке.
- Запустите отрицательный контроль на сортировке ячейки, анализируя весь образец.
- Установите ворота цитометрии потока, чтобы изолировать соответствующие популяции клеток для сортировки одной ячейки.
- Участок FSC области против SSC области и установить ворота R1 для изоляции лимфоцитов.
- Участок FSC высота против FSC ширина и установить ворота R3, чтобы исключить FSC дублетов.
- Участок SSC высота против SSC ширина и установить ворота R4, чтобы исключить SSC дублетов.
- Участок SSC высота против DAPI и установить ворота R5, чтобы изолировать живые клетки (DAPI-).
- Участок IgG против CD19 и установить ворота R6, чтобы изолировать IgGи B клетки (CD19и IgG)).
- Участок IgG против CD27 и установить ворота R7, чтобы выбрать ячейки памяти B (CD27высокий).
- Участок Антиген-PE (Ag-PE) против антигена-APC (Ag-APC) иустановить ворота R8 в качестве Ag-PE /Ag-APC- квадрант с низким количеством событий в двойных положительных ворот.
- Установите ворота цитометрии потока, чтобы изолировать соответствующие популяции клеток для сортировки одной ячейки.
- Соберите равное количество клеток памяти из антиген-положительного образца (Ag)для определения сигнала к шуму.
- Сортировать одиночные ячейки, имеющие фенотип CD19и CD27, Ag-PEи Ag-APC (Gate R8) в подготовленные 96-колодские ПЦР пластины.
- Обложка пластин с алюминиевой лентой колодки, центрифуга в течение 1 мин на 400 х г, и хранить при -80 градусов по Цельсию для будущего клонирования.
- Подготовьте 48 скважин из 96-колодных плит ПЦР для сортировки.
3. Клонирование одноклеточных элементов
-
Обратная транскрипция реакций
- Снимите одну пластину сортировки ячейки B с 80 градусов по Цельсию. Оттепель на льду в течение 2 мин. Спин пластины в течение 2 мин на 3300 х г, чтобы бассейн содержимое в нижней части скважины перед открытием.
- Подготовьте dNTP/буфер мастер смесь с 10% неионических моющих средств и олиго (dT) в качестве обратного грунтовки. Добавьте смесь фермента к каждому хорошо содержащему одиночную клетку и НЕMIX.
- Инкубировать 5 мин при 65 градусах Цельсия. Добавьте ферментную мастер-миксу, содержащую 100 мМ DTT, ингибитор RNase и фермент обратной транскриптазы, чтобы довести общий объем до 20 кл. Инкубировать 10 мин при 50 градусах Цельсия, затем 10 мин при 80 градусах Цельсия.
- Переход на лед до начала пЦР-шагов
-
Многоступенчатые вложенные реакции ПЦР
- Использование отдельных вложенных реакций ПЦР используется для усиления тяжелой цепи (HC) и легкой цепи (LC).
- Для первого раунда усиления ПЦР (Шаг I) используйте пул передних грунтовок и один обратный грунтовку для усиления переменных областей HC и LC в отдельных реакциях ПЦР(рисунок 2). По замыслу, пул передних праймеров будет усиливать большой процент последовательностей лидера зародышей (L), а реверсивная грунтовка специфична для постоянных областей каждой цепи, включая каппу (я) и лямбу (я) для легких цепей16.
- Используйте 2,5 злпродукта кДНК в качестве шаблона для каждой реакции ПЦР (Шаг I). Добавьте грунтовки к конечной концентрации реакции в размере 1 мкм для каждого из них. Удвойте 10-разный буфер полимеразы до концентрации 2x, доведите окончательный объем до 25 л за реакцию. Выполнить HC PCR реакции, используя 94 C /4 мин; 94 C /15 s, 55 КС /20 с, 68 КС /60 с; 68 C /3 мин " Запустите реакции LC PCR, используя 98 C/4 мин; 98 C /15 s, 72 C /20 s, 72 C/60 s; 72 C/3 мин.
- Используйте 2,5 зл и l продукта Step I в качестве шаблона для каждой (Шаг II) ПЦР реакции. Добавьте пул форвардных праймеров, специфичную для области HC и LC (no и) framework 1 и пул реверсивных грунтовок, специфичную для области соединения каждой цепи антител. Удвойте 10-разлимную буфер полимеразы до концентрации 2x и запустите 50 циклов ПЦР, каждый из которых использует те же параметры, что и Шаг I.
- Запустите завершенные реакции ПЦР на 1% агарозный гель, чтобы визуализировать положительные удары усиления. Восстановить парные ампиктоны (как тяжелые, так и светлые цепи из одной клетки) и изолировать через гель экстракции. Определите концентрацию фрагмента ДНК через OD260 для точных расчетов перевязочной смеси.
- Объедините восстановленные тяжелые и светлые фрагменты цепи с фрагментом связующим звеном и хепольной основой выражения, используя реакцию перевязки 4 фрагментов с помощью коммерческого комплекта.
- Преобразование реакций перевязки в химически компетентные бактерии с помощью коммерческого комплекта. После того, как покрывали на антибиотик пластины, добавить 4 мл роста средств (с антибиотиком) в остальной культуре преобразования, и инкубировать 37 градусов по Цельсию в одночасье при 250 об/мин. Это "культура перевязки микса".
- Подготовка мини-prep ДНК из ночной перевязки смеси культур с помощью коммерческого комплекта, и определить в результате плазмид ной концентрации ДНК.
4. ELISA Экран для подтверждения антиген-специфическисть
-
Трансфект 10 мкг мини-прежней ДНК с катионным липидным реагентом в 10 мл суспензии 293 клетки, и инкубировать в течение 3-4 дней при 37 градусах Цельсия (8% CO2) при повороте при 125 об/мин. Для сбора урожая, центрифуги культуры 10 мин на 1000 х г и восстановить уточнил средств массовой информации.
- Измерьте концентрацию IgG в супернатантах через сродство к протеину А.
- Проверьте каждый IgG (в супернатанте) на 20 мкг/мл фермент-связанных адсорбент анализа (ELISA) против отдельных пептидов, используемых для рода, захваченных на стрептавидиновые пластины или актин в качестве отрицательного контроля. Используйте коза анти-человеческого пероксидазы вторичного антитела против человека IgG Fab для обнаружения рекомбинантных клонов. После вычитания фона, OD sgt; 0.5 определяется как экран положительный.
- Подтвердите положительные хиты экрана, выполняя дополнительные ELISA, используя разбавления супернатанта IgG для создания кривой концентрации, начиная с 20 мкг на мл, и построение против OD для каждого антигена, показывающего реактивность в начальном экране ELISA.
Результаты
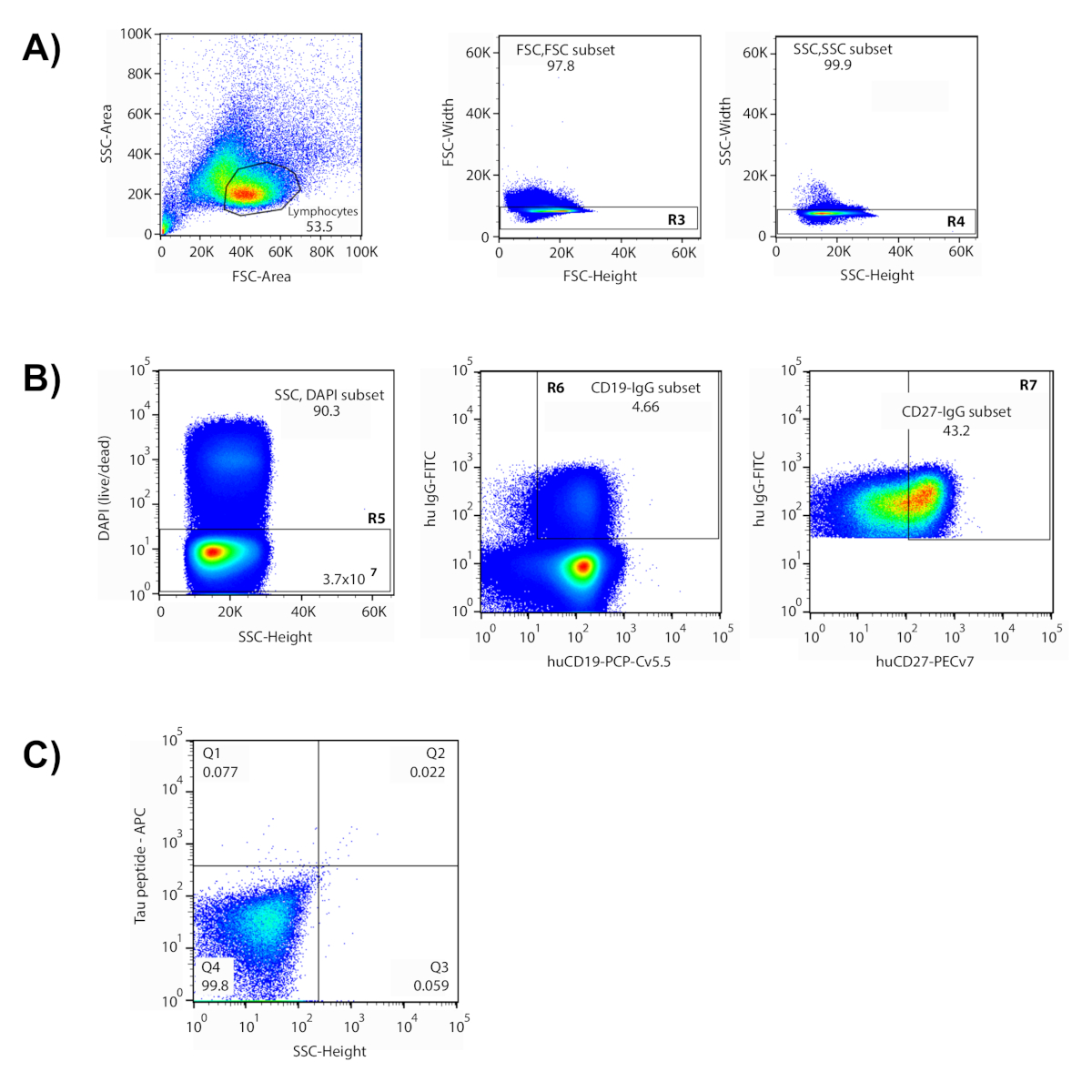
Этот метод охватывает многоступенчатый процесс изоляции антигенных антител от человеческих доноров. В репрезентативных данных, показанных здесь, клетки были инкубированы с бассейном флуоресцентно помеченных пептидов, представляющих несколько различных областей тау белка, в том числе фосфорилированных пептидов для имитации хосфорилирования сайтов. Эти пептиды были использованы в качестве "приманки" для выявления клеток, которые реактивна с тау эпитопы (ы) интерес. В рамках подготовки к сортировке для идентификации использовалась панель флуоресцентно маркированных фенотипических маркеров для идентификации различных популяций клеток в обогащенной популяции входящих в группу входящих в группу В. Серия ворот цитометрии были разработаны, чтобы изолировать целевые клетки памяти B(рисунок 1). Лимфоциты были изолированы на основе их размера клетки и детализации с использованием переднего рассеяния (FSC) и бокового рассеяния (SSC) участков в потоке цитометрии6,7. После исключения нескольких клеток ("дублеты") и мертвых клеток, фенотипические маркеры позволили сегрегации IgGи памяти B-клеток через IgG, CD19 (B-клетки) и CD27 (память). При таком подходе маркер CD27 не распределяется на две отдельные популяции, поэтому для конечных ворот включены 45% лучших 45% клеток CD27и выражения. Наконец, клетки, дважды положительные для APC и PE фторфоров распространять в верхнем правом квадранте gating графа, что свидетельствует о реактивности обеих помеченных версий пептидов. Клетки, попавший в нарисованные ворота, были изолированы и отсортированы по отдельным колодцам 96-колодца. Использование антигена с двумя различными метками увеличивает соотношение сигнала к шуму и уменьшает количество ложных срабатываний в последующем процессе молекулярной биологии. Отсортированные ячейки составляли около 0,1% клеток памяти B в конечных воротах и 0,001% от исходного образца ячейки.
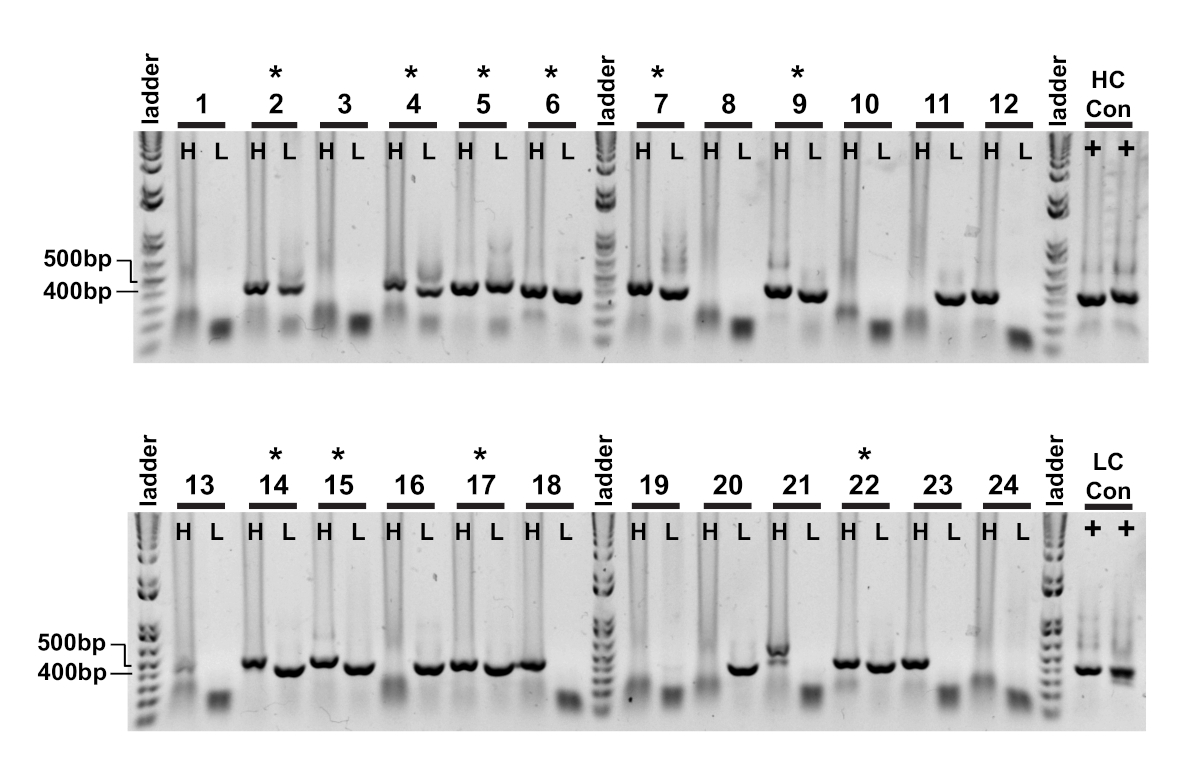
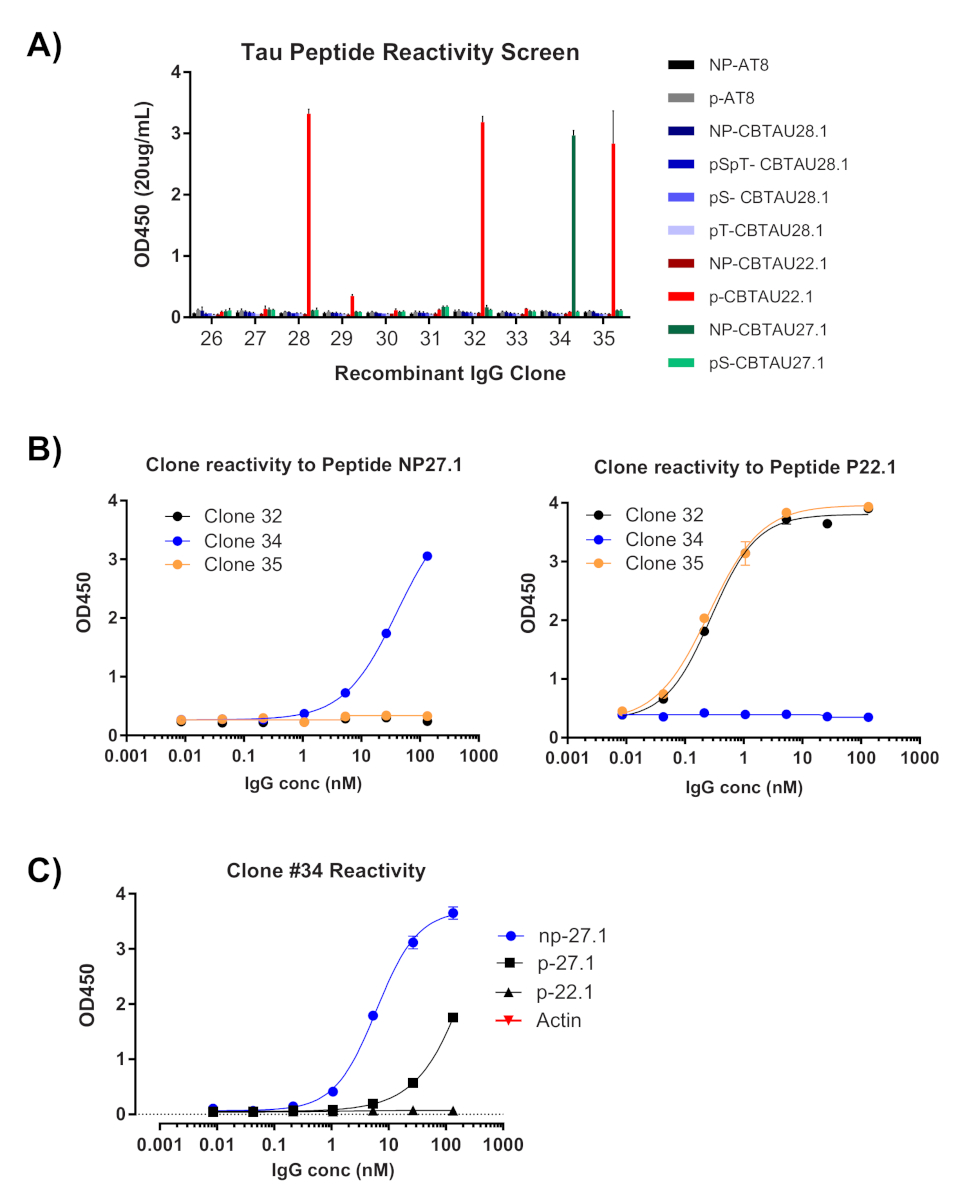
Первое считывание одноклеточного клонирования является подтверждением усиления соответствующих тяжелых и легких переменных цепей(рисунок 2). Так как парное восстановление обоих ампилонов желательно, реакции ПЦР оцениваются бок о бок на гель агарозы, и соответствующие пары вырезаются из геля и фрагментов ДНК извлечены. Типичная эффективность усиления составляет 30-50% тяжелой цепи и 50-70% легкой (каппа) цепи. Восстановление парных ампиктонов, как правило, между 25-40% эффективности. Эти показатели варьируются в зависимости от донорских пулов, и репрезентативные данные(рисунок3) являются примером очень эффективного усиления из 24 одиночных клеток (42% парного восстановления). После клонирования IgG вектор экспрессии IgG трансфицируется в эмбриональные почки человека (HEK-293), которые используются для максимального выражения. Использование среды, свободной от сыворотки, уменьшает загрязняющие белки из рекомбинантных антител и помогает свести к минимуму шум в последующих связывающих анализах. Восстановленные антитела проверяются на исходной панели тау пептидов и забил для реактивности ELISA (Рисунок 4). Первоначальный порог OD 0.5 выше фона используется для обозначения положительной реактивности антигена, а белок «Актин» используется в качестве контроля для неспецифической связывания. Если цитометрия потока и сортировки шаги использовать смешанный пул пептидов, скрининг ELISA является первым шагом deconvoluting конкретных реактивности восстановленных IgGs. Три из 10 IgGs асссеянели продемонстрировали реактивность против фосфорилированного пептида CBTAU22.1, и один был реактивным к нефосфорилированной CBTAU27.1 (Рисунок 4A). Дополнительное подтверждение было завершено с использованием кривой концентрации тех же рекомбинантных образцов антител против пептидов, определенных на первоначальном экране, и дополнительного пептида в качестве отрицательного контроля(рисунок 4В). Для каждого положительного удара отдельный плазмидный клон был изолирован от преобразованного пула и подтвержден тем же методом ELISA. Только клон 34 показано данные, представленные, и в то время как реактивность к нефосфорилированной CBTAU27.1 было подтверждено, более низкая сродство связывания также наблюдается с фосфорилированным 27,1 пептид (Рисунок 4C).

Рисунок 1 : Изоляция антиген-реактивных одиночных клеток через цитометрию потока. (A) Показано репрезентативное участок, где популяция лимфоцитов содержалась в нарисованных воротах. (B) Ворота на основе переднего и бокового рассеяния (FSC-высота v FSC-ширина (R3); SSC-высота v SSC- ширина (R4)) были использованы для исключения дублетов. Только ячейки внутри нарисованных ворот оценивались в последующих участках. Клетки DAPIбыли признаны мертвыми и исключенными (R5). В этом эксперименте были допрошены 3,7 х 107 "живых" клеток. Большинство живых клеток были В-клетки (CD19)и IgGq клетки (4.66%) были изолированы от этой популяции (R6). Лучшие 43,2% CD27-выражающие ячейки памяти были включены в ворота отбора (R7). (C) Квадрант 2 (No2) содержал клетки, реактивные как помеченные антигены (пептид-APC v пептид-PE), и эти клетки были отсортированы и восстановлены индивидуально в 96-хорошую пластину. Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой цифры.
{kind=link}

Рисунок 2 : Усиление ПЦР восстанавливает тяжелую и легкую переменную последовательность IgG для клонирования. (A) Как тяжелые (VH) и легкие (Vk или Vl) переменные области цепи восстанавливаются с помощью вложенных реакций ПЦР. Шаг I праймеры усиливают вперед от родной последовательности лидера и обратного изнутри постоянной области. Несколько стрелок представляют собой пул между 7-12 грунтовки, которые используются для обеспечения широкого охвата возможных зародышевых линий. Шаг II праймеры вложены, специфические для крайних концов переменной открытой рамки чтения, и добавить последовательность к концам амблонов, которые гомологичны к соседней последовательности в векторе выражения. (B) Связной состоит из постоянной световой цепи (Ck или Cl), а затем тяжелые цепи промоутер и неродной сигнал пептид последовательности. Ампликоны, связующие и плазмидные позвоночника одновременно сливки через перекрывающиеся гомологичные последовательности, генерируемые во время шага II PcR усиления, и изолированы как нетронутыми плазмиды. Рекомбинантные тяжелые и легкие цепи в конечном векторе экспрессии управляются независимыми (и идентичными) промоутерами ЦМВ и переводятся как отдельные белки. Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой цифры.
{kind=link}

Рисунок 3 : Тяжелые и легкие ампиконы переменной цепи IgG извлекаются из одиночных ячеек. Вложенные реакции ПЦР (Шаг II) были непосредственно загружены на 1% геля агарозы и визуализированы. Тяжелая цепь (H) и каппа световой цепи (L) ПЦР реакции из той же ячейки были загружены в соседних скважинах так парные ампликоны были легко наблюдаться. Успешные тяжелые и легкие цепи продукции примерно 400 bp и 350 bp соответственно. Успешное восстановление парных ампелонов обозначается (кв). Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой цифры.
{kind=link}

Рисунок 4 : Антигенная специфичность подтверждается рекомбинантными IgGs. Плазмиды, содержащие восстановленные тяжелые и световые цепные последовательности, трансфицируются в клетки HEK для выражения. Через четыре дня рекомбинантные антитела оцениваются ELISA на реактивность. (A) Концентрация IgG в уточненных носителях была измерена и разбавлена до 20 мкг/мл. Бассейн пептидов, используемых для сортировки, был индивидуально оценен с использованием 40 рмоль на скважину в стрептавидиновых пластинах. Все IgGs были протестированы в дублирующих скважинах. (B) Клоны, которые отображаются OD450 измерения йgt;0.5 были переоценены (клоны #32, #34, #35) с помощью 1:5 разбавления шаги против пептидов, определенных на экране. (C) После подтверждения в качестве хита, один клон плазмида был изолирован от клона #34 преобразованы перевязки, транссексуалов, и подтверждено ELISA. То же самое было сделано для клонов #32 и #35 (данные не показаны). Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой цифры.
{kind=link}
Обсуждение
Представленный здесь метод сочетает в себе цитометрию потока и клонирование одноклеточных клеток, а методы, которые мы описываем здесь, мы основыем на методах, ранее разработанных Тиллером и коллегами11. Их работа описывает восстановление и клонирование моноклональных антител для изучения репертуара В-клеток у людей на уровне отдельных клеток. Мы адаптировали основные компоненты их процесса, чтобы позволить восстановление антиген-специфических моноклональных антител из популяции клеток памяти В, в том числе многоступенчатой стратегии праймеризации усиления. Основной модификацией является добавление маркированного антигена "приманка". Дополнительные адаптации к опубликованному протоколу, в том числе (но не ограничиваясь) изменение хребета вектора клонирования в одно выражение плазмида, дополнительное охват зародышевой линии (как последовательность лидеров, так и рамка 1), трансфекция суспензии HEK293 клеток для более высокой экспрессии, а также использование полимеразы высокой точности во время усиления ПЦР.
Для описанных здесь методов наиболее важные шаги находятся вблизи перехода между цитометрией потока и клонированием одноклеточных ячеек. Во-первых, необходимо правильное размещение отсортированных одиночных ячеек в тарелку. Правильное настройка задержки сбросом на сортаторявляется ключевым шагом. Экологические факторы, такие как низкая влажность, также должны быть приняты во внимание, как мы обнаружили, что наши восстановления эффективности значительно снизиться, если статический пистолет используется на целевой сортировки пластин. После размещения клеток, пластины центрифугируются, чтобы убедиться, что клетки связались с 10 зл буфера в нижней части скважин. Все эти меры имеют решающее значение для молекулярной биологии часть, чтобы быть успешным. Если условия являются неоптимальными для обратной реакции транскрипции одной клетки, pcR усиление даже к-актин может быть трудным. Сила представленного метода заключается в способности допрашивать миллионы клеток и сортировать только те, которые соответствуют критериям реактивности антигена против антигена выбора. Это требует, чтобы соотношение сигнала к шуму было как можно выше, что делается путем оптимизации концентрации приманки перед сортировкой. Двойная этикетка приманки используется для уменьшения восстановления ложноположительных клеток, которые могут произойти, если один из флюорофоров имеет высокий фон. Использование более чем одного антигена приманки может сделать сигнал: оптимизация шума сложнее, но возможность допроса нескольких пептидов одновременно является еще одним преимуществом этого метода.
При низкой эффективности восстановления используется набор вложенных праймеров с актином, которые подтверждают наличие шаблона cDNA. Недостатком этого подхода является то, что трудно определить, не было ли ячейки, присутствуютвой в колодце или обратная реакция транскрипции не удалось, что делает устранение неполадок трудно. Иногда происходит обратное, и эффективность выше, чем ожидалось. Типичное восстановление для тяжелой цепи составляет от 30-45% и световые цепи выше, между 40-60%. Каждая реакция ПЦР (Шаг I и II) использует 50 циклов для усиления из одной ячейки. Большое количество общих циклов также делает этот метод восприимчивым к загрязнению. Последовательность анализа ампиконов может быть использована для определения того, загрязнитель был введен, или необычно высокий уровень восстановления был законно достигнута.
Полезность этого метода для восстановления местных человеческих антител против антигена выбора имеет несколько существенных ограничений. Во-первых, только растворимые белки, которые могут быть помечены могут быть использованы в качестве "приманки" для потока цитометрии. Для видов, представленных в репрезентативных результатах, мы использовали серию синтезированных перекрывающихся пептидов из тау-белка, так как использование всего белка оказалось трудным. Использование линейных пептидов, вероятно, не является оптимальным, так как это может ограничить выявление антител против нелинейных или структурных эпитопов. Тем не менее, мы смогли определить несколько уникальных антител против Тау, и они в настоящее время проходят дальнейшую оценку8,9. Другим ограничением является низкая пропускная стоимость восстановления молекулярной биологии и клонирования ИгГов. Первоначальная стратегия сортировки позволяет скрининг миллионов клеток, но последующая обработка молекулярной биологии состоит из нескольких этапов. Продолжающиеся усилия по оптимизации для внедрения новых технологий, таких как Gibson Assembly10, упростили несколько шагов, но узким местом остается клонирование отдельных тяжелых и легких цепных пар.
Метод "BSelex" использует BCR, выраженный на поверхности клеток памяти В, чтобы определить клетки, которые отображают реактивность антигена, а затем восстановить эти отдельные клетки через цитометрию потока11,12. Из-за этой зависимости от BCR, метод ограничен отсеком клетки памяти B, и не захватывает клетки высвобождения антител (ASCs) such as plasmablasts. Тем не менее, этот подход может быть выгодным для восстановления более широкого репертуара антиген-специфических иммуноглобинов по сравнению с ASCs. Исследования вакцинации против гриппа показывают, что в то время как реактивные ASCs может доминировать небольшое количество расширенных клонов Клеток B, антиген-специфической памяти B клеточной популяции редко клональный12. Рецептор Т-клеток (ТКР) на поверхности Т-клеток обладает сходством с рецептором В-клеток в расположении генов и рекомбинации для максимального разнообразия. Одноклеточные подходы, аналогичные тому, что описано в методе, представленном здесь, были разработаны для оценки репертуара Т-клеток, включая восстановление и одноклеточное клонирование альфа- и бета-цепей13. Тем не менее, распознавание TCR требует, чтобы пептиды были представлены молекулами MHC, добавляя значительную сложность маркированному подходу к приманке для определения пептид-специфических Т-клеток. В отличие от В-клеток, Т-клетки не проходят созревание сродства всей переменной области, поэтому идентификация короткой области CDR3 и некоторой фланговой последовательности — это все, что требуется для идентификации и восстановления. Наконец, этот метод описывает идентификацию молекул IgG с использованием цитометрии потока, но можно также использовать альтернативные фенотипические маркеры для идентификации В-клеток различных изотипов.
В настоящее время разрабатываются методы, связанные с подключением огромного объема данных, собранных в ходе секвенирования следующего поколения, с функциональным анализом специфичности антигена, однако эти методы все еще совершенствуются. Несмотря на свои ограничения, описанный здесь метод использовался для выявления антителс потенциальной терапевтической ценностью как для инфекционных, так и для неинфекционных заболеваний 8,14,15,и представляет собой надежный подход к восстановлению соответствующих антиген-специфических IgGs от людей, без обширных манипуляций В-клеток или обширной культуры клеток.
Раскрытие информации
Авторам нечего раскрывать
Благодарности
Авторы хотели бы поблагодарить Люси Шаммас, Марта Коста, Джули Ким, Нэнси Эредиа и Джереми Маседо за тщательное тестирование многих модификаций метода и уточнение текущей платформы BSelex.
Материалы
| Name | Company | Catalog Number | Comments |
| MoFlo Astrio EQ, Cell Sorter | MoFlo | cell sorting | |
| Biotinylated peptides | New England Peptide | Cell sorting "bait" | |
| Micro Bio-spin P30 gel column | BioRad | #7326223 | |
| Streptavidin (R-PE) | Thermo Scientific | SA10041 | |
| Streptavidin (APC) | Thermo Scientific | S32362 | |
| CD22 MicroBeads, human | Miltenyi | #130-046-401 | |
| LS columns | Miltenyi | #130-042-401 | |
| Pre separation filters | Miltenyi | #130-041-407 | |
| PerCp-Cy5.5 mouse anti-human CD19 | BD Biosciences | BD#340951 | |
| PE-Cy7 mouse anti-human CD27 | BD Biosciences | #560609 | |
| FITC mouse anti-human IgG | BD Biosciences | #560952 | |
| DAPI | Life Technologies | #D21490 | |
| RNaseOUT | Life Technologies | #10777-019 | |
| Bovine serum albumin, Fraction V | Sigma | #A4503 | |
| RPMI media | Hyclone | #SH30096.01 | |
| HI FBS (Fetal bovine serum) | Invitrogen | #10082147 | |
| Mastercycler Gradient | Eppendorf | Model #6325 | PCR machine |
| PCR 96-well plates | Phenix | MPS-500 | |
| PCR Plate mats | Phenix | SMX-PCR96 | |
| Advantage UltraPure PCR Deoxynucleotide Mix | Clontech | #639125 | |
| Superscript IV First-strand synthesis system | Invitrogen | #18091050 | |
| Phusion High Fidelity Polymerase | Thermo Scientific | F-531-L | |
| Platinum polymerase | Invitrogen | #10966108 | |
| Nuclease-free water | QIAGEN | #129114 | |
| Oligonucleotide primers | IDT | assorted | Primers for all PCR steps |
| Gel Extraction Kit | QIAGEN | #28706 | |
| PCR Purification kit | QIAGEN | #28106 | |
| Miniprep Kit | QIAGEN | #27106 | |
| NEBuilder HiFi DNA Assembly Cloning Kit | New England Biolabs | E5520S | Gibson assembly |
| Expi293 Expression Media | Invitrogen | #A14351-01 | |
| ExpiFectamine 293 Transfection Kit | Invitrogen | #A14525 | |
| Opti-MEM Media | Invitrogen | #31985070 | |
| CO2 incubator (Multitron) | |||
| Octet RED384 System | Pall/ Forte Bio | ||
| Pierce Streptavidin Coated High Binding Capacity Clear 96-well Plates | Thermo Scientific | #15500 | |
| Corning Costar 96-well Polystyrene Plate High Binding Half Area | Corning | #3690 | |
| Goat Anti-human IgG Fab Secondary Antibody | Jackson Labs | #109-036-097 | |
| Goat Anti-mouse HRP Secondary Antibody | Jackson Labs | #115-035-072 | |
| SureBlueTM TMB Microwell Peroxidase Substrate (1-Component) | KPL | #52-00-03 | |
| TMB Stop Solution | KPL | #50-85-06 |
Ссылки
- Su, K. Y., Watanabe, A., Yeh, C. H., Kelsoe, G., Kuraoka, M. Efficient Culture of Human Naive and Memory B Cells for Use as APCs. Journal of Immunology. 197 (10), 4163-4176 (2016).
- Corti, D., Lanzavecchia, A. Broadly neutralizing antiviral antibodies. Annual Review of Immunology. 31, 705-742 (2013).
- Steinitz, M., Klein, G., Koskimies, S., Makel, O. EB virus-induced B lymphocyte cell lines producing specific antibody. Nature. 269 (5627), 420-422 (1977).
- Bloom, A. D., Nakamura, F. T. Establishment of a tetraploid, immunoglobulin-producing cell line from the hybridization of two human lymphocyte lines. Proceedings of the National Academy of Sciences of the United States of America. 71 (7), 2689-2692 (1974).
- Olsson, L., Kaplan, H. S. Human-human hybridomas producing monoclonal antibodies of predefined antigenic specificity. Proceedings of the National Academy of Sciences of the United States of America. 77 (9), 5429-5431 (1980).
- Lechner, J., et al. Alterations in Circulating Immune Cells in Neovascular Age-Related Macular Degeneration. Scientific Reports. 5, 16754(2015).
- Loken, M. R., Brosnan, J. M., Bach, B. A., Ault, K. A. Establishing optimal lymphocyte gates for immunophenotyping by flow cytometry. Cytometry. 11 (4), 453-459 (1990).
- Pascual, G., et al. Immunological memory to hyperphosphorylated tau in asymptomatic individuals. Acta Neuropathologica. 133 (5), 767-783 (2017).
- van Ameijde, J., et al. Enhancement of therapeutic potential of a naturally occurring human antibody targeting a phosphorylated Ser(422) containing epitope on pathological tau. Acta Neuropathologica Communications. 6 (1), 59(2018).
- Gibson, D. G., et al. Enzymatic assembly of DNA molecules up to several hundred kilobases. Nature Methods. 6 (5), 343-345 (2009).
- Tiller, T., et al. Efficient generation of monoclonal antibodies from single human B cells by single cell RT-PCR and expression vector cloning. Journal of Immunological Methods. 329 (1-2), 112-124 (2008).
- Wrammert, J., et al. Rapid cloning of high-affinity human monoclonal antibodies against influenza virus. Nature. 453 (7195), 667-671 (2008).
- Han, A., Glanville, J., Hansmann, L., Davis, M. M. Linking T-cell receptor sequence to functional phenotype at the single-cell level. Nature Biotechnology. 32 (7), 684-692 (2014).
- Apetri, A., et al. A common antigenic motif recognized by naturally occurring human VH5-51/VL4-1 anti-tau antibodies with distinct functionalities. Acta Neuropathologica Communications. 6 (1), 43(2018).
- Jones, H. G., et al. Structural basis for recognition of the central conserved region of RSV G by neutralizing human antibodies. PLoS Pathogens. 14 (3), 1006935(2018).
- Janssen Vaccines & Prevention B.B. US patent. , U.S. Patent Application No. 20170210787, Published July 27, 2017 (2017).
Перепечатки и разрешения
Запросить разрешение на использование текста или рисунков этого JoVE статьи
Запросить разрешениеThis article has been published
Video Coming Soon
Авторские права © 2025 MyJoVE Corporation. Все права защищены