
Method Article
전 세포 전압 클램프 기록을위한 전기 생리 학적 방법
요약
Drosophila melanogaster photoreceptor의 전체 세포 기록은 다양한 조건 에서 자연스런 어두운 범프, 양자 범프, 빛에 대한 거시적 반응 및 전류 - 전압 관계를 측정 할 수있게합니다. D. melanogaster 유전자 조작 도구와 결합하여이 방법은 유비쿼터스 이노시톨 - 지질 신호 전달 경로와 그 표적 인 TRP 채널에 대한 연구를 가능하게합니다.
초록
Drosophila melanogaster 광 수용체의 전체 세포 전압 클램프 기록은 단일 분자 수준에서 이노시톨 - 지질 신호 및 과도 수용 수용체 (Transient Receptor Potential, TRP) 채널을 연구하기 위해 D. melanogaster 분자 유전학의 사용을 가능하게하는 무척추 동물 영상 전달 분야를 혁신했습니다. 소수의 연구실에서는이 강력한 기법을 익혔으며 고도로 제어 된 조건에서 빛에 대한 생리적 반응을 분석 할 수 있습니다. 이 기술은 세포 내 및 세포 외 매체를 제어 할 수 있습니다. 막 전압; 및 다양한 이온 성 또는 pH 지시약과 같은 약리학 적 화합물의 세포 내 및 세포 외 매체로의 신속한 적용을 포함한다. 예외적으로 높은 신호 대 잡음비로,이 방법은 어두운 자발적 및 광 유도 단일 전류 (자발적 및 양자 범프)와 거시적 인 빛 유도 전류 (LIC)를 죄로부터 측정 할 수 있습니다gle D. melanogaster 광 수용체. 이 프로토콜은 전기 생리 학적 및 광학적 기록을 모두 포함하는이 기술을 수행하는 데 필요한 모든 주요 단계를 아주 자세하게 설명합니다. 목욕실에서 손상되지 않고 생존 가능한 생체 외 고립성 구강 내막염 제거를위한 파리 망막 해부 절제술이 기술되었다. 전체 셀 및 형광 이미징 측정을 수행하는 데 필요한 장비도 자세히 설명합니다. 마지막으로, 확장 된 실험 동안이 섬세한 준비를 사용함에있어서 함정이 설명됩니다.
서문
약 100 년 전에 시작된 초파리 Drosophila melanogaster ( D. melanogaster) 에 대한 광범위한 유전 연구는 D. melanogaster fly를 복잡한 생물학적 과정의 유전 적 해부를위한 매우 유용한 실험 모델로 확립했다. 아래에서 설명하는 방법은 D. melanogaster 분자 유전학의 누적 된 출력과 전체 셀 패치 클램프 녹음의 높은 신호 대 잡음 비율을 결합합니다. 이 조합은 네이티브 환경과 단일 분자의 최고 해상도 모두에서 이노시톨 - 지질 시그널링 및 TRP 채널 조절 및 활성화의 모델로서 D. melanogaster 광 변환의 연구를 허용한다.
D. melanogaster photoreceptors에 대한 전체 세포 기록 방법의 적용은 무척추 동물의 phototransduction에 대한 연구에 혁명을 일으켰습니다. 이 방법은 Hardie 1 과 indep가 개발했습니다.Ranganathan과 동료들에 의해 2 ~ 26 년 전에 끝내었고 , D. melanogaster 의 광범위한 유전자 조작 도구를 이용하고 phototransduction과 inositol-lipid signaling의 메커니즘을 밝히기 위해 사용되었다. 처음에는이 기술은 해부학 과정에서 빛 민감도가 급격히 감소하고 개복 수확량이 낮아 세부적인 정량 연구가 어려웠습니다. 나중에 패치 피펫에 ATP와 NAD를 첨가하면 장기간의 정량적 레코딩을위한 준비의 적합성이 크게 증가했습니다. 그 후, 분자 수준에서 신호 전달 메커니즘의 광범위한 특성이 실현되었다.
현재, D. melanogaster 광전 변환은 phosphoinositide 신호 및 TRP 채널이 단일 분자 분해능으로 생체 외에서 연구 될 수있는 몇 안되는 시스템 중 하나입니다. D. melanogaster 광전 변환과 저를 만듭니다 .thodology는이 메커니즘을 매우 민감한 모델 시스템으로 연구하기 위해 개발되었습니다. 이 프로토콜은 D. melanogaster 망막을 해부하고 주위 안료 (glia) 세포에서 분리 된 ommatidia를 기계적으로 제거하는 방법을 설명합니다. 이것은 photoreceptor 세포 몸에 기가 씰과 전체 세포 패치 클램프의 형성을 가능하게합니다. 다행히 대부분의 신호 전달 단백질은 횡문근 막에만 국한되며 확산되지 않습니다. 또한 신호 구획과 세포 사이에는 칼포틴 (calciumphotin)이라고 불리는 움직일 수없는 칼슘 2+ 완충액이 있으며, 미포 질에서 Na + / Ca 2+ 교환기 (CalX)의 발현 수준이 높습니다. 함께 rhabdomere, calphotin 완충액 및 CalX의 높은 발현에 단백질 감금은 필수 구성 요소의 손실없이 상대적으로 연장 된 ( 즉 ~ 20 분까지) 전체 세포 녹음을 허용합니다빛에 대한 높은 감도를 유지하면서 광전도 공정의 다음 프로토콜은 고립 된 눈꺼풀 아래를 얻는 방법과 광 변환 캐스케이드의 고유 특성을 보존하는 것처럼 보이는 전체 세포 기록을 수행하는 방법을 설명합니다. 분리 된 바퀴벌레 ( Periplaneta americana ) 6 및 크리켓 ( Gryllus bimaculatus ) 7 외피에 대한 전체 세포 패치 클램프 실험은 D. melanogaster 에 대해 기술 된 것과 유사하게 수행 되었다. 또한 파일 클램프 ( 리마 scabra )와 가리비 ( Pecten의 irradians )의 dissociated photoreceptors에 대한 패치 클램프 실험은 전체 셀 8 및 단일 채널 측정을 허용 디 melanogaster 에 실시와 약간 다른 방식으로 수행되었다 9 . 여기,이 기술을 사용하여 디 melanogaster 에서 얻은 주요 업적이 설명되어 있습니다. 토론 나는이 기법의 몇 가지 함정과 한계에 대한 설명은 없습니다.
프로토콜
1. 시약 준비
참고 : 표 1-4 의 지침에 따라 모든 솔루션을 준비하십시오.
- Extracellular Solution (ES 또는 ES-0Ca 2+ ; 필요에 따라 표 1 참조)과 함께 10 mL 주사기를 채우고 얼음 위에 보관하십시오.
- Trituration Solution (TS; 표 2 참조, 즉 ES 또는 ES-0Ca 2+ + FBS와 자당) 한 병을 준비하고 얼음 위에 보관하십시오.
- 실험을위한 충분한 ES를 준비하고 필요할 때까지 얼음 위에서 유지하십시오.
참고 : 연속 목욕 관류가 필요하지 않으므로 ES의 부피는 수십 mL 이상이어야합니다. - 22 μm PVDF 필터를 사용하여 전극 팁이 길게 뾰족해진 1 mL 주사기에 세포 내 용액을 넣으십시오 ( 표 3 또는 표 4 참조). 얼음 위에 올려 놓으세요.
2. 해부 도구의 일반 설정

그림 1 : 격리 된 개안 약품 준비에 필요한 도구와 장치. 사진은 위의 상세한 프로토콜에 설명 된 바와 같이 격리 된 수정란 작성을 위해 필요한 다양한 장치를 보여줍니다. 두 개의 비이커는 튜브 ( C )에 연결된 분쇄 피펫 ( D )을 청소하기 위해 물 ( A )로 채우고 다른 하나는 에탄올 ( B )로 채 웁니다. 해부 도구는 2 쌍의 벌금과 1 쌍의 거친 족집게 ( F )와 망막 scooper ( E )입니다. 망막 스커 퍼를 준비하기 위해 바늘 ( I )의 상단을 평평하게하기 위해 바이스 ( G )로 작용하는 두 개의 선반 도구 사이에서 마이크로 해부 바늘 ( H )을 누릅니다. 그런 다음 그것은 긴 pi에 연결됩니다 접착제 ( J )를 사용하여 바늘 홀더 ( E )에 장착 된 금속의 에이스. 면도날 칩 및 홀더 : 면도날 홀더 ( K )를 사용하여 면도날의 작은 삼각형 칩을 분리하여 장착하십시오. 해부 작용 영역은 2 개의 광 가이드 ( N )를 갖는 쌍안경 ( L ) 및 시원한 적색 광원 ( M )으로 구성된다. 트위저 ( F ), 망막 스커퍼 ( E ), 비커 ( A 및 B ), 적색 필터 ( Q )가있는 플래시 라이트, 섬세한 와이퍼 ( R ) 등 해부 도구가 쌍안의 양쪽에 있습니다. ES, FBS - ES, 세포 내 솔루션 주사기, 60mm 페트리 접시 ( S ), 그리고 전극 홀더 ( P )와 얼음 양동이도 테이블에 배치됩니다. 기록 전극은 수평 풀러 ( T )를 사용하여 당겨진다.e.com/files/ftp_upload/55627/55627fig1large.jpg "target ="_ blank ">이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
- retinae를 분리하기 위해 망막 scooper를 구성하십시오.
- 두 개의 바이스 턱 사이에 마이크로 해부 바늘 (곤충 바늘, 길이 12mm, 직경 0.1mm)의 지점 (1-4mm)을 삽입하고 작은 망치로 두드려 평평하게하십시오.
- 마이크로 해부 식 바늘 홀더에 편평한 바늘을 장착하십시오 ( 재료 표 참조).
- 한 쌍의 족집게를 사용하여 ~ 2.5 mm의 곡률을 가진 후크를 형성하기 위해 바늘의 평평한 끝을 휘게하십시오.
- 당김 분리를위한 마찰 피펫을 만듭니다.
- 개방형 화염 위에 1.2 x 0.68 mm (OD x ID) 유리 모세관을 놓고 열을 줄이기 위해 그것을 닦습니다.
- 현미경으로 모세관 개구의 크기를 측정합니다. 생성 된 구강 분열 pipettes을 seve로 정렬n 개의 그룹 ( 예 : 0.2-0.5 mm). 별도의 적합한 용기 ( 예 : 시험관)에 보관하십시오.
- 하나의 작은 비이커에 이중 증류수 (DDW)를 채우고 다른 작은 비이커에 에탄올 70 %를 채우십시오. 각 비이커를 파라핀 필름 시트로 덮으십시오.
- DDW 비커를 덮고있는 파라핀 필름 시트에 하나의 작은 구멍을 뚫습니다 ( 그림 1A 참조 ).
- 에탄올 비커를 덮고있는 파라핀 필름 시트에 7 개의 작은 구멍을 뚫고 0에서 6까지의 구멍에 번호를 매 깁니다 ( 그림 1B 참조).
- 가장 큰 구멍을 가진 피펫은 0 번 구멍에 위치하고 가장 작은 구멍을 가진 피펫은 6 번 구멍에 위치하도록 각 파라핀 필름 구멍에 각 크기 그룹의 하나의 둥근 마찰 피펫을 놓습니다.
- 플라스틱 200 μL 피펫 팁을 길이 35 cm, 1.57 x 1.14 mm (OD x ID)의 폴리에틸렌 튜브에 연결하십시오. 코 넥크가장 큰 구멍 ( 예 : 0.46 - 0.5mm)을 가진 눈꺼풀 연화 피펫 중 하나에 튜빙의 다른 끝을 놓고 마찰 피펫의 뾰족한 끝 부분이 튜빙 안으로 향하지 않도록하십시오.
- 유리 모세관이 들어있는 1 x 0.58 mm (OD x ID) 붕규산 필라멘트에서 패치 클램프 피펫을 당겨 전체 세포 기록 피펫을 준비합니다.
참고 : 칼륨 글루코 네이트 기반 세포 내 용액 (IS1)을 사용하는 경우 피펫의 저항은 8-15 MΩ이어야합니다. 임의의 적합한 패치 피펫 풀러 (patch peller pulller )가 사용될 수있다 (예를 들어, 표 (Table of Materials ) 참조). 화염 연마는 필요하지 않습니다. - 용융 파라핀 또는 고진공 실리콘 그리스를 사용하여 커버 슬립 (재료 표 참조)을 욕조 챔버 바닥에 고정하여 기록 챔버 ( 그림 2 참조)를 준비하십시오. 전극 접근 및 재관류를 허용하는 임의의 수제 또는 상업용 챔버를 사용하십시오.
- 거꾸로 된 현미경의 무대에 목욕을 마운트합니다. 관류 시스템 튜브, 흡입 시스템 및 그라운드 ( 예 : Ag-AgCl 와이어 / 펠렛)를 욕조 에 놓으십시오 (재료 표 및 그림 2 참조).
- 망막 해부 및 ommatidia 격리 단계 동안 사용하기 위해 작업 표면에 좋은 # 5 족집게 ( 그림 1 )의 두 쌍을 놓으십시오.
3. D. melanogaster 양육
- Raise D. Melanogaster 는 19-24 ° C에서 표준 옥수수 음식을 담은 병에서 낮은 인구 밀도 ( 즉 , 6 온스 병에서 ~ 20 파리)로 날아 오른다.
참고 : 어두운 적응 파리에서 작업하는 것이 바람직합니다. 빛에 대한 높은 민감도를 유지하기 위해 돌연변이 파리에서 망막 변성을 방지하고 다양성을 줄입니다. - 실험 전 적어도 24 시간 동안 어둠 속에서 파리를 뒤덮습니다.
참고 : 실험에 사용 된 파리는ntly eclosed (2 시간 이내), 여전히 부드럽고, 창백하고, meconium을 표시합니다. 개미는 또한 빛에 대한 감수성이 10 세에 의존적이지만 번데기로 쉽게 준비 할 수 있습니다.
4. 망막 해부 및 당뇨병 격리 : 옵션 1
참고 : 준비를 올바르게 볼 수있는 증폭을 사용하여 입체 줌 현미경에서 다음 단계를 모두 수행하십시오 ( 그림 1 참조).
- 뒤집힌 60mm 배양 접시에 ES-0Ca 2+ 4 방울과 TS 용액 1 방울을 놓습니다.
- 거친 족집게를 사용하여 날개 또는 몸체로 새 비행을 포착합니다 (<2 시간 후 eclosion). 이 시점부터 모든 절차를 20 ± 1 ° C의 어두운 조도에서 신속하게 수행하십시오.
- 거친 족집게로 파리를 잡는 동안 첫 번째 쌍의 섬세한 족집게를 사용하여 파리 머리를 신체에서 분리하십시오. 서브첫 번째 ES-0Ca 2+ 드롭에서 헤드를 움직입니다.
- 두 번째 쌍의 미세 핀셋을 사용하여 화살 평면을 따라 머리를 반으로 해부합니다. 이 단계가 끝날 때 두 눈이 손상되지 않도록하십시오.
- 헤드의 절반을 두 번째 ES-0Ca 2+ 드롭으로 옮기고 나머지 절반은 세 번째 ES-0Ca 2+ 드롭으로 옮깁니다.
- 미세 핀셋을 사용하여 가능한 한 눈 주위의 많은 조직을 제거하고 망막에 해를 입히지 않도록하십시오.
- 섬세한 핀셋으로 각막 가장자리를 단단히 잡고 망원경으로 망막을 퍼덕 거리십시오.
참고 :이 단계가 완료되면 각막은 손상되지 않은 상태로 유지되며 손상되지 않은 망막과 분리됩니다. - DDW로 튜빙에 연결된 마찰 피펫을 헹구고 네 방울에서 소량의 ES-0Ca 2+ 로 피펫을 채 웁니다.
참고 :이 단계는 새로운 수정란 제거 피펫을 사용하거나 제거 할 때마다 수행해야합니다.m 에탄올 비커 (피펫을 채우는 용액은 망막이 잠긴 용액과 일치해야 함). - 부드럽게 피펫으로 고립 된 망막을 그리기 위해 입으로 대기음. 피펫에 공기 방울이 생기지 않도록 각별히주의하십시오.
- 격리 된 망막을 TS 방울로 옮깁니다. 두 번째 눈에도 4.6-4.10 단계를 수행하십시오.
- 섬세한 물티슈를 사용하여 ES-0Ca 2+ 의 물방울을 닦아 내고 페트리 접시에 두 망막이 모두 들어있는 TS 방울을 남겨 둡니다. 페트리 접시 상단에 여섯 방울의 TS를 더 넣으십시오. 두 망막을 다른 TS 방울 중 하나로 옮깁니다.
- 마찰 작은 피펫을 작은 지름의 피펫으로 교체하십시오. 피펫을 채우기 위해 솔루션으로 TS를 사용하여 단계 4.8에서 설명한대로 린스.
- 신속하고 반복적으로 대기 중 망막 전체에서 색소 세포를 제거한 격리 된 개 피부의 분리를 시작하기 위해 솔루션에서 두 망막을 모두 대기음으로내어 놓습니다.
참고 : 격리 된 옴단점은 TS 강하에서 볼 수 있으며 격리 과정이 진행됨에 따라 TS 강하가 덜 반투명하게됩니다. - 남은 망막을 다음 TS 드롭으로 옮깁니다. 전 TS 드롭 (격리 된 ommatidia를 포함)으로 피펫을 채우고 목욕 방으로 드롭을 expirate.
- 최대 고립 고립증을 얻으려면 4.12-4.14 단계를 반복하십시오. 격리 된 개머리판이 가라 앉고 목욕실 바닥에 달라 붙을 때까지 약 1 분 정도 기다립니다.
- 재관류 시스템을 사용하여 1.5 MM 칼슘 2 + 와 목욕탕 챔버에 ES - 0Ca 2 + 의 흐름을 시작합니다. 챔버가 바닥에서 상단으로 용액으로 완전히 채워지고 바닥이 용액에 완전히 잠겨 있는지 확인하십시오. 목욕을 4 ~ 5 번 씻으십시오.
5. 망막 절개술과 당뇨병 격리 : 옵션 2
참고 : 입체 줌 현미경을 사용하여 다음 단계를 모두 수행하십시오.( 그림 1 참조).
- 면도날 칩과 홀더를 준비하십시오. 면도날 홀더 ( 그림 1 )를 사용하여 면도날의 작은 삼각형 칩을 분리하여 장착하십시오.
- 제조 업체의 지침에 따라 실리콘 해부 접시 / 블록을 준비합니다 ( 재료 표 참조).
- 해부학을 위해, 실리콘 해부 블록에 ES 솔루션의 큰 방울 (<0.5 ML)을 만듭니다. TS 솔루션의 두 개의 "저장소"방울 (~ 50 μL 각각)을 60 mm 페트리 접시에 추가하십시오
- 얼음으로 덮인 유리 튜브에 새롭게 닫힌 (2 시간 이내) 플라이를 고정시키고 핀셋을 사용하여 날개로 잡아 당깁니다. 이 시점부터 모든 절차를 20 ± 1 ° C의 어두운 조도에서 신속하게 수행하십시오.
- 족집게로 파리를 잡고, 홀더에 장착 된 면도날 칩을 사용하여 파리의 머리를 자릅니다. 벌레 핀 (12mm 길이,지름 0.1mm)을 핀셋과 연결하고 눈 사이에 머리를 관통시킵니다.
- 잠시 머리를 70 % 에탄올에 담그십시오. 이것은 기포가 머리 / 눈 표면 상에 형성되는 것을 방지한다. 실리콘 해부 접시에 ES 드롭 아래 머리를 핀.
- 눈의 앞쪽 여백 라인을 따라 톱질 동작을 사용하여 면도날 칩을 사용하여 두 눈을 잘라냅니다.
- 각막의 모서리를 단단히 조여줍니다.
- scooper를 사용하여 망막을 스쿠프합니다.
참고 :이 단계가 완료되면 각막은 손상되지 않은 상태로 유지되며 손상되지 않은 망막과 분리됩니다. - 망막을 손상시키지 않으면 서 핀셋과 스커 퍼를 사용하여 부착 된 공기 주머니와 과도한 뇌 조직을 부드럽게 제거하십시오.
참고 : 격리 retinae의 준비는 또한 비 특정 retinae 단백질 11 , 전체 탑재 histology 및 전체 망막 이미징의 웨스턴 블럿 분석에 유용합니다. 패치 클램프 녹음은 ph전체 망막의 otoreceptors 12 . - 가장 큰 직경을 가진 분쇄 피펫을 가져 와서 튜빙에 연결하고 부드러운 흡입 ( 예 : 입에 의한)에 의해 페트리 접시에있는 저수지 방울 중 하나에서 작은 양의 TS로 피펫을 채우십시오. 새로운 당김 분쇄 피펫을 사용할 때마다이 단계를 수행하십시오.
- 부드럽게 입으로 두 retinae 위에 TS를 날려 부드럽게 흡입을 사용하여 피펫으로 고립 된 망막을 그립니다. 공기 방울이 피펫으로 들어 가지 않도록주의하십시오.
- 작은 방울 (~ 20 μL)을 형성하고, 저수지 방울 중 하나에서 TS로 한두 번 그들을 씻어 페트리 접시에 고립 된 망막을 전송합니다.
- 20-25 분 동안 어둠 속에서 망막을 품어주세요.
- ommatidia trituration 피펫을 작은 지름의 구멍이있는 피펫으로 교체하십시오 (피펫을 채우기 위해 4.6 단계의 저수지 방울 중 하나에서 새 TS를 사용하십시오).
- 급속하게격리 된 개머리판 독액 분리를 시작하기 위해 작은 방울 (~ 20 μL)로 두 망막을 기음과 만료하십시오.
참고 : 첫 번째 단계에서는 주변 색소 glia가 분해되어 용액에 눈에 띄는 작은 부스러기가 남습니다. - 상당한 양의 작은 찌꺼기가 쌓인 후에 많은 ommatidia가 분리되기 전에 저수지 방울 중 하나에서 신선한 TS를 사용하고 망막을 새로운 작은 방울로 옮깁니다.
- 직경이 작은 분쇄 피펫을 선택하고 백필을 채우고 분쇄하십시오.
참고 : 개머리판 구균이 분리되기 시작함에 따라 입체 현미경의 강력한 힘에 의해 그 긴 형태가 명확하게 보여야합니다. 필요한 경우, ommatidia의 좋은 수확량이 보일 때까지 더 작은 지름으로 분쇄 pipettes을 계속 변경하십시오 - ommatidia의 적당한 수확량이 보이고 드롭이 더 이상 반투명하지 않으면 피펫을 격리 된 개머리판을 포함한 전체 드롭으로 채우고 드롭을 부드럽게ES로 미리 채워진 욕실 챔버의 바닥.
- 격리 된 개머리판멍이 가라 앉고 목욕실의 바닥에 정착되도록 약 1 분 정도 기다리십시오.
- 관류 시스템을 사용하여 목욕탕으로 ES의 흐름을 시작합니다. 챔버가 바닥에서 위까지 용액으로 완전히 채워져 있고 바닥이 용액에 완전히 잠겨 있는지 확인하십시오. 목욕을 4 ~ 5 번 씻으십시오.
참고 : 이후, 새로운 피펫 피펫을 소개하기 전에 목욕을 잠시 플러시해야하지만, 연속 관류는 필요하지 않습니다.
6. 전체 셀 기록

그림 2 : 전기 생리학 및 광학 설정 개요. 설정에는 검은 색으로 칠해진 패러데이 새장 ( F )이 포함되어 있습니다. 앞면은 내부에 구리 메쉬가 달린 검은 커튼으로 덮여 있습니다. 이 설정━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━ 역 형광 현미경 ( A )은 방진 대 ( E )에 고정되어있다. 광 수용체 기록 장치는 관류 입력 ( Q ), 흡입 피펫 ( S ) 및은 - 염화 염화물 그라운드 ( P )가있는 수제 아크릴 유리 욕조 챔버 ( O )로 구성됩니다. 목욕 챔버는 수제 어댑터로 현미경 스테이지 ( T )에 장착됩니다. 레코딩 피펫은 은색 염화 와이어를 통해 아크릴 헤드 홀더 ( C )에 연결된 아크릴 유리 홀더에 연결됩니다. 헤드 스테이지는 정밀한 XYZ 기계식 미세 조작기 ( B )에 장착 된 거친 미세 조작기에 장착됩니다. 관류 시스템 ( D )은 주사기 세트로 구성되는 반면, 리퀴의 흐름d는 핀치 밸브 ( H )에 의해 제어됩니다. 랙은 패러데이 케이지 내의 모든 장비가 연결된 동일한 중앙 접지에 전기적으로 연결되어 있으며 패치 클램프 앰프 ( N ), 오실로스코프 ( J ), 펄스 / 함수 발생기 ( K ), A ~ D 변환기 ( L ), 관류 제어기 ( I ) 및 필터 휠 및 셔터 제어기 ( M )를 포함한다. 이미징 실험의 경우 냉각 된 (-110 ° C, 재료 표 참조) CCD 카메라가 측면 포트 ( G )를 통해 연결됩니다. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
{kind=link}
참고 : 다음 단계를 수행하는 동안 희미한 적색 조명 만 사용하고 조명에 대한 외부 노출은 최소화하십시오 ( 즉, 빠르게 작동하고개안을보기를 요구하지 않는 작업을 수행 할 때 개안을보기 위해 사용되는 해부 광원 및 챔버 적색 조명을 끕니다. 또한 표준 전기 생리 학적 프로토콜에 따라 다음 단계를 모두 수행하십시오.
- 거꾸로 된 현미경 (40X 대물 렌즈) 아래, 조심스럽게 입욕제를 검사하고 실험을위한 적절한 개안 약을 선택하십시오.
- 개 모세관의 외막이 부드럽고 손상되지 않도록하고, 장축이 전극 접근 방향 ( 그림 3A 참조 )에 대해 거의 직각을 이루며, 개 모세포의 원위부가 어떤 방향으로 둘러싸여 있지 않은지 확인하십시오 과잉 조직. 선택한 외과 의사를 대물 렌즈의 광학 축 (시야 중심)에 배치하여 균일 한 조명을 보장하십시오.
- 세포 내 용액 (IS1 또는 IS2)으로 패치 피펫을 채 웁니다.
참고 : To mea강도 응답 및 범프 분석이 IS1을 사용하는지 확인하십시오. 감광 채널의 역전 전위를 측정하려면 IS2를 사용하십시오. - 전극 홀더에 패치 피펫을 장착하십시오.
- 전극 홀더에 연결된 튜브를 통해 입으로 피펫으로 불어 넣어 정압으로 채 웁니다. 튜브 밸브를 닫아 압력을 유지하십시오.
- micromanipulator를 사용하여 전극을 욕조에 넣으십시오.
- ommatidium에서 작은 딤플 (패치 피펫의 긍정적 인 압력으로 인해)이 관찰 될 때까지 전극과 개미 사이에 접촉이 없도록 개나 개머리판의 원위부 가까이에 전극을 놓습니다.
- 기록 소프트웨어를 엽니 다 (재료 표 참조). "멤브레인 테스트 모듈"을 열어 2mV의 연속 제곱 전압 펄스를 100Hz 속도로 적용하십시오.
- 적절한 kno를 조정하여 접합 전위를 "0"으로 설정합니다.패치 클램프 증폭기에서 제곱 펄스의베이스를 "제로"전류로 설정하십시오
참고 : 전기 생리 학적 설정에는 증폭기 ( 예 : 2 단계 증폭)에 연결된 헤드 스테이지 ( 즉, 1 단계 증폭)가 포함됩니다. 증폭 된 아날로그 신호는 PC 컴퓨터에 설치된 소프트웨어로 제어되는 A / D 변환기를 사용하여 디지털 신호로 변환됩니다. - 전극 홀더에 연결된 튜브의 밸브를 열어 피펫의 양압을 해제하십시오. 부드럽게 세포 막에 피펫의 연결로 이어지는 튜브 빠는 피펫에 부정적인 압력을 만듭니다. 튜브 밸브를 닫아 압력을 유지하십시오.
- 컴퓨터 화면에서 본 전극 저항이 100 - 150 MΩ으로 올라 갔는지 확인하십시오. 전극 홀더에 연결된 튜브의 밸브를 수동으로 열어 피펫의 음압을 해제하십시오.
- 엘발열 저항이 적어도 1-2 GΩ으로 상승합니다.
참고 :이 시점에서 전극과 감광체 사이에 씰이 형성되었습니다. - 패치 클램프 앰프의 해당 노브를 조정하여 피펫의 용량 성 전류를 상쇄하십시오.
- 전극에 부정적인 압력의 빠르고, 짧고, 강력 한 관찰을 만들고, 전극 홀더에 연결된 튜브에서 입을 빨아서 감광체 막으로 "끊어"전체 셀 구성을 만듭니다. 또는 "Zap 버튼"을 사용하여 "0.1 ms"의 지속 시간으로 시작하는 짧은 직사각형 전기 펄스를 적용하거나 두 가지 방법의 조합을 적용하십시오.
참고 : 전체 세포 구성의 생성은 피펫 커패시턴스의 급격한 증가로 나타납니다 (일반적으로 야생형 R1-6 광 수용체의 경우 ~ 60pF, ~ 20pF의 커패시턴스 만 있으면 R7 광 수용체로부터의 기록, 위의 커패시턴스 ~ 90 pF는 두 개의 녹음을 나타냅니다.photoreceptors). - 패치 클램프 증폭기의 적절한 노브를 사용하여 포토 리셉터의 유지 전위를 필요한 전압 (일반적으로 -70mV)으로 설정합니다.
참고 : 밀봉을 얻은 후에 (단계 6.11) 전체 셀 구성이 이루어지기 전에이 단계를 수행 할 수 있습니다. - 커패시 티브 전류 및 직렬 저항 (25 MΩ보다 큰 측정 된 직렬 저항 값은 전극 피펫이 막혀 있음을 나타냄)을 오프셋하고 필요한 경우 ( 즉 , 큰 전류의 경우) 패치 클램프 증폭기의 적절한 노브를 사용하여 직렬 저항 보상을 적용하십시오 .
- 최대 어둠과 전기 절연을 얻으려면 패러데이 케이지의 검은 색 앞막을 닫으십시오.
- 소프트웨어를 사용하여 기록 과정을 시작하고 원하는 실험 절차에 따라 빛 자극 및 / 또는 약리학 물질을 투여하십시오.
7. Wh 동시셀 레코딩 및 칼슘 2 + 이미징
- 유전자 암호화 된 칼슘 2 + 표시기의 경우 GCaMP6f를 표현하는 D. melanogaster 파리를 사용하여 위에서 설명한 것처럼 각을 분리하십시오. 형광 측정을 위해 CCD 카메라 ( 재료 표 참조)를 사용하고 현미경에 적절한 여기 및 방출 필터와 이색 성 거울 ( 표 4 참조) 이 갖추어져 있는지 확인하십시오 4 .
- 외인성 칼슘 2+ 지시약 ( 그림 4 , 재료 표 참조)의 사용을 위해서는 위에서 언급 한 것처럼 개안을 분리하십시오. 또한 피펫 용액에 20-100 μM 칼슘 지시약이 있는지 확인하십시오.
- 이미징 소프트웨어 ( 재료 표 참조)를 사용하여 40 Hz의 속도로 이미지를 수집하십시오. 10 초 어두운 시간 동안 이미지 수집을 수행 한 다음 강렬한 2 초 조명 sti 수행Mulation.
- 이미징 소프트웨어를 사용하고 관심 영역 (ROI)을 정의하십시오. 투자 수익 (ROI)에서 형광 강도를 측정합니다. 어두운 형광 (F D )을 평균하고 빛 (F L ) 자극 (F L - F D ) 동안 형광 녹음에서 빼십시오. 빛 자극 (F L 0 )의 시작 부분에서 형광 강도에 따라 이러한 측정을 표준화하십시오.
결과
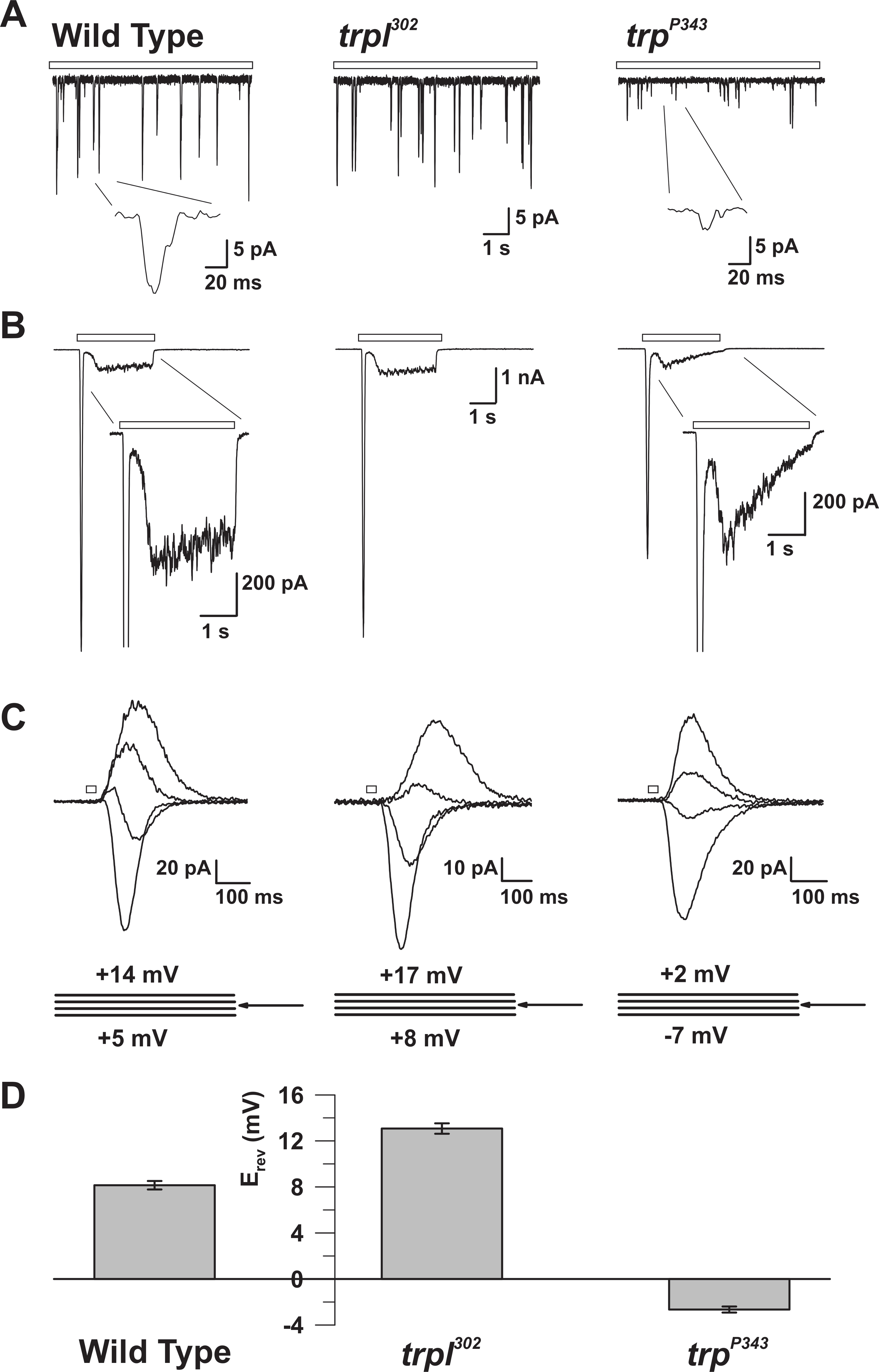
기술 된 방법은 정의 된 조건 하에서 빛에 대한 거시적 인 응답을 생성하기 위해 합계하는 자발적이고 광 유발 된 양자 범프를 생성하는 기본 단위 전류의 정확한 기록을 가능하게했다. 그것은 또한 중요한 신호 분자 ( 그림 3 과 5 ) 14 , 15 , 16 , 17 , 18에 결함이있는 야생형과 돌연변이 파리 사이의 비교를 허용했습니다. 또한, bi-ionic 조건 하에서 reversal potential을 측정하는 능력은 TRP와 TRP-like (TRPL) 채널의 기본적인 생물 물리적 특성을 밝혀냈다. 그것은 또한 Ca 2+ 를 변형시킨 TRP의 세공 영역에서 아미노산 치환 효과를 측정 할 수있게 해 주었다투자율 20 .
패치 클램프 전체 셀 기록에 의해 얻어진 빛 응답은 적어도 4 차수의 광 강도에 선형 적으로 의존합니다. 이것은 ERG 및 세포 내 기록 방법을 사용하여 해결할 수 없습니다. 따라서, 증가하는 강도의 간단한 섬광에 대한 일련의 반응 및 강도 반응 함수의 플롯은 광 세기가 증가함에 따라 플래시 응답의 엄격한 선형성을 나타내었다. 엄격한 선형성은 적어도 수백 pA까지 유지되지만, 이후에는 선형성 또는 고장을 일으키는 클램프 제어인지 여부는 논쟁의 여지가 있습니다 ( 그림 6 ). 이러한 결과는 빛에 대한 거시적 인 반응이 빛에 대한 단일 응답 ( 즉, 양자 범프)의 선형 합계임을 시사한다.
그것은 빛 자극 ind를 어둡게하는 전압 기록을 사용하여 잘 확립되어왔다.대부분의 무척추 동물에서 개별적인 전압 변동 ( 즉, 양자 범프)이 발생합니다. D. melanogaster 양자 범프는 범프 18 의 피크에서 ~ 15 TRP 채널과 ~ 2 TRPL 채널의 공동 개통으로 인해 발생합니다. 각각의 범프는 단일 광자의 흡수에 의해 생성되는 반면,보다 강렬한 빛에 대한 거시적 인 반응은 이들 기본 응답의 합계 14 , 21 이다. 범프는 자극 조건이 동일한 경우에도 대기 시간, 시간 경과 및 진폭이 크게 다릅니다. 범프 발생은 포아송 (Poisson) 통계에 의해 기술 된 확률 과정 (stochastic process)으로, 효과적으로 흡수 된 각각의 광자는 단지 하나의 범프만을 유도한다. 단일 - 광자 - 단일 범프 관계는 캐스케이드의 각 단계가 효율적인 "턴온"메커니즘뿐만 아니라 똑같이 효과적인 "턴 오프"메커니즘을 포함 할 것을 요구합니다. 기능적 이점은매우 민감한 광자 카운터는 시각적 시스템에 필요한 감도와 시간 해상도에 매우 적합한 빠른 과도 응답을 제공합니다. 능동적 인 photopigment ( 즉, metarhodopsin, M) 또는 그것의 타겟 인 G q α가 불 활성화되지 않고 빛이 꺼진 후에도 계속적으로 범프가 생성 될 때 효율적인 턴 오프 메커니즘에 대한 요구 사항이 드러난다. 3 ) 15 , 22 , 23 , 24 .
범프는 미세 융모에서 TRP / TRPL 채널의 협력 활동을 나타냅니다. 이와 같이, 채널 활성화의 임의의 가설은 협력 채널 활성화를 또한 설명해야한다. 최근 하디 (Hardie)와 동료들은 빛이 광 수용체의 빠른 수축을 불러 일으킨다는 것을 보여 주었는데, 이는 감광제e 채널 (TRP / TRPL)은 기계적으로 게이팅 될 수 있습니다 25 . 이 기계적 활성화는 PLC 매개 PIP 2 가수 분해에 의해 방출 된 관찰 된 양성자와 함께 TRP / TRPL 채널의 개방을 촉진하고 범프 생산의 협동 특성을 설명한다. 현재 D. melanogaster 광 수용체는 phosphoinositide 신호 전달 및 TRP 채널 을 생체 내 에서 연구 할 수있는 몇 가지 시스템 중 하나이므로 D. melanogaster 광전 변환을 만들고이 메커니즘을 연구하는 방법이 개발되어 매우 유용한 모델 시스템입니다.

그림 3 : inaC P209 와 inaD P215 돌연변이 체는 빛과 단일 양자 범프에 대한 거시적 반응의 느린 응답 종료를 나타냅니다. ( A ) 고립 된 개 모세 혈관 예비형광 루시퍼 옐로우 CH 염료 (여기 : 430 nm, 방출 : 540 nm)로 채워진 패치 피펫으로 배양 한 후 전체 세포를 녹음 하였다. 형광 염료는 하나의 광 수용체 세포 몸체를 확산 및 표지하고 광 수용체 세포 몸체는 길쭉한 축색 돌기에서 분리되지만 생존력을 유지한다는 점에 유의하십시오. 이 준비는 전체 세포 녹음 및 영상 실험을 동시에 수행하는 데 적합합니다. ( BD ) 상단 패널 : WT, inaC P209 및 inaD P215 돌연변이 플라이에서 연속 희미한 빛 (열린 막대)에 대한 전체 셀 전압 클램프 된 양자 범프 응답. 범프의 느린 종결은 WT 파리에 비해 inaC P209 및 inaD P215 돌연변이에서 관찰됩니다. 아래의 삽 입은 단일 범프의 확대 된 모양을 표시합니다. 하단 패널 : 정규화 된 전체 세포는 위의 야생형의 500ms 광 펄스 (1.5 x 10 5 광자 / 초)에 대한 거시적 인 응답을 기록했습니다. 돌연변이 파리. (EG) 상단 패널 : 야생형, arr2 3 및 ninaC P235 돌연변이 파리에서 단일 광자 반응을 유발하는 짧은 (1 ms) 희미한 빛에 대한 전체 셀 전압 클램프 된 양자 범프 응답. 단일 광자 흡수에 대한 반응으로 arr2 3 및 ninaC P235 돌연변이 파리에서 관찰 된 범프 트레인에 주목하십시오. 하단 패널 : 전체 셀 전압은 대응하는 돌연변이 체에서 500 ms 광 펄스 (1.5 x 10 4 광자 / s)에 대한 정규화 된 응답을 고정했습니다. arr2 3 에서 관찰 된 거시적 반응의 느린 종결에 주목하십시오 및 ninaC P235 돌연변이 체는 WT에 비하여 파리. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
{kind=link}
d / 55627 / 55627fig4.jpg "/>
그림 4 : 신호 유도 된 칼슘 2+ 주입 후 세포질 칼슘 2+ 역학은 칼 포틴에 의해 영향을받습니다. 야생형과 Cpn 1 % 의 광 수용체 이미지의 시계열은 빛 자극 동안 Ca 2+ 지시약의 형광을 나타냅니다. 원시 강도 영상은 잘못된 색상 코딩 (막대 = 10 μm, 화살촉은 피펫을 나타냄)을 사용하여 플롯됩니다. Weiss 외의 허가를 받아 증쇄 된 그림 . 4 . 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
{kind=link}

그림 5 : WT, trp 및 trpl 돌연변이 체의 전기 생리 학적 성질. ( A ) 전체 셀 전압 클램프 리코WT, trp 302 및 trp P343 null 돌연변이 플라이의 연속적인 희미한 빛 (열린 막대)에 대한 응답으로 양자 범프의 돌출부. trp P34 3 범프의 크게 감소 된 진폭이 관찰됩니다. 삽입 : wildtype과 trp P343 null 돌연변이 플라이의 확대 된 단일 양자 범프가 표시됩니다. ( B ) 야생형의 3 초 광 펄스 및 해당 돌연변이 체에 대한 반응으로 전체 세포 전압 클램프 기록. trp P343 돌연변이 체의 일시적인 정상 상태 응답이 관찰됩니다. 삽입 : WT 및 trp P343 돌연변이 체의 확대 된 빛 반응이 표시됩니다. ( C ) 역전 전위 (E rev )를 중심으로 측정 한 3 mV의 전압 단계에서 20 ms 광 펄스에 반응하여 유도 된 상기 플라이 변형의 중첩 된 광 유도 전류 군. ( D ) wildtype의 평균 E rev 와 다양한 mutan을 플로팅하는 히스토그램ts. 오류 막대는 SEM입니다. WT의 역전 전위 (E rev )는 TRP 만 나타내는 trpl 302 의 양의 전자 레인지와 TRPL 만 나타내는 trp P343 null 돌연변이의 전자 레인지 사이에 있습니다. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
{kind=link}

그림 6 : 플래시 응답은 빛의 강도가 증가함에 따라 엄밀히 선형입니다.
빛의 강도가 증가하는 간단한 섬광에 대한 현재의 일련의 반응과 짧은 빛의 섬광의 증가하는 강도에 대한 빛의 반응의 피크 진폭의 의존성에 대한 플롯. 이 관계는 플래시 응답과 증가하는 광 강도 간의 엄격한 선형성을 나타냅니다. 이 엄격한 선형성빛의 강도가 4 차수 이상에 걸쳐서 적어도 수백 pA를 유지하는 반면 선형성이든 아니면 이후에 고장 나게되는 클램프 제어인지는 논쟁의 여지가있다. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
{kind=link}
| pH | 7.15 (NaOH로 조절) |
| 시약 | 농도 (mM) |
| NaCl | 120 |
| KCl | 5 |
| MgCl2 | 4 |
| TES | 10 |
| 프롤린 | 25 세 |
| 알라닌 | 5 |
| -20 ° C에서 보관하십시오. | |
| 참고 :이 솔루션은 명목상 Ca 2+ 가 없지만 Ca 2+ 완충액이 첨가되지 않았으므로 약 5 - 10μM의 Ca 2+가 추가 됩니다. Extracellular solution (ES) = ES-0Ca 2+ 에 1.5 또는 1 M 재고 용액에서 CaCl 2 를 첨가하여 만든 1.5 mM CaCl 2의 ES-0Ca 2+ . | |
표 1 : Ca +2 없는 세포 외 용액 (ES). 화학적 설명 및 Ca +2가 없는 ES의 생산에 필요한 특정 양.
| 시약 | 양 |
| FBS | 15 mL |
| 자당 | 1.5 g |
| 1.5 mL 바이알에 150 μL 분량으로 나누어 -20 °에 보관하십시오 76, C. | |
| 연화 액 (TS) | 해부 동안 사용 된 용액과 일치하도록 원액 150 mL의 1 바이알에 1,350 mL의 ES 또는 ES-0Ca 2+ 를 채 웁니다. |
표 2 : 태아 소 혈청 (FBS) + 자당 스톡 용액. 태아 소 혈청 (FBS) + 자당 스톡 솔루션의 생산에 필요한 화학적 설명 및 특정 양.
| pH | 7.15 (KOH로 조정) |
| 시약 | 농도 (mM) |
| 글루 콘산 칼륨 (Kglu) | 140 |
| MgCl2 | 2 |
| TES | 10 |
| ATP 마그네슘 염 (MgATP) | 4 |
| GTP 나트륨 염 (Na2GTP) | 0.4 |
| β- 니코틴 아미드 아데닌 디 뉴클레오타이드 수화물 (NAD) | 1 |
| -20 ° C에서 보관하십시오. |
표 3 : 세포 내 용액 (IS1). 화학적 설명과 IS1 생산에 필요한 특정 양은 주로 강도 응답 및 양자 범프 측정에 사용됩니다.
| pH | 7.15 (CsOH로 조정) |
| 시약 | 농도 (mM) |
| CsCl | 120 |
| MgCl2 | 2 |
| TES | 10 |
| ATP 마그네슘 염 (MgATP) | 4 |
| GTP 나트륨 염 (Na2GTP) | 0.4 |
| β- 니코틴 아미드 아데닌 디 뉴클레오타이드 수화물 (NAD) | 1 |
| 테트라 에틸 - 염화 암모늄 (TAE) | 15 명 |
| -20 ° C에서 보관하십시오. |
표 4 : 세포 내 용액 (IS2). 광학적 유도 전류의 역전 전위 측정에 주로 사용되는 세포 내 용액 IS2를 생성하는 데 필요한 화학적 설명 및 특정 양.
토론
D. melanogaster 광 수용체에 대한 전체 세포 기록의 응용은 TRP 채널 27 , 28 , 29 및 INAD 30 , 31 , 32 스캐 폴드 단백질과 같은 새로운 신호 전달 단백질의 발견 및 기능적 해명을 가능하게했다. 이 기법을 처음 도입 한 이래로 이온 반응 메커니즘에 대한 장기적인 기본 문제를 해결할 수있었습니다. 이것은 멤브레인 전압과 세포 및 세포 내 이온 조성물 19 , 28 을 정확하게 제어 할 수있는 능력이 부여 되었기 때문에 발생했습니다.
D. melanogaster 에서의 패치 클램핑 기술의 주요 장애는 격리 된 개 구균 제거제의 취약성이다정액. 상세한 연구에 따르면 광전도 기계 장치의 무결성은 ATP의 지속적인 공급, 특히 빛에 노출되는 동안 ATP의 대량 소모로 이어지는 것이 중요합니다. 불행히도 패치 피펫으로 광 수용체 막에 도달하는 데 필요한 안료 ( 즉, glia) 세포의 기계적 스트라이핑은 ATP 생산에 필요한 주요 대사 물질을 제거합니다 33 . 기록 피펫으로의 외인성 ATP의 적용은 다량의 ATP에 대한 요건을 부분적으로 충족시킨다. ATP의 공급 부족은 TRP 채널의 자발적인 활성화와 빛 활성화 된 채널에서 광 전달 메커니즘의 해리로 연결되어 세포질 Ca 2+의 증가와 빛에 대한 정상적인 반응의 폐지를 초래한다. 이러한 일련의 사건은해부 절차에 의해 광 수용체가 아니라 오히려 ATP의 세포 고갈. 이러한 일련의 사건이 발생하지 않도록하고 정상적인 빛 반응을 유지하려면 광 수용체를 강렬한 빛에 노출 시켜서는 안되며 많은 양의 ATP를 섭취해야합니다. 또한, NAD는 기록 피펫에 포함되어야하며 아마도 미토콘드리아에서 ATP 생산을 촉진해야한다. 자발적 및 양자 범프의 측정을 위해서는 희미한 조명 만 사용되기 때문에 위의 어려움은 최소화됩니다. 실제적으로 안정된 전체 세포 기록은 ~ 20 ~ 25 분 동안 유지 될 수 있지만, 반응 속도론이이 기간 동안 느려지는 경향이있다. 해리 된 각화 증의 단일 제제는 최대 2 시간 동안 생존 할 수있다.
고립 된 개조 제제의 또 다른 단점은 TRP의 접근 불가능 성으로 해석되는 미생물의 접근 불가능 성이며,TRPL 채널을 녹음 피펫에 연결하여 단일 채널 녹음을 방지합니다. 그들이 개발 한 방법을 사용하여, Bacigalupo와 동료들은 횡문 면도 37 에서 단일 채널 활동을 직접 기록하는 데 성공했다. 그러나,이 채널 활성은 조직 배양 세포에서 이종적으로 발현 된 TRPL 채널의 것과는 다르며, 고립 된 개 구균 34 에서 얻은 샷 잡음 분석에서 파생 된 TRP 채널 활성과는 다릅니다. 아마도,이 방법을 사용할 때 해부 절차는 광 수용체 세포를 크게 손상 시켰습니다.
공개
저자는 공개 할 것이 없습니다.
감사의 말
이 연구의 실험적 부분은 미국 이스라엘 바이 사이언스 재단 (BM 및 IL), 이스라엘 과학 재단 (ISF), 독일 - 이스라엘 프로젝트 개발 (DIP) (BM으로), 그리고 생명 공학 및 생물 과학 연구위원회 (BBSRC 교부금 번호 : BB / M007006 / 1 및 BB / D007585 / 1)에서 RCH
자료
| Name | Company | Catalog Number | Comments |
| 10 mL syringe | |||
| 5 mL syringe | |||
| 1 mL syringe with elongated tip | |||
| Petri dish | 60 mm | ||
| Syringe filters | Millex | 22 µm PVDF filter | |
| Capillaries (for omatidia separation) | Glass, 1.2 x 0.68 mm (~7.5 cm each) | ||
| Polyethylene Tubing | Becton Dickinson | 1.57 x 1.14 mm (35 cm) | |
| 2 small beakers | 50 mL or less | ||
| 2 paraffin film sheets | Parafilm M | ~5 x 5 cm | |
| Bath chamber | home-made | ||
| Cover slips | 22 x 22 mm No. 0 | ||
| Paraplast Plus | Sigma | Paraffin – polyisobutylene mixture | To glue the coverslip onto the bottom of the bath chamber |
| Ground | Warner Instruments | 64-1288 | Hybrid Assembly Ag-AgCl Wire Assembly |
| Headless micro dissection needle | Entomology, 12 mm | ||
| Micro dissecting needle holder | |||
| Vise | |||
| 2 fine tweezers + 1 rough tweezers | Dumont #5, Biology | 0.05 x 0.02 mm, length 110 mm, Inox | |
| Stereoscopic zoom Microscope | Nikon | SMZ-2B | |
| Cold light source | Schott | KL1500 LCD | |
| Filter (Color) for cold light source | Schott | RG620 | |
| Delicate wipers | Kimtech | Kimwipes | |
| Electrode holder | Warner Instruments | QSW-T10P | Q series Holders compatible with Axon amplifiers, straight body style |
| Silver Wire | Warner Instruments | 0.25 mm diameter, needs to be chloridized | |
| Micromanipulator | Sutter Instruments | MP 85 | Huxley-Wall Style Micromanipulator |
| Faraday cage | home made | Electromagnetic noise shielding and black front curtain | |
| Anti-vibration Table | Newport | VW-3036-OPT-01 | |
| Osilloscope | GW | GOS-622G | |
| Perfusion system | Warner Instruments | VC-8P | Pinch valve control system |
| Perfusion valve controller | Scientific instruments | BPS-8 | |
| Suction system | |||
| Amplifier | Molecular Device | Axopatch-1D | |
| Head-stage | Molecular Device | CV - 4 | Gain: x 1/100 |
| A/D converter | Molecular Device | Digidata 1440A | |
| Clampex | Molecular Device | 10 | software |
| pCLAMP | Molecular Device | 10 | software |
| Light source (Xenon Arc lamp) | Sutter Instruments | Lambda LS | |
| Light detector | home made | phototransistor | |
| Filter wheel and shutter controller | Sutter Instruments | Lambda 10-2 with a Uniblitz shutter | |
| Filters (Natural density filter) | Chroma | 6,5,4,3,2,1,0.5,0.3 | |
| Filter (Color) | Schott | OG590, Edge filter | |
| Xenon Flash Lamp system | Dr. Rapp OptoElecftronic | JML-C2 | |
| Light guide | Quartz | ||
| Pulse generator | AMPI | Master 8 | |
| Microscope | Olympus | IX71, Inverted | |
| Red illumination filter (Microscope) | RG630 / RG645 ø45mm | ||
| Microscope objective | Olympus | X60/0.9 UplanFL N air or X60/1.25 UplanFI oil | |
| CCD Camera | Andor | iXon DU885K | |
| NIS Element | Nikon | AR | software |
| Ca+2 indicator | Invitrogen | Calcium green 5N | |
| Excitation & emission filters and dichroic mirror | Chroma | 19002 - AT - GFP/FITC Longpass set | |
| Vertical pipette puller | Narishige | Model PP83 | Use either vertical or horizontal puller, as preferred. |
| Horizontal pipette puller | Sutter Instrument | Model P-1000 Flaming/Brown Micropipette Puller | |
| Filament | Sutter Instrument | 3 mm trough or square box | |
| Capillaries | Harvard Apparatus | borosilicate glass capillaries | 1 x 0.58 mm |
참고문헌
- Hardie, R. C. Whole-cell recordings of the light induced current in dissociated Drosophila photoreceptors: evidence for feedback by calcium permeating the light-sensitive channels. Proc. R. Soc. Lond. B. 245, 203-210 (1991).
- Ranganathan, R., Harris, G. L., Stevens, C. F., Zuker, C. S. A Drosophila mutant defective in extracellular calcium- dependent photoreceptor deactivation and rapid desensitization. Nature. 354, 230-232 (1991).
- Martin, J. H., Benzer, S., Rudnicka, M., Miller, C. A. Calphotin: a Drosophila photoreceptor cell calcium-binding protein. Proc. Natl. Acad. Sci. U.S.A. 90 (4), 1531-1535 (1993).
- Weiss, S., et al. Compartmentalization and Ca2+ buffering are essential for prevention of light-induced retinal degeneration. J Neurosci. 32 (42), 14696-14708 (2012).
- Wang, T., et al. Light activation, adaptation, and cell survival functions of the Na + /Ca 2+ exchanger CalX. Neuron. 45 (3), 367-378 (2005).
- Weckstrom, M., Hardie, R. C., Laughlin, S. B. Voltage-activated potassium channels in blowfly photoreceptors and their role in light adaptation. J. Physiol. Lond. 440, (1991).
- Frolov, R. V., Immonen, E. V., Weckström, M. Performance of blue- and green-sensitive photoreceptors of the cricket Gryllus bimaculatus. J Comp Physiol A Neuroethol Sens Neural Behav Physiol. 200 (3), 209-219 (2014).
- Nasi, E. Whole-cell clamp of dissociated photoreceptors from the eye of Lima scabra. J Gen Physiol. 97 (1), 35-54 (1991).
- Nasi, E., Gomez, M. P. Light-activated ion channels in solitary photoreceptors of the scallop Pecten irradians. J. Gen. Physiol. 99, 747-769 (1992).
- Hardie, R. C., Peretz, A., Pollock, J. A., Minke, B. Ca 2+ limits the development of the light response in Drosophila photoreceptors. Proc. R. Soc. Lond. B. Biol. Sci. 252, 223-229 (1993).
- Kohn, E., et al. Functional Cooperation between the IP3 Receptor and Phospholipase C Secures the High Sensitivity to Light of Drosophila Photoreceptors In Vivo. J Neurosci. 35 (6), 2530-2546 (2015).
- Hevers, W., Hardie, R. C. Serotonin modulates the voltage dependence of delayed rectifier and Shaker potassium channels in Drosophila photoreceptors. Neuron. 14 (4), 845-856 (1995).
- Chen, T. W., et al. Ultrasensitive fluorescent proteins for imaging neuronal activity. Nature. 499 (7458), 295-300 (2013).
- Henderson, S. R., Reuss, H., Hardie, R. C. Single photon responses in Drosophila photoreceptors and their regulation by Ca 2. J. Physiol. Lond. 524 (Pt 1), 179-194 (2000).
- Scott, K., Zuker, C. S. Assembly of the Drosophila phototransduction cascade into a signalling complex shapes elementary responses. Nature. 395 (6704), 805-808 (1998).
- Elia, N., Frechter, S., Gedi, Y., Minke, B., Selinger, Z. Excess of G betae over G qalphae in vivo prevents dark, spontaneous activity of Drosophila photoreceptors. J. Cell Biol. 171 (3), 517-526 (2005).
- Katz, B., Minke, B. Phospholipase C-Mediated Suppression of Dark Noise Enables Single-Photon Detection in Drosophila Photoreceptors. J. Neurosci. 32 (8), 2722-2733 (2012).
- Hardie, R. C., et al. Molecular basis of amplification in Drosophila phototransduction. Roles for G protein, phospholipase C, and diacylglycerol kinase. Neuron. 36 (4), 689-701 (2002).
- Reuss, H., Mojet, M. H., Chyb, S., Hardie, R. C. In vivo analysis of the Drosophila light-sensitive channels, TRP and TRPL. Neuron. 19, 1249-1259 (1997).
- Liu, C. H., et al. In vivo identification and manipulation of the Ca 2+ selectivity filter in the Drosophila transient receptor potential channel. J. Neurosci. 27 (3), 604-615 (2007).
- Ahmad, S. T., Natochin, M., Barren, B., Artemyev, N. O., O'Tousa, J. E. Heterologous expression of bovine rhodopsin in Drosophila photoreceptor cells. Invest Ophthalmol Vis Sci. 47 (9), 3722-3728 (2006).
- Scott, K., Sun, Y., Beckingham, K., Zuker, C. S. Calmodulin regulation of Drosophila light-activated channels and receptor function mediates termination of the light response in vivo. Cell. 91 (3), 375-383 (1997).
- Liu, C. H., et al. Ca 2+ -dependent metarhodopsin inactivation mediated by calmodulin and NINAC myosin III. Neuron. 59 (5), 778-789 (2008).
- Cook, B., et al. Phospholipase C and termination of G-protein-mediated signalling in vivo. Nat. Cell Biol. 2 (5), 296-301 (2000).
- Hardie, R. C., Franze, K. Photomechanical responses in Drosophila photoreceptors. Science. 338 (6104), 260-263 (2012).
- Huang, J., et al. Activation of TRP channels by protons and phosphoinositide depletion in Drosophila photoreceptors. Curr. Biol. 20 (3), 189-197 (2010).
- Minke, B., Wu, C. F., Pak, W. L. Isolation of light-induce response of the central retinular cells from the electroretinogram of Drosophila. J. Comp. Physiol. 98, 345-355 (1975).
- Hardie, R. C., Minke, B. The trp gene is essential for a light-activated Ca2+ channel in Drosophila photoreceptors. Neuron. 8, 643-651 (1992).
- Niemeyer, B. A., Suzuki, E., Scott, K., Jalink, K., Zuker, C. S. The Drosophila light-activated conductance is composed of the two channels TRP and TRPL. Cell. 85 (5), 651-659 (1996).
- Huber, A., et al. The transient receptor potential protein (Trp), a putative store- operated Ca 2+ channel essential for phosphoinositide-mediated photoreception, forms a signaling complex with NorpA, InaC and InaD. EMBO J. 15 (24), 7036-7045 (1996).
- Shieh, B. H., Niemeyer, B. A novel protein encoded by the InaD gene regulates recovery of visual transduction in Drosophila. Neuron. 14 (1), 201-210 (1995).
- Tsunoda, S., et al. A multivalent PDZ-domain protein assembles signalling complexes in a G-protein-coupled cascade. Nature. 388 (6639), 243-249 (1997).
- Tsacopoulos, M., Veuthey, A. L., Saravelos, S. G., Perrottet, P., Tsoupras, G. Glial cells transform glucose to alanine, which fuels the neurons in the honeybee retina. J. Neurosci. 14 (3 Pt 1), 1339-1351 (1994).
- Hardie, R. C., Minke, B. Spontaneous activation of light-sensitive channels in Drosophila photoreceptors. J. Gen. Physiol. 103, 389-407 (1994).
- Agam, K., et al. Metabolic stress reversibly activates the Drosophila light-sensitive channels TRP and TRPL in vivo. J Neurosci. 20 (15), 5748-5755 (2000).
- Agam, K., Frechter, S., Minke, B. Activation of the Drosophila TRP and TRPL channels requires both Ca2+ and protein dephosphorylation. Cell Calcium. 35 (2), 87-105 (2004).
- Delgado, R., Muñoz, Y., Peña-Cortés, H., Giavalisco, P., Bacigalupo, J. Diacylglycerol activates the light-dependent channel TRP in the photosensitive microvilli of Drosophila melanogaster photoreceptors. J Neurosci. 34 (19), 6679-6686 (2014).
- Parnas, M., Katz, B., Minke, B. Open channel block by Ca2+ underlies the voltage dependence of Drosophila TRPL channel. J. Gen. Physiol. 129 (1), 17-28 (2007).
재인쇄 및 허가
JoVE'article의 텍스트 или 그림을 다시 사용하시려면 허가 살펴보기
허가 살펴보기더 많은 기사 탐색
This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. 판권 소유