
Method Article
Microarrays de DNA e RNA de alta densidade-síntese fotolitográfica, hibridação e preparação de grandes bibliotecas de ácidos nucleicos
Neste Artigo
Resumo
Neste artigo, apresentamos e discutimos novos desenvolvimentos na síntese e aplicações de Microarrays de ácidos nucleicos fabricados in situ. Especificamente, mostramos como os protocolos de síntese de DNA podem ser estendidos ao RNA e como os microarrays podem ser usados para criar bibliotecas de ácidos nucleicos recuperáveis.
Resumo
Photolithography é uma técnica poderosa para a síntese de oligonucleotídeos do ADN em corrediças de vidro, porque combina a eficiência de reações do acoplamento do monomers com a precisão e a densidade da luz UV refletida dos espelhos micrometer-feitos medida. A fotolitografia produz Microarrays que podem acomodar de centenas de milhares até vários milhões de seqüências de DNA diferentes, 100-NT ou mais, em apenas algumas horas. Com este espaço muito grande da seqüência, os microarrays são plataformas ideais para explorar os mecanismos do ácido nucleico · interações do ligand, que são particular relevantes no caso do RNA. Nós relatado recentemente na preparação de um jogo novo de phosphoramidites do RNA compatíveis com o photolithography in situ e que foram usados subseqüentemente para crescer oligonucleotides do RNA, homopolímeros assim como seqüências da misturado-base. Aqui, ilustramos detalhadamente o processo de fabricação de Microarray de RNA, desde o projeto experimental, até a configuração instrumental, síntese de matriz, desproteção e ensaio de hibridação final usando uma seqüência de modelo 25mer contendo todas as quatro bases como um exemplo. Paralelamente, vamos além de experimentos baseados em hibridação e exploramos a fotolitografia de Microarray como uma porta de entrada barata para bibliotecas de ácidos nucleicos complexos. Para isso, os microarrays de DNA de alta densidade são fabricados em um monómero sensível à base que permite que o DNA seja convenientemente clivada e recuperado após síntese e desproteção. O protocolo de fabricação é otimizado de modo a limitar o número de erros sintéticos e, para esse efeito, uma camada de solução de β-caroteno é introduzida para absorver fótons de UV que, de outra forma, podem refletir de volta para os substratos de síntese. Nós descrevemos em uma maneira passo a passo o processo completo da preparação da biblioteca, do projeto à clivagem e à quantificação.
Introdução
O uso prático de Microarrays de DNA tem sido tradicionalmente no estudo das variações nos níveis de expressão gênica entre duas populações celulares, utilizando vertentes complementares e fluorescência como método de detecção1. Ocasionalmente, os microarrays de DNA se aventuram em eventos vinculativos com ligantes de ácido não nucleico, como proteínas, com uma estratégia de permutação sistemática de sequências que oferece uma visão abrangente da paisagem vinculativa2,3, 4,5. Esta aproximação transforma eficazmente Microarrays das superfícies simples da hibridação em plataformas com a cobertura larga da seqüência, que seria um recurso para o estudo do mundo mais rico e mais complexo da estrutura e da função do RNA. Apoiado pela reação extremamente eficiente do acoplamento do monomers6, as matrizes sintetizadas in situ do ADN podem agora igualmente ser consideradas como uma fonte barata do ADN7, que está tornando-se particular relevante Considerando a demanda cada vez maior para material de ácido nucleico para o conjunto de genes8,9,nanoestruturas baseadas em DNA10, armazenamento de informações ou sequenciamento11,12. Da mesma forma, as tecnologias de sequenciamento provavelmente se beneficiarão do desenvolvimento de métodos que produzem misturas muito complexas de oligonucleotídeos de RNA13. Neste contexto, os protocolos da fabricação da disposição que permitem que os oligonucleotídeos sejam sintetizados in situ e na alta densidade são coloc idealmente para encontrar as necessidades do campo ràpida de expansão da biotecnologia ácida nucleico. Entretanto, com um campo tão diverso quanto a biotecnologia, a finalidade de cada aplicação pode exigir que o ADN no microarray esteja produzido no elevado-throughput ou com uma quantidade muito baixa de erros sintéticos14,15, ou ambos, exigindo um olhar mais atento para os protocolos de síntese de Microarrays de DNA que, historicamente, foram otimizados principalmente para ensaios de hibridação. Entrementes, a síntese in situ de Microarrays do RNA foi encontrada para ser um esforço desafiante, com a maioria da dificuldade associada com o grupo de protecção para a função 2 '-Oh, geralmente um metade do trimetilsilílicos na síntese padrão da contínuo-fase que é removida com reagentes à base de flúor, produtos químicos incompatíveis com superfícies de vidro ou silício. Estas edições e os desafios na síntese do microarray do ADN e do RNA têm sido ultimamente o assunto de um grande corpo do trabalho, no detalhe com a aproximação da fotolitografia16.
Photolithography usa a luz UV para desbloquear oligonucleotídeos antes do acoplamento e exige máscaras para construir um teste padrão da exposição UV, organizando desse modo espacialmente e controlando o crescimento de oligonucleotídeos. As máscaras físicas foram substituídas por microespelhos controlados por computador cuja inclinação reflete seletivamente a luz UV sobre o substrato microarray17,18,19. Como uma fonte de UV, usamos 365 nm luz de uma fonte de LED de alta potência20. As configurações fotolitográficas atuais são equipadas com as matrizes do micromirror que contêm 1024 × 768 espelhos, correspondendo a mais de 780.000 pontos individualmente enderezáveis ("características") em uma área pequena de apenas 1,4 cm2, ou 1080p com matrizes de 1920 × 1080, ou > 2 milhões de espelhos. Cada um dos espelhos no dispositivo, portanto, tem controle direto sobre a seqüência cultivada no recurso correspondente. Com exceção da luz UV, a fotolitografia funciona como uma técnica de síntese de fase sólida e adota a química de fosfoidite baseada em ciclo. Somente exige uma estratégia inteiramente diferente da proteção para que a síntese do RNA suceda. Nós desenvolvemos uma série nova de fosfatos luz-sensíveis do RNA que carregam grupos protegendo hydrazine-labile21. Estes monómeros permitem que o RNA seja desprotegido condições suaves que não afetam a integridade da superfície. Uma primeira etapa da desproteção usa a trietilamina para remover os grupos de protecção do fosfodiéster do ácido, quando o hidrazina for usado em um segundo, etapa separada para remover aqueles no 2 '-Oh e nas funções exocíclicas da amina. Ao fazer isso, o RNA oligonucleotídeos ~ 30-NT de comprimento e de qualquer seqüência pode agora ser sintetizado in situ em Microarrays22,23. Paralelamente, também recentemente começamos a abordar as questões de produtividade, qualidade e velocidade na fotolitografia de DNA e RNA. Foram mensuradas as eficiências de acoplamento > 99% para todos os amiditos de DNA e RNA (Figura 1) e investigaram cada etapa individual no ciclo de alongamento do oligonucleotídeo, desde o tempo de oxidação, à escolha do ativador e à exposição UV ideal24 , 25. We trouxeram em grupos de protecção 5 ' Light-sensitive novos que podem ser removidos nos segundos somente, transformando a síntese de centenas de milhares de 100mers em um processo de poucas horas-longo26. Nós também dobramos a produção de matriz de fabricação, expondo dois substratos simultaneamente27. Finalmente, nós introduzimos um monomers do descolamento que contem um grupo base-sensível do succinyl como uma maneira conveniente de Cleave, coletam e analisam oligonucleotides do ADN e do RNA, que é central à preparação28da biblioteca.
Apesar do aspecto relativamente mundano da síntese de fase sólida de DNA e RNA, especialmente para os químicos de ácidos nucleicos, a fotolitografia de Microarray continua sendo uma atualização não trivial que exige uma configuração complexa, controle cuidadoso e supervisão do processo, e instruções separadas para manuseio pós-sintético, dependendo da natureza do oligonucleotídeo e do tipo de aplicação. Neste artigo, desejamos fornecer uma apresentação detalhada de todo o procedimento passo a passo da síntese in situ de Microarrays de DNA e RNA por fotolitografia, desde o delineamento experimental até a análise dos dados, com ênfase na elaboração de instrumentos e Consumíveis. Em seguida, descrevemos os métodos de desproteção pós-sintéticos que correspondem à finalidade pretendida da fabricação de Microarray (ou seja, a hibridação ou a recuperação de bibliotecas de ácidos nucleicos).
Protocolo
1. projeto do microarray
- Escreva as sequências a serem sintetizadas em um editor de texto, 5 ' → 3 ', uma linha por sequência. Para o controle de qualidade 25mer, use a sequência "GTCATCATCATGAACCACCCTGGTC". Use letras A, C, G e T para nucleotídeos de DNA e números 5, 6, 7 e 8 para nucleotídeos de RNA.
- No caso da preparação da biblioteca, adicione um número extra (isto é, 9) no final de 3 ' de cada sequência. Isso corresponderá ao acoplamento do monómero sensível à base.
- Cada sequência deve ser seguida por uma vírgula. Atribua um nome a cada sequência após a vírgula e verifique se cada faixa segue o formato [Sequence, #sequence_name] (sem colchetes). Guarde a lista de sequências como um ficheiro. txt.
- Inicie o MatLab e carregue o programa Chipdesign. m. Execute o programa.
- Na janela do painel de propriedades, carregue o arquivo de layout do microarray de um chip . Carregue o arquivo de layout Millichip se o objetivo é sintetizar toda a matriz em quatro locais idênticos.
- Na janela do painel de propriedades , em especificação de chip, selecione selecionar contêiner e área de síntese. Em padrão, selecione o padrão que produz a quantidade correta de recursos enderetiveis, contando o número de sequências × número de repetições. Para o controle de qualidade 25mer, selecione o padrão 25:36.
- Em selecionar recipiente , selecione As características de recursos são geralmente sintetizadas nos cantos da área de síntese e são usadas para extrair dados de hibridação. Com o sistema de gravação selecionado, escreva uma sequência (5 ' → 3 ') que será sintetizada em características de recursos e que podem atuar como um controle positivo. Para o experimento 25mer, use a mesma sequência de DNA.
- Em sequência, carregar o arquivo de texto que contém todas as seqüências escritas. Certifique-se de que Randomize está selecionado. Dê um título para o experimento no título do projeto e, em seqüência de vinculador, escreva ttttt (correspondente a um vinculador T5 ).
- Pressione gerar, em seguida, encontrar os arquivos de design de matriz e máscaras (Figura 2) em MaskGen_delta_rc1/designs pasta. Certifique-se de que contém um script de exibição, uma sequência de fluxo e um arquivo de design.
- Para a preparação da biblioteca, abra o script de exibição. Adicione uma linha extra na parte inferior do script de exibição que copia exatamente a primeira linha do script (por exemplo, Exibir First_Mask. bmp 150). Isto removerá o grupo photoprotegendo terminal na extremidade 5 ' de todos os oligonucleotides.
- Inicie o programa de trabalho automático do criador de trabalhos, carregue um modelo clicando em carregar modeloe clique em Exibir script para carregar o arquivo de script de exibição que foi gerado pelo MATLAB. Pressione Generate. Esta etapa irá criar uma série de instruções, chamado de um jobfile, que irá controlar a comunicação do computador com os microespelhos e o sintetizador de DNA.
2. slide preparação e funcionalização
- Perfure uma corrediça em duas posições que correspondem à posição da tubulação da entrada e da tomada na pilha da síntese. Use um bocado do diamante de 0,9 milímetros em um router do CNC para perfurar precisamente e confiantemente. Enxágüe as lâminas perfuradas com água ultrapura e organize-as em um rack de slides.
- Limpe as superfícies sonicando as lâminas em um banho de água contendo 5% de um limpador de finalidade especial à base de amônia por 30 min a 35 ° c. Enxágüe o slide com H2o duplo destilado e transfira-os em um rack limpo e seco.
- Organize slides de microscópio perfurados e não perfurados em um rack de slides. Em um grande cilindro graduado, prepare a solução de funcionalização misturando 475 mL de etanol (EtOH) com 25 mL de ddH2O, 10 g de reagente silanizante (N-(3-trietoxisilylpropyl)-4-hidroxibutiramida) e 1 ml de ácido acético. Mexa bem até que homogêneo, em seguida, transferir em um recipiente adequado, fechado.
- Coloque a cremalheira carregada no recipiente, feche a tampa e deixe a rocha do recipiente delicadamente em um abanador orbital para 4 h na temperatura ambiente.
- Após 4 h, descartar a solução de funcionalização e substituir com 500 mL de solução de lavagem, consistindo de 475 mL de EtOH, 25 mL de ddH2O e 1 ml de ácido acético. Agitar lentamente durante 20 min à temperatura ambiente, depois descartar e substituir com 500 mL de solução de lavagem fresca.
- Após 20 minutos adicionais à temperatura ambiente, elimine a solução, seque as lâminas com um fluxo de argônio e Cure-as num forno a vácuo pré-aquecido a 120 ° c. Depois de 2 h, desligue o forno e a bomba de vácuo, mas deixe as lâminas pressão reduzida durante a noite. Em seguida, traga o forno de volta à pressão atmosférica e guarde as lâminas em um dessecador até o uso posterior.
3. preparação de reagentes de síntese e reagentes
- Coloque os pós de fosfoidite (Figura 1) da temperatura de armazenamento (-25 ou-45 ° c) à temperatura ambiente em um dessecador.
- Quando os fosfatos atingiram a temperatura ambiente, dissolvem o pó com um volume de acetonitrila ultraleve (< 30 ppm H2O) para atingir a concentração de 30 mm para o DNA padrão e fosfatos de RNA, e 50 mm para o monômero DT sensível à base ( preparação da biblioteca). Adicione um pequeno saco de peneiras moleculares para prender qualquer vestígio de humidade.
- Prepare uma solução de 1% (p/p) de imidazol em DMSO dissolvendo 11 g de imidazol em 1 L de DMSO seco. Agitar bem até dissolver completamente. Anexe a solução à porta auxiliar do sintetizador de DNA. Este será o solvente de exposição necessário para a remoção completa do grupo de 5 ′-photoprotegendo.
- Para a síntese de bibliotecas, prepare uma solução de 1% (p/v) β-caroteno em diclorometano dissolvendo 100 mg de β-caroteno em 10 mL de diclorometano. Agitar bem em uma garrafa de vidro âmbar, em seguida, embrulhe em folha de alumínio.
4. preparação e monitorização da síntese de Microarray.
- Recorde de temperatura e umidade na sala de fabricação de Microarray e certifique-se que o sintetizador de DNA está pressão suficiente de hélio.
- Ligue o LED UV e a ventoinha de arrefecimento. Fixe um medidor de intensidade UV no plano focal da luz UV de entrada e ligue-o (Figura 3a).
- No computador, inicie o software de controlador de arquivos de jobfile/Micromirror/síntese (denominado WiCell). Ative e, em seguida, inicialize o dispositivo Micromirror e carregue um arquivo de máscara todo-branco clicando com o botão direito do mouse no DMDe selecionando carregar imagem.
- Botão direito do mouse sobre o ícone UVS e selecione obturador UV aberto. Leia o valor de potência (em mW/cm2) no medidor de intensidade e conte 60 s. Depois de 60 s, leia o valor de energia novamente e anote os valores inicial e final. Feche o obturador selecionando o obturador UV perto e desligue o medidor de intensidade. Calcule o valor médio da intensidade de UV em mW/cm2.
- Calcule o tempo de exposição necessário para atingir uma energia radiante de 6 J/cm2 para 5 '-NPPOC photodeprotection (DNA e RNA microarrays) e 3 J/cm2 para 5 '-bznppoc photodeprotection (bibliotecas de DNA), simplesmente
 seguindo o relacionamento.
seguindo o relacionamento. - No software de estação de trabalho de sintetizador de DNA, crie um arquivo de sequência no editor de sequências copiando e colando o conteúdo da sequência de fluxo gerada pelo MatLab. Adicione duas etapas de lavagem extra na extremidade 3 ' (letra de ciclo padrão: "s") para lavar a superfície dos substratos antes de passar para o primeiro acoplamento. Guarde e exporte o ficheiro de sequência.
- No editor de protocolo do software Workstation, crie um protocolo que contenha um ciclo dedicado denominado após cada uma das letras e números presentes no arquivo de sequência (por exemplo, se um arquivo de sequência contiver apenas letras A, C, G e T, o arquivo de protocolo deve contêm quatro ciclos, denominados A, C, G e T).
- Ajuste o tempo de acoplamento para fosfatos de DNA (ciclos A, C, G e T) a 15 s, a 120 s para Fosfito de rU (ciclo 8) e a 300 s para fosfoidites rA, rC e rG (ciclos 5, 6 e 7). Para a preparação da biblioteca, defina o acoplamento do monômero dT clivável sensível à base para 2 × 120 s (tabela 1 e tabela 2).
- Certifique-se de que o tempo de espera entre dois eventos de comunicação do evento 2 out em cada ciclo corresponda ao tempo de exposição UV calculado para atingir a energia radiante necessária. Quanto ao arquivo de sequência, salve e exporte o arquivo de protocolo.
- No software WiCell, carregar o trabalho, seqüência e arquivos de protocolo em suas respectivas subjanelas, em seguida, clique em Enviar para enviar os arquivos de seqüência e protocolo para o SINTETIZADOR de DNA.
- No arquivo de sequência, conte o número de acoplamentos para cada phosphoramidite.
- Para medir o volume necessário (em μL) de solução de fosfoidite necessária para executar cada síntese, multiplicar o número de acoplamentos por 60. Adicionar 250 μL a cada volume para segurança e escorva da linha no sintetizador de ADN. Utilize um volume de base de 250 μL para fosfatos que exijam apenas um acoplamento único.
- Transfira rapidamente a solução para os frascos para injetáveis no sintetizador de ADN correspondente à letra/número da porta nos ciclos de protocolo.
- Prime as portas com as soluções de fosforamidite com 10 pulsos primos cada, a fim de preencher as linhas com reagente. Prime a linha de lavagem de acetonitrila antes de fixar a célula de reacção.
- Para montar a célula de síntese, coloque uma junta grossa (250 μm) perfluoroelastómero (FFKM) primeiro no bloco de quartzo da célula. Coloc uma corrediça perfurada, funcionalizados do microscópio sobre a primeira gaxeta, e verifique que os furos nas corrediças conectam com a tubulação da entrada e da tomada da pilha da síntese.
- Coloc uma segunda, gaxeta fina (de 50 μm) politetrafluoretileno (PTFE) sobre a corrediça perfurada, cercando os dois furos. Finalmente, coloque um segundo, funcionalizados, mas não perfurado slide no topo da segunda junta (Figura 3B). Coloque um quadro metálico de 4 parafusos na parte superior da célula de substrato duplo montado e aperte o parafuso para a mesma força de aperto (0,45 nm) usando uma chave de fenda de torque.
- Fixe a tubagem de entrada e saída ao sintetizador de ADN. Prima a linha de lavagem acetonitrila e verifique o fluxo adequado de acetonitrila (ACN) através dos substratos. Desmontar e remontar a célula se qualquer vazamento de ACN pode ser observado nesta fase. Medir o volume de ACN na linha de resíduos depois de passar por 7 ciclos de escorva da ACN. Este volume deve ser de 2 mL.
- Anexar a célula de síntese no plano focal de entrada de luz UV. No caso da preparação da biblioteca, prenda uma linha extra de entrada e saída à parte de trás da célula e encha a câmara traseira com a solução de β-caroteno (2 mL de solução é suficiente). Certifique-se de que não há vazamento (Figura 3C).
- Inicie a síntese clicando primeiro em executar no software WiCell. No primeiro comando WAIT no arquivo de trabalho, pressione Start no SINTETIZADOR de DNA.
- Durante a síntese, verifique regularmente se a exibição de arquivos de máscara coincide com a exposição UV e a abertura do obturador.
- Após a síntese de Microarrays regulares, desconecte a célula do sintetizador, desmontar a célula e usar uma caneta de diamante para gravar o número de síntese para as lâminas de vidro. Etch o número na face não sintetizada de cada slide. Transfira os slides em tubos de centrifugação de 50 mL e armazene-os em uma área dessecada até o uso posterior.
- Após a síntese de Microarrays da biblioteca, escorra primeiramente a solução do β-caroteno fora da câmara a seguir lave-a fluindo 2 × 5 mL de CH2CL2, a seguir escorra. Prossiga com a desmontagem de acordo com a etapa 4,21. A matriz sintetizada pode ser visível a olho nu (Figura 4).
5. desproteção do microarray do ADN
- Encha um frasco de vidro da mancha com 20 ml de EtOH e de 20 ml do etilenodiamina (EDA). Coloque os microarrays somente de DNA verticalmente no frasco, feche a tampa e deixe os slides para desproteger por 2 h à temperatura ambiente.
PRECAUÇÃO: a EDA é um líquido agudamente tóxico, corrosivo e inflamável. Trabalhe com luvas em uma capa de fumaça bem ventilada. - Após 2 h, recupere os slides usando pinças e enxague-os cuidadosamente com H2O duplo destilado.
- Seque os slides em uma centrífuga microarray por alguns segundos, em seguida, armazenar em um dessecador.
6. desproteção do microarray do RNA
- Num tubo de centrifugação de 50 mL, prepare uma solução seca de 2:3 trietilamina/ACN (20 mL e 30 mL de cada, respetivamente). Transfira uma corrediça do microarray do RNA no tubo de centrifugação, feche a tampa então enrole com filme plástico da selagem. Agitar suavemente o tubo de centrifugação num agitador orbital durante 1h e 30 min à temperatura ambiente.
- Depois de 1h e 30 min, retire o slide, lave com 2 × 20 mL de ACN seco e seque em uma centrífuga de Microarray por alguns segundos.
Cuidado: a trietilamina é um líquido agudamente tóxico, corrosivo e inflamável. Acetonitrila é tóxico e inflamável. Trabalhe com luvas em uma capa de fumaça bem ventilada.
Nota: primeira etapa de desproteção concluída. - Prepare uma solução de hidrato de hidrazina de 0,5 M em 3:2 ácido acético/piridina. Em primeiro lugar, misture 20 mL de ácido acético e 30 mL de piridina em um cilindro graduado. Aguarde a solução arrefecer antes de adicionar 1,21 mL de hidrazina hidratada. Transfira cerca de 40 mL da solução resultante para um tubo de centrifugação de 50 mL.
Cuidado: hidrazina hidrato é um líquido agudamente tóxico e corrosivo. A piridina é altamente inflamável e agudamente tóxica. O ácido acético é inflamável e corrosivo. Trabalhe com luvas em uma capa de fumaça bem ventilada. - Após a primeira etapa de desproteção, transfira a corrediça de RNA para a solução hidrazina hidratada, feche a tampa e enrole com filme plástico de vedação. Agitar suavemente o tubo num agitador orbital durante 2 h à temperatura ambiente. Depois de 2 h, retire a lâmina, lave com 2 × 20 mL de ACN seco e seque-a em uma centrífuga de Microarranjo durante alguns segundos.
Nota: segunda etapa de desproteção concluída. - Se a micromatriz de RNA também contiver nucleotídeos de DNA, prossiga com uma terceira etapa de desproteção. Num tubo de centrifugação de 50 mL, misture 20 ml de EDA com 20 mL de EtOH. Adicione o microarray DNA/RNA à solução 1:1 EDA/EtOH e deixe à temperatura ambiente por 5 min.
- Após 5 min, retire o slide, lave com 2 × 20 mL de água estéril, em seguida, seque em uma centrífuga Microarray e armazenar em um dessecador.
7. hibridação com uma vertente complementar fluorescently-etiquetada
- Descongelar a albumina sérica de bovinos acetilados (10 mg/mL) e a vertente complementar com o rótulo Cy3 (100 nM) e aquecer até à temperatura ambiente.
- Em um tubo de microcentrífuga estéril de 1,5 mL, misture 150 μL de 2x tampão MES (200 mM 2-(N-morpholino) ácido etanólico; 1,8 M NaCl; 40 mm EDTA; 0, 2% Tween-20) com 26,7 ΜL de DNA rotulado com Cy3, 13,4 ΜL de BSA acetilada e 110 μL de H2O. mistura estéril e Vórtice. Dobre o volume se ambos os slides devem ser usados para a hibridação.
- Pré-aqueça um forno da hibridação a uma temperatura abaixo do Tm do duplex mas elevado bastante para assegurar a boa discriminação entre seqüências do cheio-fósforo e do não-fósforo. Para o controle de qualidade 25mer, ajuste a temperatura a 42 ° c (Tm de 59 ° c).
- Coloc com cuidado uma câmara autoadesiva da hibridação do μL 300 sobre a área da síntese em cada corrediça e Pipet na solução da hibridação preparada acima. Cubra os furos da câmara com pontos adesivos e enrole a corrediça inteira na folha de alumínio.
- Coloque a corrediça do microarray no forno da hibridação, cubra-a e deixe-a girar delicadamente na temperatura selecionada da hibridação para 2 h.
- Após 2 h, retire o slide, remova a folha de alumínio e cuidadosamente rasgar a câmara de hibridização. Transfira as lâminas para um tubo de centrifugação contendo 30 mL de tampão de lavagem não rigoroso (NSWB; 0,9 M NaCl, 0, 6 M de fosfato, 6 mM de EDTA, 0, 1% Tween20, pH 7,4) e agitar vigorosamente durante 2 min à temperatura ambiente.
- Transfira o slide para um tubo de centrifugação contendo 30 mL de tampão de lavagem rigoroso (SWB; 100 mM MES, 0,1 M NaCl, 0, 1% Tween20) e agite vigorosamente durante 1 min.
- Finalmente, transfira o slide para um tubo de centrífuga contendo 30 mL de tampão de lavagem final (FWB; 0,1 x citrato salino de sódio) e agitar por alguns segundos. Seque o slide em uma centrífuga microarray.
- Coloque o microarray seco, área de síntese voltada para baixo, no porta-lâmina do scanner de Microarray. Para duplexes rotulados com Cy3, faça uma varredura com uma resolução de 5 μm com um comprimento de onda de excitação de 532 nm, um filtro de 575 nm e um fotomultiplicador de 350. Salve a varredura de alta resolução como um arquivo de imagem. tif (Figura 5a).
8. extração e análise de dados
- Antes da extração de dados, gire a varredura de matriz salva em um editor de imagem para situar a cadeia mais longa de recursos de edição no canto superior esquerdo da digitalização. Salve a imagem girada.
- Inicie o Nimblescane, em seguida, prima File | Abra e carregue a verificação de matriz. Em seguida, clicando em procurar na subseção de arquivo de design, carregar o. arquivo de projeto FDN que foi gerado automaticamente durante o experimento da experiência de Microarray. Em seguida, clique em abrir.
- No modo de exibição, clique em contraste/brilho automático . Clique no ícone alinhar manualmente acima da digitalização. Coloque quatro marcadores em forma de quadrado nos quatro cantos da digitalização e clique no ícone verde agora novamente. Extraia os dados da hibridação estalando na análise, relata então o relatório da sonda .
- Abra o. arquivo de relatório de sonda em um editor de planilha. Mantenha as colunas B e I e descarte o restante antes de prosseguir com o cálculo dos valores médios e do desvio padrão dos dados extraídos (Figura 5B).
9. Biblioteca deprotection, clivagem e recuperação
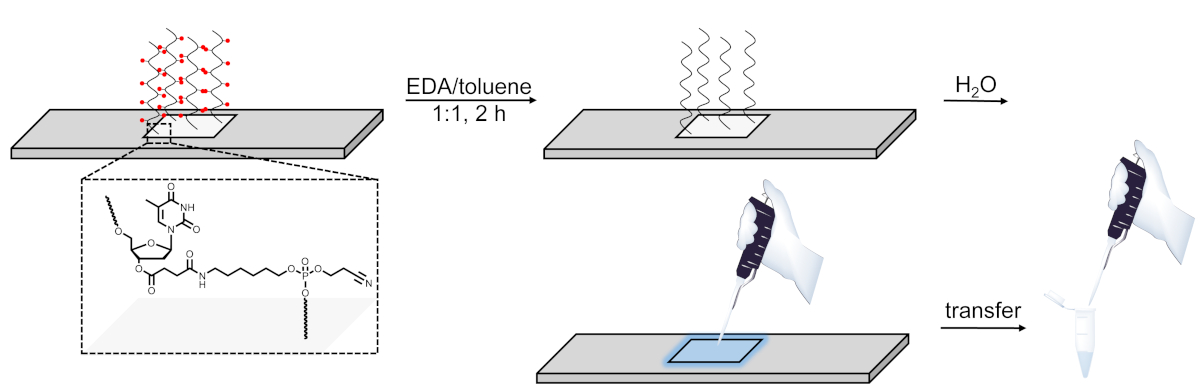
- Para desproteger e Cleave bibliotecas de DNA, prepare uma solução de 1:1 Dry EDA/tolueno em um tubo de centrífuga de 50 ml. Mergulhe o slide na solução de clivagem, feche o slide e enrole com filme plástico de vedação, em seguida, Gire suavemente em um agitador orbital para 2 h à temperatura ambiente.
- Depois de 2 h, retire a lâmina e lave com 2 × 20 mL de ACN seca escrupulosamente. Retire o slide e deixe secar ao ar.
- Com uma pipeta, aplique 100 μL de H2o estéril sobre a área agora discernable da síntese. Pipet a solução para cima e para baixo algumas vezes antes de transferi-lo em um tubo de microcentrífuga de 1,5 mL. Repita o processo e combine o eluído do microarray no mesmo tubo (Figura 6).
- Evate o 2 × 100 μl do eluído da microplaqueta à secura a seguir redissolver-se em 10 μL de H2o. da nuclease-livre meça a absorvância em um espectrofotômetro UV-VIS.
- Desalt a biblioteca do ADN em uma ponta da pipeta de 10 μL equipada com a resina C18. Primeiramente, transforme o eluído da microplaqueta em uma solução tamponada do acetato do de de 0,1 M (Teaa).
- Molhar a resina aspirando 3 x 10 μL de H2o/ACN 1:1, e equilibrar a resina por lavagem com 3 x 10 μL de tampão teaa de 0,1 M. Vincular o DNA por pipetagem do chip eluído 10 vezes para cima e para baixo através da resina. Lave a resina com 3 x 10 μL de tampão TEAA de 0,1 M, 3 x 10 μL de H2O.
- Elute o ADN demolhado da ponta lavando a resina com 10 μL de H2O/ACN 1:1. Seque a solução dessalinada para baixo e redissolva em 10 μL de H2o estéril. Meça a absorvância num espectrofotômetro UV-VIS (Figura 7). Evate a biblioteca para secura e armazene a-20 ° c até o uso mais adicional.
Resultados
Hibridação do microarray do ADN e do RNA
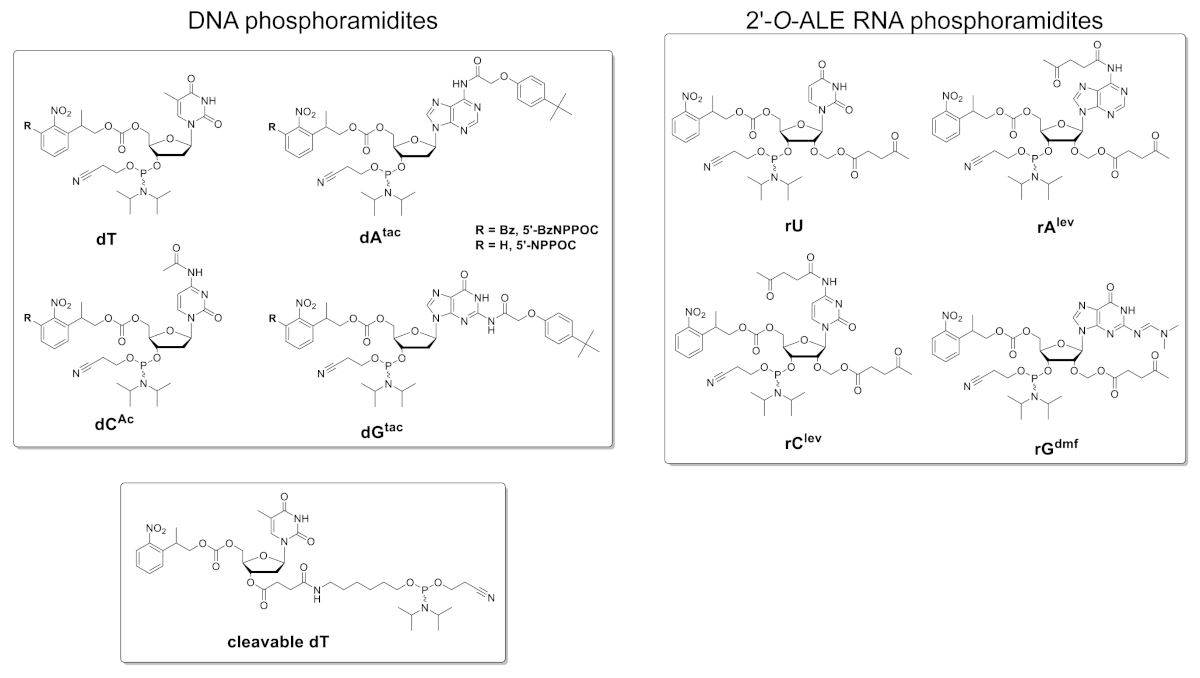
A Figura 5 mostra os resultados de um ensaio da hibridação executado em um microarray que contem as versões do ADN e do RNA de uma seqüência 25mer (5 '-GTCATCATCATGAACCACCCTGGTC-3 ' no formulário do ADN). A varredura na Figura 5a aparece em um formato de greenscale correspondente ao espectro de excitação/emissão de fluorescência Cy3, com intensidade de fluorescência registrada em unidades arbitrárias entre 0 e 65536. O design da matriz seguiu o layout de recurso 25:36 descrito na seção de protocolo. A digitalização é mostrada após a orientação adequada da matriz, com o canto superior esquerdo preenchido com a cadeia mais longa de recursos de características. Aqui, as características da propriedade contêm a versão do ADN do 25mer e devem, no princípio, sempre dar um sinal positivo da fluorescência a fim executar o alinhamento da varredura e a extração dos dados. O microarray hibridizado deve aparecer uniformemente brilhante, com as bordas da área de síntese sendo no entanto geralmente mais brilhante do que o centro (até 50% mais brilhante). A grande quantidade de repetições de sequências, distribuídas aleatoriamente em toda a área, reduz o impacto de artefatos espaciais. Aqui, cada seqüência (DNA e RNA) foi sintetizada em 2.000 locais aleatórios. Há tipicamente baixo ruído da fluorescência (fundo), < 50 u.a., que conduz a uma relação do sinal/ruído na ordem de 200:1 a 800:1 em ensaios da hibridação. Após a extração dos dados, intensidades de fluorescência são médias e plotadas ± DP.
Há variabilidade significativa nos valores de fluorescência absoluta entre experimentos. Aqui, mostramos os resultados de três sínteses independentes utilizando os mesmos parâmetros de fabricação e o mesmo manuseio pós-sintético. O DNA 25mer, quando hibridizado a sua vertente complementar Cy3-etiquetada do ADN, produzirá os sinais da fluorescência que variam em qualquer lugar de 20.000 a 30.000, muito raramente acima ou abaixo. O RNA 25mer, quando hibridizado ao mesmo complemento Cy3-etiquetado do ADN, dará intensidades da fluorescência nas características correspondentes que variam de 15.000 a 20.000. Entretanto, a intensidade da fluorescência dos duplex do RNA/ADN cairá ocasionalmente abaixo de 8.000, quando os duplex correspondentes do ADN/ADN ainda fluorescência dentro da escala 20000-30000. Em tais casos, os resultados para o RNA podem ser considerados como secundário-optimal. Uma síntese ou falha de hibridação, seja para DNA ou RNA, será imediatamente perceptível durante a digitalização a partir da óbvia falta de fluorescência. Há umas oportunidades múltiplas para que a síntese do RNA falhe ou seja parcialmente suceder e serão esboçadas na divisória da discussão.
Desproteção, clivagem e recuperação da biblioteca
Dependendo da complexidade e da densidade da biblioteca, a forma e o contorno da matriz sintetizada podem ser vistos sem ampliação, mas iluminação adequada (Figura 4), com o DNA ainda em forma protegida. Após a desproteção com EDA/tolueno, e antes da adição de água, a fim de coletar a biblioteca clivada, a área de síntese contendo os oligonucleotídeos agora desprotegidos pode se destacar como uma zona hidrofílica, quando o resto da lâmina de vidro aparecerá coberto com uma camada hidrofóbica turva. A observação direta da área de síntese depende da área total utilizada para sintetizar oligonucleotídeos: maior uso da área de síntese corresponderá a maior chance de distinção clara entre regiões hidrofílicas e hidrofóbicas na superfície. Inversamente, bibliotecas sintetizadas usando menos espelhos e com recursos menores podem não ser imediatamente observáveis.
Da mesma forma, a quantidade de DNA recuperado após a dessalinagem é diretamente proporcional à área total usada para síntese. Se todas as características são usadas para a síntese do oligonucleotide, a clivagem e o procedimento da recuperação devem render entre 25 e 30 pmol do ADN. Um uso de 10% da área de síntese, portanto, vai pagar apenas cerca de 3 pmol de DNA.

Figura 1 . As estruturas químicas do DNA e do RNA phosphoramidites usaram-se na síntese do oligonucleotide pela fotolitografia do Microarray. Os grupos de protecção fotossensível padrão de nitrophenylpropyloxycarbonyl (nppoc) no 5 '-Oh são usados na síntese regular do microarray do ADN e do RNA para finalidades da hibridação. Para a síntese de bibliotecas complexas do ADN, mais fotolábeis Benzyl-nppoc (bznppoc) é preferido no 5 '-Oh, porque bznppoc é removido duas vezes tão rapidamente quanto nppoc, que reduz significativamente o tempo total da síntese do Microarray. Os oligonucleotídeos do ADN para bibliotecas igualmente exigem o acoplamento de um monômero ligante do descolamento na extremidade 3 '. Este monómero, que carrega uma função de éster succinílico, será clivada durante a desproteção, permitindo que o DNA seja coletado do microchip. Por favor clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 2 . Exemplo de uma máscara como um arquivo de imagem enviado para o dispositivo Micromirror durante a exposição UV. Os pixels brancos correspondem aos espelhos que serão inclinados na posição "on", refletindo a luz UV sobre a célula de síntese. Pixels pretos correspondem a "off" espelhos, onde a luz UV será refletida longe da célula. Os pixéis brancos permitirão conseqüentemente para o acoplamento do monomers entrante seguinte nos oligonucleotídeos encontrados nas características correspondentes nos substratos de vidro. Os oligonucleotides sintetizados nas características cujos espelhos correspondentes são, neste arquivo da máscara, os pixéis pretos permanecerão inertes entretanto durante o evento de acoplamento seguinte. Por favor clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 3 . Fotografias do microarray fotolitografia óptica e configuração de síntese. (A) circuito óptico para exposição A UV. A luz UV do UV-LED é homogeneizada pela primeira vez através de um tubo de luz retangular-cross-section, em seguida, reflete sobre os microespelhos. Os microespelhos que foram inclinados em uma posição "OFF" refletirão a luz UV longe da célula de síntese, mas os microespelhos na posição "ON" refletirão a luz na célula de síntese, situada no plano focal, pela primeira passagem através de um relé 1:1 Offner sistema de imagem. (B) a célula de síntese, uma vez montada, consiste em uma corrediça perfurada colocada primeiramente no bloco de quartzo da pilha, separada por uma gaxeta grossa de PTFE (não mostrada). Um segundo, não-perfurou a corrediça é posicionada então sobre a corrediça perfurada, separada por uma gaxeta fina de PTFE. Uma armação metálica (não mostrada) mantém a montagem unida. (C). para a preparação da biblioteca, uma vez que a célula de síntese é anexada no plano focal de entrada de luz UV, a câmara localizada entre o bloco de quartzo e o slide perfurado é preenchido com uma solução de 1% de β-caroteno em ch2CL2. Para isso, um tubo de entrada e saída adicional é anexado ao bloco de quartzo e a solução laranja flui do mais à direita para a posição mais à esquerda. O fluxo de reagentes e solventes para a síntese é mostrado em setas brancas. Por favor clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 4 . A área de síntese é geralmente visível a olho nu. Aqui, uma biblioteca de DNA pode ser vista na superfície de vidro logo após a síntese, com o DNA ainda em forma protegida. Por favor clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 5 . Os ensaios da hibridação às seqüências do ADN 25mer e do RNA sintetizados in situ em Microarrays. (A) varredura da fluorescência do ADN e do microarray hibridizados inteiros do RNA. Os oligonucleotídeos do ADN e do RNA 25mer são hibridizados a suas costas complementares Cy3-etiquetadas. A matriz foi digitalizada com um laser em um comprimento de onda de excitação de 532 nm, com resolução de 5 μm. (B) intensidades de fluorescência (unidades arbitrárias) do DNA: DNA e RNA: duplex de DNA em três experimentos distintos. Os dados verdes claros para oligonucleotídeos sintetizados in situ do RNA podem ser considerados suboptimal, quando comparados à intensidade da fluorescência das seqüências correspondentes do ADN. As barras de erro são SD. por favor clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 6 . Representação esquemática do procedimento de desproteção, clivagem e recuperação para bibliotecas de DNA sintetizadas em Microarrays. As sequências de DNA são cultivadas em um nucleosídeo DT clivável sensível à base (mostrado na área ampliada). Após a síntese, a desproteção dos oligonucleotídeos do ADN (os grupos de protecção da base são representados como esferas vermelhas) em EDA/Toluene sae do material deprotected ligado electrostaticamente à superfície e pode então pipetado para fora aplicando uma pequena quantidade de água na área sintetizada. Por favor clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 7 . Espectro representativo da absorvência (220-350 nanômetro) de uma biblioteca cleaved, demolhado do ADN que contem 4.000 seqüências diferentes, 100-NT no comprimento. Um total de 940 ng do ADN foi isolado de uma única síntese da disposição, correspondendo a 30 pmol do total do ADN, ou 15 pmol por o substrato de vidro. Por favor clique aqui para ver uma versão maior desta figura.
{kind=link}

Tabela 1. Protocolo representativo do ciclo para o acoplamento/oxidação/photodeprotection de 5 '-bznppoc-da, supondo que o monomers correspondente estêve carregado no porto "a". O tempo de acoplamento (em segundos) é mostrado na linha "monômero de casal". O tempo de fotodeproteção UV, aqui correspondente a uma energia radiante de 3 J/cm2 (fotoquímica bznppoc), é calculado como o tempo decorrido entre os dois sinais de comunicação "evento 2 out".

Tabela 2. Protocolo representativo do ciclo para o acoplamento/oxidação/photodeprotection de 5 '-nppoc-ra, supondo que o monomers correspondente do RNA estêve carregado no porto "a". O tempo de acoplamento (em segundos) é mostrado na linha "monômero de casal". O tempo de fotodeproteção UV, aqui correspondente a uma energia radiante de 6 J/cm2 (fotoquímica nppoc), é calculado como o tempo decorrido entre os dois sinais de comunicação "evento 2 out".
Discussão
A síntese de DNA e RNA de fase sólida é o pão e a manteiga de todos os laboratórios de química de ácidos nucleicos, e embora a adição do componente de fotolitografia seja reconhecidamente uma operação complexa, a fabricação de Microarray mediada pela luz UV também é um processo muito confiável . É, além, o único método disponível para a síntese in situ do RNA em Microarrays. Ainda assim, como em qualquer procedimento experimental em vários estágios, há um amplo espaço para o erro humano.
Talvez o passo mais crítico é o acoplamento de um phosphoramidite, como ele precisa ser uma reação química constantemente de alto rendimento, a fim de pagar oligonucleotídeos com poucos erros sintéticos. Em nosso protocolo da síntese do microarray, o acoplamento do monomers é ainda mais crucial à qualidade total da síntese desde que o processo da fabricação contorna tampando e impede a purificação do oligonucleotide. As eficiências Stepwise do acoplamento acima de 99% foram calculadas para todos os phosphoramidites fotossensível do ADN e do RNA, mesmo para tempos de acoplamento muito curtos (15 s)24 mas uns rendimentos mais baixos do acoplamento podem ocasionalmente ocorrer, particular no caso de DG amidites. A estabilidade dos fosfatos solubilizados à temperatura ambiente tem sido investigada antes e mostrou-se depender da natureza da nucleobase, com fosfatos de guanosina propensas a degradação extensiva em apenas uma questão de dias29, a 30. Mas, quando armazenada a-25 ° c, a dG fosamidites dissolvidos na ACN como solução de 30 mM foi encontrada estável durante várias semanas. No entanto, a instabilidade relativa das soluções de fosfoidite da dG à temperatura ambiente significa que não devem ser mantidas ligadas ao sintetizador de ADN durante vários dias.
Para Fosfito de RNA, o rendimento do acoplamento é muito dependente da qualidade de fosfoidite (que pode ser avaliado por espectroscopia de RMN de 31P) e tempo de acoplamento. Os tempos de acoplamento de 5 minutos para rA, rG, rC e 2 min para rU parecem necessários. Certamente, nós encontramos que encurtando o tempo da condensação a 2 minutos para todos os phosphoramidites do RNA conduzidos aos sinais significativamente mais baixos da hibridação.
O sintetizador de DNA em si, bem como os reagentes e solventes, certamente precisa ser tão limpo quanto possível, a fim de alcançar o maior rendimento de síntese de oligonucleotídeo. No entanto, materiais insolúveis, sais ou partículas, podem se acumular ao longo do tempo nas linhas e tubulações do sistema de parto, levando a uma diminuição gradual no consumo de reagentes e reactantes. Quando uma limpeza geral do sintetizador não resolver um volume de saída baixo, um aumento no número de pulsos pode ser uma solução alternativa. Particularmente útil no caso de baixo consumo de fosfoidite, a linha no protocolo de acoplamento correspondente ao bombeamento de uma mistura de fosfoidite e ativador (terceira linha da subseção de acoplamento na tabela 1 e tabela 2) pode ser modificada, de 6 a 9 pulsos sem qualquer efeito negativo apreciável na qualidade da síntese. Além disso, o número de pulsos de ativador necessário para trazer a mistura de amidite/ativador para o substrato de síntese (atualmente 6, quarta linha na subseção de acoplamento, ver tabela 1 e tabela 2) depende do próprio sintetizador de DNA, bem como em comprimento da tubagem na célula de síntese. Este número pode ser ajustado após a substituição do monomers com uma solução colorida e contando o número de pulsos necessários para empurrar a mistura colorida para o substrato de vidro para acoplamento.
O método descrito nisto permite a síntese do ADN e do RNA de prosseguir simultaneamente, no mesmo microarray. Os híbridos do ADN e do RNA podem igualmente ser preparados sem nenhuma mudança aos protocolos da fabricação da disposição, e contanto que o protocolo da desproteção da três-etapa for seguido. Entretanto, deve-se notar que os microarrays RNA-somente exigem uma desproteção em duas etapas: um decyanoethylation primeiramente com o et3N seguido pelo hidroxila e pela desproteção baixa com hydrazine. Os nucleobases do ADN foram encontrados para ser desprotegidos incompletamente aquelas circunstâncias, e precisam a etapa adicional em EDA a fim efetuar a remoção completa de grupos do phenoxyacetyl (PAC). Este tratamento extra com EDA é mais curto (5 min) do que para a desproteção padrão de Microarrays de DNA31, mas é suficiente para conduzi-lo para a conclusão após os tratamentos de Trietilamina e hidrazina. Além, um tempo de reação curto com EDA limita a exposição de um oligonucleotide inteiramente-deprotected do RNA às condições básicas.
Uma vantagem da síntese in situ da disposição do RNA sobre métodos alternativos como a mancha ou a transcrição32do ADN,33,34 são a habilidade de armazenar o microchip sintetizado do RNA na forma protegida até o uso, assim evitando o risco de degradação potencial do RNA. Os procedimentos pós-sintéticos para RNA, por outro lado, implicam que os consumíveis e reagentes são mantidos estéreis e que o manuseio é realizado condições livres de RNase. Da nota, nós encontramos que a adição de inibidor de RNase à mistura da hibridação não rende uns sinais mais fortes da hibridação para as características do RNA.
A síntese de bibliotecas de DNA em um monómero sensível à base é mais complexa do que a síntese de algumas seqüências de controle em uma superfície, e como tal é certamente mais propensos a erros de design. No entanto, assumindo que o design da sequência (ou seja, a natureza e o número de sequências) está correto, transformando esta lista em uma coleção de máscaras de exposição e uma série ordenada de ciclos de acoplamento continua a ser um processo simples. No entanto, variações importantes da síntese de Microarray padrão existem e são críticas para uma fabricação bem-sucedida de uma matriz de biblioteca de alta densidade.
Primeiro, um monômero DT sensível à base é acoplado como o primeiro monomers após a síntese do vinculador. O rendimento de acoplamento deste monómero (Figura 1) foi encontrado relativamente baixo, cerca de 85%28, razão pela qual são envidados esforços para melhorar a sua taxa de incorporação, quer aumentando a sua concentração na ACN de 30 mm para 50 mm, ou repetindo o etapa de acoplamento: duas reações de acoplamento consecutivas usando monómeros frescos, ou dois ciclos de acoplamento separados mas consecutivos.
A segunda alteração é a adição de uma solução de β-caroteno na câmara traseira da célula de síntese, que absorve convenientemente a luz de 365 nm. Esta é uma modificação importante da instalação do photolithography porque impede que a luz UV reflita para trás no substrato da disposição. Na verdade, depois de atravessar o meio intersticial entre os substratos, saídas de luz UV de entrada através do slide perfurado e atinge o bloco de quartzo da célula. As equações de Fresnel prevêem que ~ 4% da luz UV do incidente perpendicularmente refletirá de cada uma das três interfaces de ar-vidro a jusante (lado da saída do substrato 2ND e ambos os lados do bloco de quartzo) e de volta para o substrato de síntese, levando a exposição não intencional de oligonucleotides fotoprotegidos. A difração e a dispersão também contribuem para a fotoproteção "off-Target" e, portanto, para a inserção de nucleotídeo, que afeta diretamente a taxa de erro de síntese, mas essas contribuições são muito menores que as reflexões e podem ser abordadas principalmente por reduzindo a densidade de síntese (deixando lacunas entre as feições). Nós encontramos que o nível de solução do β-caroteno na câmara mais baixa da pilha tende a diminuir ligeiramente somente durante os primeiros minutos da síntese da disposição, e conseqüentemente precisa de ser monitorado e readjusted.
Finalmente, a terceira mudança é a solução de desproteção, substituindo EtOH por tolueno, que mantém a biblioteca de DNA clivada ligada à superfície, presumivelmente através de interações eletrostáticas. A aplicação de uma pequena quantidade de água na área de síntese após a lavagem da ACN permite que a biblioteca seja convenientemente recolhida. O processo é entretanto somente bem sucedido se o índice de água no EDA e no tolueno for mínimo, tornando o ácido nucleico inteiramente insolúvel no cocktail do desproteção. Alternativamente, as bibliotecas do ADN podem ser clivadas fora da microplaqueta usando a amônia9,10,14,35, a seguir desprotegidas mais por aquecer o ADN-contendo a solução aquosa da amônia a 55 ° c durante a noite. A recuperação de bibliotecas do ADN que usam a amônia não é entretanto compatível com RNA. Os oligonucleotídeos do RNA em uma carcaça base-cleavable podem ser eluída da superfície usando o mesmo procedimento de EDA/Toluene descrito acima, mas somente no penúltimo estágio após a estratégia de desproteção do dois-passo de et3N e de hidrazina28.
As estratégias alternativas para recuperar associações do oligonucleotide dos Microarrays sem a necessidade para um tratamento básico específico existem, são no princípio compatíveis com photolithography e confiam no uso das enzimas. Por exemplo, um único nucleotide do deoxyuracil pode ser o alvo do uracil-ADN glicolisase (UDG) e extirpado do descanso da seqüência do ADN, ou uma única unidade do RNA pode ser reconhecida pelo tipo 2 enzimas de RNase H e pela ligação 5 do fosfodiéster ao RNA clivada , liberando o 5 ' DNA parte23.
Nós temos agora um método poderoso, de confiança e high-density para a síntese do ADN, do RNA, e dos Microarrays híbridos do ADN/RNA. Estes podem não somente serir como plataformas para a hibridação ou os ensaios de ligação36, mas igualmente representam uma maneira rápida e barata de produzir bibliotecas complexas do ácido nucleico. Para o armazenamento de dados digital baseado em DNA, a fotolitografia de Microarray pode se tornar uma solução potencial para o gargalo de "escrita" (ou seja, para a codificação de informações por síntese). O sucesso na codificação digital no ADN e no conjunto do gene de de novo depende da fidelidade da seqüência que, a nível da síntese, traduz na taxa de erro. Os erros sintéticos e óticos em nossos protocolos atuais da fabricação da disposição serão discutidos e relatados em em outra parte. Paralelamente, os esforços estão agora em andamento para aumentar ainda mais a escala de fabricação e a taxa de transferência.
Divulgações
Os autores certificam que não têm nenhuma afiliação com nenhuma organização para fins lucrativos.
Agradecimentos
Este trabalho foi apoiado pelo Austrian Science Fund (FWF concede P23797, P27275 e P30596) e pela Swiss National Science Foundation (Grant #PBBEP2_146174).
Materiais
| Name | Company | Catalog Number | Comments |
| Slide functionalization | |||
| Acetic acid >99.8% | Sigma | 33209 | For RNA deprotection |
| CNC router | Stepcraft | 300 CK | |
| Ethanol absolute | VWR | 1.07017.2511 | For deprotection and functionalization |
| N-(3-triethoxysilylpropyl)-4-hydroxybutyramide | Gelest | SIT8189.5 | Silanizing reagent |
| Nexterion Glass D microscope slides | Schott | 1095568 | |
| Polymax 1040 | Heidolph | Orbital shaker | |
| Proclean 507 Ultrasonic water bath | Ulsonix | To clean slides after drilling | |
| Tickopur RW 77 Special Purpose Cleaner | Sigma | Z860086 | To clean slides after drilling |
| Microarray synthesis | |||
| 0.25 M dicyanoimidazole in ACN | Biosolve | 0004712402BS | Activator |
| 0.7 XGA DMD | Texas Instruments | Digital Micromirror Device | |
| 20 mM I2 in pyridine/H2O/THF | Sigma | L860020-06 | Oxidizer |
| 250 μm thick Chemraz 584 perfluoroelastomer | FFKM | Lower teflon gasket | |
| 2'-O-ALE RNA phosphoramidites | ChemGenes | ||
| 365 nm high-power UV-LED | Nichia | NVSU333A | |
| 5'BzNPPOC DNA phosphoramidites | Orgentis | ||
| 5'NPPOC DNA phosphoramidites | FlexGen | ||
| Acetonitrile | Biosolve | 0001205402BS | For DNA synthesis |
| Amidite Diluent for DNA synthesis | Sigma | L010010 | For dissolving phosphoramidites |
| Cleavable dT | ChemGenes | Base-sensitive monomer for library preparation | |
| DMSO | Biosolve | 0004474701BS | As exposure solvent |
| DNA and RNA microarray deprotection | |||
| Ethylenediamine >99.5% | Sigma | 3550 | For deprotection |
| Expedite 8909 | PerSeptive Biosystems | DNA synthesizer | |
| Hydrazine hydrate 50-60% hydrazine | Sigma | 225819 | For RNA deprotection |
| Imidazole | Sigma | 56750 | |
| Industrial Strength lower-density PTFE tape | Gasoila | Thin, upper teflon gasket | |
| Pyridine >99% | Sigma | P57506 | For RNA deprotection |
| Triethylamine >99% | Sigma | T0886 | For RNA deprotection |
| β-carotene | Sigma | C9750 | For library preparation |
| Hybridization and scanning | |||
| 20x Sodium Saline Citrate | Roth | 1054.1 | |
| 5'Cy3-labelled complementary strand | Eurogentec | For duplex hybridization | |
| Biopur Safe-Lock microcentrifuge tube | Eppendorf | ||
| BSA (10 mg/mL) | Promega | R3961 | |
| EDTA molecular biology grade | Promega | H5031 | |
| GenePix 4100A | Molecular Devices | Microarray scanner | |
| Hybridization oven | Boekel Scientific | 230500 | |
| MES monohydrate | Sigma | 69889 | |
| MES sodium | Sigma | M3058 | |
| NaCl >99.5% | Sigma | 71376 | |
| SecureSeal SA200 hybridization chamber | Grace BioLabs | 623503 | |
| Spectrafuge mini | Labnet | C1301 | Microarray centrifuge |
| Tween-20 molecular biology grade | Sigma | P9416 | |
| Data extraction | |||
| Excel | Microsoft | For data extraction | |
| MatLab | MathWorks | Microarray design | |
| NimbleScan 2.1 | Roche NimbleGen | ||
| Desalting and quantification | |||
| NanoDrop One Spectrophotometer | Thermo Scientific | ||
| Toluene | Merck | ||
| ZipTip C18 | Millipore | ZTC18s008 | Desalting pipet tips |
Referências
- Bumgarner, R. Current protocols in molecular biology. 101, 22(2013).
- Berger, M. F., Bulyk, M. L. Universal protein-binding microarrays for the comprehensive characterization of the DNA-binding specificities of transcription factors. Nature Protocols. 4 (3), 393-411 (2009).
- Tietjen, J. R., Donato, L. J., Bhimisaria, D., Ansari, A. Z. Sequence-specificity and energy landscapes of DNA-binding molecules. Methods in enzymology. 497, 3-30 (2011).
- Katilius, E., Flores, C., Woodbury, N. W. Exploring the sequence space of a DNA aptamer using microarrays. Nucleic Acids Research. 35 (22), 7626-7635 (2007).
- Franssen-van Hal, N. L. W., et al. Optimized Light-Directed Synthesis of Aptamer Microarrays. Analytical Chemistry. 85 (12), 5950-5957 (2013).
- Matteucci, M. D., Caruthers, M. H. Nucleotide Chemistry .4. Synthesis of Deoxyoligonucleotides on a Polymer Support. Journal of the American Chemical Society. 103 (11), 3185-3191 (1981).
- Eroshenko, N., Kosuri, S., Marblestone, A. H., Conway, N., Church, G. M. Gene Assembly from Chip-Synthesized Oligonucleotides. Current Protocols in Chemical Biology. 2012, (2012).
- Kosuri, S., et al. Scalable gene synthesis by selective amplification of DNA pools from high-fidelity microchips. Nature Biotechnology. 28 (12), 1295(2010).
- Richmond, K. E., et al. Amplification and assembly of chip-eluted DNA (AACED): a method for high-throughput gene synthesis. Nucleic Acids Research. 32 (17), 5011-5018 (2004).
- Schmidt, T. L., et al. Scalable amplification of strand subsets from chip-synthesized oligonucleotide libraries. Nature Communications. 6, (2015).
- Grass, R. N., Heckel, R., Puddu, M., Paunescu, D., Stark, W. J. Robust Chemical Preservation of Digital Information on DNA in Silica with Error-Correcting Codes. Angewandte Chemie International Edition. 54 (8), 2552-2555 (2015).
- Erlich, Y., Zielinski, D. DNA Fountain enables a robust and efficient storage architecture. Science. 355 (6328), 950-953 (2017).
- Garalde, D. R., et al. Highly parallel direct RNA sequencing on an array of nanopores. Nature Methods. 15 (3), 201-206 (2018).
- Cleary, M. A., et al. Production of complex nucleic acid libraries using highly parallel in situ oligonucleotide synthesis. Nature Methods. 1 (3), 241-248 (2004).
- LeProust, E. M., et al. Synthesis of high-quality libraries of long (150mer) oligonucleotides by a novel depurination controlled process. Nucleic Acids Research. 38 (8), 2522-2540 (2010).
- Lietard, J., Damha, M. J., Somoza, M. M. Enzymatic and Chemical Synthesis of Nucleic Acid Derivatives. Fernández-Lucas, J. , Wiley Online Books. (2018).
- Fodor, S. P. A., et al. Spatially Addressable Parallel Chemical Synthesis. Science. 251 (4995), 767-773 (1991).
- Singh-Gasson, S., et al. Maskless fabrication of light-directed oligonucleotide microarrays using a digital micromirror array. Nature Biotechnology. 17 (10), 974-978 (1999).
- Pease, A. C., et al. Light-Generated Oligonucleotide Arrays for Rapid DNA-Sequence Analysis. Proceedings of the National Academy of Sciences of the United States of America. 91 (11), 5022-5026 (1994).
- Holz, K., Lietard, J., Somoza, M. M. High-Power 365 nm UV LED Mercury Arc Lamp Replacement for Photochemistry and Chemical Photolithography. ACS Sustainable Chemistry & Engineering. 5 (1), 828-834 (2017).
- Lackey, J. G., Mitra, D., Somoza, M. M., Cerrina, F., Damha, M. J. Acetal Levulinyl Ester (ALE) Groups for 2′-Hydroxyl Protection of Ribonucleosides in the Synthesis of Oligoribonucleotides on Glass and Microarrays. Journal of the American Chemical Society. 131 (24), 8496-8502 (2009).
- Lackey, J. G., Somoza, M. M., Mitra, D., Cerrina, F., Damha, M. J. In-situ chemical synthesis of rU-DNA chimeras on chips and enzymatic recognition. Chimica Oggi-Chemistry Today. 27 (6), 30-33 (2009).
- Lietard, J., Ameur, D., Damha, M., Somoza, M. M. High-density RNA microarrays synthesized in situ by photolithography. Angewandte Chemie International Edition. 57 (46), 15257-15261 (2018).
- Agbavwe, C., et al. Efficiency, Error and Yield in Light-Directed Maskless Synthesis of DNA Microarrays. Journal of Nanobiotechnology. 9, (2011).
- Sack, M., et al. Express photolithographic DNA microarray synthesis with optimized chemistry and high-efficiency photolabile groups. Journal of Nanobiotechnology. 14, (2016).
- Kretschy, N., Holik, A. K., Somoza, V., Stengele, K. P., Somoza, M. M. Next-Generation o-Nitrobenzyl Photolabile Groups for Light-Directed Chemistry and Microarray Synthesis. Angewandte Chemie International Edition. 54 (29), 8555-8559 (2015).
- Sack, M., Kretschy, N., Rohm, B., Somoza, V., Somoza, M. M. Simultaneous Light-Directed Synthesis of Mirror-Image Microarrays in a Photochemical Reaction Cell with Flare Suppression. Analytical Chemistry. 85 (18), 8513-8517 (2013).
- Lietard, J., et al. Base-cleavable microarrays for the characterization of DNA and RNA oligonucleotides synthesized in situ by photolithography. Chemical Communications. 50 (85), 12903-12906 (2014).
- Krotz, A. H., et al. Solution stability and degradation pathway of deoxyribonucleoside phosphoramidites in acetonitrile. Nucleosides Nucleotides & Nucleic Acids. 23 (5), 767-775 (2004).
- Hargreaves, J. S., Kaiser, R., Wolber, P. K. The Degradation of Dg Phosphoramidites in Solution. Nucleosides Nucleotides & Nucleic Acids. 34 (10), 691-707 (2015).
- McGall, G. H., et al. The efficiency of light-directed synthesis of DNA arrays on glass substrates. Journal of the American Chemical Society. 119 (22), 5081-5090 (1997).
- Collett, J. R., et al. Functional RNA microarrays for high-throughput screening of antiprotein aptamers. Analytical Biochemistry. 338 (1), 113-123 (2005).
- Buenrostro, J. D., et al. Quantitative analysis of RNA-protein interactions on a massively parallel array reveals biophysical and evolutionary landscapes. Nature Biotechnology. 32 (6), 562-568 (2014).
- Wu, C. -H., Holden, M. T., Smith, L. M. Enzymatic Fabrication of High-Density RNA Arrays. Angewandte Chemie International Edition. 53 (49), 13514-13517 (2014).
- Tian, J., et al. Accurate multiplex gene synthesis from programmable DNA microchips. Nature. 432 (7020), 1050-1054 (2004).
- Lietard, J., et al. Mapping the affinity landscape of Thrombin-binding aptamers on 2'F-ANA/DNA chimeric G-Quadruplex microarrays. Nucleic Acids Research. 45 (4), 1619-1632 (2017).
Reimpressões e Permissões
Solicitar permissão para reutilizar o texto ou figuras deste artigo JoVE
Solicitar PermissãoThis article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. Todos os direitos reservados