
Method Article
Визуализация АТФ-синтазы димеров в митохондриях с помощью электронной Cryo-томографии
В этой статье
Резюме
Мы приводим протокол о том, как собирать и обрабатывать электронов крио-томограммы всей митохондрий. Методика обеспечивает детальное представление о структуре, функции и организации больших мембраны белковых комплексов в родных биологических мембран.
Аннотация
Электронно-крио-томографии является мощным инструментом в структурной биологии, способны визуализировать трехмерную структуру биологических образцов, таких как клеток, органелл, мембранных везикул, или вирусов на молекулярном подробно. Для достижения этой цели, водный образец быстро остеклованные в жидком этана, который сохраняет его в, замороженных гидратированных состоянии, близком к родным. В электронном микроскопе, серия тент записываются при температуре жидкого азота, из которого 3D томограммы реконструируются. Отношение сигнал-шум от объема томографического по своей природе низкой. Узнаваемый, повторяющиеся черты усиливаются subtomogram усреднения, при помощи которых отдельные subvolumes вырезаются, выровненной и усредненного для уменьшения шума. Таким образом, 3D-карты с разрешением 2 нм или выше могут быть получены. Подгонка имеющихся структур высокого разрешения в 3D-объем, то производит атомные модели белковых комплексов в их родной среде. Здесь мы показываем, как мы используем электронную крио-tomograpHY для изучения на месте организацию крупных мембраны белковых комплексов в митохондриях. Мы обнаружили, что АТФ-синтазы организованы в строки димеров вдоль изогнутых высокой вершинах внутренней мембраны крист, в то время как комплекс я случайным образом распределены в мембране областей по обе стороны от рядов. По subtomogram усреднения мы получили структуру митохондриального АТФ-синтазы димера в крист мембраны.
Введение
Митохондрии являются силовые дома клетке. Путем преобразования электрохимический протонный градиент через внутреннюю митохондриальную мембрану в энергию химических связей, митохондриальная АТФ-синтазы производит большую часть АТФ, что приводит клеточные процессы. Для того чтобы понять механизмы, лежащие преобразования митохондриального энергетического, мы должны определить структуру АТФ-синтазы на месте, и, чтобы узнать, как она устроена и распространяется в внутренней мембране митохондрий. Хотя с высоким разрешением структуры большинство митохондриальных АТФ-синтазы компонентов 1-3 и карт с низким разрешением целого комплекса 4 доступны, важно установить структуру и конформацию рабочего фермента в мембране. Распределение АТФ-синтазы в внутренней мембране митохондрий было принято считать случайными, но рано нахождения 5 и наши собственные первые результаты 6 указано, что это не тон так. Последующие исследования, проведенные в нашей группе и другие 7 подтвердили, что АТФ-синтазы устроена длинными рядами димеров вдоль плотно изогнутых хребтов внутренней митохондриальной мембраны крист 8, в то время как протонные насосы электрон-транспортной цепи, кажется, расположены по обе стороны строк 9. Это устройство имеет важные последствия для механизмов преобразования митохондриального энергии.
Методика, которую мы использовали для определения этой договоренности является электронной крио-томографии (крио-ET). Крио-ET в настоящее время является единственным методом, который обеспечивает точные трехмерные (3D) объемы клеток, клеточных отсеках или органеллы в молекулярным разрешением. Крио-ET особенно подходит для изучения больших комплексов в биологических мембранах, так как мембраны появляются с хорошим контрастом и легко проследить в объемах 3D томографических.
Другие методы для изучения 3D структуры клеток или органеллES не обеспечивают молекулярную деталь. Супер-разрешение световой микроскопии 10, 11 превосходно на выявление позиции или расстояния между светоизлучающих ярлыками к белкам интерес с точностью в десятки нм, но не раскрывает структуру самого белка, даже при низком разрешении . Просвечивающей электронной микроскопии из последовательных секций 12 или блок-лицевой изображений методом сканирующей электронной микроскопии 13 пластиковых встраиваемый биологических образцов открывается вид с низким разрешением клеточных объемов, но аналогичным образом не раскрывают молекулярную деталь. Атомно-силовая микроскопия 14 может в принципе поставить молекулярную или даже атомное разрешение, но только на поверхности объектов на атомно-плоской, твердой подложке. Наконец, рентгеновской томографии 15 или рассеяния интенсивных рентгеновских импульсов от лазеров на свободных электронах 16 вряд ли выявить структуру крупных, сложных, апериодических объектов, таких как целые клетки или органеллы в мolecular разрешение в обозримом будущем. Таким образом, в настоящее время не существует альтернативы крио-ET для изучения 3D-структуры клетки или органеллы в нанометровым разрешением.
Крио-ET является методом выбора для исследования структуры и конформации ассоциированных с мембранами белков сборок в том числе комплекса ядерной поры 17, шип гриппа образует комплекс 18, и жгутики моторных белков 22, 23, но и организация целых бактериальных клеток 19 и вступление патогенных вирусов, таких как ВИЧ и клеток 20-23. Крио-ET имеет неоценимое значение для визуализации нитевидные белки и их взаимодействия в клетке, в том числе нитей актина 24 или аксонем 25. Разрешение может быть повышена до 2 нм или лучше по subtomogram среднем 26, в результате чего subvolumes повторных, правильными чертами вырезаются из объема томографического и усредняются имидже одночастичномметоды переработкой.
Крио-ET включает в себя приобретение ряда проекционных изображений тонкой образца (<250 нм), принятым на различных углах наклона в электронном микроскопе (ПЭМ). Образец должен быть тонким, чтобы электроны, которые сильно взаимодействуют с веществом, не разбросаны не более одного раза. Многократное рассеяние делает качество полученного изображения трудно интерпретировать и уменьшает контраст. Изображения выбранной области образца были выровнены относительно друг друга и прогнозируемых в 3D-пространстве с помощью подходящего компьютерной программы, генерации 3D объем образца. Выравнивание изображений автоматизированного золотыми координатных меток, которые смешиваются с образцом до замораживания. В идеальном случае 10 или более равномерно распределены координатных меток должны присутствовать в каждом изображении, чтобы добиться хорошего согласования.
Для наблюдения молекулярную деталь, образцы окунуться заморозки в жидком этана, который сохраняет свой родной гашеной состояние. Замораживания в жидкомэтана так быстро (~ 10 5 ° С / сек) 27, что вода не кристаллизуется, но остается в стекловидной, стекло, как состояние. Ледяной кристалл повреждает формирования чувствительные биологические структуры. Как биологические образцы страдают от радиационных повреждений, есть предел, к общему числу рассеивающих событий образец может терпеть. Изображения, таким образом, приобрела в режиме низких доз: сфера интересов идентифицируется при малом увеличении (1,500X) с дозой электронов ниже 1 е - / нм 2 (режим поиска). Затем изображение фокусируется на большем увеличении, от прочих интересных (режим фокусировки). Только когда изображение приобрела, область интересов облучают более высокой дозе электронов (режиме экспозиции).
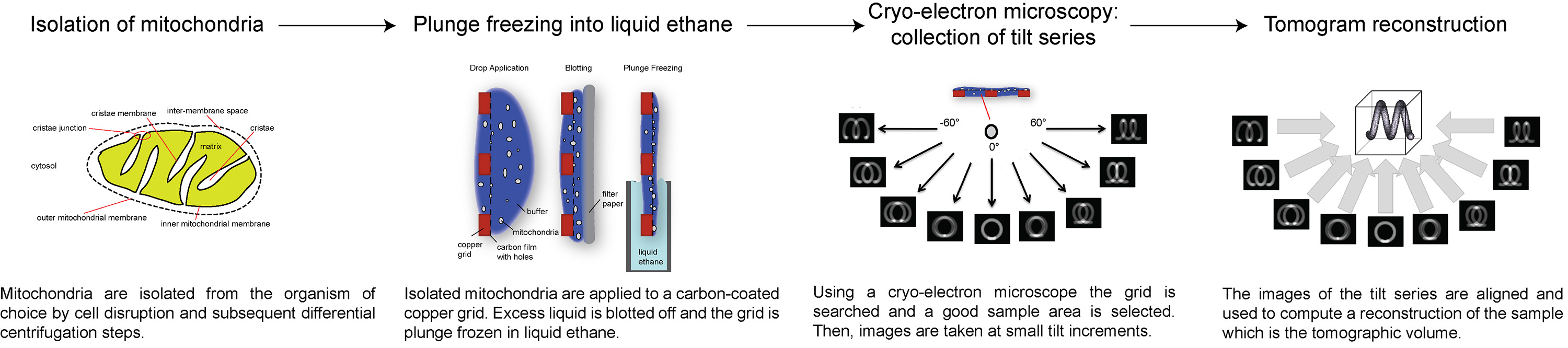
Здесь мы представляем обзор того, как сбор и обработка электронных криопроводники томограммы, используя АТФ-синтазы димеры в внутренней мембране митохондрий в качестве примера. Следующий протокол описывает, как подготовить митохондрии для крио-ЭТ, как настроитьи собрать наклона серии с определенной общей дозы электронов, и как обрабатывать серию наклона для получения 3D объем интересующей области. Обзор процедуры показан на рисунке 1.
протокол
1 Получение митохондрий из клеток или тканей дифференциальным центрифугированием
В этом разделе описывается общая процедура для выделения интактных митохондрий из различных эукариотических организмов. Точное буферные компоненты и скорость центрифугирования должны быть оптимизированы для каждой ткани / изученных видов.
- Перерыв клеток в изотоническом буфере (например, 250 мМ сахарозы, 10 мМ HEPES, рН 7,4), с использованием стекла шаровой мельницы (грибковые мицелий) 28, ферментативное расщепление клеточной стенки (Saccharomyces CEREVISIAE) 29, на подшипниках гомогенизатора (один -клетка эукариоты / Культура клеток / нематоды) 30 или блендер (животных или растительных тканей) 31.
- Удалить клеточного дебриса путем фильтрации через муслина с последующим низкоскоростным центрифугированием (2000 х г, 4 ° C, 10 мин).
- Соберите супернатанта и осадка митохондрии от высокоскоростного центрифугирования (9000 мкг в, 4 ° C, 10 мин).
- При необходимости используйте изотонический шаг градиента плотности для дополнительной очистки митохондриальной фракции 32.
2 Получение митохондрий для Electron Cryo-томографии
В следующем разделе описывается, как получить замороженные гидратированных образцов для крио-ЭТ. ПРИМЕЧАНИЕ: Метод предполагает использование исключительно холодной жидкого азота и этана, которые могут вызвать серьезные ожоги кожи. Защитные очки и перчатки крио-защиты необходимо носить. Жидкость этана, который также легковоспламеняющиеся, необходимо обращаться в вытяжной шкаф.
- Ресуспендируют осаждают митохондрии в 250 мМ трегалозы, 10 мМ HEPES буфере при рН 7,4 до концентрации приблизительно 5 мг / мл общего белка.
- Тлеющего разряда дырявые углерода EM сетки, боковые углерода до, в вакуумной устройства в соответствии с инструкциями производителя.
- Сжижение несколько миллилитров этана, направляя поток этана газа на внутренней стороне жидкого азота прохладыред алюминия контейнер.
- Смешайте белок-сопряженных золотую Fiducial дорог 1: 1 с митохондриальной суспензии и сразу же применить 3 мкл для тлеющего разряда EM сеткой, в пинцетом.
- Поместите пинцет в остекловыванию устройства, например, самодельные гильотины. Пятно от избытка жидкости с клином фильтровальной бумаги (~ 5 сек или до жидких остановок распространения) и сразу же погружают сетку в жидкий этан, отпустив курок.
- Трансфер сетку от жидкого этана в жидкий азот. Во время передачи, удаления избытка этана из сетки с фильтровальной бумагой. Поместите кончик клиновидного кусочек фильтровальной бумаги в жидкую этана. Когда жидкость этан поднимается, мягко тянуть сетку до фильтровальной бумаге, но держать его ниже уровня жидкости перед. Жидкость этана удаляется из сетки под действием капиллярных сил и как только вся жидкость этан был удален, немедленно передать сетку в жидкий азот.
- Поместите витрифицированном сетку в ящике для хранения сетки,й хранить в жидком азоте для последующего использования в случае необходимости.
3 Запись томографических Tilt серии
В следующем разделе описывается, как настроить и собрать томографическое наклона серию митохондрии с Polara электронного микроскопа, оснащенного энергетической фильтра после столбца и ПЗС-камеры. Подобные протоколы используются со всеми электронными крио-микроскопов, оснащенных ПЗС или прямых детектора электронов камер.
- Совместите микроскоп
- Вставьте тест образец например, графита и золота острова на дырявой углеродной пленки.
- Выберите режим поиска в системе низких доз микроскопа.
- Принесите образец eucentric высоте. Это точка минимального движения ху при наклоне держатель образца. Сосредоточьте достопримечательность при 0 ° наклона, наклоните этап до 20 ° и повторно отцентрировать точку, изменяя Z-высоты. Вернуться в 0 ° и повторять, пока боковое смещение не минимизируется.
- Выберитережим экспозиции. Выберите нужный увеличение сбора томограммы (например, 25,000X на детекторе ≈ 0,6 нм образца размер пикселя).
- Выберите небольшой конденсатор диафрагмы (50-70 мм) и выберите размер пятна и интенсивность пучка так, чтобы луч просто шире, чем устройство обработки изображений и дает пикселя чтение 60 е - / пиксел (CCD) или 14 е - / пиксель / сек (прямой электронный детектор, режим подсчета).
- Центр конденсатор диафрагмы.
- Найти Gaussian фокусировки, например, точку минимальной контрастностью. Сброс чтение микроскоп расфокусировки и правильные точки разворота и центр вращения в соответствии с инструкциями изготовителя.
- Наберите в нужное расфокусировки для записи томограмму. ПРИМЕЧАНИЕ: Высокая расфокусировки (8 мкм) увеличивает контраст, но уменьшает разрешение, в то время как низкий расфокусировки (2-4 мкм) увеличивает разрешение за счет контраста.
- За пустой отверстие, генерировать новую ссылку усиления и выравнивания энергии фильтр в соответствии с производителеминструкции.
- Совместите поиска и воздействия режимы. В режиме экспозиции, центр достопримечательность и переключения в режим поиска. Выберите увеличения из 1,500X (0,033 мкм / пиксель образца на детектор) и расфокусировки 100 мкм (для повышения контрастности). Принесите достопримечательность спине к центру с помощью смещения изображения катушки.
- В режиме поиска, настроить размер пятна и интенсивность пучка так, чтобы луч просто шире, чем устройство обработки изображений и дает пикселя чтение ~ 20 е - / пиксел (CCD) или ~ 8 е - / пиксел / сек (прямой детектора электронов, счетном режиме).
- Поиск прекрасный образец Area
- Вставьте сетку с замороженных гидратированных митохондрий в электронном микроскопе при температуре жидкого азота (обратитесь к инструкции EM-изготовителя).
- В режиме поиска, поиск сетку для области соответствующей толщины льда и образцов качества. Возьмите поиска 6 сек образ перспективных направлений для определения пригодности для сбора томограммы. Botч внутренней и внешней митохондриальной мембраны должна быть видна при этом увеличении.
- Запись томографического Tilt серии
- После того, как хорошая область образец найден, наклоните этап ± 60 ° для определения максимального диапазона наклона, который доступен без обструкции воздействия или зона фокусировки сетевыми баров или льда комков.
- На соседнем льдом заполнено отверстие внешнего сходства, переключиться в режим экспозиции и регулировки времени интенсивности пучка или захвата изображений, так что каждый записанное изображение имеет электронный дозу 30-50 е - / пиксел для ПЗС или 6-8 е - / пиксель / с прямые электронные детекторы, режим подсчета.
- Рассчитать коэффициент распределения дозы (I 0 / I 60) путем деления средневзвешенное количество электронов для 1 сек изображения, полученного при 0 ° с, что из 60 ° изображения. Это отношение описывает увеличение срока экспозиции, необходимое для поддержания постоянной количество электронов каждого изображения при увеличении угла наклона (время экспозиции = 1 / сОС (α), где N (I 0 / I 60) = 2 л). Отношение также служит хорошим указанием толщины льда. Хорошие томограммы митохондрий обычно записываются с I 0 / I 60 = 2,3-2,6.
- За пустой отверстие, приобрести 1 сек изображение в режиме экспозиции и обратите внимание на количество электронов в расчете на одну 2. Учитывая соотношение дозы распределения, рассчитать общее количество изображений, которые можно записать для определенного общей дозы электронов (например, <40 е - / 2 для определения структуры и ~ 160 э - / 2 для морфологии).
- Определите соответствующий интервал наклона для сбора томограммы путем деления общего диапазон наклона (например, 120 ° для ± 60 °) на общее количество изображений, рассчитанных в 3.3.4.
- Настройка и записать томограмму с параметрами, определенных выше, с использованием соответствующих автоматического сбора данных программное обеспечение 33, 34. Серия Tilt обычно начинаются на ± 20 ° и пройти через 0 ° до достижения высоких наклоны в целях максимизации информационного наполнения низкого наклона изображений, который разрушенные увеличением электронного дозы.
4 Создание и сегментация томографических томах
В этом разделе описывается томографические объемы митохондрий сгенерированы наклона серии и, как объемы присутствуют для общего просмотра.
- Сохраните серию томографическое наклона к соответствующему каталогу. Генерация стека изображений и конвертировать в нужный формат файла с программным обеспечением с открытым исходным кодом, таких как dm2mrc или tif2mrc (IMOD пакета), который преобразует .dm3, .dm4 или .tif файлы MRC стеков. MRC стеки необходимы для томографической реконструкции с УПМ 35. Другие пакеты требуют различных форматов.
- Совместите изображения и генерировать томограмму, следуя инструкциям, подробно описанные в учебнике IMOD ( http://bio3d.colorado.edu/imod/doc/etomoTutorial.html ).
- Повышение контрастности томограммы, используя нелинейный фильтр анизотропной диффузии распределенную с УПМ. Этот фильтр хорошо работает для мембран и мембранных ассоциированных частиц, таких как АТФ-синтазы.
- Для визуализации, вручную сегмент томограмма с использованием коммерчески доступных программ, например, Amira. Назначение вокселей, соответствующие внутренней или наружной мембраны и генерировать поверхности. С помощью опции кликер в EM-пакета плагин для Amira 36 отметьте расположение частиц АТФ-синтазы.
5 Subtomogram Усреднение АТФ-синтазы димеров и Место рентгеновских структур
В следующем разделе описывается, как subtomogram средние АТФ-синтазы димеров может быть получен.
- Использование отмеченные частицы в качестве входных и соответствующего программного пакета, таких как «Частиключицы Оценка для программы электронной томографии ', вычислить среднее subtomogram.
- Для оценки разрешения, сравнить два независимо определены subtomogram средние от Фурье оболочки корреляции 37.
- Если есть возможность, док известно рентгеновские структуры в среднем subtomogram по твердого тела установки, либо вручную, либо с помощью автоматического последовательные док программ, как те, в программе Chimera 38.
Результаты
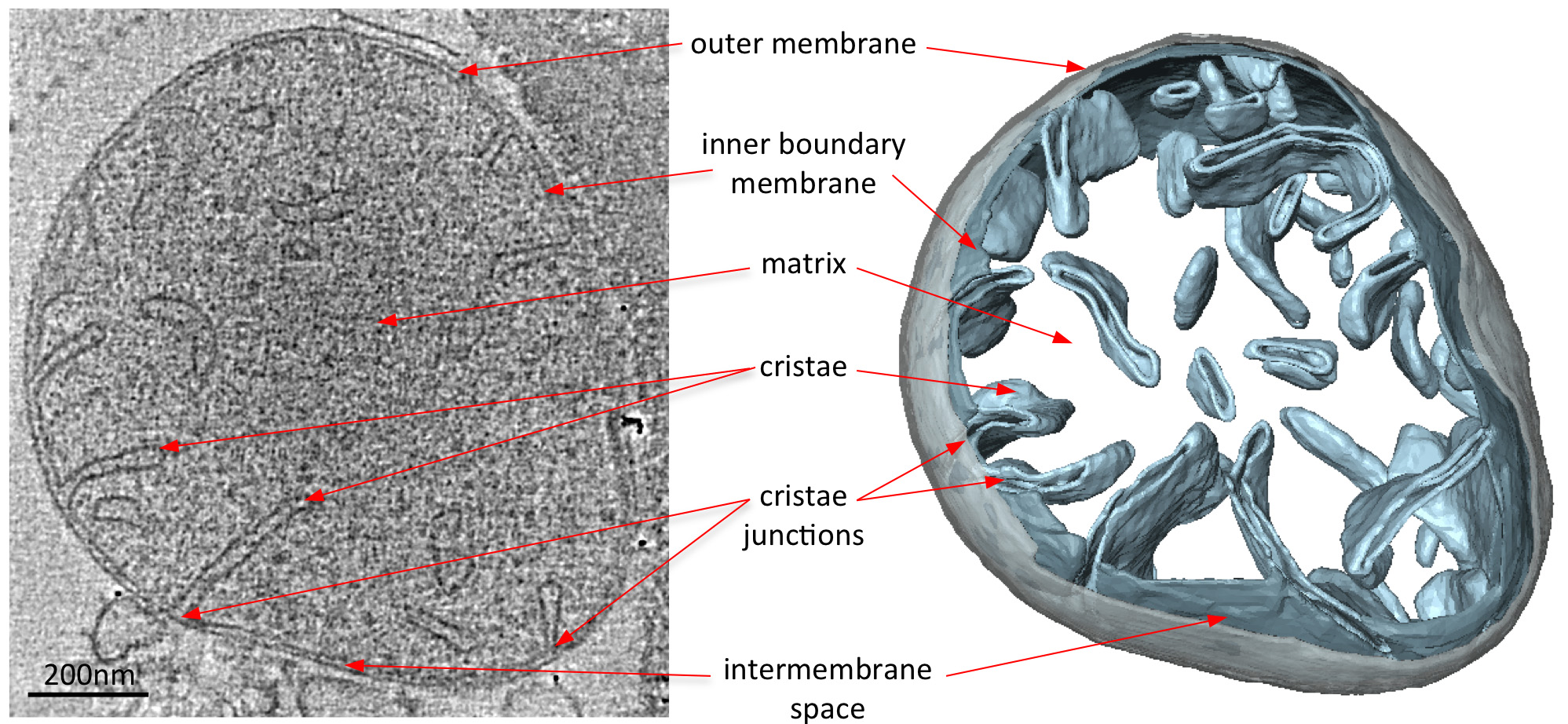
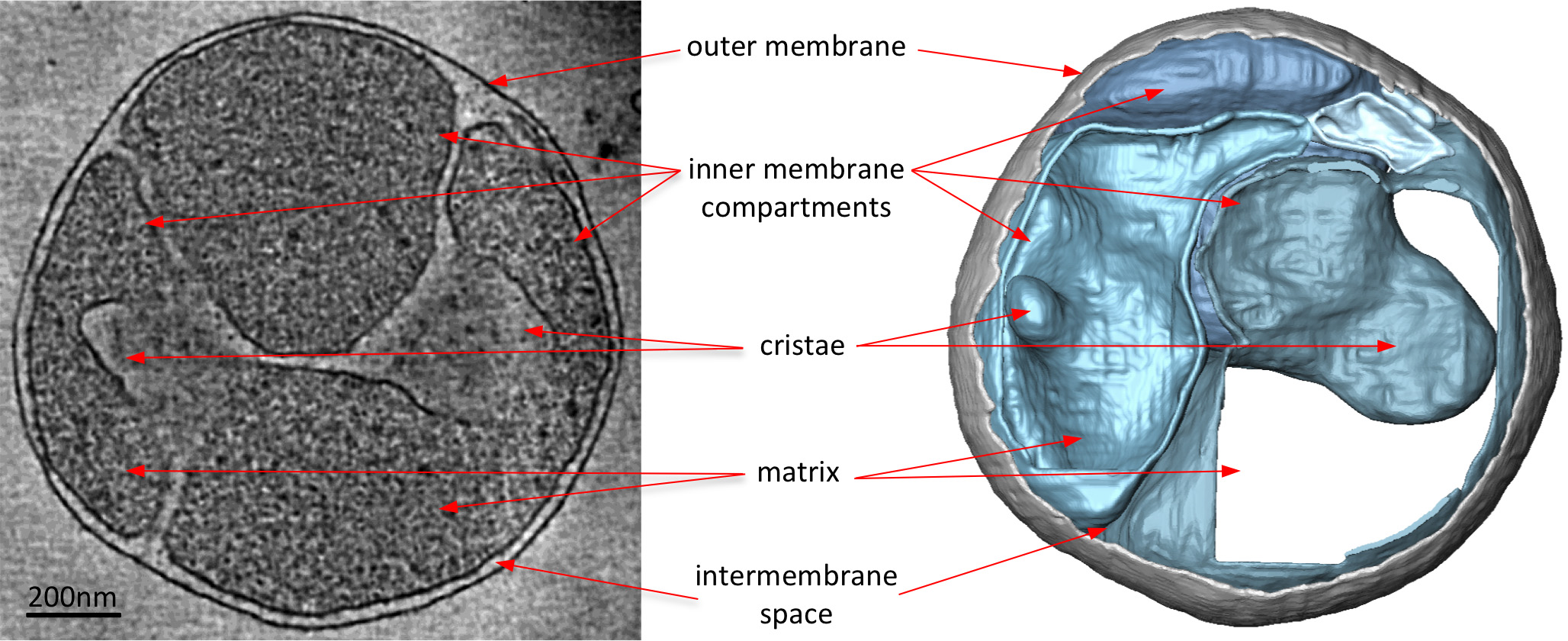
Электронные криопроводники томограммы митохондрий четко выявить 3D морфологию органелл (Рисунок 2). Руководство сегментация мембран в объеме томографического иллюстрирует структуру крист в митохондриях. При получении изображений митохондрии из различных штаммов дрожжей с нокаутом, которые не имеют определенные белковые компоненты, действие этих белков на крист морфологии может быть оценена. 3 показан митохондрии от штамма дрожжей не хватает субъединицы АТФ-синтазы е. Этот компонент комплекса синтазы АТФ требуется для димеризации митохондриального АТФ-синтазы. Митохондрии из этого штамма не имеют нормальную слоистую крист митохондрий дикого типа (фигура 2), а вместо этого содержат ряд внутренних мембранных отсеков. Эти отсеки либо лишены крист или содержать мелкие шарообразную форму мембранные инвагинации (рисунок 3).
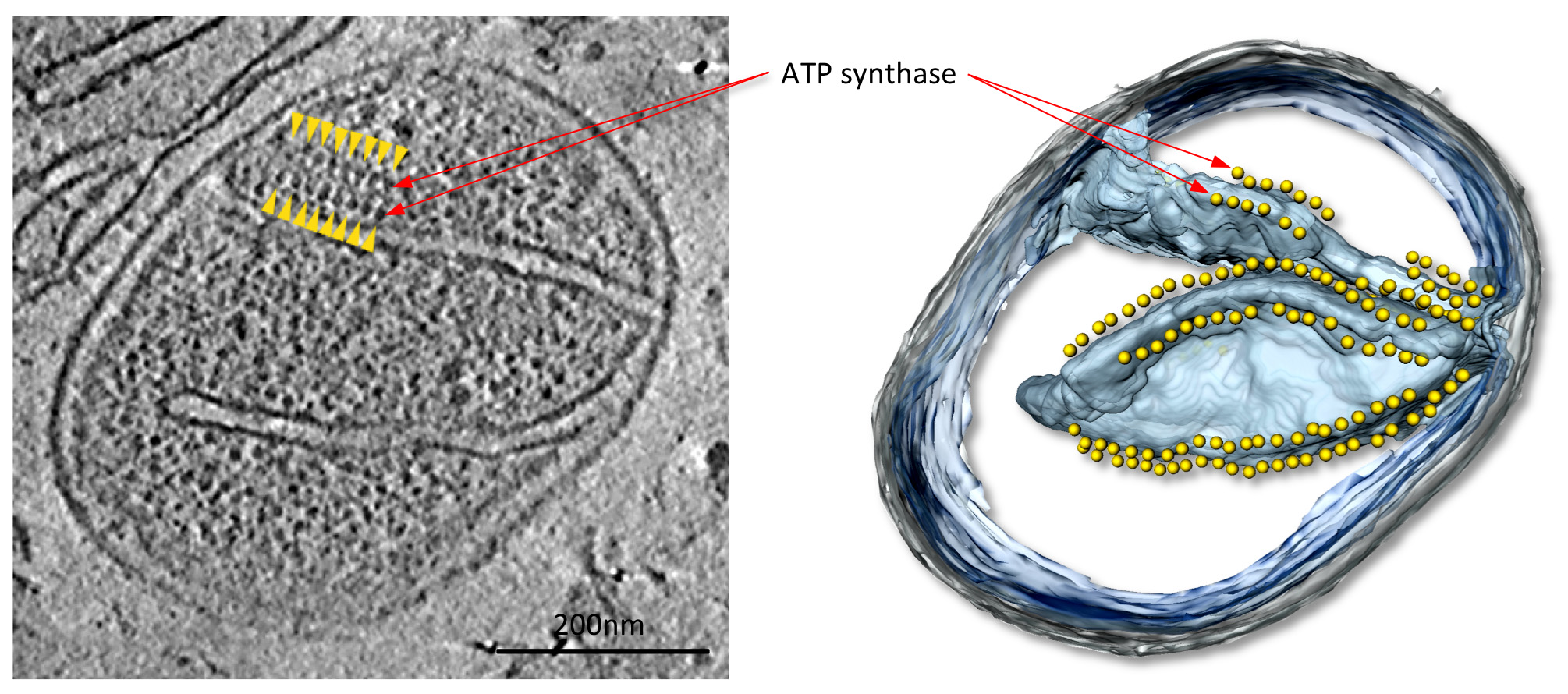
В Томаograms с хорошим контрастом, крупных митохондрий белковых комплексов, в данном случае АТФ-синтазы димеров, хорошо видны (Рис 4; Фильм 1). Структуры комплексов могут быть определены при разрешении нм 2-3 по subtomogram усреднения (фиг.5; Movie 2). Средние объемы могут быть помещены обратно в томограммы, чтобы оценить организацию отдельных комплексов по отношению друг к другу и в других белковых комплексов в мембране (фиг.6; Movie 3).

Рисунок 1. блок-схема, показывающая этапы электронного крио-томографии. Кликните здесь, чтобы посмотреть увеличенное изображение.
{kind=link}

Рисунок 2 Морфология митохондрии от дикого типа S. CEREVISIAE. Центральный срез через объем томографического из дикого типа С. CEREVISIAE митохондрия (слева) и соответствующая поверхность оказанные объем (справа). Сегментированный объем внешней мембране будет серым и объемы внутренней границе и крист мембраны в светло-голубой. По материалам Дэвис и др 8. Нажмите, чтобы увеличить изображение.
{kind=link}

Рис.3 митохондрии из S. CEREVISIAE штамм хватает субъединицу, необходимое дляАТФ-синтазы димеризации. Нарежьте через томографического объема (слева) и сопровождающих поверхности оказанные объем (справа) митохондрии от S. CEREVISIAE напряжение отсутствует необходимый для АТФ-синтазы димеризации на белок субъединицы е. По сравнению с фиг.2, митохондрии из мутантного штамма отсутствует нормальный слоистую крист митохондрий дикого типа. Вместо этого, митохондрия имеет много внутренней мембране карманов либо не крист или шарообразную форму гребешков. Таким образом электрон крио-томография подчеркивает изменения в мембраны морфологии в связи с генными удалений. По материалам Дэвис и др 8. Нажмите, чтобы увеличить изображение.
{kind=link}

Figurе 4 Митохондрии из гриба P. гусиной. Нарежьте через томографического объема (слева) и сопутствующей поверхности оказанные объема (справа) митохондрии от гифомицетов П. гусиной. В этом томограммы, ряды 10 нм частиц (желтые стрелки) расположены над высоко загнутыми мембранных хребтов во внутренней мембране крист (см фильм 1). Эти частицы были идентифицированы как АТФ-синтазы димеров по subtomogram усреднения. От Дэвис и др 9. Нажмите здесь, чтобы увеличить изображение.
{kind=link}

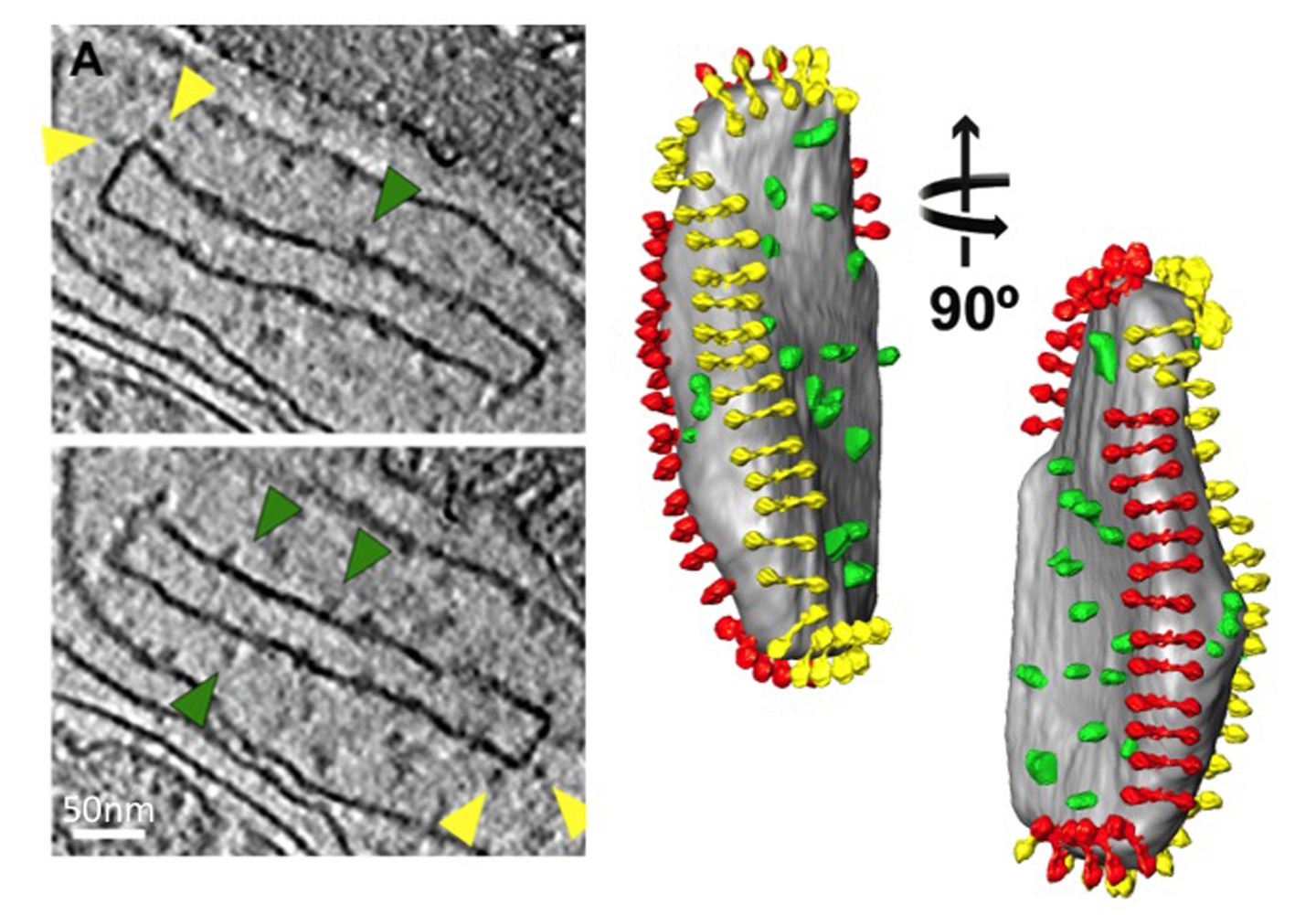
Рисунок 5 Структура и организация митохондриальной АТФ-синтазы. Боковые и вид сверху, показывающий плотность электронов АТФ-синтазы димера из S. с erevisiae как определяется subtomogram усреднения со встроенными атомных моделей (слева). Внутренней мембраны митохондрий пузырек показывает организацию АТФ-синтазы димеров в рядах (справа). На рисунке был сгенерирован путем установки subtomogram среднем синтазы димера АТФ в сегментированной объема мембранной везикулы, используя координаты, вычисленные при усреднении. По материалам Дэвис и др 8. Атомные модели: F 1 / ротор-кольцо [PDB: 2WPD] 39 (синий и фиолетовый); олигомицин чувствительной придающего белка-OSCP [PDB: 2BO5] 40 (зеленый); периферическая фрагмент стебля [PDB: 2CLY] 1 с N-концевых остатков [PDB: 2WSS] 2 (желтый и красный) (см фильм 2). Нажмите сюда, чтобы посмотреть увеличенное изображение.
{kind=link}
_upload / 51228 / 51228fig6highres.jpg "ширина =" 500 "/>
Рисунок 6 Изолированные Криста пузырек из P. гусиной митохондрия. Ломтики от объема томографической (слева) и сопутствующей поверхности оказанные объема (справа) о Криста пузырька из P. гусиной. Плотности Белковые выступающие из мембраны хорошо видны. Плотность указанные желтыми стрелками являются АТФ-синтазы димеры, указанный subtomogram усреднения. Зеленые стрелки указывают на плотностей, определенных маркировки антител как NADH дегидрогеназы (комплекс 1; подробнее см 9). Сегментация плотностей белка показывает свою организацию в крист, с синтазы димеров АТФ (красные и желтые), образующих строки вдоль высоко загнутыми крист хребтов и NADH дегидрогеназы комплексов (зеленые) в мембранных регионах по обе стороны рядами (см Фильм 3). От Дэвис и др 9.28 / 51228fig7highres.jpg "целевых =" _blank "> Нажмите здесь, чтобы увеличить изображение.
Фильм 1 Electron крио-томограмма P. гусиной митохондрия. фильм показывает последовательные срезы через объем томографического взятой из митохондрии от гифомицетов П. гусиной. Ряды АТФ-синтазы обозначены желтыми стрелками. Поверхность вынесено, сегментированный объем показано расположение АТФ-синтазы (желтые сферах) по отношению к структуре 3D крист. Внешний мембрана, серый; Внутренняя граница мембраны, прозрачно-голубой; крист мембраны, непрозрачный синий. От Дэвис и др 9. Смотрите также Рисунок 4. Нажмите здесь, чтобы посмотреть видео.
Фильм 2 Subtomogram среднем митохондриальной АТФ-синтазы димера от S. CEREVISIAE со встроенными атомных моделей. Средняя былрассчитывается от 121 subvolumes. Плотность отображается на трех уровнях контурных: 1с - сетка, 2s - светло-серый и 3s - темно-серый. Атомные модели были оснащены в плотности с помощью последовательного подходят рутину в Химера. Атомные модели: F 1 / ротор-кольцо [PDB: 2WPD] 39 (синий и фиолетовый); олигомицин чувствительной придающего белка-OSCP [PDB: 2BO5] 40 (зеленый); периферическая фрагмент стебля [PDB: 2CLY] 1 с N-концевых остатков [PDB: 2WSS]. 2 (желтый и красный) Нажмите здесь, чтобы посмотреть видео.
По материалам Дэвис и др 8. Смотрите также Рисунок 5.
Фильм 3 Изолированные гребни пузырек из P. гусиной митохондрии. Последовательные срезы через объем томографического показаны с последующим сегментированного поверхностно-визуализации объема. Плотности Белковые выступающие из мембранныхе отчетливо видны. Красные и желтые плотности являются АТФ-синтазы димеры. Зеленые плотности являются NADH дегидрогеназы (комплекс I), как определено маркировки антител. От Дэвис и др 9. Смотрите также Рисунок 6. Нажмите здесь, чтобы посмотреть видео.
Обсуждение
Протокол, представленные здесь, представляет собой введение в крио-ET и subtomogram усреднение митохондрий, но по существу та же процедура может быть применена к любой другой отсек клетки или мембраны. Для получения наилучших данных, критические шаги в ходе процедуры являются пробоподготовки, стратегию приобретения окунуться процесс замораживания и данных. Качество сэмплирования, которая имеет решающее значение для успеха, зависит от оптимизированного протокола замерзания обеспечить подходящую толщину льда, который имеет первостепенное значение для хорошей контрастностью изображения. Оптимальная стратегия сбора данных зависит от инструмента и образца. Параметры, которые будут оптимизированы включают электронный дозу на изображение, схему наклона и расфокусировки. Приобретение хорошего томограмму хороший образец делает все дальнейшая обработка шаги легче и обеспечивает удовлетворительный конечный результат.
Крио-ET сочетании с subtomogram усреднения и атомной модели фитинга содержит подробную информацию о том, как белковые комплексы расположены в гоEIR родной сотовой среда. Техника в равной степени подходит для исследования структуры крупных мембраны белковых комплексов, таких как supercomplexes дыхательной цепи (1.7 MDA), АТФ-синтазы димеров (2x500 кДа), или поровый комплекс ядерной (~ 120 MDA) 8, 9, 17. Организация АТФ-синтазы димеров в ряды не могут наблюдаться с помощью методов высокого разрешения, таких как рентгеновская кристаллография, ЯМР или одночастичном крио-ЭМ, потому что димерных рядов нарушается путем экстракции моющего средства, что является необходимым шагом в изоляции и очистка мембраны белковых комплексов.
Расположение АТФ-синтазы в ряды димеров вдоль гребней крист является универсальным организация принцип митохондрий у всех видов. Протон-насосные комплексы электрон-транспортной цепи, в частности комплекса I (NADH дегидрогеназы), расположены в мембранных областях обе стороны от строк 8,9. Эта организация дыхательной цепи имеет глубокое влияние на митохондриальных биоэнергетики. Если димерных рядов не могут образовывать связи с отсутствие димерных конкретных белковых субъединиц, клетки обладают более длительного времени генерации и снижение клеточного пригодность, как это наблюдалось в дрожжевой мутанта, показанной на рисунке 3 41. С ростом интереса к митохондриальных заболеваний, подробное понимание молекулярных основ правящей ультраструктуры митохондрий и функции имеет первостепенное значение. Электрон крио-томография обеспечивает связь между структурой белка определяется методов высокого разрешения, и распределения и расположения этих белков в мембране в масштабе нанометров. Это делает крио-ЭТ является важным инструментом для понимания митохондриальной структуры и функции в норме и патологии.
Дальнейшие технические усовершенствования и улучшения, в крио-ЭТ включают в себя стратегии белок-маркировки для идентификации положения Proteiп субъединицы в макромолекулярных комплексов или расположение небольших или менее различных белков (<0,5 MDA) в клетках. Кроме того, методы обработки гибрид EM, которые сочетают subtomogram усреднение с анализа одного частиц или спиральной реконструкции недавно определяется белковые структуры в ~ 8 Å 42, 43. Эти методы обработки в настоящее время ограничивается очищенных белков, которые хорошо разделенных во льду или образуют спиральные узлы и в настоящее время не применяется в переполненных сотовых сред, как митохондрии и крист мембраны. Для сбора данных, новые вычислительные и аппаратные средства разрабатываются для автоматизированного сбора томограмма, увеличить контрастность изображения и снизить общую требуемую электронную дозу. Единственное фундаментальное ограничение в крио-ЭТ является излучение повреждение образца электронным пучком. Это означает, что только очень низкие дозы электронов может быть использован для записи каждого изображения из серии томографическое наклона, что приводит к плохой сигнал TСоотношение что в конечном итоге ограничивает достижимую разрешение о-шум. Новые прямые электронные детекторы, выпущенные менее года назад, в настоящее время революцию в поле одночастичном крио-ЭМ 44, 45. Эти новые детекторы обеспечивают более высокую контрастность и лучшее разрешение при низких электронных дозах. Для электронного крио-томографии, это означает, что наклон ряд с меньшими размерами шагом или даже двумя томограмм оси могут быть собраны без опасений по поводу чрезмерного радиационного повреждения (один ПЗС эквивалентно в электронном дозы до 5 прямых образов детектора электронов). Обширные объемы данных, полученных этими детекторами создавать свои собственные проблемы в данных обработки, переработки и хранения, которые будут иметь преодолеть.
Кроме того, фазовые пластины, которые работают на аналогичных принципах, которые используются обычно в световой микроскопии, чтобы добавлять фазового контраста, в настоящее время разрабатываются для просвечивающей электронной микроскопии 46, 47. Это должно позволить томограммы быть собраны ближе, чтобы сфокусировать и, следовательно, с более высоким разрешением, в то же время сохраняя при этом низким разрешением признаки, необходимые для выравнивания и интерпретации томографических объемов. Взятые вместе, эти технологические достижения значительно расширить спектр биологических вопросов, которые могут быть решены с помощью крио-ЭТ.
Раскрытие информации
Авторы заявляют, что они не имеют конкурирующие финансовые интересы.
Благодарности
Эта работа была поддержана Общества Макса Планка (КМД, BD, AWM, туберкулеза, TBB, DJM, и WK), Кластер Превосходства Франкфурте "макромолекулярных комплексов», финансируемого Немецкого исследовательского (WK) и докторской ЕМВО долгосрочных Братство (VAMG).
Материалы
| Name | Company | Catalog Number | Comments |
| Sucrose | Roth | 4621.2 | |
| Trehalose | Sigma-Aldrich | T9449 | |
| HEPES | Sigma-Aldrich | H3375 | |
| Zymolase | biomol | Z1000.1 | |
| Density gradient medium | Sigma-Aldrich | D1556 or P1644 | |
| Protein A gold | Aurion | 810.111 | 10 nm |
| Ethane | Air Liquide | P0502S02R0A001 | |
| Nitrogen | Air Liquide | I4150RG | |
| Consumables | |||
| Filter paper | Whatman, GE Healthcare | 1004 090 | Nr 4 |
| Tweezers | Dumont | T506 | Non-magnetic, pattern #5 |
| EM combined test grid | Plano GmbH | S142 | grapite and gold islands on holey carbon film |
| Holey carbon grids | Quantifoil Micro Tools GmbH or C-flat | R2/2 300 Cu mesh | |
| Cryo grid boxes | Plano GmbH | 160-42 | Alternative home-made metal boxes |
| Potter | Sigma-Aldrich | P7984 | |
| Blender | Waring | 8011S | |
| Glass bead mill - bead beater | BioSpec Products | 1107900-105 | Including 0.5 mm beads |
| Ball-bearing (Balch) homogeniser | isobiotec | H8 | |
| Centrifuge | Thermo Scientific | 46915 | Sorvall RC6+ |
| Glow-discharge apparatus | Bal-Tec A.G. | CTA 010 | |
| Grid vitrification device | FEI, Gatan or Leica | Alternative home-made plunger | |
| Transmission electron microscope | FEI or Jeol | 300 keV | |
| Direct electron detector | Gatan | K2 summit | 4k x 4k pixels, counted mode |
| Charge coupled device (CCD) | Gatan | US4000 | 4k x 4k pixels (alternative to Direct electron detector) |
| Energy filter | Gatan or Jeol | ||
| Software | |||
| Image acquisition software | Gatan | ||
| Tomogram acquisition software | Gatan, FEI, USCF, or UC-Boulder | 4 alternatives | |
| Tomogram processing software | FEI, UCSF or UC-Boulder | 3 alternatives | |
| Subtomogram averaging software | UC-Boulder, or University of Basel | 2 alternatives | |
| Tomogram visualization software | UC-Boulder, FEI, or University of Utah | 3 alternatives | |
| Visualization software plugin | Goethe university | http://www.biophys.uni-frankfurt.de/frangakis/ | |
| Structure visualization software | UCSF | http://www.cgl.ucsf.edu/chimera/ | |
| Numerical calculation software | MathWorks | http://www.mathworks.de/academia/ | |
Ссылки
- Dickson, V. K., Silvester, J. A., Fearnley, I. M., Leslie, A. G., Walker, J. E. On the structure of the stator of the mitochondrial ATP synthase. The EMBO journal. 25, 2911-2918 (2006).
- Rees, D. M., Leslie, A. G., Walker, J. E. The structure of the membrane extrinsic region of bovine ATP synthase. Proceedings of the National Academy of Sciences of the United States of America. 106, 21597-21601 (2009).
- Watt, I. N., Montgomery, M. G., Runswick, M. J., Leslie, A. G., Walker, J. E. Bioenergetic cost of making an adenosine triphosphate molecule in animal mitochondria. Proceedings of the National Academy of Sciences of the United States of America. 107, 16823-16827 (2010).
- Baker, L. A., Watt, I. N., Runswick, M. J., Walker, J. E., Rubinstein, J. L. Arrangement of subunits in intact mammalian mitochondrial ATP synthase determined by cryo-EM. Proceedings of the National Academy of Sciences of the United States of America. 109, 11675-11680 (2012).
- Allen, R. D., Schroeder, C. C., Fok, A. K. An investigation of mitochondrial inner membranes by rapid-freeze deep-etch techniques. The Journal of cell biology. 108, 2233-2240 (1989).
- Strauss, M., Hofhaus, G., Schroder, R. R., Kühlbrandt, W. Dimer ribbons of ATP synthase shape the inner mitochondrial membrane. The EMBO journal. 27, 1154-1160 (2008).
- Dudkina, N. V., Oostergetel, G. T., Lewejohann, D., Braun, H. P., Boekema, E. J. Row-like organization of ATP synthase in intact mitochondria determined by cryo-electron tomography. Biochimica et biophysica acta. 1797, 272-277 (2010).
- Davies, K. M., Anselmi, C., Wittig, I., Faraldo-Gomez, J. D., Kühlbrandt, W. Structure of the yeast F1Fo-ATP synthase dimer and its role in shaping the mitochondrial cristae. Proceedings of the National Academy of Sciences of the United States of America. 109, 13602-13607 (2012).
- Davies, K. M., et al. Macromolecular organization of ATP synthase and complex I in whole mitochondria. Proceedings of the National Academy of Sciences of the United States of America. 108, 14121-14126 (2011).
- Wurm, C. A., et al. Nanoscale distribution of mitochondrial import receptor Tom20 is adjusted to cellular conditions and exhibits an inner-cellular gradient. Proceedings of the National Academy of Sciences of the United States of America. 108, 13546-13551 (2011).
- Schermelleh, L., Heintzmann, R., Leonhardt, H. A guide to super-resolution fluorescence microscopy. The Journal of cell biology. 190, 165-175 (2010).
- Austin, J. R. 2nd, Staehelin, L. A. Three-dimensional architecture of grana and stroma thylakoids of higher plants as determined by electron tomography. Plant physiology. 155, 1601-1611 (2011).
- Ohta, K., et al. Beam deceleration for block-face scanning electron microscopy of embedded biological tissue. Micron. 43, 612-620 (2012).
- Buzhynskyy, N., Sens, P., Prima, V., Sturgis, J. N., Scheuring, S. Rows of ATP synthase dimers in native mitochondrial inner membranes. Biophysical journal. 93, 2870-2876 (2007).
- Larabell, C. A., Nugent, K. A. Imaging cellular architecture with X-rays. Current opinion in structural biology. 20, 623-631 (2010).
- Miao, J., Hodgson, K. O., Sayre, D. An approach to three-dimensional structures of biomolecules by using single-molecule diffraction images. Proceedings of the National Academy of Sciences of the United States of America. 98, 6641-6645 (2001).
- Maimon, T., Elad, N., Dahan, I., Medalia, O. The human nuclear pore complex as revealed by cryo-electron tomography. Structure. 20, 998-1006 (2012).
- Meyerson, J. R., et al. Determination of molecular structures of HIV envelope glycoproteins using cryo-electron tomography and automated sub-tomogram averaging. Journal of visualized experiments : JoVE. (58), (2011).
- Chen, S., et al. Electron cryotomography of bacterial cells. Journal of visualized experiments : JoVE. (39), (2010).
- Bharat, T. A., et al. Structural dissection of Ebola virus and its assembly determinants using cryo-electron tomography. Proceedings of the National Academy of Sciences of the United States of America. 109, 4275-4280 (2012).
- Bennett, A. E., et al. Ion-abrasion scanning electron microscopy reveals surface-connected tubular conduits in HIV-infected macrophages. PLoS pathogens. 5, e1000591(2009).
- Liu, J., et al. Cellular architecture of Treponema pallidum: novel flagellum, periplasmic cone, and cell envelope as revealed by cryo electron tomography. Journal of molecular biology. 403, 546-561 (2010).
- Liu, J., et al. Intact flagellar motor of Borrelia burgdorferi revealed by cryo-electron tomography: evidence for stator ring curvature and rotor/C-ring assembly flexion. Journal of bacteriology. 191, 5026-5036 (2009).
- Patla, I., et al. Dissecting the molecular architecture of integrin adhesion sites by cryo-electron tomography. Nature cell biology. 12, 909-915 (2010).
- Heuser, T., et al. Cryoelectron tomography reveals doublet-specific structures and unique interactions in the I1 dynein. Proceedings of the National Academy of Sciences of the United States of America. 109, E2067-E2076 (2012).
- Briggs, J. A. Structural biology in situ-the potential of subtomogram averaging. Current opinion in structural biology. 23, (2013).
- Cheng, D., Mitchell, D. R. G., Shieh, D. B., Braet, F. Current Microscopy Contributions to Advances in Science and Technology. Méndez-Vilas, A. 2, Formatex. (2012).
- Brust, D., et al. Cyclophilin D links programmed cell death and organismal aging in Podospora anserina. Aging cell. 9, 761-775 (2010).
- Gregg, C., Kyryakov, P., Titorenko, V. I. Purification of mitochondria from yeast cells. Journal of visualized experiments : JoVE. (30), (2009).
- Bhaskaran, S., et al. Breaking Caenorhabditis elegans the easy way using the Balch homogenizer: an old tool for a new application. Analytical biochemistry. 413, 123-132 (2011).
- Pavlov, P. F., Rudhe, C., Bhushan, S., Glaser, E. In vitro and in vivo protein import into plant mitochondria. Methods in molecular biology. 372, 297-314 (2007).
- Graham, J. M., et al. Ch. 3, Current protocols in cell biology. Bonifacino, J. S. , John Wiley and Sons. (2001).
- Zheng, S. Q., et al. UCSF tomography: an integrated software suite for real-time electron microscopic tomographic data collection, alignment, and reconstruction. Journal of structural biology. 157, 138-147 (2007).
- Mastronarde, D. N. Automated electron microscope tomography using robust prediction of specimen movements. Journal of structural biology. 152, 36-51 (2005).
- Kremer, J. R., Mastronarde, D. N., McIntosh, J. R. Computer visualization of three-dimensional image data using IMOD. Journal of structural biology. 116, 71-76 (1996).
- Pruggnaller, S., Mayr, M., Frangakis, A. S. A visualization and segmentation toolbox for electron microscopy. Journal of structural biology. 164, 161-165 (2008).
- Scheres, S. H., Chen, S. Prevention of overfitting in cryo-EM structure determination. Nature methods. 9, 853-854 (2012).
- Pettersen, E. F., et al. UCSF Chimera--a visualization system for exploratory research and analysis. Journal of computational chemistry. 25, 1605-1612 (2004).
- Dautant, A., Velours, J., Giraud, M. F. Crystal structure of the Mg.ADP-inhibited state of the yeast F1c10-ATP synthase. The Journal of biological chemistry. 285, 29502-29510 (2010).
- Carbajo, R. J., et al. Structure of the F1-binding domain of the stator of bovine F1Fo-ATPase and how it binds an alpha-subunit. Journal of molecular biology. 351, 824-838 (2005).
- Paumard, P., et al. The ATP synthase is involved in generating mitochondrial cristae morphology. The EMBO journal. 21, 221-230 (2002).
- Bharat, T. A., et al. Structure of the immature retroviral capsid at 8 A resolution by cryo-electron microscopy. Nature. 487, 385-389 (2012).
- Bartesaghi, A., Lecumberry, F., Sapiro, G., Subramaniam, S. Protein secondary structure determination by constrained single-particle cryo-electron tomography. Structure. 20, 2003-2013 (2012).
- Bai, X. C., Fernandez, I. S., McMullan, G., Scheres, S. H. Ribosome structures to near-atomic resolution from thirty thousand cryo-EM particles. eLife. 2, e00461(2013).
- Li, X., et al. Electron counting and beam-induced motion correction enable near-atomic-resolution single-particle cryo-EM. Nature methods. , (2013).
- Danev, R., Nagayama, K. Optimizing the phase shift and the cut-on periodicity of phase plates for TEM. Ultramicroscopy. 111, 1305-1315 (2011).
- Walter, A., et al. Practical aspects of Boersch phase contrast electron microscopy of biological specimens. Ultramicroscopy. 116, 62-72 (2012).
Перепечатки и разрешения
Запросить разрешение на использование текста или рисунков этого JoVE статьи
Запросить разрешениеСмотреть дополнительные статьи
This article has been published
Video Coming Soon
Авторские права © 2025 MyJoVE Corporation. Все права защищены