
Method Article
Visualizzazione di ATP sintetasi Dimeri nei mitocondri da Electron Cryo-tomografia
In questo articolo
Riepilogo
Vi presentiamo un protocollo di come raccogliere e elettrone processo di crio-tomografie di tutta mitocondri. La tecnica fornisce approfondimenti dettagliati sulla struttura, funzione, e l'organizzazione di grandi complessi proteici di membrana nelle membrane biologiche autoctone.
Abstract
Electron crio-tomografia è un potente strumento per la biologia strutturale, in grado di visualizzare la struttura tridimensionale di campioni biologici, come le cellule, organelli, vescicole di membrana, o virus al dettaglio molecolare. Per raggiungere questo obiettivo, il campione acquoso viene rapidamente vetrificato in etano liquido, che conserva in uno stato congelato-idratata close-to-native. Nel microscopio elettronico, serie inclinazione sono rilevati alla temperatura dell'azoto liquido, da cui tomografie 3D sono ricostruiti. Il rapporto segnale-rumore del volume tomografica è intrinsecamente bassa. Riconoscibile, caratteristiche ricorrenti sono arricchite da subtomogram media, con la quale i singoli subvolumes sono tagliati fuori, allineati e media per ridurre il rumore. In questo modo, le mappe 3D con una risoluzione di 2 nm o migliore può essere ottenuto. Una forma di strutture ad alta risoluzione disponibili al volume 3D allora produce modelli atomici di complessi di proteine nel loro ambiente nativo. Qui mostriamo come usiamo elettrone crio-tomograpHY per studiare in situ organizzazione di grandi complessi proteici di membrana in mitocondri. Troviamo che sintasi ATP sono organizzati in file di dimeri lungo apici molto curve della membrana interna creste, mentre io complesso è distribuito in modo casuale nelle regioni di membrana su entrambi i lati delle righe. Con subtomogram media abbiamo ottenuto una struttura mitocondriale ATP sintasi dimero all'interno della membrana creste.
Introduzione
I mitocondri sono i power-case del cellulare. Con la conversione di un gradiente protonico elettrochimico attraverso la membrana mitocondriale interna in energia legame chimico, la ATP sintetasi mitocondriale produce la maggior parte del ATP che guida i processi cellulari. Al fine di comprendere i meccanismi che stanno dietro la conversione di energia mitocondriale, abbiamo bisogno di determinare la struttura della ATP sintasi in situ, e per scoprire come è organizzato e distribuito nella membrana mitocondriale interna. Sebbene strutture ad alta risoluzione della maggior parte dei componenti mitocondriali ATP sintasi 1-3 e cartine bassa risoluzione dell'intero complesso 4 sono disponibili, è importante stabilire la struttura e conformazione dell'enzima di lavoro nella membrana. La distribuzione della ATP sintasi nella membrana mitocondriale interna è stato ampiamente ipotizzato essere casuale, ma uno dei primi 5 e trovare i nostri risultati iniziali 6 indicato che questo non è tche caso. Studi successivi del nostro gruppo e altri 7 hanno confermato che l'ATP sintasi è organizzato in lunghe file di dimeri lungo i crinali strettamente curve della membrana mitocondriale interna creste 8, mentre le pompe protoniche della catena di trasporto degli elettroni sembrano essere situato su entrambi i lati delle file 9. Questa disposizione ha importanti implicazioni per i meccanismi di conversione energetica mitocondriale.
La tecnica che abbiamo utilizzato per determinare tale disposizione è crio-elettroni tomografia (crio-ET). Cryo-ET è attualmente l'unico metodo che fornisce accurate tridimensionale (3D) I volumi delle cellule, compartimenti cellulari o organuli a risoluzione molecolare. Cryo-ET è particolarmente adatto per studiare i grandi complessi di membrane biologiche, perché le membrane appaiono con un buon contrasto e sono facili da tracciare in volumi tomografiche 3D.
Altri metodi per studiare la struttura 3D delle cellule o organelles non forniscono dettagli molecolare. Super-risoluzione microscopia ottica 10, 11 è eccellente a rivelare la posizione o distanze tra etichette luminescenti collegati a proteine di interesse con una precisione di decine di nm, ma non rivela la struttura della proteina stessa, anche a bassa risoluzione . Microscopia elettronica a trasmissione di sezioni seriali di 12 o di imaging block-face da microscopia elettronica a scansione 13 di campioni biologici di plastica-embedded offrono viste a bassa risoluzione di volumi cellulari, ma allo stesso modo non rivelano dettaglio molecolare. Microscopia a forza atomica 14 può in linea di principio di fornire risoluzione molecolare o atomica, ma solo alla superficie degli oggetti su un atomicamente piatta sostegno, solido. Infine, tomografia a raggi X 15 o dispersione di impulsi di raggi X intensi da laser ad elettroni liberi 16 è improbabile per rivelare la struttura di grandi e complessi, oggetti aperiodici come le cellule intere o organelli a mRisoluzione olecular nel prossimo futuro. Quindi allo stato attuale, non vi è alcuna alternativa al crio-ET per studi sulla struttura 3D di cellule o organuli a risoluzione nanometrica.
Cryo-ET è il metodo di scelta per l'esame della struttura e la conformazione dei gruppi di proteine associate alla membrana, tra cui il complesso del poro nucleare 17, il picco dell'influenza complessi 18, e flagelli proteine motrici 22, 23 ma anche l'organizzazione di intere cellule batteriche 19 e l'entrata di virus patogeni come l'HIV nelle cellule 20-23. Cryo-ET ha un valore inestimabile per la visualizzazione di proteine filamentose e le loro interazioni nella cellula, compresi i filamenti di actina 24 o axonemes 25. La risoluzione può essere migliorata a 2 nm o meglio subtomogram media di 26, in base al quale subvolumes di ripetute, lineamenti regolari sono tagliati fuori da un volume tomografica e media per singola particella Image Protecniche di lavorazione.
Cryo-ET comporta l'acquisizione di una serie di immagini di proiezione di un esemplare sottile (<250nm) prese a diversi angoli di inclinazione in un microscopio elettronico a trasmissione (TEM). Il campione deve essere sottile in modo che gli elettroni, che interagiscono fortemente con la materia, sono sparsi in non più di una volta. Scattering multiplo rende le immagini risultanti difficili da interpretare e riduce il contrasto. Immagini dell'area campione selezionato sono allineati l'uno rispetto all'altro e proiettati in uno spazio 3D da un programma per computer adatto, generando un volume 3D del campione. L'allineamento delle immagini è aiutata da marker fiduciali oro, che si mescolano con il campione prima del congelamento. Marker fiduciali Idealmente 10 o più equamente distribuiti dovrebbero essere presenti in ogni immagine per ottenere un buon allineamento.
Per osservare dettaglio molecolare, i campioni sono tuffo congelati in etano liquido, che conserva il loro stato idratato nativo. Congelamento di liquidoetano è così veloce (~ 10 5 ° C / sec) 27 che l'acqua non cristallizza, ma rimane in uno stato simile al vetro vetrificato. Cristalli di ghiaccio danneggia formazione strutture biologiche sensibili. Come campioni biologici soffrono di danni da radiazioni, c'è un limite al numero totale di eventi di scattering il campione può tollerare. Le immagini vengono quindi acquisiti in modalità a basso dosaggio: Una zona di interesse è identificata a basso ingrandimento (1,500X) con una dose inferiore a 1 elettrone e - / nm 2 (modalità di ricerca). L'immagine viene poi messo a fuoco con un ingrandimento maggiore, l'area di interesse (modalità di messa a fuoco). Solo quando l'immagine viene acquisita, l'area di interesse viene irradiata con una dose elevata di elettroni (modo di esposizione).
Qui, vi presentiamo una panoramica di come raccogliere e di elettroni processo di crio-tomografie, utilizzando ATP sintasi dimeri nella membrana mitocondriale interna come un esempio. Il seguente protocollo descrive come preparare i mitocondri per crio-ET, come impostaree raccogliere una serie di inclinazione con una dose specifica di elettroni totale, e come elaborare la serie di inclinazione per ottenere un volume 3D della zona di interesse. Una panoramica della procedura è illustrata nella figura 1.
Protocollo
1 Preparazione di mitocondri da cellule o tessuti mediante centrifugazione differenziale
Questa sezione descrive una procedura generale per l'isolamento dei mitocondri intatti da vari organismi eucarioti. I componenti del tampone e velocità di centrifugazione precise devono essere ottimizzate per ogni tessuto / specie studiate.
- Rompere le cellule in tampone isotonico (ad esempio, saccarosio 250 mm, 10 HEPES mM pH 7.4), utilizzando un mulino vetro tallone (micelio fungino) 28, digestione enzimatica della parete cellulare (Saccharomyces cerevisiae) 29, un omogeneizzatore-cuscinetto a sfere (singolo -cell eucarioti / cultura cellule / nematodi) 30, o un frullatore (tessuti animali o vegetali) 31.
- Rimuovere detriti cellulari mediante filtrazione attraverso mussola seguita da centrifugazione a bassa velocità (2000 xg, a 4 ° C, 10 min).
- Raccogliere il surnatante e pellet mitocondri mediante centrifugazione ad alta velocità (9.000 xg, a 4 ° C, 10 min).
- Se necessario, utilizzare un gradiente di densità passo isotonica per la purificazione ulteriore della frazione mitocondriale 32.
2 Preparazione di mitocondri per Electron Cryo-tomografia
La sezione seguente descrive come ottenere i campioni congelati-idratata per crio-ET. NOTA: Il metodo prevede l'uso di azoto liquido estremamente freddo ed etano, che può causare gravi ustioni. Occhiali di sicurezza e guanti di crio-protezione devono essere indossati. Etano liquido, che è anche infiammabile, deve essere gestito in una cappa aspirante.
- Risospendere mitocondri pellet in trealosio 250 mM, 10 mM tampone HEPES a pH 7,4 ad una concentrazione di circa 5 mg / ml di proteine totali.
- Glow-scarico griglie EM carbonio bucata, laterali carbonio alto, in un dispositivo a depressione secondo le istruzioni del produttore.
- Liquefare pochi millilitri di etano dirigendo un flusso di gas etano sul lato interno di un fresco azoto liquidocontenitore di alluminio ed.
- Mescolare oro proteina A-coniugato sospensione fiduciale 1: 1 con sospensione mitocondriale e applicare immediatamente 3 microlitri di una griglia EM bagliore-scarico in pinzetta.
- Mettere le pinzette in un dispositivo di vetrificazione, ad esempio, una ghigliottina fatta in casa. Asciugare il liquido in eccesso con un cuneo di carta da filtro (~ 5 secondi o fino a quando si ferma liquidi diffusione) e immergersi immediatamente la griglia in etano liquido rilasciando il grilletto.
- Trasferire la griglia da etano liquido in azoto liquido. Durante il trasferimento, rimuovere l'eccesso di etano dalla griglia con carta da filtro. Posizionare la punta di un pezzo a forma di cuneo di carta filtro in etano liquido. Come la etano liquido sale, trascinare delicatamente la griglia la carta da filtro ma tenerlo sotto la parte frontale del liquido. Etano liquido viene rimosso dalla rete per azione capillare e una volta che tutto etano liquido è stato rimosso, trasferire immediatamente la griglia in azoto liquido.
- Posizionare la griglia vetrificata in una scatola di immagazzinaggio griglia, unand conservare in azoto liquido per un uso successivo come appropriato.
3 Registrazione di tomografica Tilt Series
La sezione seguente descrive come impostare e raccogliere una serie di inclinazione tomografica di un mitocondrio con un microscopio elettronico Polara dotato di un filtro di energia post-colonna e camera CCD. Protocolli simili sono utilizzati con tutti i microscopi elettronici crio-dotati di CCD o telecamere dirette rivelatore di elettroni.
- Allineare il microscopio
- Inserire provino ad esempio, grafite e le isole d'oro su pellicola di carbonio bucata.
- Selezionare la modalità di ricerca nel sistema di basse dosi del microscopio.
- Portare campione altezza eucentrica. Questo è il punto di minimo movimento xy quando si inclina il supporto del campione. Centrare un punto di interesse a 0 ° di inclinazione, inclinare la fase a 20 ° e ricentrare il punto alterando lo z-altezza. Torna a 0 ° e ripetere fino a spostamento laterale è ridotto al minimo.
- Selezionaremodalità di esposizione. Scegli ingrandimento desiderato per la raccolta di un tomogramma (ad esempio, 25,000X sul rivelatore ≈ 0,6 nm CAMPIONE dimensione dei pixel).
- Scegli una piccola apertura del condensatore (50-70 mm) e selezionare la dimensione dello spot e l'intensità del fascio in modo che il fascio è solo più largo del dispositivo di imaging e dà una lettura di 60 pixel e - / pixel (CCD) o 14 e - / pixel / sec (rivelatore di elettroni diretto, modalità conteggio).
- Apertura del condensatore Center.
- Trova gaussiana fuoco ad esempio, il punto di contrasto minimo. Ripristino lettura microscopio defocus e corretti punti di articolazione e il centro di rotazione secondo le istruzioni del produttore.
- Quadrante in defocus desiderata per la registrazione tomogram. NOTA: Alta defocus (8 micron) aumenta il contrasto, ma riduce la risoluzione, considerando che la bassa defocus (2-4 micron) aumenta la risoluzione a scapito del contrasto.
- Su un buco vuoto, generare un nuovo guadagno di riferimento e allineare filtro di energia secondo il fornitore diistruzioni.
- Allineare ricerca e di esposizione modi. Nel modo di esposizione, centrare un punto di interesse e passare alla modalità di ricerca. Selezionare ingrandimento di 1,500X (0,033 micron / pixel di campione sul rivelatore) e la sfocatura di 100 micron (per aumentare il contrasto). Punto di interesse schiena Portare a centrare utilizzando bobine di spostamento immagine.
- Nel modo di ricerca, regolare la dimensione dello spot e l'intensità del fascio in modo che il fascio è solo più largo del dispositivo di imaging e dà una lettura pixel di ~ 20 e - / pixel (CCD) o ~ 8 e - / pixel / sec (rivelatore di elettroni diretto, modalità conteggio).
- Trovare un buon Area campione
- Inserire la griglia con mitocondri congelati-idratata nel microscopio elettronico a temperatura dell'azoto liquido (fare riferimento alle istruzioni del produttore EM).
- Nel modo di ricerca, cercare la griglia per le zone di spessore del ghiaccio e del campione di qualità appropriato. Prendete un 6 sec ricerca di immagini di settori promettenti per determinare l'idoneità per la raccolta tomogram. Both la membrana mitocondriale interna ed esterna deve essere visibile in questo ingrandimento.
- Registrazione di un tomografica Tilt Series
- Una volta che una buona zona esemplare si trova, inclinare la fase ± 60 ° per determinare la portata massima inclinazione che è disponibile senza alcun ostacolo di esposizione o di messa a fuoco da bar griglia o grumi di ghiaccio.
- Su un buco pieno di ghiaccio nelle vicinanze di aspetto simile, passare alla modalità di esposizione e regolare il tempo di intensità del fascio o l'acquisizione delle immagini in modo che ogni immagine registrata ha una dose di 30-50 elettrone e - / pixel per CCD o 6-8 e - / pixel / s rivelatori di elettroni diretti, modalità di conteggio.
- Calcolare il rapporto di distribuzione della dose (I 0 / I 60) dividendo il numero medio di elettroni per un'immagine 1 sec acquisita a 0 ° con quella di un'immagine di 60 °. Questo rapporto descrive l'aumento del tempo di esposizione necessario per mantenere costante il numero di elettroni per ogni immagine con l'aumentare dell'angolo di inclinazione (tempo di esposizione = 1 / cos (α) n dove (I 0 / I 60) = 2 n). Il rapporto serve anche come una buona indicazione dello spessore del ghiaccio. Buone tomogrammi di mitocondri sono generalmente registrati con un I 0 / I 60 = 2,3-2,6.
- Su un buco vuoto, acquisire un'immagine 1 sec in modalità di esposizione e notare il conteggio elettronico per Å 2. Tenendo conto del rapporto di distribuzione della dose, calcolare il numero totale di immagini che possono essere registrate per una dose totale di elettroni specifico (ad esempio, <40 e - / a 2 per la determinazione della struttura & ~ 160 e - / A 2 per morfologia).
- Determinare l'intervallo di inclinazione appropriata per la raccolta tomografia dividendo il range di inclinazione totale (ad esempio, 120 ° per ± 60 °) per il numero totale di immagini calcolati in 3.3.4.
- Impostare e registrare un tomogramma con i parametri determinati in precedenza utilizzando il software appropriato raccolta automatica dei dati 33, 34. Serie Tilt sono di solito avviati a ± 20 ° e passare attraverso 0 ° prima di raggiungere elevate inclinazioni al fine di massimizzare il contenuto informativo delle immagini a bassa inclinazione, che viene distrutto aumentando la dose di elettroni.
4 Creazione e segmentazione di volumi tomografiche
Questa sezione descrive come volumi tomografiche di mitocondri sono generati da serie tilt e come sono presenti per la visualizzazione generale dei volumi.
- Salvare la serie tomografico inclinazione a una directory appropriata. Generare una serie di immagini e convertire in un formato di file appropriato con il software open-source, come ad esempio dm2mrc o tif2mrc (pacchetto IMOD), che convertono .dm3, .dm4 o tif file stack MRC. pile MRC sono necessari per la ricostruzione tomografica con IMOD 35. Altri pacchetti richiedono diversi formati.
- Allineare le immagini e generare un tomogramma seguendo la procedura descritta nel tutorial IMOD ( http://bio3d.colorado.edu/imod/doc/etomoTutorial.html ).
- Migliora il contrasto del tomogramma utilizzando il filtro anisotropico diffusione non lineare distribuito con MDI. Questo filtro funziona bene per le membrane e particelle associate alla membrana, come ATP sintasi.
- Per la visualizzazione, segmento manualmente il tomogramma utilizzando programmi disponibili in commercio per esempio, Amira. Assegnare voxel corrispondenti alla membrana interna o esterna e generare una superficie. Utilizzando l'opzione clicker nel plugin EM-pacchetto per AMIRA 36 contrassegnare la posizione delle particelle ATP sintetasi.
5. Subtomogram della media dell'ATP sintasi dimeri e montaggio di strutture X-Ray
La sezione seguente descrive come medie subtomogram di ATP sintasi dimeri possono essere ottenuti.
- Utilizzando le particelle contrassegnate come input e un pacchetto software appropriato, come il 'ParteStima icolo per il programma Electron Positron ', calcolare una media subtomogram.
- Per un preventivo risoluzione, confrontare due medie subtomogram determinati autonomamente da Fourier shell correlazione 37.
- Se disponibili, dock noto strutture a raggi X nel medio subtomogram di corpo rigido raccordo, manualmente o utilizzando le routine di docking automatico sequenziale come quelli nel programma Chimera 38.
Risultati
Electron crio-tomografie di mitocondri rivelano chiaramente la morfologia 3D del organello (Figura 2). Segmentazione manuale delle membrane in un volume tomografica illustra la struttura delle creste in un mitocondrio. Con l'imaging mitocondri provenienti da diversi ceppi di lievito eliminazione diretta che mancano alcuni componenti proteiche, l'effetto di queste proteine su creste morfologia può venire valutato. Figura 3 mostra un mitocondrio da un ceppo di lievito privo di ATP sintetasi subunità e. Questo componente è necessario del complesso ATP sintasi per la dimerizzazione della ATP sintasi mitocondriale. I mitocondri da questo ceppo manca la normale lamellare creste dei mitocondri di tipo selvatico (Figura 2) e invece contiene una serie di compartimenti membrana interna. Questi comparti sono o privi di creste o contengono piccole invaginazioni di membrana a forma di palloncino (Figura 3).
In tomograms con un buon contrasto, grandi complessi di proteine mitocondriali, in questo caso ATP sintasi dimeri, sono facilmente visibili (figura 4; Movie 1). Le strutture dei complessi possono essere determinati a 2-3 nm risoluzione da subtomogram media (figura 5; Movie 2). I volumi medi possono essere riposti nella tomografia per valutare l'organizzazione dei singoli complessi rispetto all'altro e ad altri complessi proteici nella membrana (Figura 6; film 3).

Figura 1 Flow-chart che mostra le fasi di elettroni crio-tomografia. Clicca qui per vedere l'immagine ingrandita.
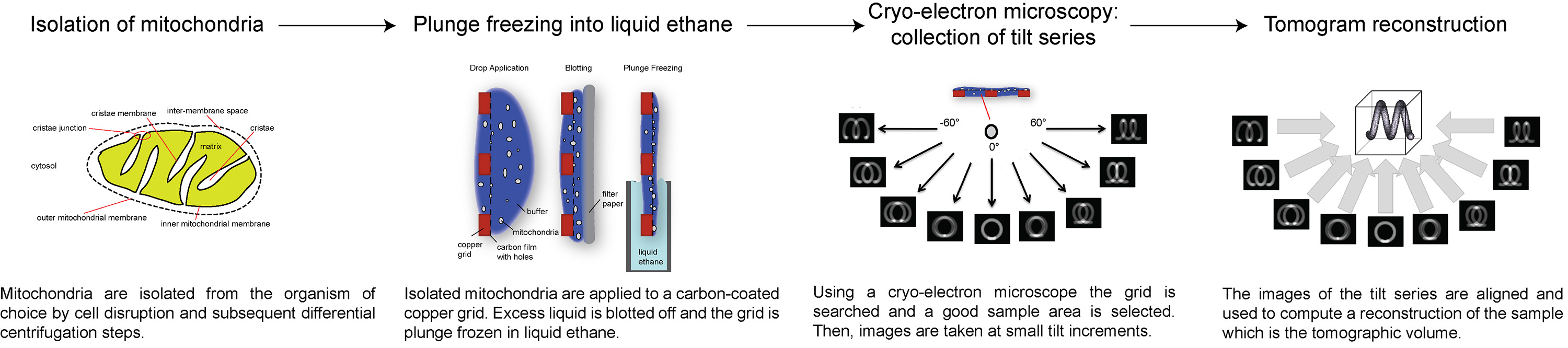{kind=link}

Figura 2 Morfologia di un mitocondrio di tipo selvatico S. Cerevisiae. Fetta centrale attraverso un volume tomografica di un tipo selvatico S. mitocondrio cerevisiae (a sinistra) e il corrispondente volume-superficie di rendering (a destra). Il volume segmentato della membrana esterna è visualizzata in grigio e volumi del limite interno e creste membrane in azzurro. Adattato da Davies et al 8. Clicca qui per vedere l'immagine ingrandita.
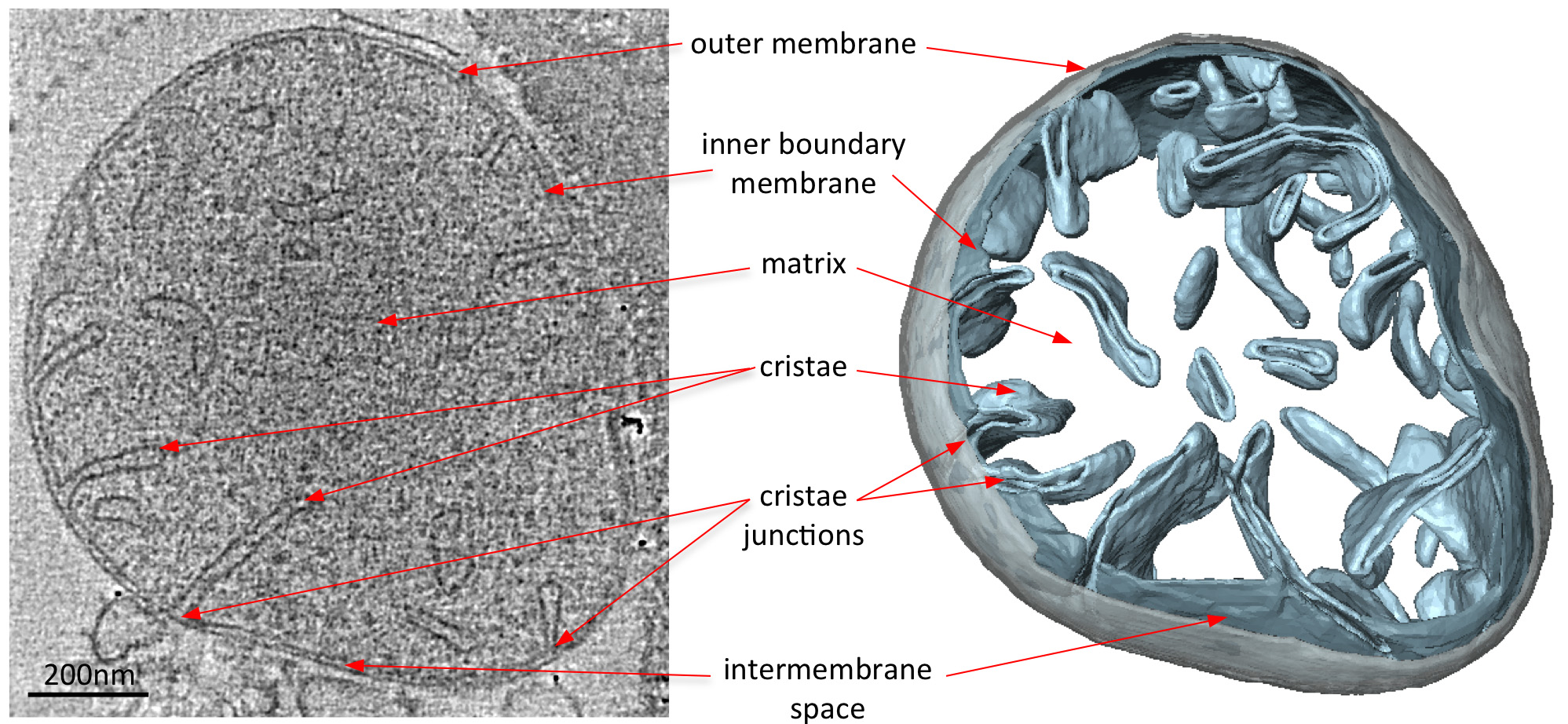{kind=link}

Figura 3 mitocondrio da una S. Cerevisiae ceppo manca una subunità necessarie perATP sintasi dimerizzazione. Tagliate attraverso il volume tomografica (a sinistra), che accompagna il volume-superficie di rendering (a destra) di un mitocondrio da una S. Cerevisiae ceppo manca la proteina subunità e necessario per l'ATP sintasi dimerizzazione. Quando confrontato con la figura 2, il mitocondrio dal ceppo mutante manca la normale lamellare creste dei mitocondri tipo selvatico. Invece, il mitocondrio ha molti compartimenti di membrana interna sia con senza creste o creste a forma di palloncino. Così elettrone crio-tomografia evidenzia alterazioni della morfologia della membrana a causa di delezioni geniche. Adattato da Davies et al 8. Clicca qui per vedere l'immagine ingrandita.
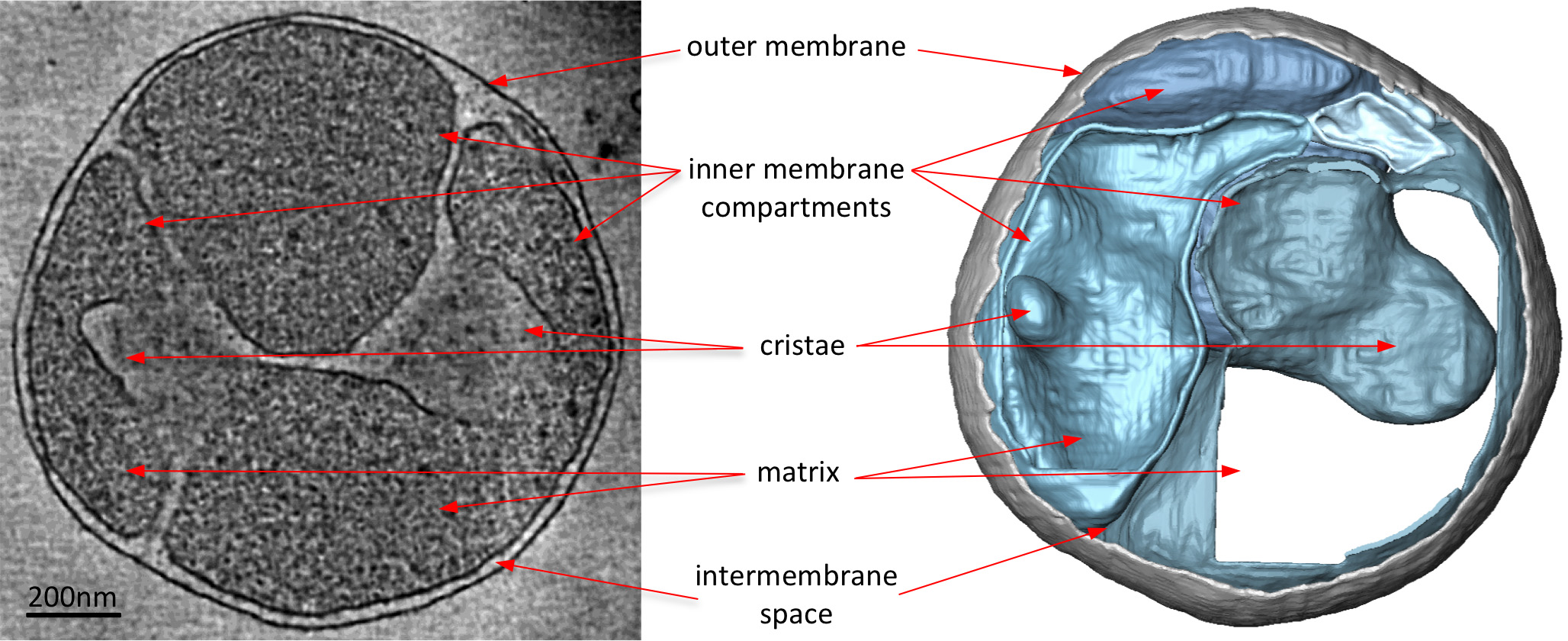{kind=link}

Figure 4 mitocondrio dal fungo P. anserina. Affettare attraverso il volume tomografica (a sinistra) e di volume-superficie di rendering di accompagnamento (a destra) di un mitocondrio dal fungo filamentoso P. anserina. In questo tomogramma, filari di 10 particelle nm (frecce gialle) si trovano sopra le creste molto curve di membrana in membrana creste interna (vedi filmato 1). Queste particelle sono state identificate come dimeri ATP sintasi di subtomogram media. Da Davies et al 9. Clicca qui per vedere l'immagine ingrandita.
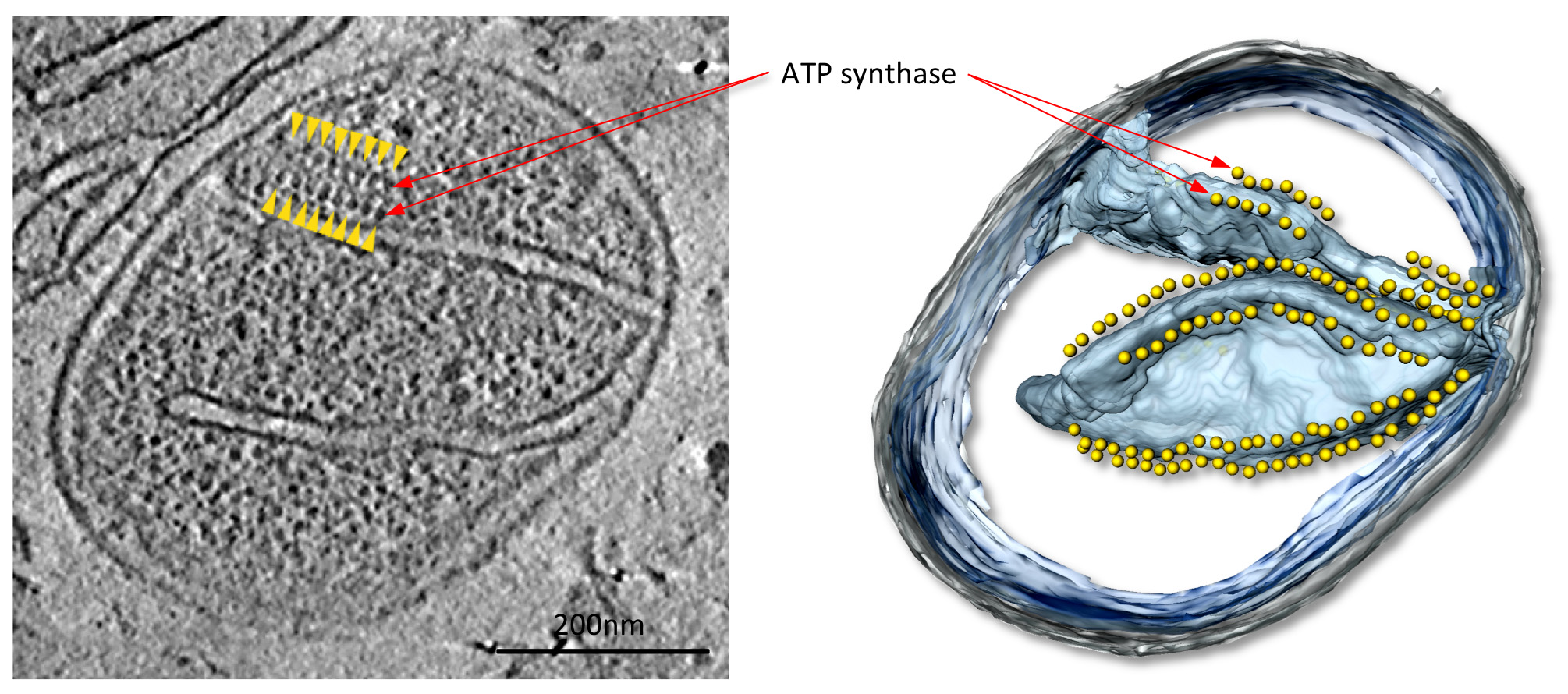{kind=link}

Figura 5 Struttura e organizzazione della ATP sintasi mitocondriale. Laterale e vista dall'alto che mostra la densità elettronica di un dimero ATP sintasi da S. c erevisiae come determinato dal subtomogram media con modelli atomici a muro (a sinistra). Mitocondriale vescicola membrana interna che mostra l'organizzazione di ATP sintasi dimeri in righe (a destra). Il dato è stato generato posizionando la media subtomogram della sintasi dimero ATP nel volume segmentato della vescicola membrana, utilizzando le coordinate calcolate durante media. Adattato da Davies et al 8. Modelli atomici: F 1 / rotore-ring [PDB: 2WPD] 39 (blu e viola); oligomicina sensitive conferendo proteina-OSCP [PDB: 2BO5] 40 (verde); frammento gambo periferico [PDB: 2CLY] 1 con residui N-terminali da [PDB: 2WSS] 2 (giallo e rosso) (vedi Movie 2). Clicca qui per vedere l'immagine ingrandita.
{kind=link}
_upload / 51228 / 51228fig6highres.jpg "width =" 500 "/>
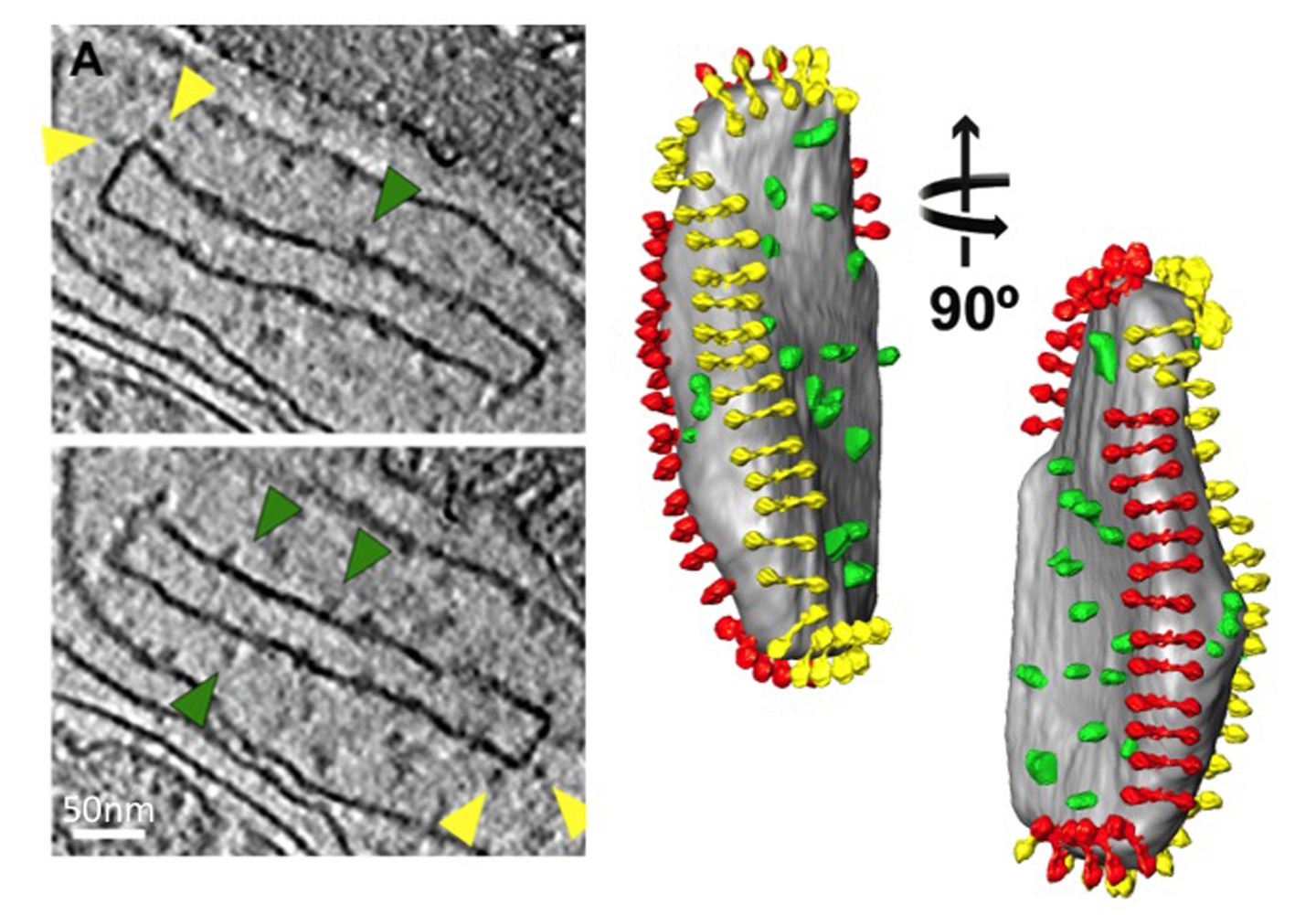
Figura 6 Isolato Crista vescicola da un P. anserina mitocondrio. Fette da un volume tomografica (a sinistra) e di volume-superficie di rendering di accompagnamento (a destra) di una vescicola crista da P. anserina. Densità di proteine che sporgono dalla membrana sono chiaramente visibili. Le densità indicati da frecce gialle sono dimeri ATP sintetasi, come individuate dalla subtomogram media. Frecce verdi indicano densità individuati mediante l'etichettatura degli anticorpi come NADH deidrogenasi (complesso 1, per i dettagli vedere 9). Segmentazione delle densità di proteine rivela la loro organizzazione in creste, con i dimeri ATP sintasi (rosso e giallo) che formano righe lungo i crinali creste molto curve e complessi NADH deidrogenasi (verdi) nelle regioni di membrana su entrambi i lati delle righe (vedi Movie 3). Da Davies et al 9.28 / 51228fig7highres.jpg "target =" _blank "> Clicca qui per vedere l'immagine ingrandita.
Movie 1 Electron crio-tomogramma di un P. anserina mitocondrio. Il filmato mostra le fette successive attraverso un volume tomografica presa di un mitocondrio dal fungo filamentoso P. anserina. Righe di ATP sintasi sono indicati da frecce gialle. Il volume segmentato superficie rendering mostra la posizione di ATP sintasi (sfere gialle) in relazione alla struttura creste 3D. Membrana esterna, grigio; membrana limite interno, blu trasparente; creste membrane, blu opaco. Da Davies et al 9. Vedere anche la Figura 4. Clicca qui per vedere il video.
Movie 2 media Subtomogram della sintasi mitocondriale dimero ATP da S. Cerevisiae con modelli atomici a muro. La media è statacalcolata da 121 subvolumes. La densità viene visualizzata a tre livelli di contorno: 1s - maglia, 2s - grigio chiaro e 3s - grigio scuro. Modelli atomici sono stati montati nella densità utilizzando la routine in forma sequenziale in Chimera. Modelli atomici: F 1 / rotore-ring [PDB: 2WPD] 39 (blu e viola); oligomicina sensitive conferendo proteina-OSCP [PDB: 2BO5] 40 (verde); frammento gambo periferico [PDB: 2CLY] 1 con residui N-terminali da [PDB: 2WSS]. 2 (giallo e rosso) Clicca qui per vedere il video.
Adattato da Davies et al 8. Vedere anche Figura 5.
Movie 3 Isolati creste vescicola da un P. anserina mitocondrio. fette successive attraverso il volume tomografica sono presenti, seguito dal volume-superficie resi segmentato. Densità di proteine sporgenti dal membrane sono chiaramente visibili. Densità rossi e gialli sono dimeri ATP sintetasi. Densità verdi sono NADH deidrogenasi (complesso I), come determinato mediante l'etichettatura degli anticorpi. Da Davies et al 9. Vedere anche Figura 6. Clicca qui per vedere il video.
Discussione
Il protocollo qui presentato fornisce un'introduzione alla crio-ET e subtomogram media dei mitocondri, ma essenzialmente la stessa procedura può essere applicata a qualsiasi altro compartimento cellulare o membrana. Per ottenere i migliori dati possibili, fasi critiche nel corso della procedura sono la preparazione del campione, la strategia di acquisizione processo di congelamento tuffo e dati. Qualità del campione, che è fondamentale per il successo, dipende da un protocollo di congelamento ottimizzato per garantire lo spessore del ghiaccio adatto, che è di fondamentale importanza per il buon contrasto dell'immagine. La strategia ottimale di acquisizione dati dipende dallo strumento ed il campione. Parametri da ottimizzare includono dose di elettroni per immagine, sistema di inclinazione e sfocatura. Acquisire una buona tomogramma di un buon campione rende ogni ulteriore trasformazione passaggi più facili e garantisce un risultato finale soddisfacente.
Cryo-ET combinato con subtomogram media e montaggio di modello atomico fornisce dettagli su come complessi proteici sono disposti in thambiente cellulare nativo EIR. La tecnica è ideale per indagare la struttura di grandi complessi proteici di membrana, come supercomplessi catena respiratoria (1.7 MDA), dimeri ATP sintasi (2x500 kDa), o il complesso del poro nucleare (~ 120 MDA) 8, 9, 17. L'organizzazione di ATP dimeri sintasi in righe non può essere osservato con tecniche ad alta risoluzione come la cristallografia a raggi X, NMR o singola particella crio-EM, perché le righe dimero sono interrotti per estrazione detergente, che è un passo necessario per l'isolamento e purificazione di complessi proteici di membrana.
La disposizione della ATP sintasi in file di dimeri lungo crinali creste è un principio organizzativo universale dei mitocondri in tutte le specie. I complessi protone-pompaggio della catena di trasporto degli elettroni, in particolare complesso I (NADH deidrogenasi), sono situati nelle zone di membrana lati delle righe 8,9. Questa organizzazione della catena respiratoria ha un impatto profondo sulla bioenergetica mitocondriale. Se le righe dimero non possono formare a causa dell'assenza di subunità proteiche specifica-dimero, le cellule presentano tempi di generazione più lunghi e ridotta idoneità cellulare, come osservato nel mutante di lievito mostrato in Figura 3 41. Con il crescente interesse per le malattie mitocondriali, una dettagliata comprensione delle basi molecolari che governano ultrastruttura e funzione mitocondriale è di fondamentale importanza. Electron crio-tomografia fornisce un collegamento tra struttura proteica determinata mediante metodi ad alta risoluzione, e la distribuzione e la disposizione di queste proteine nella membrana sulla scala dei nanometri. Questo rende crio-ET uno strumento essenziale per comprendere la struttura e la funzione mitocondriale in salute e malattia.
Ulteriori sviluppi tecnici e miglioramenti in crio-ET includono strategie di proteina-etichettatura per identificare la posizione di Protein subunità in complessi macromolecolari o la posizione delle proteine distinte più piccole o meno (<0,5 MDA) nelle cellule. Inoltre, i metodi di lavorazione EM ibridi, che combinano subtomogram media con l'analisi singola particella o ricostruzione elicoidale hanno recentemente determinato strutture proteiche a ~ 8 A 42, 43. Questi metodi di lavorazione sono attualmente limitati alle proteine purificate, che sono ben separati in ghiaccio o si formano gruppi elicoidali e non sono attualmente applicabili agli ambienti cellulari affollati come i mitocondri e le membrane creste. Per la raccolta dei dati, nuovi strumenti informatici e hardware sono in fase di sviluppo per l'acquisizione tomogram automatizzato, aumentare il contrasto dell'immagine e ridurre la dose totale richiesta di elettroni. L'unica limitazione fondamentale in crio-ET è danni da radiazioni al campione dal fascio di elettroni. Ciò significa che solo dosi molto basse di elettroni possono essere usati per registrare ogni immagine di una serie tomografica inclinazione, con conseguente scarsa segnale-trapporto che in ultima analisi, limita la risoluzione ottenibile o-rumore. I nuovi rivelatori di elettroni diretto, uscito meno di un anno fa, stanno rivoluzionando il campo di singolo-particella crio-EM 44, 45. Questi nuovi rivelatori forniscono un maggiore contrasto e una migliore risoluzione a dosi più basse di elettroni. Per elettrone crio-tomografia, questo significa che serie tilt con dimensioni di passo più piccole o addirittura tomogrammi biassiali possono essere raccolti senza preoccupazioni per i danni da radiazioni eccessive (una sola immagine CCD è equivalente a dosaggio elettronico a 5 immagini dirette rivelatore di elettroni). Le grandi quantità di dati prodotti da questi rilevatori di creare le proprie sfide nella gestione dei dati, trasformazione e stoccaggio, che dovranno essere superate.
Inoltre, le piastre di fase, che lavorano su principi simili a quelli utilizzati di routine in microscopia ottica per aumentare il contrasto di fase, sono attualmente in fase di sviluppo per la microscopia elettronica a trasmissione 46, 47. Ciò dovrebbe permettere tomogrammi da rilevare più vicino al fuoco e quindi a maggiore risoluzione, mentre allo stesso tempo conservando bassa risoluzione caratteristiche necessarie per l'allineamento e l'interpretazione dei volumi tomografiche. Presi insieme, questi progressi tecnologici espandere notevolmente la gamma di questioni biologiche che possono essere affrontati con crio-ET.
Divulgazioni
Gli autori dichiarano di non avere interessi finanziari in competizione.
Riconoscimenti
Questo lavoro è stato sostenuto dalla Max Planck Society (KMD, BD, AWM, la tubercolosi, TBB, DJM, e WK), il Cluster di Eccellenza Francoforte "complessi macromolecolari", finanziato dalla Deutsche Forschungsgemeinschaft (WK) e un post-dottorato EMBO a lungo termine Fellowship (VAMG).
Materiali
| Name | Company | Catalog Number | Comments |
| Sucrose | Roth | 4621.2 | |
| Trehalose | Sigma-Aldrich | T9449 | |
| HEPES | Sigma-Aldrich | H3375 | |
| Zymolase | biomol | Z1000.1 | |
| Density gradient medium | Sigma-Aldrich | D1556 or P1644 | |
| Protein A gold | Aurion | 810.111 | 10 nm |
| Ethane | Air Liquide | P0502S02R0A001 | |
| Nitrogen | Air Liquide | I4150RG | |
| Consumables | |||
| Filter paper | Whatman, GE Healthcare | 1004 090 | Nr 4 |
| Tweezers | Dumont | T506 | Non-magnetic, pattern #5 |
| EM combined test grid | Plano GmbH | S142 | grapite and gold islands on holey carbon film |
| Holey carbon grids | Quantifoil Micro Tools GmbH or C-flat | R2/2 300 Cu mesh | |
| Cryo grid boxes | Plano GmbH | 160-42 | Alternative home-made metal boxes |
| Potter | Sigma-Aldrich | P7984 | |
| Blender | Waring | 8011S | |
| Glass bead mill - bead beater | BioSpec Products | 1107900-105 | Including 0.5 mm beads |
| Ball-bearing (Balch) homogeniser | isobiotec | H8 | |
| Centrifuge | Thermo Scientific | 46915 | Sorvall RC6+ |
| Glow-discharge apparatus | Bal-Tec A.G. | CTA 010 | |
| Grid vitrification device | FEI, Gatan or Leica | Alternative home-made plunger | |
| Transmission electron microscope | FEI or Jeol | 300 keV | |
| Direct electron detector | Gatan | K2 summit | 4k x 4k pixels, counted mode |
| Charge coupled device (CCD) | Gatan | US4000 | 4k x 4k pixels (alternative to Direct electron detector) |
| Energy filter | Gatan or Jeol | ||
| Software | |||
| Image acquisition software | Gatan | ||
| Tomogram acquisition software | Gatan, FEI, USCF, or UC-Boulder | 4 alternatives | |
| Tomogram processing software | FEI, UCSF or UC-Boulder | 3 alternatives | |
| Subtomogram averaging software | UC-Boulder, or University of Basel | 2 alternatives | |
| Tomogram visualization software | UC-Boulder, FEI, or University of Utah | 3 alternatives | |
| Visualization software plugin | Goethe university | http://www.biophys.uni-frankfurt.de/frangakis/ | |
| Structure visualization software | UCSF | http://www.cgl.ucsf.edu/chimera/ | |
| Numerical calculation software | MathWorks | http://www.mathworks.de/academia/ | |
Riferimenti
- Dickson, V. K., Silvester, J. A., Fearnley, I. M., Leslie, A. G., Walker, J. E. On the structure of the stator of the mitochondrial ATP synthase. The EMBO journal. 25, 2911-2918 (2006).
- Rees, D. M., Leslie, A. G., Walker, J. E. The structure of the membrane extrinsic region of bovine ATP synthase. Proceedings of the National Academy of Sciences of the United States of America. 106, 21597-21601 (2009).
- Watt, I. N., Montgomery, M. G., Runswick, M. J., Leslie, A. G., Walker, J. E. Bioenergetic cost of making an adenosine triphosphate molecule in animal mitochondria. Proceedings of the National Academy of Sciences of the United States of America. 107, 16823-16827 (2010).
- Baker, L. A., Watt, I. N., Runswick, M. J., Walker, J. E., Rubinstein, J. L. Arrangement of subunits in intact mammalian mitochondrial ATP synthase determined by cryo-EM. Proceedings of the National Academy of Sciences of the United States of America. 109, 11675-11680 (2012).
- Allen, R. D., Schroeder, C. C., Fok, A. K. An investigation of mitochondrial inner membranes by rapid-freeze deep-etch techniques. The Journal of cell biology. 108, 2233-2240 (1989).
- Strauss, M., Hofhaus, G., Schroder, R. R., Kühlbrandt, W. Dimer ribbons of ATP synthase shape the inner mitochondrial membrane. The EMBO journal. 27, 1154-1160 (2008).
- Dudkina, N. V., Oostergetel, G. T., Lewejohann, D., Braun, H. P., Boekema, E. J. Row-like organization of ATP synthase in intact mitochondria determined by cryo-electron tomography. Biochimica et biophysica acta. 1797, 272-277 (2010).
- Davies, K. M., Anselmi, C., Wittig, I., Faraldo-Gomez, J. D., Kühlbrandt, W. Structure of the yeast F1Fo-ATP synthase dimer and its role in shaping the mitochondrial cristae. Proceedings of the National Academy of Sciences of the United States of America. 109, 13602-13607 (2012).
- Davies, K. M., et al. Macromolecular organization of ATP synthase and complex I in whole mitochondria. Proceedings of the National Academy of Sciences of the United States of America. 108, 14121-14126 (2011).
- Wurm, C. A., et al. Nanoscale distribution of mitochondrial import receptor Tom20 is adjusted to cellular conditions and exhibits an inner-cellular gradient. Proceedings of the National Academy of Sciences of the United States of America. 108, 13546-13551 (2011).
- Schermelleh, L., Heintzmann, R., Leonhardt, H. A guide to super-resolution fluorescence microscopy. The Journal of cell biology. 190, 165-175 (2010).
- Austin, J. R. 2nd, Staehelin, L. A. Three-dimensional architecture of grana and stroma thylakoids of higher plants as determined by electron tomography. Plant physiology. 155, 1601-1611 (2011).
- Ohta, K., et al. Beam deceleration for block-face scanning electron microscopy of embedded biological tissue. Micron. 43, 612-620 (2012).
- Buzhynskyy, N., Sens, P., Prima, V., Sturgis, J. N., Scheuring, S. Rows of ATP synthase dimers in native mitochondrial inner membranes. Biophysical journal. 93, 2870-2876 (2007).
- Larabell, C. A., Nugent, K. A. Imaging cellular architecture with X-rays. Current opinion in structural biology. 20, 623-631 (2010).
- Miao, J., Hodgson, K. O., Sayre, D. An approach to three-dimensional structures of biomolecules by using single-molecule diffraction images. Proceedings of the National Academy of Sciences of the United States of America. 98, 6641-6645 (2001).
- Maimon, T., Elad, N., Dahan, I., Medalia, O. The human nuclear pore complex as revealed by cryo-electron tomography. Structure. 20, 998-1006 (2012).
- Meyerson, J. R., et al. Determination of molecular structures of HIV envelope glycoproteins using cryo-electron tomography and automated sub-tomogram averaging. Journal of visualized experiments : JoVE. (58), (2011).
- Chen, S., et al. Electron cryotomography of bacterial cells. Journal of visualized experiments : JoVE. (39), (2010).
- Bharat, T. A., et al. Structural dissection of Ebola virus and its assembly determinants using cryo-electron tomography. Proceedings of the National Academy of Sciences of the United States of America. 109, 4275-4280 (2012).
- Bennett, A. E., et al. Ion-abrasion scanning electron microscopy reveals surface-connected tubular conduits in HIV-infected macrophages. PLoS pathogens. 5, e1000591(2009).
- Liu, J., et al. Cellular architecture of Treponema pallidum: novel flagellum, periplasmic cone, and cell envelope as revealed by cryo electron tomography. Journal of molecular biology. 403, 546-561 (2010).
- Liu, J., et al. Intact flagellar motor of Borrelia burgdorferi revealed by cryo-electron tomography: evidence for stator ring curvature and rotor/C-ring assembly flexion. Journal of bacteriology. 191, 5026-5036 (2009).
- Patla, I., et al. Dissecting the molecular architecture of integrin adhesion sites by cryo-electron tomography. Nature cell biology. 12, 909-915 (2010).
- Heuser, T., et al. Cryoelectron tomography reveals doublet-specific structures and unique interactions in the I1 dynein. Proceedings of the National Academy of Sciences of the United States of America. 109, E2067-E2076 (2012).
- Briggs, J. A. Structural biology in situ-the potential of subtomogram averaging. Current opinion in structural biology. 23, (2013).
- Cheng, D., Mitchell, D. R. G., Shieh, D. B., Braet, F. Current Microscopy Contributions to Advances in Science and Technology. Méndez-Vilas, A. 2, Formatex. (2012).
- Brust, D., et al. Cyclophilin D links programmed cell death and organismal aging in Podospora anserina. Aging cell. 9, 761-775 (2010).
- Gregg, C., Kyryakov, P., Titorenko, V. I. Purification of mitochondria from yeast cells. Journal of visualized experiments : JoVE. (30), (2009).
- Bhaskaran, S., et al. Breaking Caenorhabditis elegans the easy way using the Balch homogenizer: an old tool for a new application. Analytical biochemistry. 413, 123-132 (2011).
- Pavlov, P. F., Rudhe, C., Bhushan, S., Glaser, E. In vitro and in vivo protein import into plant mitochondria. Methods in molecular biology. 372, 297-314 (2007).
- Graham, J. M., et al. Ch. 3, Current protocols in cell biology. Bonifacino, J. S. , John Wiley and Sons. (2001).
- Zheng, S. Q., et al. UCSF tomography: an integrated software suite for real-time electron microscopic tomographic data collection, alignment, and reconstruction. Journal of structural biology. 157, 138-147 (2007).
- Mastronarde, D. N. Automated electron microscope tomography using robust prediction of specimen movements. Journal of structural biology. 152, 36-51 (2005).
- Kremer, J. R., Mastronarde, D. N., McIntosh, J. R. Computer visualization of three-dimensional image data using IMOD. Journal of structural biology. 116, 71-76 (1996).
- Pruggnaller, S., Mayr, M., Frangakis, A. S. A visualization and segmentation toolbox for electron microscopy. Journal of structural biology. 164, 161-165 (2008).
- Scheres, S. H., Chen, S. Prevention of overfitting in cryo-EM structure determination. Nature methods. 9, 853-854 (2012).
- Pettersen, E. F., et al. UCSF Chimera--a visualization system for exploratory research and analysis. Journal of computational chemistry. 25, 1605-1612 (2004).
- Dautant, A., Velours, J., Giraud, M. F. Crystal structure of the Mg.ADP-inhibited state of the yeast F1c10-ATP synthase. The Journal of biological chemistry. 285, 29502-29510 (2010).
- Carbajo, R. J., et al. Structure of the F1-binding domain of the stator of bovine F1Fo-ATPase and how it binds an alpha-subunit. Journal of molecular biology. 351, 824-838 (2005).
- Paumard, P., et al. The ATP synthase is involved in generating mitochondrial cristae morphology. The EMBO journal. 21, 221-230 (2002).
- Bharat, T. A., et al. Structure of the immature retroviral capsid at 8 A resolution by cryo-electron microscopy. Nature. 487, 385-389 (2012).
- Bartesaghi, A., Lecumberry, F., Sapiro, G., Subramaniam, S. Protein secondary structure determination by constrained single-particle cryo-electron tomography. Structure. 20, 2003-2013 (2012).
- Bai, X. C., Fernandez, I. S., McMullan, G., Scheres, S. H. Ribosome structures to near-atomic resolution from thirty thousand cryo-EM particles. eLife. 2, e00461(2013).
- Li, X., et al. Electron counting and beam-induced motion correction enable near-atomic-resolution single-particle cryo-EM. Nature methods. , (2013).
- Danev, R., Nagayama, K. Optimizing the phase shift and the cut-on periodicity of phase plates for TEM. Ultramicroscopy. 111, 1305-1315 (2011).
- Walter, A., et al. Practical aspects of Boersch phase contrast electron microscopy of biological specimens. Ultramicroscopy. 116, 62-72 (2012).
Ristampe e Autorizzazioni
Richiedi autorizzazione per utilizzare il testo o le figure di questo articolo JoVE
Richiedi AutorizzazioneThis article has been published
Video Coming Soon
Personale delle biblioteche
Copyright © 2025 MyJoVE Corporation. Tutti i diritti riservati