
Method Article
Visualizzare le interazioni proteina-DNA in cellule vive batteriche Uso fotoattivati singola molecola di monitoraggio
In questo articolo
Riepilogo
Fotoattivati microscopia localizzazione (PALM) in combinazione con inseguimento singola molecola permette l'osservazione diretta e la quantificazione delle interazioni proteina-DNA in cellule di Escherichia coli vive.
Abstract
Interazioni proteina-DNA sono al centro di molti processi cellulari fondamentali. Ad esempio, la replicazione del DNA, trascrizione, riparazione, e l'organizzazione dei cromosomi sono regolati da proteine che legano il DNA che riconoscono strutture o sequenze di DNA specifiche. Negli esperimenti in vitro hanno contribuito a generare modelli dettagliati per la funzione di molti tipi di proteine che legano il DNA, ma , i meccanismi esatti di questi processi e la loro organizzazione nel complesso contesto della cellula vivente rimangono molto meno comprensibili. Abbiamo recentemente introdotto un metodo per quantificare le attività di riparazione del DNA in cellule di Escherichia coli vive con fotoattivati localizzazione Microscopy (PALM) in combinazione con inseguimento singola molecola. Il nostro approccio generale identifica singoli eventi che legano il DNA dal cambiamento nella mobilità di una singola proteina upon associazione con il cromosoma. La frazione di molecole legate fornisce una misura quantitativa diretta per l'atto proteinaivity e l'abbondanza di substrati o siti di legame a livello di singola cellula. Qui, descriviamo il concetto del metodo e dimostrare la preparazione del campione, l'acquisizione dei dati e procedure di analisi dati.
Introduzione
Questo protocollo descrive la misurazione diretta delle interazioni proteina-DNA in cellule viventi Escherichia coli. La tecnica utilizza la variazione del coefficiente di diffusione di una singola proteina fluorescente come si lega il cromosoma (Figura 1). Per illustrare il metodo che usiamo DNA polimerasi I (Pol1), una proteina di legame al DNA prototipo che colma le lacune di DNA in replicazione filamento in ritardo e percorsi di riparazione escissione 1.
L'avvento del super-risoluzione microscopia a fluorescenza consente la visualizzazione di strutture molecolari in cellule con risoluzione nanometrica. Fotoattivati localizzazione Microscopy (PALM) impiega proteine fluorescenti che possono essere attivate da uno stato oscuro iniziale ad uno stato fluorescente (Figura 2). Solo un sottoinsieme di tutte le molecole marcate viene attivata in qualsiasi momento per determinare le loro posizioni in maniera sequenziale, indipendentemente tegli concentrazione totale di molecole marcate nel campione 2. La precisione di localizzazione per molecola dipende principalmente dalle dimensioni del fluorescente Point Spread Function (PSF), il numero di fotoni raccolti, e il segnale di fondo 3. Molte applicazioni di questo metodo si concentrano sulla migliore visualizzazione delle strutture cellulari. La realizzazione che Palm può essere combinato con singola molecola inseguimento 4 aperto nuove strade per seguire direttamente il movimento dei numeri arbitrari di proteine marcate nelle cellule viventi. Aumento della sensibilità e risoluzione temporale di microscopi a fluorescenza permettono ora il monitoraggio delle proteine fluorescenti diffondenti singoli nel citoplasma batterico 5.
Qui, impieghiamo PAmCherry, una proteina fluorescente ingegnerizzato che converte irreversibilmente da uno stato non fluorescente stato iniziale ad uno fluorescente durante l'irradiazione con luce 405 nm 6. Fluorofori PAmCherry attivati possono essere immagined da eccitazione a 561 nm e monitorati per diversi fotogrammi fino photobleaching. Dimostriamo la capacità del metodo per identificare transitori eventi che legano il DNA di singole proteine utilizzando una fusione di Pol1 e PAmCherry. Il trattamento di cellule con methanesulfonate metile (MMS) provoca danni al DNA metilazione che viene trasformato in substrati DNA gapped da enzimi di riparazione di base escissione. Il nostro metodo mostra chiaramente vincolante delle singole molecole Pol1 in risposta al danno MMS 7.
Protocollo
1. Cell Culture
Utilizzare provette di coltura sterili e relativi puntali. Il E. coli AB1157 polA-PAmCherry porta un C-terminale PAmCherry fusione di Pol1. La fusione è stato inserito nella posizione cromosomica nativo sostituendo il gene wild-type utilizzando lambda-Red ricombinazione come descritto in Datsenko et al. 8 funzionalità della proteina di fusione è stata confermata come giudicato da tassi di crescita cellulare e la sensibilità all'agente che danneggiano il DNA metil methanesulfonate (MMS). Ulteriori informazioni sulla costruzione del ceppo cella in cui si trovano Uphoff et al. 7, Datsenko et al. 8, e Reyes-Lamothe et al. 9 colture cellulari sono coltivate in terreno minimo M9 per ridurre autofluorescenza ed evitare le particelle di fondo sul vetrino del microscopio. In alternativa, un terreno specifico nutriente può essere utilizzato 10.
- Streak E. coli ceppo AB1157 polA-PAmCherry da uno stock di glicerolo congelato in una Luria Broth (LB) piastra di agarosio con antibiotici selettivi (qui, 25 mcg / ml kanamicina) e incubare a 37 ° C durante la notte.
- Inoculare una coltura LB 5 ml da una singola colonia di cellule e crescere a 37 ° C agitando a 220 rpm per 3 ore.
- Diluire la cultura 1:10.000 in 5 ml di terreno minimo (M9 medio, aminoacidi MEM + prolina, vitamine MEM, 0,2% glicerolo) e incubare a 37 ° C in agitazione a 220 rpm durante la notte.
- Il mattino seguente, misurare la densità ottica (OD) utilizzando uno spettrofotometro e diluire la cultura in 5 ml di mezzo fresco minima OD 0,025. Crescere per 2 ore a 37 ° C in agitazione a 220 rpm di prima fase esponenziale (OD 0.1).
- Concentrato 1 ml di cellule in una provetta da 1,5 ml microcentrifuga per centrifugazione a 2300 xg per 5 min. Rimuovere il surnatante e risospendere il pellet cellulare in 20 microlitri di media residua e vortex.
2. Microscopio diapositiva Preparatione
- Preparare una soluzione di agarosio a bassa fluorescenza 1,5% in dH 2 O. Utilizzare un forno a microonde per sciogliere l'agarosio fino a quando la soluzione è limpida. Mescolare 500 ml di soluzione di agarosio fuso con 500 ml di terreno minimo 2x pipettando gentilmente su e giù un paio di volte.
- Distribuire la soluzione di agarosio in modo uniforme sul centro di un vetrino da microscopio (n spessore 1.5). Questo deve essere fatto in fretta prima che l'agarosio si raffredda, evitando le bolle.
- Appiattire il pad con un secondo vetrino (No spessore 1.5). Per rimuovere le particelle fluorescenti sfondo, vetrini sono stati precedentemente bruciato in un forno a 500 ° C per 1 ora. Coprioggetto Burned possono essere conservati per settimane a temperatura ambiente rivestito in lamina di alluminio.
- Per gli esperimenti danno del DNA, preparazione di un pad agarosio contenente MMS 100 mm. Seguire la procedura dei passi 2.1-2.3, ma aggiungere 8,3 microlitri MMS a 500 pl di mezzo M9 prima della miscelazione con 500 ml di fuso agarosio 1,5%, per una concentrazione finale di 100 mM MMS. (Attenzione! MMS è tossico e mutageno e deve essere trattato con i guanti, mascherina, occhiali e camice da laboratorio).
- Rimuovere la slitta superiore del pad e aggiungere 1 ml di sospensione cellulare concentrata sul pad. Immobilizzare le cellule coprendo il tappetino con un vetrino inutilizzato bruciato (No spessore di 1,5, corrispondente alla specifica obiettivo microscopio) e premendo delicatamente sul vetrino. Le cellule dovrebbero essere esposte entro 45 min di immobilizzazione prima che si asciuga pad agarosio. Per prevenire l'essiccazione durante gli esperimenti più lunghi, pad agarosio possono essere sigillati con guarnizioni in silicone.
- Per gli esperimenti di danno al DNA, incubare cellule immobilizzate sul pad agarosio contenente MMS 100 mM per 20 minuti in un contenitore umidificato a temperatura ambiente prima di imaging.
3. Preparazione Microscopia Data Acquisition
PALM basa sul rilevamento e localizzazione precisa di proteine fluorescenti singoli. La sensibilità e l'allineamento ottimaleil microscopio è critica per la qualità dei dati. Singola molecola microscopi a fluorescenza tipicamente impiegano Total Internal Reflection (TIR) illuminazione per migliorare il rapporto segnale-rumore di soli giochi a fluorofori all'interno di una sezione sottile sopra la superficie del vetrino. Qui, imaging all'interno E. coli richiede altamente illuminazione inclinata 11, che può essere realizzato su un microscopio TIRF leggermente diminuendo l'angolo della luce di eccitazione. Imaging PAmCherry richiede inoltre un laser fotoattivazione 405 nm e un laser di eccitazione 561 nm. L'emissione di fluorescenza è registrata su un elettrone moltiplicando CCD (EMCCD) fotocamera con un ingrandimento risultante in una lunghezza di pixel di 114,5 nm / pixel. Per una precisione ottimale localizzazione, la dimensione dei pixel deve corrispondere approssimativamente alla larghezza deviazione standard del PSF garantirà un campione sufficiente senza diffondere il segnale su troppi pixel. Figura 3 mostra uno schema di una configurazione PALM minima. Film 1dà l'impressione del processo di costruzione microscopio su misura; vedere Uphoff et al 7 per una descrizione dettagliata dello strumento..
- Esecuzione di allineamento microscopio di routine. Misurare la 405 nm e 561 nm intensità laser ad onda continua di fronte al obiettivo. Regolare l'intensità 561 nm a 3,5 mW (~ 400 W / cm 2) e 405 nm intensità di 10 μW (~ 1 W / cm 2). Usare una ruota continuamente variabile filtro a densità neutra che permette la regolazione graduale dell'intensità di 405 nm 0-10 μW. Spegnere l'illuminazione laser fino all'inizio dell'esperimento.
- Mettere il campione sul palco microscopio e portare le cellule a fuoco in modalità microscopia a luce trasmessa (Figura 4A). Il guadagno della telecamera EMCCD deve essere spento per evitare danni alla fotocamera da sovraesposizione.
- Definire un FOV ritagliata per ridurre la dimensione dei dati e aumentare la velocità di read-out fotocamera.
- Coprire il campione dalla luce ambientale e prosch sul EMCCD guadagno della telecamera.
- Impostare il frame rate a 15.26 msec / telaio (compresi 0.26 msec fotocamera tempo di lettura). Vedere "Tempo di esposizione e di eccitazione intensità" nella sezione di discussione.
- Visualizzare i dati della fotocamera per controllare il segnale fondo scuro (Figura 4B).
- Accendere il laser 561 nm e controllare il segnale di eccitazione sfondo (Figura 4C).
- Accendere il laser 405 nm per fotoattivazione delle proteine di fusione Pol1-PAmCherry e aumentare l'intensità fino alla comparsa PSF fluorescenza.
- Regolare l'angolo del fascio di eccitazione per illuminare solo una sezione sottile del campione vicino alla superficie coprioggetto.
- A tal fine, il fascio laser viene focalizzato sul piano focale posteriore di un obiettivo 100X 1.4 NA (Figura 3). Traducendo la lente di focalizzazione perpendicolare al fascio sposta la distanza dal centro dell'obiettivo causando il fascio per uscire dell'obiettivo sotto un angolo.
- Aim per massimizzare l'intensità di fluorescenza e minimizzare il segnale di fondo. Si noti che il rigoroso TIR eccitazione è ottimale per fluorofori immagine entro 100 nm della superficie coprioggetto, tuttavia, di imaging DNA-binding proteine associate alla E. coli nucleoide richiede più profonda illuminazione fino a 0,8 micron.
4. Acquisizione Dati
Qui, descriviamo il protocollo generale per l'acquisizione di un filmato PALM. La stessa procedura si applica per l'imaging proteine di fusione Pol1-PAmCherry in intatto E. coli cellule e sotto trattamento danni al DNA continuo con MMS. Applicazione del metodo di proteine di fusione di peso o di numero di copia molecolare diverso per ogni cella richiederà diverse impostazioni di acquisizione (vedere la sezione Discussione).
- Trovare un nuovo campo di vista (FOV) di cellule in modalità microscopia luce trasmessa e fuoco l'immagine. Prendete uno snapshot telecamera per registrare l'(4A Figura) contorni cellulari.
- Coprire il campione dalla luce ambientale e accendere la EMCCD guadagno della telecamera.
- Accendere il laser 561 nm e sbiancare la autofluorescenza cellulare e macchie di sfondo sul vetrino per alcuni secondi prima di iniziare l'acquisizione dati. Per le cellule coltivate e ripreso in M9 medio e utilizzando vetrini bruciato di solito c'è molto poco fluorescenza di fondo, tuttavia, prebleaching potrebbe essere utile per l'imaging cellule in un terreno di coltura ricco come LB. Si noti che l'illuminazione intensa è tossico per le cellule così prebleaching deve essere mantenuta al minimo.
- Avviare l'acquisizione di un filmato PALM in continuo 561 nm di eccitazione a 15.26 msec / telaio.
- Accendere il laser 405 nm e aumentare gradualmente l'intensità nel corso del film, arrivando fino a 1 W / cm 2. Evitare superiori 405 nm intensità che causano autofluorescenza cellulare. Prestare attenzione alla densità delle molecole fluorescenti - è importante mantenere i tassi bassi di attivazione tale che PSF sono chiaramenteisolato in ogni fotogramma (figure 4D-F).
- Registrare 10.000 fotogrammi / film (a seconda del numero di molecole da acquisire per cella); un film richiede in genere 2-3 minuti e richiede 0,5-1 GB di spazio su disco a seconda delle dimensioni del FOV.
- Ripetere la procedura di acquisizione per più FOV. Si noti che ogni FOV può essere ripreso solo una volta perché fluorofori PAmCherry ottenere fotoattivati e sbiancato irreversibile.
5. Analisi dei dati
Un quadro di analisi automatizzata dei dati e robusto è essenziale per le prestazioni e l'efficienza del metodo. Usiamo software personalizzato scritto in MATLAB.
- Effettuare l'analisi localizzazione utilizzando algoritmi descritti in Crocker et al. 12, Holden et al. 13, HoldenI et al. 14, e Wieser et al. 15 PSF vengono prima identificati in un'immagine filtrata passa banda utilizzando un kernel gaussiano con 7 pixeldiametro s (Figura 5A). Posizioni candidati corrispondono ai PSF con intensità di picco dei pixel di sopra di 4,5 volte la deviazione standard del segnale di fondo (Figura 5B). Il pixel localmente luminoso per candidato PSF serve come ipotesi iniziale per il montaggio di una funzione gaussiana ellittica (Figura 5C). I parametri atti liberi sono: posizione x, posizione y, x larghezza, y-width, angolo di rotazione, ampiezza e offset sfondo. La maschera gaussiana ellittica rappresenta molecola durante il tempo di esposizione, che sfuma e si deforma il PSF.
- Tracciare le risultanti (x, y) localizzazioni da tutti i frame del film PALM sull'immagine microscopia luce trasmessa della stessa FOV. Localizzazioni di Pol1-PAmCherry dovrebbero apparire nella zona centrale di E. coli (Figura 6a). Se molti localizzazioni appaiono esterna delle cellule, la soglia di localizzazione è stato troppo basso o il campione conteneva particelle fluorescenti sfondo.
- Per l'analisi di inseguimento automatico, l'attuazione MATLAB dell'algoritmo descritto in Crocker et al. 12 può essere usato (vedere "analisi Diffusion" nella sezione Discussione). Posizioni che appaiono in fotogrammi successivi all'interno di una finestra di inseguimento definita dall'utente sono collegati a formare una traiettoria. Nel caso in cui più localizzazioni verificano nella stessa finestra, tracce vengono assegnati esclusivamente minimizzando la somma delle lunghezze di passo. Per una discussione dettagliata delle varie considerazioni nel calcolo coefficienti di diffusione dai dati di monitoraggio singola particella, vedere Wieser et al 15.
- L'algoritmo utilizza un parametro di memoria per tenere conto di transitori lampeggiante o localizzazioni perse durante un brano. Qui, abbiamo impostato il parametro di memoria di 1 fotogramma; valori superiori possono essere utilizzati per il monitoraggio fluorofori con longevi stati scure.
- Scegliere una finestra di monitoraggio adeguato sulla base dei seguenti punti di calibrazione. Per Pol1, usiamo 0,57 micron (5 pixel).
- Eseguire l'algoritmo di tracking per una serie di tracciare parametri della finestra. Calcolare il numero di tracce misurate per cella in funzione della finestra di monitoraggio per identificare la finestra di inseguimento più piccola possibile che non contempla brani (Figura 6B).
- Tracciare le tracce risultanti dell'immagine microscopia luce trasmessa della stessa FOV su per visualizzare la distribuzione spaziale della molecola movimento all'interno delle cellule. Brani Pol1 dovrebbero visualizzare diffusione confinati all'interno delle cellule singole (figure 6C-D).
- Se una frazione di brani sembra attraversare tra cellule questo suggerisce che molecole separate stati erroneamente collegati perché la finestra inseguimento è stato scelto troppo grande e / o il tasso di fotoattivazione è troppo elevato (Figura 6E).
- Tracciare la distribuzione cumulativa delle lunghezze di passo tra le localizzazioni consecutivi (Figura 6F). La curva si alza e satura intoppi per sufficientemente ampie finestre di monitoraggioma mostra un bordo di taglio se la finestra è stata scelta troppo piccola.
- Per analizzare le caratteristiche di diffusione di Pol1, calcola lo spostamento quadratico medio (MSD) tra localizzazioni consecutivi per ogni traccia con un totale di N passi):
MSD = 1 / (N-1) Σ i = 1 N-1 (X i +1 - x i) 2 + (y i +1 - y i) 2.
Includi ascolti solo con almeno 4 punti (N ≥ 5 localizzazioni) per ridurre l'incertezza statistica nei valori MSD. - Tracciare una curva di valori MSD su una gamma di tempi di latenza calcolando spostamenti in più fotogrammi (Figura 6G). La forma della curva MSD può aiutare a classificare il movimento molecolare osservata (Figura 6H).
- Calcolare il coefficiente di diffusione apparente D * per traccia dal MSD:
D * = MSD / (4 Dt) - σ loc 2 / At.
Il secondo tehm corregge l'errore di localizzazione stimata (qui, σ loc = 40 nm e At = 15.26 msec, vedere Wieser et al. 15). - Tracciare un istogramma dei valori * D misurati da tutte le tracce nella (Figura 7A) FOV.
- Identificare molecole Pol1 individuali che appaiono legato al cromosoma base al valore * D misurata per traccia. Separare le popolazioni di limite (distribuzione tagliente centrato in D * ~ 0 micron 2 / sec) e le molecole che diffondono liberamente (ampia distribuzione centrato in D * ~ 0,9 micron 2 / sec) impostando una soglia D * <0,15 micron 2 / sec ( barre rosse nelle figure 7A e 7D).
- Effettuare la localizzazione, il monitoraggio e l'analisi diffusione per Pol1 in cellule danneggiate (Figure 7A-C), e in cellule in trattamento DNA danno con MMS (figure 7D-E). La frazione di brani rilegati fornisce una misura quantitativa diretta del DNUn'attività riparazione di Pol1 in vivo.
Risultati
Il concetto di fotoattivati singola molecola inseguimento per studiare le interazioni proteina-DNA in vivo è illustrato in Figura 1. Proteine di fusione PAmCherry vengono rilevati in tempo reale E. coli in maniera sequenziale per fotoattivanti singole molecole stocasticamente con luce 405 nm con una frequenza inferiore a una molecola per cella alla volta. Molecole attivate sono esposte in continuo 561 nm di eccitazione. Movimento molecolare nella cella può essere seguito collegando localizzazioni vicine in una serie di fotogrammi fino fotoscolorimento irreversibile. Poiché la diffusione di proteine che legano il DNA è rallentato dopo il legame del cromosoma, il coefficiente di diffusione apparente D * ottenuta per traccia riporta direttamente sulle singole interazioni proteina-DNA.
La Figura 2 mostra fotoattivazione di proteine di fusione Pol1-PAmCherry in diretta E. coli. L'influenza dell'intensità 405 nm on la densità delle molecole fluorescenti può essere visto in Figura 4. Si noti che la densità non dipende soltanto dall'intensità 405 nm ma in aggiunta per il numero di molecole che sono disponibili per l'attivazione, la vasca delle molecole residue è esaurito nel corso di un film PALM.
Analisi localizzazione viene eseguita per ogni fotogramma di un film PALM come illustrato nella Figura 5. Abbiamo misurato la precisione di localizzazione utilizzando molecole immobili in cellule fissate o molecole associate in cellule vive. Le nostre impostazioni di acquisizione ha dato una precisione di localizzazione di σ loc = 40 nm, in accordo con la previsione teorica 3.
Le risultanti localizzazioni Pol1 occupano la zona centrale della cella (Figura 6A), ampiamente ricapitolati l'organizzazione spaziale della E. coli nucleoide 7. La maggior parte dei Pol1 ascolti nelle cellule danneggiate visualizzare diffusione come mostrato nella Figura 6C. Una tipica cellula contiene diverse centinaia di tracce Pol1 (Figura 6D), in linea con il numero di copie di circa 400 molecole Pol1 per E. . coli cella 1 Figure 6B e 6E-F fornire indicazioni sulla scelta di un parametro finestra di monitoraggio adatto - se la finestra di monitoraggio è troppo grande, diverse molecole sono più probabilità di diventare erroneamente collegato a una traccia, se la finestra di monitoraggio è troppo piccola, brani con passi più lunghi sarà diviso. La curva di MSD per Pol1 aumenta linearmente per tempi brevi lag e saturi in tempi di latenza più lunghi a causa di confinamento delle cellule (Figura 6G). Diversi tipi di moto molecolare possono essere identificati mediante analisi MSD. Movimento diretto dà una curva parabolica, moto browniano è caratterizzata da una linea retta, la curva di diffusione confinato raggiunge un plateau; un offset della curva MSD per particelle immobile rappresenta lincertezza ocalization (Figura 6H). Ulteriori informazioni sulla singola particella consigli di monitoraggio e la risoluzione dei problemi può essere trovato in Arnauld et al 16.
Abbiamo già applicato il metodo per misurare l'attività di riparazione del DNA di Pol1 in risposta al danno alchilazione del DNA esogeno 7. Il D * istogramma di Pol1 ascolti nelle cellule danneggiate mostra una popolazione dominante di diffondere molecole (Figure 7A-C). Una piccola frazione di 2,7% molecole Pol1 legate probabilmente è coinvolto nella replica filamento in ritardo e la riparazione dei danni al DNA endogeno. Sotto continuo 100 danni MMS mM, la popolazione di brani con D * ~ 0 micron 2 / sec aumenta al 13,8% (Figura 7D). Queste tracce rappresentano le singole molecole Pol1 dello spettacolo sintesi di riparazione del DNA per colmare le lacune a singolo nucleotide come parte del percorso di riparazione di base-escissione. Le posizioni dei brani rilegati mostrano le posizioni dei singoliDanno al DNA e siti di riparazione (Figura 7E-F).

Figura 1. Rappresentazione grafica del metodo. (A) La proteina PAmCherry fluorescente può essere fotoattivato da uno stato iniziale non fluorescente su irradiazione con luce 405 nm. Lo stato brillante è eccitato a 561 nm ed emette fluorescenza circa 600 nm fino a quando i candeggianti fluoroforo irreversibile. (B) Il controllo del tasso di fotoattivazione permette l'imaging solo un singolo stocasticamente activated protein PAmCherry di fusione per cella, in qualsiasi momento, mentre il arbitrariamente grande piscina di molecole che non sono ancora stati attivati o sono già state sbiancate resti in uno stato di buio. (C) La posizione della molecola fluorescente è determinato dal centro della PSF isolata e tracked per diversi fotogrammi fino photobleaching. (D) Tracce di molte molecole vengono registrati in maniera sequenziale. (EF) L'interazione di una proteina legante il DNA con una sequenza bersaglio cromosomico o struttura arresta il moto diffusivo casuale. Molecole legati e non legati si distinguono per il coefficiente di diffusione apparente D * estratto da singole tracce. La frazione risultante di molecole legate dà una misura quantitativa per l'attività di una proteina legante il DNA in vivo. Clicca qui per vedere l'immagine ingrandita.
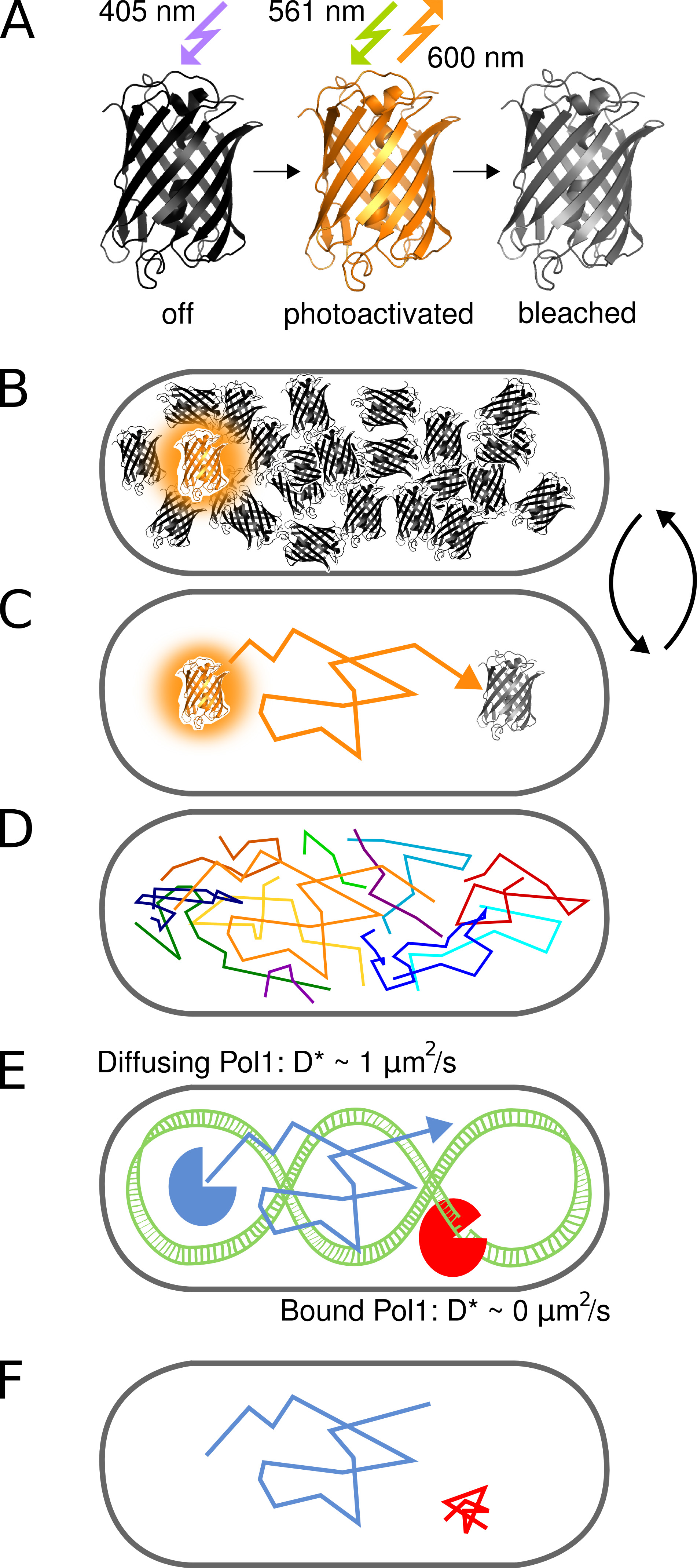{kind=link}

Figura 2. Fotoattivazione di Pol1-PAmCherry in diretta E. coli. Bar Scala: 1 micron. Schemi vengono visualizzati sotto ogni pannello. ( A) luce immagine al microscopio di cellule immobilizzate su un pad agarosio trasmessa. (B) Phototactivating un singolo fluoroforo PAmCherry in una cella. (C) Un tasso fotoattivazione alto aumenta il numero di molecole fluorescenti. (D) Integrato PAmCherry fluorescenza da un film PALM. Clicca qui per vedere l'immagine ingrandita.
{kind=link}

Figura 3. Schema di una configurazione minima PALM per le proteine fotoattivanti e di imaging PAmCherry fusione D1:. Specchio dicroico (es. 550-pass lungo nm). D2: specchio dicroico (es. 570-pass lungo nm). L1: Lente di collimazione. L2: lente TIR. L3: obiettivo Tube.1177/51177fig3highres.jpg "target =" _blank "> Clicca qui per vedere l'immagine ingrandita.

Figura 4. Immagini rappresentative da un filmato PALM con 15.26 msec / telaio Barre di scala:. 1 micron. (A) immagine luce delle cellule immobilizzate su un pad agarosio trasmessa. (B) immagine di sfondo scuro misurata sulla fotocamera EMCCD con i laser spento. (C) l'immagine di sfondo di eccitazione in continuo 561 nm di eccitazione prima fotoattivazione. (DF) Aumento dell'intensità 405 nm porta a tassi più elevati di Fotoattivazione PAmCherry, impressi in continuo 561 nm di eccitazione. Le aree in scatola sono riportati di seguito ingrandita. (D) a bassa intensità di 405 nm (<1 μW) attivi pochissime molecole fluorescenti per FOV. (E) Medium 405risultati intensità nm (~ 2 μW) Fotoattivazione in una buona densità PSF per la localizzazione e l'analisi di monitoraggio. (F) superiore 405 nm intensità (~ 10 μW) attiva più di una molecola fluorescente in alcune cellule, che oscura la localizzazione e l'analisi di monitoraggio. Clicca qui per vedere l'immagine ingrandita.
{kind=link}

Figura 5. Illustrazione delle analisi di localizzazione. Bar Scala: 1 micron (A) passa-banda filtraggio rimuove il rumore dei pixel spuri e appiattisce gradienti di intensità attraverso il FOV.. (B) PSF candidati sono identificati nell'immagine filtrata basata su una soglia definita dall'utente che è scelto per minimizzare i falsi rilevamenti positivi e falsi negativi. Il threshold corrisponde all'intensità minima di un pixel candidato divisa per la bassa deviazione standard (segnale-rumore SNR). (C) Il pixel localmente luminoso che passa la soglia serve come ipotesi localizzazione iniziale (croce arancione) per una bidimensionale ellittica Gaussiana. Barre di scala: 0,5 micron. La localizzazione super-risoluzione risultante (croce blu) ha una precisione media di σ loc = 40 nm. Clicca qui per vedere l'immagine ingrandita.
{kind=link}

.. Figura 6 Illustrazione delle analisi di monitoraggio bar Scala: 1 micron. (A) Tutte le localizzazioni rilevate di Pol1-PAmCherry in una cella esempio. (B) Numero brani rilevareEd in cella esempio in funzione della finestra di monitoraggio. Finestre di monitoraggio piccole divisi traiettorie molecola, che porta a artefatta elevato numero di tracce. La linea tratteggiata indica la nostra scelta per il parametro window inseguimento (0,57 micron, 5 pixel) - questo dà un buon compromesso tra il rilevamento della distribuzione completa di passaggi e mantenendo le traiettorie di diverse molecole intatte. (C) Esempio traccia di una singola molecola Pol1-PAmCherry. (D) Tutte le tracce misurati visualizzati in colori casuali. (E) Monitoraggio artefatti se si sceglie la finestra inseguimento troppo grande (qui 0,8 micron, 7 pixel) o la densità di PSF per frame è troppo alto. (F) distribuzioni cumulative delle lunghezze di passo per finestre inseguimento: 0.34 micron (3 pixel, linea rossa), 0,57 micron (5 pixel, linea blu) e 0,80 micron (7 pixel, linea verde). Notare che la finestra di monitoraggio 0.34 micron taglia passi più lunghi di 0.34 micron che chiaramentetronca la distribuzione completa di passi. La finestra 0.57 micron inseguimento rileva quasi la stessa distribuzione di passi come fa la finestra inseguimento 0,80 micron. Curva (G) MSD mostra diffusione limitata di Pol1. Curve (H) Schema MSD per il moto diretto, moto browniano, la diffusione limitata, e le particelle immobile. Clicca qui per vedere l'immagine ingrandita.
{kind=link}

Figura 7. La misura diretta dell'attività di riparazione del DNA di Pol1 in diretta E. . coli cellule bar Scala: 1 micron. (A) Istogramma della apparente coefficiente di diffusione D * per tutte le tracce di 4 o più fasi di un FOV di cellule danneggiate (N = 4.162 brani). La frazione di molecole classificate come b ound è evidenziato in rosso. (BC) Tracce di Pol1-PAmCherry sono mostrati un'immagine al microscopio a luce trasmessa su. Tracce classificate come vincolati in base al loro coefficiente di diffusione sono mostrati in rosso. (D) D * istogramma per le tracce Pol1 misurati in cellule immobilizzate su un pad agarosio con MMS 100 mM e incubate per 20 minuti prima di imaging (N = 2.128 brani). La popolazione di molecole legate impegnati nella riparazione del DNA è mostrato in rosso. (EF) Pol1-PAmCherry ascolti immagine al microscopio a luce trasmessa che mostra le tracce dei singoli eventi Pol1 di riparazione del DNA in rosso. Clicca qui per vedere l'immagine ingrandita.
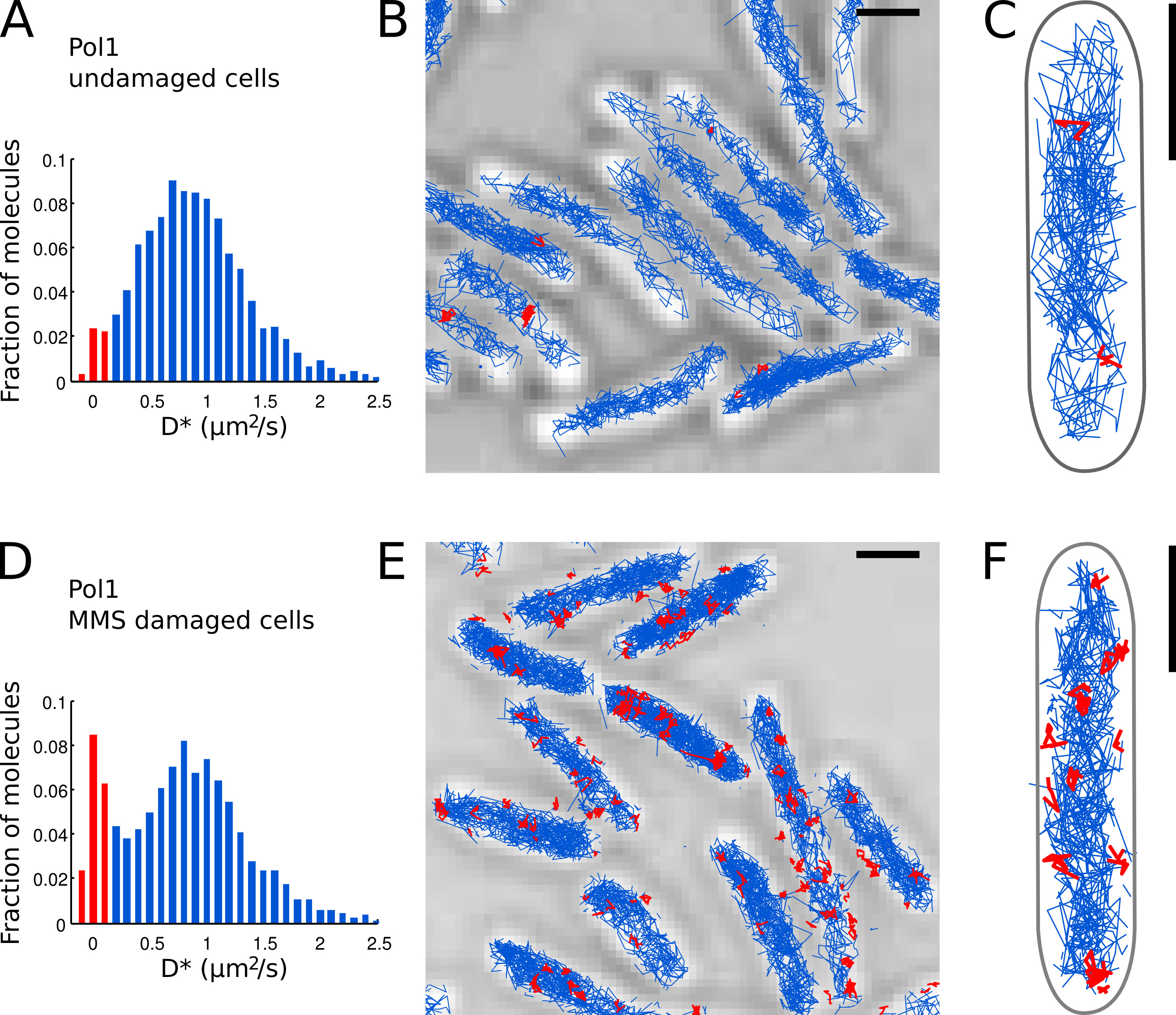{kind=link}

Movie 1. Costruire una configurazione PALM su misura.JoVE_Uphoff_Movie1.avi "target =" _blank "> Clicca qui per vedere il video.
Discussione
Discutiamo diverse considerazioni chiave per il successo dell'esperimento.
Scelta e l'espressione della proteina di fusione fluorescente: Vi è una grande tavolozza di proteine fluorescenti photoactivatable e photoswitchable 17. La scelta specifica dipende dalle caratteristiche del microscopio, in particolare i laser e filtri disponibili. La combinazione di 405 nm e 561 nm è ideale per proteine fluorescenti photoactivatable comuni. Abbiamo scelto PAmCherry 6 perché è monomerica e mostrato alcuna aggregazione nelle cellule. Inoltre, fotoattivazione irreversibile consente di conteggiare il numero di fluorofori attivati per misurare il numero di copie per cellula proteina. Invece di esprimere la proteina di fusione da un plasmide, preferiamo inserimento cromosomica del gene codificante per la proteina di fusione al wild-type locus. Questo assicura la completa sostituzione della proteina di interesse con il fluorescente versione e mantenendo il livello di espressione wild-type.
Tasso fotoattivazione: è importante regolare il tasso di fotoattivazione tale che in media meno di una molecola per cella è nello stato fluorescente in qualsiasi fotogramma del film. Questo dipende dall'intensità 405 nm e il numero di molecole lasciato da attivare. A densità molto bassa di imaging, tuttavia, non tutte le molecole saranno ripreso prima della fine del film o filmati molto lunghi devono essere acquisita. Il numero di fotogrammi registrati al film dipende dal numero di copie di proteine di fusione per cellula e la media photobleaching durata PAmCherry alle condizioni di eccitazione utilizzati. Il numero di copie di Pol1 è ~ 400 molecole / cella 1 e il valore medio della distribuzione vita photobleaching esponenziale era ~ 4 fotogrammi. Aumentando l'intensità 405 nm gradualmente, l'attivazione viene uniformemente distribuito sulle 10.000 fotogrammi del film. Pertanto, ogni cella è chiavied da molecole fluorescenti per un totale di ~ 1.600 fotogrammi, garantendo poca sovrapposizione di PSF e complicazioni inseguimento in un film di 10.000 fotogrammi.
Tempo di esposizione e di eccitazione intensità: Primo, i tempi di esposizione devono essere sufficientemente breve da osservare PSF nitide con poco movimento sfocatura. Tuttavia, il frame rate dovrebbe essere scelto per produrre il movimento molecolare osservabile tra i fotogrammi successivi al di là dell'incertezza localizzazione, altrimenti fotoni cruciali sono sprecati dal sovracampionamento pista. Il movimento delle molecole non legate deve essere campionato a intervalli di tempo sufficientemente lungo da essere chiaramente distinguibile dal moto apparente delle molecole legate a causa dell'incertezza localizzazione. Quando è impostato il tempo di esposizione, l'intensità PSF deve essere regolata. La precisione di localizzazione di un PSF aumenta con il numero di fotoni rilevati lungo la durata di un frame. Elevate intensità di eccitazione aumentano i bu tasso di emissione di fotonit anche il tasso photobleaching e segnale di fondo. Utilizzare l'intensità di eccitazione più bassa che dà la precisione di localizzazione desiderata. Per Pol1-PAmCherry abbiamo scelto 15.26 msec / telaio e 3,5 mW 561 nm di eccitazione (400 W / cm 2). È importante confermare vitalità cellulare per le condizioni di imaging particolare monitorando la crescita cellulare e la morfologia prima e dopo l'acquisizione dei dati (vedere informazioni supplementari Uphoff et al. 7).
Pol1 presenta un tempo di legame di ~ 2 sec per un substrato DNA gapped in vivo 7; ci aspettiamo quindi la maggior parte delle molecole di essere sia nello stato associato o non associato per l'intera durata della traccia. Molecole legate appaiono sostanzialmente immobile perché i siti cromosomiche hanno un coefficiente di diffusione di diversi ordini di grandezza inferiore (~ 10 -5 micron 2 / sec, Elmore et al. 18) che Pol1 diffusione nel citoplasma (~ 1 micron 2 / sec).
Analisi Diffusione: L'apparente coefficiente di diffusione D * viene calcolato dalla MSD delle singole tracce, in media su un minimo di 4 punti (5 fotogrammi) per ridurre l'errore statistico. Si noti che ~ 75% di molecole candeggiare entro meno di 5 fotogrammi per le condizioni di imaging descritte. Tali brani brevi non forniscono certezza statistica sufficiente per distinguere molecole legati e non legati. Tuttavia, le relative frazioni di molecole legati e non legati che informano l'attività della proteina sono indipendenti dal numero totale dei brani analizzati.
È utile per spiegare la errore di localizzazione PSF (σ loc) nel calcolo di D * perché l'incertezza aggiunge un passo casuale apparente per ciascuna localizzazione di una molecola 15.
Per migliorare la classificazione delle molecole legate e diffondenti, si consiglia di calcolo D * both da spostamenti solo stadio, e gli spostamenti sul tempo di due telai. E 'quindi possibile impostare due soglie * D distinti: D * (15 msec) <0,15 micron 2 / sec e D * (30 msec) <0,075 micron 2 / sec.
Si noti che D * è un coefficiente di diffusione apparente che è influenzata da confinamento delle cellule dei brani e movimento di sfocatura dovuta al trasudamento durante il tempo di esposizione. Per estrarre i coefficienti di diffusione imparziali precise, si è dimostrato utile per confrontare il moto osservato ai dati simulati sulla base di un modello stocastico browniano movimento 5,7. Dati simulati possono anche essere usati per testare le procedure di analisi dei dati.
Le potenziali applicazioni di questo metodo: Abbiamo descritto un approccio generale per la visualizzazione e la quantificazione delle interazioni proteina-DNA in vivo dalla variazione della mobilità di una proteina dopo il legame al cromosoma. Le attività di DNA-oRNA-binding proteins coinvolte nella riparazione, replicazione, trascrizione e manutenzione cromosoma possono così seguire in tempo reale a livello di singola cellula con una risoluzione spaziale di sotto del limite di diffrazione ottica. Fotoattivati inseguimento singola molecola si estende metodi di monitoraggio tradizionali che si limitano a poche molecole marcate per cella. Un metodo alternativo che misura la diffusione molecolare in vivo è Fluorescence Recovery After Photobleaching (FRAP). Mentre FRAP è molto utile per misurare le caratteristiche di diffusione globali a grandi cellule, è limitato nella sua capacità di risolvere diverse specie molecolari con differenti mobilità in un ambiente spazialmente eterogeneo, soprattutto per le piccole cellule batteriche.
Abbiamo applicato fotoattivati singola molecola di monitoraggio per misurare le attività che legano il DNA e le localizzazioni subcellulari di una gamma di diverse proteine in E. coli compresi Pol1, ligasi del DNA, proteine Fis, DNApolimerasi III 7, così come la manutenzione strutturale dei cromosomi proteine MukB, E, e F 19. Anticipiamo il metodo può essere adattato anche ad altri tipi di cellule.
Divulgazioni
Gli autori dichiarano di non avere interessi finanziari in competizione.
Riconoscimenti
Riconosciamo Justin Pinkney e Johannes Hohlbein aiuto per la costruzione del microscopio su misura e Seamus Holden per il software di localizzazione. Rodrigo Reyes-Lamothe si ringrazia per la fornitura del E. coli ceppo. La ricerca è stata finanziata dalla Commissione Europea Settimo programma quadro di Grant FP7/2007-2013 SALUTE-F4-2008-201418, UK Biotecnologie e Scienze Biologiche Research Council di Grant BB/H01795X/1, e il Consiglio europeo Research Grant 261.227 a ANK. DJS è stato finanziato da un programma Wellcome trust di Grant WT083469. SU è stato sostenuto da una borsa di studio di dottorato MathWorks.
Materiali
| Name | Company | Catalog Number | Comments |
| E. coli strain carrying a chromosomal insertion for a PAmCherry DNA-binding fusion protein | created by Lambda-Red recombination | ||
| MEM amino acids | Invitrogen | 11130-051 | minimal medium supplement |
| MEM vitamins | Invitrogen | 11120-052 | minimal medium supplement |
| Agarose | BioRad | 161-3100 | certified molecular biology grade |
| Microscope coverslips No 1.5 thickness | Menzel | BB024060SC | remove background particles by heating slides in furnace at 500 °C for 1 hr |
| Methyl methanesulfonate (MMS) | Sigma-Aldrich | 129925 | CAUTION: toxic |
| PALM setup | home-built | described in Uphoff et al.7 | |
| MATLAB | MathWorks | for data analysis and visualization | |
| Localization software | custom-written, available online | http://www.physics.ox.ac.uk/Users/kapanidis/Group/Main.Software.html | MATLAB and C++ software package that can be adapted for localization analysis. Cite Holden et al.13 if used in publication. |
| Tracking software | available online | http://physics.georgetown.edu/matlab/ | MATLAB implementation by Blair and Dufresne. Cite Crocker & Grier12 if used in publication. |
Riferimenti
- Friedberg, E. C. . DNA Repair and Mutagenesis. , (2006).
- Betzig, E., et al. Imaging intracellular fluorescent proteins at nanometer spatial resolution. Science. 313 (5793), 1642-1645 (2006).
- Mortensen, K. I., Churchman, L. S., Spudich, J. A., Flyvbjerg, H. Optimized localization analysis for single-molecule tracking and super-resolution microscopy. Nat. Methods. 7 (5), 377-381 (2010).
- Manley, S., et al. High-density mapping of single-molecule trajectories with photoactivated localization microscopy. Nat. Methods. 5 (2), 155-157 (2008).
- English, B. P., et al. Single-molecule investigations of the stringent response machinery in living bacterial cells. Proc. Natl. Acad. Sci. U.S.A. 108 (31), (2011).
- Subach, F. V., et al. Photoactivatable mCherry for high-resolution two-color fluorescence microscopy. Nat. Methods. 6 (2), 153-159 (2009).
- Uphoff, S., Reyes-Lamothe, R., Garza de Leon, F., Sherratt, D. J., Kapanidis, A. N. Single-molecule DNA repair in live bacteria. Proc. Natl. Acad. Sci. U.S.A. 110 (20), 8063-8068 (2013).
- Datsenko, K. A., Wanner, B. L. One-step inactivation of chromosomal genes in Escherichia coli K-12 using PCR products. Proc. Natl. Acad. Sci. U.S.A. 97 (12), 6640-6645 (2000).
- Reyes-Lamothe, R., Possoz, C., Danilova, O., Sherratt, D. J. Independent positioning and action of Escherichia coli replisomes in live cells. Cell. 133 (1), 90-102 (2008).
- Neidhardt, F. C., Bloch, P. L., Smith, D. F. Culture medium for enterobacteria. J. Bacteriol. 119 (3), 736-747 (1974).
- Tokunaga, M., Imamoto, N., Sakata-Sogawa, K. Highly inclined thin illumination enables clear single-molecule imaging in cells. Nat. Methods. 5 (2), 159-161 (2008).
- Crocker, J. C., Grier, D. G. Methods of digital video microscopy for colloidal studies. J. Colloid. Interface. Sci. 179 (1), 298-310 (1996).
- Holden, S. J., et al. Defining the limits of single molecule FRET resolution in TIRF microscopy. Biophys. J. 99 (9), 3102-3111 (2010).
- Holden, S. J., Uphoff, S., Kapanidis, A. N. DAOSTORM: an algorithm for high-density super-resolution microscopy. Nat. Methods. 8 (4), 279-280 (2011).
- Wieser, S., Schütz, G. J. Tracking single molecules in the live cell plasma membrane-Do's and Don't's. Methods. 46 (2), 131-140 (2008).
- Arnauld, S., Nicolas, B., Hervé, R., Didier, M. Multiple-target tracing (MTT) algorithm probes molecular dynamics at cell surface. Protoc. Exch. , (2008).
- Lippincott-Schwartz, J., Patterson, G. H. Photoactivatable fluorescent proteins for diffraction-limited and super-resolution imaging. Trends Cell Biol. (11), 555-565 (2009).
- Elmore, S., Müller, M., Vischer, N., Odijk, T., Woldringh, C. L. Single-particle tracking of oriC-GFP fluorescent spots during chromosome segregation in Escherichia coli. J. Struct. Biol. 151 (3), 275-287 (2005).
- Badrinarayanan, A., Reyes-Lamothe, R., Uphoff, S., Leake, M. C., Sherratt, D. J. In vivo architecture and action of bacterial Structural Maintenance of Chromosome proteins. Science. 338 (6106), 528-531 (2012).
Ristampe e Autorizzazioni
Richiedi autorizzazione per utilizzare il testo o le figure di questo articolo JoVE
Richiedi AutorizzazioneThis article has been published
Video Coming Soon
Personale delle biblioteche
Copyright © 2025 MyJoVE Corporation. Tutti i diritti riservati