Method Article
cd8+ t 细胞诱导体外肿瘤细胞凋亡的实时检测--研究肿瘤浸润髓内细胞的免疫抑制作用
* 这些作者具有相同的贡献
摘要
我们在这里描述了一个方案,研究细胞的预激活 cd8+ t 细胞对癌细胞通过实时显微镜检测凋亡癌细胞。该方案可以研究骨髓细胞诱导的 t 细胞抑制背后的机制, 并评估旨在通过阻断免疫抑制骨髓细胞来补充 t 细胞的化合物。
摘要
肿瘤 cd8+ t 细胞的肿瘤杀伤能力及其有效的肿瘤浸润是成功免疫疗法的关键因素。几项研究表明, 肿瘤浸润骨髓细胞 (如骨髓源性抑制细胞 (mdsc) 和肿瘤相关巨噬细胞 (tam)) 抑制肿瘤微环境中 cd8+ t 细胞的细胞毒性, 并针对这些调节髓质细胞可以改善免疫疗法。在这里, 我们提出了一个体外检测系统, 以评估免疫抑制免疫的影响, 单核 mdsc 和 tam的抗肿瘤能力的 cd8 + t 细胞。为此, 我们首先培养出具有抗 cdcd28 激活抗体的天真的脾脾 cd8 + t 细胞, 在存在或不存在抑制细胞的情况下激活抗体, 然后在含氟的情况下, 与靶向癌细胞共同培养预激活 t 细胞板状物-3 基板。实时荧光显微镜检测癌细胞底物中的荧光, 作为 t 细胞诱导肿瘤细胞凋亡的指标。在本试验中, 我们可以成功地检测 cd8+ t 细胞对肿瘤细胞凋亡的增加及其与 tam 或 mdsc 的预培养抑制。该功能分析可用于通过调节髓细胞研究 cd8+ t 细胞抑制机制, 并通过高通量筛选确定可药物的靶点。
引言
众所周知, cd8+ t 细胞在发挥其充分的细胞毒性时, 可以消除肿瘤细胞。t 细胞受体 (tcr) 激活后, cd8+ t 细胞增殖并分化为细胞毒性效应细胞。膨胀和激活的 cd8+ t 细胞分泌细胞毒性颗粒, 包括穿孔素和粒酶, 这些颗粒被转移到靶细胞中, 并引发各种裂解途径, 如酪蛋白酶-3 介导的凋亡1。cd8+ t 细胞还可以通过激活靶细胞上的受体诱导肿瘤细胞凋亡, 如肿瘤坏死因子α (tn-α) 受体、第一凋亡信号配体 (fasl) 或 tnf 相关凋亡诱导配体 (trail)。此外, 激活的 cd8+ t 细胞通过 fasl 受体1的向上调节分泌干扰素-γ (ifn-γ), 从而抑制肿瘤细胞增殖, 提高肿瘤细胞对 cd8+ t 细胞的敏感性。鉴于 cd8+ t 肿瘤杀伤能力的潜力, 已经制定了几种提高其细胞毒性的策略 (例如, 检查点抑制剂、癌症疫苗接种和嵌合抗原受体 (car) 表达 t 细胞的采用转移)并对某些类型的癌症2有显著的治疗作用.然而, 越来越多的证据表明, 肿瘤浸润免疫细胞, 如调节 t 细胞、骨髓源性抑制细胞 (mdsc) 和肿瘤相关巨噬细胞 (tam), 可以抑制 cd8+ t 细胞功能并限制疗效。3,4, 5.为了改善这种免疫治疗, 重要的是要了解免疫抑制细胞如何限制 cd8+ t 细胞毒性。cd8+ t 细胞抑制机制的确定以及克服其可吸毒的靶点, 将需要体外检测的开发和利用。
测量 cd8+ t 细胞毒性的金标准方法是铬释放法, 其中确定了由 cd8+ t 细胞裂解的目标细胞释放放射性探针 (51cr) 的方法 6.然而, 这种检测有几个缺点, 包括相对较低的灵敏度, 高背景, 无法检测早期凋亡事件, 危险的处置问题, 以及有限的兼容性与自动液体处理和检测, 以支持更高的吞吐量应用程序。另一种常见的方法是流式细胞仪分析, 其中检测目标肿瘤细胞的凋亡与环素 v 结合7。在本试验中, 除了目标细胞凋亡外, 还可以检测其他参数, 如使用碘化物 (pi) 或 7-氨基放线霉素 d (7-aad) 检测靶细胞死亡, 以及 cd107a 或 cd69 表达所指示的效应细胞激活。.然而, 与铬释放试验相比, 这种检测需要大量的抑制细胞。它还需要粘附靶细胞的分离和分解, 这可能会影响结果。事实上, 铬释放法或流式细胞测定并不常用于研究抑制细胞对 t 细胞功能的影响。相反, 通过稀释预加载到 t 细胞中的荧光染料 (如 cfse) 来测量 t 细胞增殖, 经常被用来评估抑制细胞对 cd8+ t 细胞功能的抑制作用。从培养的 t 细胞检测 ifn-γ的产生是评估抑制细胞对 t 细胞活化8,9的影响的另一种标准方法。然而, 这些检测的结果不一定与 cd8+ t 细胞的目标细胞杀伤能力相关。
我们在这里提出了一个替代功能的分析, 以评估抑制细胞, 特别是巨噬细胞转移性肿瘤, 对 cd8+ t 细胞的细胞毒性的影响。该方法通过检测荧光诱导的荧光检测肿瘤细胞凋亡, 确定 cd8 + t 细胞的细胞毒性, 在有或不存在抑制细胞的情况下, 在抗 cdn/cd28 激活抗体的存在下, 通过荧光检测肿瘤细胞凋亡基板 6使用自动延时显微镜 (图 1)。与其他方法相比, 该协议具有多种优点;它只需要少量的细胞, 能够以高灵敏度检测粘附肿瘤细胞死亡, 能够实时图像效应与目标相互作用, 并可进行高通量筛选。
在该方案中, 从小鼠转移性肿瘤中分离出的转移相关巨噬细胞 (mam) 及其祖细胞单核细胞 mdsc (m-mdsc) 被用作抑制细胞。在转移性乳腺癌的小鼠模型中, 明显的巨噬细胞群体为 fn! 80高ly6g-cd11b高Ly6G低积累在含有转移性肿瘤的肺中。这种巨噬细胞种群很少出现在正常的肺中, 因此被称为转移性巨噬细胞 (max)10。在这些小鼠模型中, 另一个骨髓细胞群, 定义为 fn-cd11b 高 Ly6G高,也主要积累在转移性肺, 在那里它产生了 mom11.根据其特性, cd11b 高 ly6c高mam祖细胞可能代表 m-mdsc12。
研究方案
所有涉及老鼠的程序都是根据联合王国内政部的许可 (p526c60b3) 进行的。有关商业试剂和设备的信息列于材料表中。
1. 在细胞核中表达红色荧光蛋白的靶细胞的制备
- 从适当的来源获取目标小鼠癌细胞系。
注: 在本协议中, 使用了 e0771 小鼠乳腺肿瘤细胞 (e0771-g)13的高度转移衍生物。父母 e0771 细胞来源于 c57bl6 小鼠14。 - 用 dulbecco 的改良鹰培养基 (dmem) 解冻并维护一个 e0771-g 细胞小瓶, 其中包括在37°c、95% 湿度和 5% co 2 的细胞培养孵化器中的 10% (v/v) 胎儿牛血清 (fbs).
注: 应确认细胞对支原体呈阴性反应。为此, 如上文所述 (在没有抗生素和抗菌药物的情况下), 将 e0771 细胞 (或待检测的细胞) 培养 2-3天, 收集500μl 培养基。以 12, 419 x克的速度离心培养基, 以消除细胞碎片, 并将上清液转移到新的管中。根据制造商的指示, 使用市售支原体检测试剂盒 (请参阅材料表) 和 pcr15确定支原体污染。 - 种子 5 x10 3 e0771-g 细胞每口井进入12孔板, 并在37°c、95% 湿度和 5% co 2 的孵化器中用 10% (v/v) 隔夜培养 fbs-dmem 细胞.
注: 如果目标细胞的增殖率较低 (种群加倍时间大于 36小时), 细胞的数量可增加到 1 x 104. - 将介质替换为1毫升的 10% (v/v) fbs-dmem, 包括10μgml 聚乙烯, 并添加25μl 的慢病毒颗粒 (1 x10 6 tuml), 编码核限制的红色荧光蛋白 (mkate2, 见材料表)。
- 在37°c、95% 湿度和 5% co2的孵化器中培养细胞24小时。
- 将培养基替换为 10% (v/v) fbs-dmem, 并在37°c、95% 湿度和 5% co 2 的孵化器中培养24-48 的细胞.
- 当细胞开始表达红色荧光蛋白时, 将培养基替换为 10% (v/v) fbs-dmem, 包括 1μgl puromycin, 并培养细胞, 直到它们融合了80-90%。
注: 在靶细胞类型之间, puromycin 的浓度会有所不同, 应使用未经转染的细胞进行优化。 - 将存活的细胞亚培养为1-3 个通道, 用 10% (v/v) fbs-dmem (包括 1μgl puromycin), 并在液氮气相储存系统中冷冻储备储存, 直至使用。
2. 从小鼠肿瘤中分离抑制细胞
注: 在本协议中, 抑制细胞 (即mum 和 m-mdpc) 与 e0771-g 细胞建立的含有转移瘤的肺分离。组织离解和细胞分选的条件应优化, 以分离不同组织中的细胞。
- 将 1 x 106个癌细胞 (e0771-g) 注入异基因 (c57bl6) 的尾静脉, 雌性, 7-10 大的小鼠。
- 14天后, 分离含有转移性肿瘤的肺, 并准备单细胞悬浮液从灌注肺通过酶消化, 如前面所述 11.
注: 在这个方案中, 四只小鼠被注射癌细胞, 他们的转移性肺被结合, 以获得足够的抑制细胞。 - 用抗小鼠 ccmu/cd32 抗体在冰上培养30分钟的单细胞悬浮液, 并用荧光抗体在冰上涂抹 30分钟, 用荧光抗体涂抹在 cd45、f480、cd11b、ly6c 和 Ly6C (参见材料表) 上再涂抹 30分钟10,11,13岁
- 用含有 2% (w/v) 的 pbs 1 ml 清洗一次染色细胞, 用含有 2% (w/v) 的 500–1000μl pbs 重新悬浮细胞颗粒。
- 添加3μm 的 dapi, 并对 m-mdsc 进行排序(dap/cda+ f481\80+ly6g/cd11b高Ly6G) 和 mam (dapi–cd45+ fn2\ 80 + ly6g-cd11b 高ly6c低) 使用细胞分拣器 (补充图 1)。
注: ly6c 电平的阈值是根据常驻肺泡巨噬细胞 (rmac)的阈值确定的, 以区分制造 (ly6c 低) 和 m-mdcs (ly6c 高) 为基础。通过流式细胞仪测量分类细胞的纯度, 预期纯度超过90%。 - 用含有 20% (v/v) fbs 的 400μl dmem 重新移植分类细胞, 1% (v/v) penicilin/streptomin, 2 mm l-谷氨酰胺, 1% (v/v) 非必要氨基酸, 1 mm 丙酮酸钠, 和 50 nm 2-甲基甲醇 (称为 enriched-dmem, e-dmem)。
- 使用 trypan 蓝色排除方法16计数活单元的数量, 并使用 e-dmem 调整到 2 x10 6 cells/mL。
- 将细胞放在冰上, 直到使用。
3. 从小鼠脾脏中分离 cd8 + t 细胞
- 将脾脏从与目标癌细胞系 (即本协议中的 c57bl6 小鼠) 合酶上分离出来, 如下所示:
- 用二氧化碳吸入使动物安乐死 。
- 将动物放在干净的解剖板上, 用70% 的乙醇擦拭皮肤。
- 用剪刀割伤腹部皮肤以暴露脾脏
- 分离位于胃低度的脾脏, 并将其放入含有5毫升冰凉 pbs 的管中。
- 使用无菌5毫升注射器的内部柱塞, 在50毫升管上设置的100μm 细胞过滤器上研磨脾脏。
- 使用 pbs 的总10毫升将细胞通过过滤器。
- 以 337 x g 离心细胞悬浮液 5分钟 , 并吸入上清液。
- 在含有 2 mL edta 和 0.5% (w/v) bsa (运行缓冲液) 的 pbs 1 毫升中重新使用细胞颗粒, 并通过40μm 的细胞过滤器进行过滤。
- 计算活动单元格号, 并使用运行缓冲区调整为 1 x 108 cellse/ml。
注: 保留少量的细胞作为浓缩前的样品, 以便进行纯度检查。 - 使用负选择试剂盒和磁性分选器 (请参阅材料表) 丰富 cd8+ t 细胞。
- 将脾细胞的 1 x 10 8 (1 ml) 转移到5毫升聚苯乙烯圆底管中。
- 加入50μl 的生物素化抗体, 在室温下孵育10分钟。
- 加入125μl 链霉素共轭磁珠, 在室温下孵育5分钟。
- 加入1.325 毫升的运行缓冲器, 然后通过移液轻轻混合。
- 将管子放入磁铁中, 在室温下孵育2.5 分钟。
- 拿起磁铁, 将浓缩的细胞悬浮液倒入新的试管中。
- 用200μl 的 e-dmem (即,含有 20% (v/v) fbs、1% (v/v) penicilin/streptomin、2 mm l-谷氨酰胺、1% (v/v) 非必要氨基酸、1 mm 丙酮酸钠和 50 nm 2-氨基乙醇) 的 dmem。
- 用 e-dmem 计数活细胞的数量, 并调整到 2 x10 6 细胞。将细胞保持在 37°c, 放在二氧化碳孵化器中, 直到使用。
注: 保留少量的细胞作为浓缩后的样品, 以便进行纯度检查。 - 流式细胞仪测定 cd8+ t 细胞的纯度, 如下所示:
- 从浓缩前 (第3.6 步) 或浓缩后 (第3.9 步) 样品中提取 1 x 104个电池, 并使用运行缓冲液将每个细胞的总体积调整为100μl。
- 用抗小鼠 ccmu/cd32 抗体在冰上培养单个细胞悬浮液 30分钟, 用荧光抗体在 cd45、cd3、cd4 和 cd8 (参见材料表) 染色30分钟。
- 用含有 2% (w/v) bsa 的 500μl pbs 清洗染色细胞, 并用含有 2% (w/v) bsa 的 500–1000μl pbs 重新悬浮细胞颗粒。
- 加入3μm 的 dapi, 并确定 cd3+cd4–cd8+细胞在 cd45+细胞总数中的百分比.
4. 分离 cd8+ t 细胞的活化和扩展
- alquot 1 x 10 5 细胞, 每 50μl cd8+ t 细胞 (在步骤3.9 中制备) 进入 u 底部96孔板的井。
- 每 50μl抑制细胞 (在步骤2.7 中制备) 或 50μl e-dmem 在井中添加 1个 x 10 5个细胞。
- 制备活化介质, 包括 e-dmem、4 x 104 uml 集落刺激因子 1 (csf-1)、240 uml 白细胞介素-2 (il-2)、8μgml 抗小鼠 cd3 抗体和16μgml 抗小鼠 cd28 抗体。
注: csf-1 不是 t 细胞激活所必需的, 但对于此协议中抑制细胞 (即 mam 和 m-mdsc) 的生存至关重要。因此, 它被保留在 t 细胞与靶向癌细胞的共培养中, 以保持培养条件的一致性。由于 csf-1 在所有组织中都含有纳米摩尔浓度, 并且在体内是单环巨噬细胞活力所必需的,因此这是这些细胞的生理环境。 - 加入50μl 的活化介质 (步骤 4.3) 和50μl 的 e-dmem, 并对其进行检测或不添加试剂。
- 将板材放入37°c、95% 湿度和 5% co2的孵化器中, 培养细胞4天。
5. 与预激活的 cd8+ t 细胞共培养靶细胞的建立
- 在适合显微镜的平底96孔板的井中加入30μl 的1:100 稀释生长因子--减少可溶性基底膜基质 (材料表), 并在37°c 下在 co2 孵化器中孵育至少 1小时.
- 准备目标细胞 (即 e0771-g 细胞表示核限制的红色荧光蛋白)。
- 在室温下用0.05% 的胰蛋白酶 edta 对靶细胞进行1年的培养, 并通过温和的移液法收获靶细胞。
- 加入9毫升的 dmem, 包括 10% (v/v) fbs, 离心细胞悬浮在 337 x克5分钟。
- 用500μl 的 e-dmem 重新悬浮细胞, 并计算活细胞的数量。
注: 在计算目标细胞之前, 通过40μm 的电池过滤器对细胞进行过滤, 以产生单个细胞悬浮液。 - 通过添加 e-dmem,将密度调整为 2 x 10 4 cellsl (= 每 50μl1 x 10 3细胞), 并将细胞保持在冰上。
- 准备效应细胞 (即预激活的 cd8+ t 单元)。
- 通过移液将96孔板 (步骤 4.5) 中的细胞彻底重新移植, 并将浮动的 cd8+ t 细胞转移到新的 1.5 ml 管中。
注: mam 和 m-mdcs 紧紧地粘附在井上, 不会被移液器分离。 - 为了收集剩余的细胞, 在井中加入200μl 的 pbs, 并在5.3.1 的步骤转移到试管中。
- 用 e-dmem 1 毫升 (337 x g 的离心机) 清洗细胞 5分钟, 吸气并丢弃上清液), 并用 100μl e-dmem 重新悬浮细胞。
- 计数活细胞的数量 (使用色氨酸蓝色排除方法), 将密度调整为 1.6 x10 5 细胞 (= 4 x10 3 cellss\ 25μl), 并将细胞保持在冰上。
- 通过移液将96孔板 (步骤 4.5) 中的细胞彻底重新移植, 并将浮动的 cd8+ t 细胞转移到新的 1.5 ml 管中。
- 在步骤5.1 中从板的每口井中吸气基底膜基质。
- 在每口井中加入 1 x 10 3 细胞50μl 的靶细胞 (步骤 5.2.4), 搅拌均匀。
注: 为避免介质蒸发引起的边缘效应, 应仅使用96井板的内部60口井进行分析。 - 加入 25μl e-dmem, 包括 4 x10 3 ueml il-2 和10μm 荧光型 caspase-3 基板 (参见材料表)。
- 将 4 x10 3 细胞/cd8 + t 细胞 (步骤 5.3.4) 加入适当的井中, 搅拌良好。
注: 井中存在过多的细胞会增加分析的难度。在该模型中, 4: 1 效应因子: 目标比 (总细胞数 5 x10 3 cells/well well) 是最优的, 但81:1 的比率是次优的。需要包含以下四个控制的井有助于数据分析: 用于共培养井 (1 x10 3 cells井) 的密度的目标细胞, 在培养基中与卡斯帕西-3 底物和不含有卡斯帕西-3 底物的目标细胞, 效应细胞 (1 x10 3 细胞)在介质中具有和不带卡斯帕西-3 基板。 - 在所有空井 (特别是板周围的井) 中加入200μl 的 pbs 或无菌水, 以减少实验井中介质的蒸发。
- 将印版设置为延时荧光显微镜, 该显微镜保持在37°c、95% 湿度和 5% co2。
注: 以交叉模式摇晃板, 使所有细胞均匀分布, 然后让板在室温下保持在平坦表面上 10-20, 然后转移到孵化器。
6. 细胞成像
注: 详细的图像采集设置将因使用的显微镜和荧光而异;为了获得最佳结果, 应采用以下一般采集参数。
- 在显微镜下使用适当的自动对焦例程, 获取覆盖96孔板每个实验井总表面积至少25% 的图像。
- 设置显微镜以获取相对比的图像, 以及适用于核限制红色荧光蛋白 (mkate2) 的荧光通道和适用于绿色荧光活性 case-3 基板的荧光通道 ((图 2)。
- 每1至3小时捕获实验井中的图像, 时间至少为72小时。
7. 使用图像分析软件进行图像分析
注: 详细的图像分析设置将因所使用的软件而异 (请参阅材料表);应采用以下一般分析程序, 以获得最佳结果。
- 确定是否需要光谱非混合来分离目标细胞核发出的荧光信号和凋亡核发出的荧光信号, 并在必要时建立分析协议以实现这一目标;
- 查看在绿色荧光通道中没有通道中没有通道中的 c s 皮亚斯-3 基板的介质中只包含目标细胞 (mkate2-tat 标记) 的控制井。
- 观察红色原子核 (原子核呈绿色) 是否发出绿色荧光
- 如果绿色荧光在原子核中明显可见, 请访问成像软件中的光谱解混合控制, 并增加从绿色中去除的红色百分比, 直到绿色信号消失 (根据我们的经验, 这通常是6-7 的明亮 mkate2荧光)。
- 如果绿色荧光在任何原子核中都不明显, 则不需要光谱解混。
注: 这种光谱不混合校正试验也可以使用含有卡斯帕西-3 基板介质中的目标细胞井进行。然而, 目标细胞的自发凋亡水平通常很低 (不到 10%), 导致一些靶向细胞核由于激活的酪蛋白酶-3 而不是荧光出血而发出绿色荧光。在这些条件下, 当绿色荧光在所有原子核中都是明显的时, 真正的荧光出血是明显的。 - 查看在红色和绿色通道中没有通道中的无通道中的基板中仅含有效应细胞 (未标记的细胞核) 的控制井。
- 观察原子核是否发出绿色或红色荧光。如果红色和绿色荧光在各自的渠道是不明显的没有光谱解开是必要的。
注: 根据我们的经验, 鼠标 cd8+ t 细胞不是自动荧光的, 因此这些细胞不需要光谱解混。这种光谱解混校正试验也可以使用含有酪醚-3 基板的介质中的效应电池井进行。然而, 通常有一定程度的自发凋亡, 导致效应细胞核发出绿色荧光, 由于激活的酪蛋白-3。在这些条件下, 当绿色荧光在所有原子核中都是明显的时, 真正的荧光出血是明显的。
- 使用荧光背景减法 (例如 tophat), 并为样本提供相关参数, 以解决两个荧光通道中的荧光物体。
注: 在绿色通道中, 我们使用了 tophat (原子核半径 = 10.0μm, 绿色荧光阈值 = 0.7个绿色校准单元)。在红色通道中, 我们使用了 tophat (原子核半径 = 10.0μm, 红色荧光阈值 = 0.5 红色校准单元)。 - 使用适当的边缘分裂参数来解决单个荧光核。
注: 我们没有在绿色或红色荧光通道中使用边缘分裂。 - 使用仅包含目标细胞的井 (带有酪蛋白酶底物) 的红色通道中的图像来确定目标核的最小大小。
注: 我们使用了 80μm2。 - 使用仅含有效应细胞的井 (含酪蛋白基板) 的绿色通道中的图像来确定凋亡效应核的平均大小。
注: 在这些实验中, 单凋亡效应核的范围为40–80微米2 。 - 建立一个分析程序, 使用适当的最小尺寸限制 (例如, 最小面积、最小直径) 来计算荧光靶核的数量 (图 2, 目标检测掩码)。
注: 我们使用了最小核子面积 (红色荧光) = 80μm2。 - 建立一个分析程序来计算凋亡细胞核的数量, 这些细胞核的大小大于凋亡效应核的平均大小 (图 2, 大小受限凋亡掩码)。
注: 尺寸过滤器设置为 80μm2 (即只计算出大于此尺寸的绿色荧光区域, 因此不包括单个凋亡效应核)。利用这些参数可以提高分析的准确性, 计算凋亡靶核, 虽然一些凋亡效应细胞的集合体可以计数。 - 建立一个分析程序, 通过计数细胞核来计算凋亡靶细胞的数量, 其中红色荧光信号 (从步骤 7.6) 和大小限制绿色荧光信号 (从步骤 7.7) 显著地共定位 (图2, r重叠掩码)。
注: 适当的联合定位范围可能从目标核平均大小的30% 到100% 不等。重叠尺寸过滤器设置为 40μm 2。 - 确定每个实验条件下每个实验过程中凋亡靶核的数量以及井内的靶细胞核数。
8. 使用计算和绘图软件进行数据分析
- 绘制实验井在整个时间过程中在步骤7.9 中获得的凋亡靶核数。如果在每个实验条件下使用多个井, 则以图形方式将结果显示为均值±标准偏差。
- 通过将每口井中的凋亡细胞数量除以每个时间点每口井中的目标细胞数量来计算目标细胞种群的凋亡分数。
- 绘制步骤8.2 中获得的结果图。
- 确定每个曲线的曲线下区域 (auc)。如果需要, 也可以确定每条曲线的种群凋亡分数的峰值以及峰值发生的时间点。使用适当的统计测试确定为实验条件确定的 auc 是否有显著差异。
注: 未配对的 t 检验与 welch 的校正应用于 auc 的结果。
结果
该方法基于简单的靶向癌细胞与效应因子 cd8 + t 细胞的联合培养, 这些细胞在存在抗 cdn/cd28 激活抗体的情况下, 通过或不预先培养了抑制细胞。它检测 cd8+ t 细胞诱导的癌细胞凋亡随着时间的推移, 随着时间的推移, 共培养, 从而能够评估抑制细胞对 cd8+ t 细胞的细胞毒性的影响。
通常情况下, 癌细胞在激活核靶向酪蛋白酶生物传感器后, 在没有抑制细胞的情况下与抗体预激活的 caspase+ t 细胞接触时, 其细胞核中的绿色荧光增加 (图 3;补充电影 1)。细胞凋亡启动后, 可在至少15小时内检测出酪蛋白酶底物的绿色荧光。随着时间的推移, 也观察到了效应细胞 cd8+ t 细胞的一些自发凋亡, 即使这些细胞是在分离中培养的 (图 4;补充电影 2)。然而, cd8+ t 细胞的细胞核大小小于癌细胞, 因此, 通过大小限制图像分析方法, 可以将凋亡的 "效应" 细胞排除在凋亡的 "靶标" 细胞计数之外 (图 2和图 4)。虽然一些靶向癌细胞在没有绿色荧光的情况下呈小圆形, 但这并不影响分析, 因为这些细胞正在经历有丝分裂而不是凋亡 (补充电影 3), 因此被排除在凋亡的 "目标" 之外单元格计数由一个红色/绿色重叠掩码 (补充图 2)。即使在单一培养中, 也偶尔会发现靶向癌细胞的自发凋亡 (补充电影 3)。然而, 靶向癌细胞与预激活的 cd8+ t 细胞的联合培养使肿瘤细胞凋亡高于肿瘤细胞单培养细胞的自发性凋亡水平 (图 4)。一般来说, 当使用目标癌细胞与效应细胞的最佳比率时, 可以观察到凋亡靶细胞数量的峰值 (图 5a)。当数据表示为目标细胞群的凋亡分数时, 这个峰值就更加明显 (图 5b)。在本实验中, 靶向肿瘤细胞的基础凋亡在24小时达到高峰 (凋亡分数 = 0.08), 但 cd8+ t 细胞诱导的凋亡在17小时时达到最高水平 (凋亡分数 = 0.08)。
我们进一步发现, 用 mum 或 m-mdsc 预先培养的 cd8+ t 细胞可以与靶癌细胞接触, 但这种接触似乎导致癌细胞凋亡的情况比 cd8+ t 细胞在未激活时的情况下发生的情况下减少。抑制细胞 (图 3和图 4;补充电影4和补充电影 5)。虽然 cd8+ t 细胞的预培养与骨髓细胞的偶尔诱导癌细胞凋亡, 但也有一些未被刺激的靶癌细胞在细胞的过程中经历细胞凋亡的情况。实验 (补充电影 6)。与这些发现一致的是, 用 cd8+ t 细胞预先培养的髓质细胞 (mdsc-e 在23小时时凋亡分数 = 0.38, mam-e 在20小时时 0.25) 培养的凋亡癌细胞的峰值分数明显低于癌细胞的峰值分数用 cd8+ t 细胞培养, 而这些细胞不是用抑制细胞预先培养的 (图 6)。

图 1: 显示实验过程的方案.用抗 cd松/cd28激活抗体培养出具有或不具有转移相关巨噬细胞 (max) 或单核细胞骨髓源性抑制细胞 (m-mdsc) 的肠道脾脏。4天后, 在氟生成型 caspase-3 底物的情况下, 收集漂浮的 cd8+ t 细胞, 并与靶向癌细胞共同培养。在实时荧光显微镜下检测凋亡癌细胞。所显示的图像是使用实时细胞成像平台获得的 (请参阅材料表)。请点击这里查看此图的较大版本.
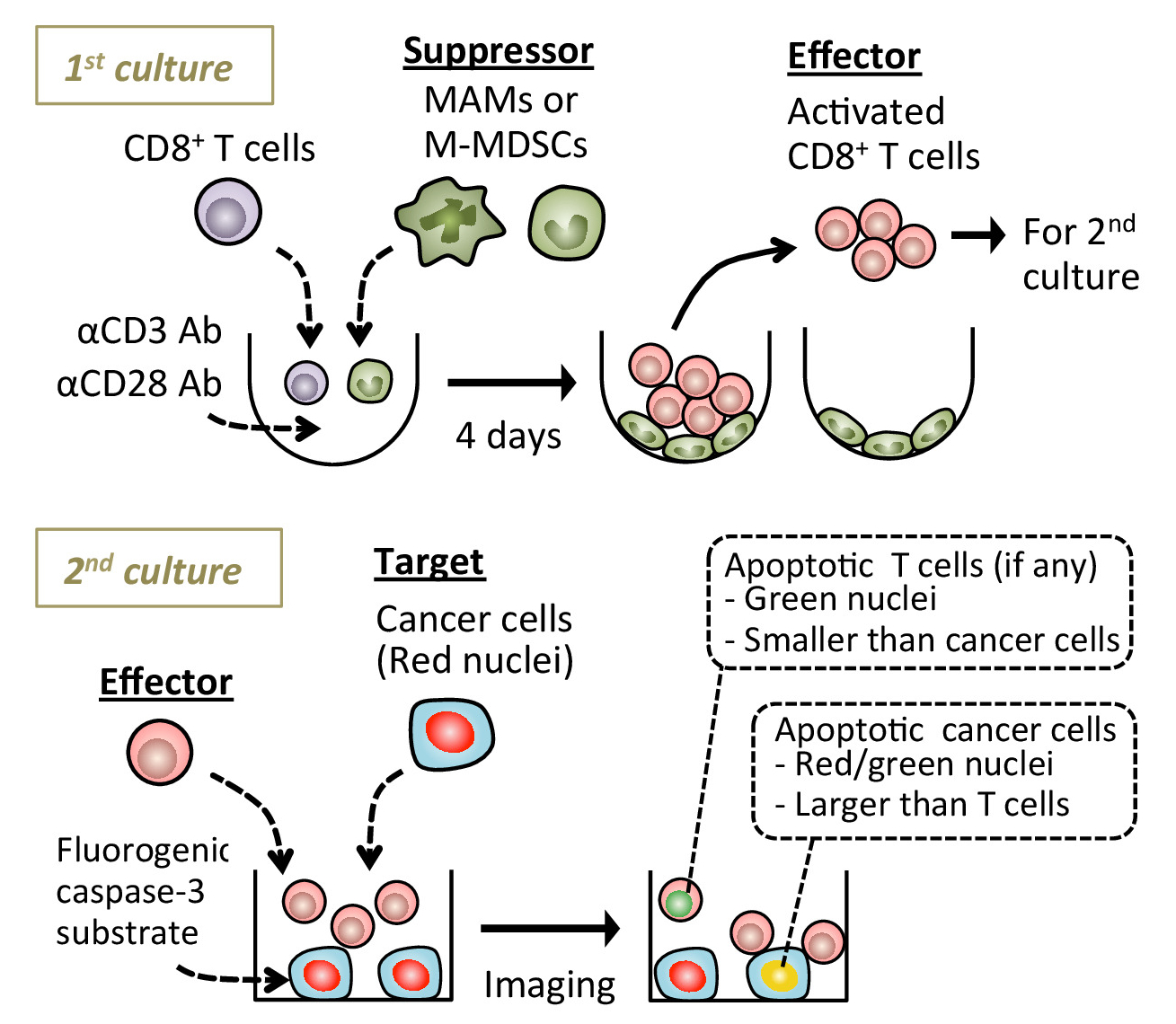{kind=link}

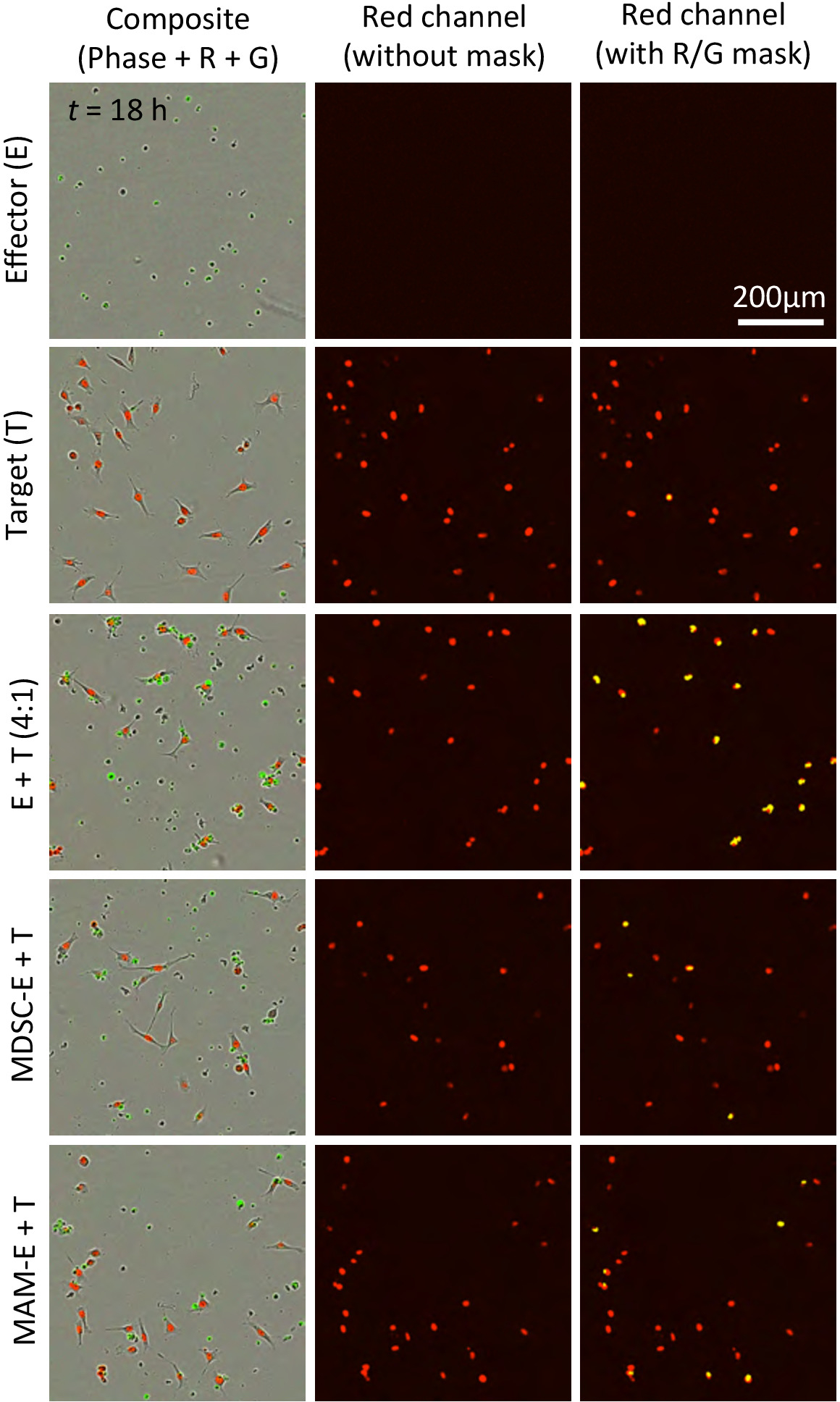
图 2: 不同于凋亡效应细胞的凋亡靶细胞的鉴定.顶行: 在红色通道中采集图像, 可以通过目标检测掩码 (粉红色分析掩码) 识别目标细胞核。中间行: 在绿色通道中获得的图像表示凋亡效应和目标细胞。大小限制凋亡掩模 (分体分析掩模; 大于 80μm2) 允许将单个凋亡效应细胞排除在分析之外。底部行: 复合图像合并红色和绿色通道与相位对比度图像 (左) 或绿色重叠掩码 (右)。用红色荧光 (黄色分析掩模) 识别共定位、大小受限的绿色荧光, 通过排除凋亡效应细胞 (白色) 的集合体, 可以更准确地检测凋亡目标核 (黄色箭头)箭头)。请点击这里查看此图的较大版本.
{kind=link}

图 3: cd8+ t 细胞与癌细胞之间的相互作用。目标 e0771-g _ nlr 细胞 (t) 的具有代表性的延时电影中的结果与效应 cd8 + t 细胞 (e) 在4:1 有效目标比的情况下共同培养。mdsc-e 和 mam-e 分别表示用 m-mdsc 和 mam 预先培养的效应细胞。显示复合图像 (包括来自相位对比度、红色和绿色通道的图像)。箭头正在通过不同的字段和时间点跟踪相同的单元格。黄色箭头: 与效应器相关并经历细胞凋亡的目标, 白色箭头: 与效应器相关联但不发生凋亡的靶标。请点击这里查看此图的较大版本.
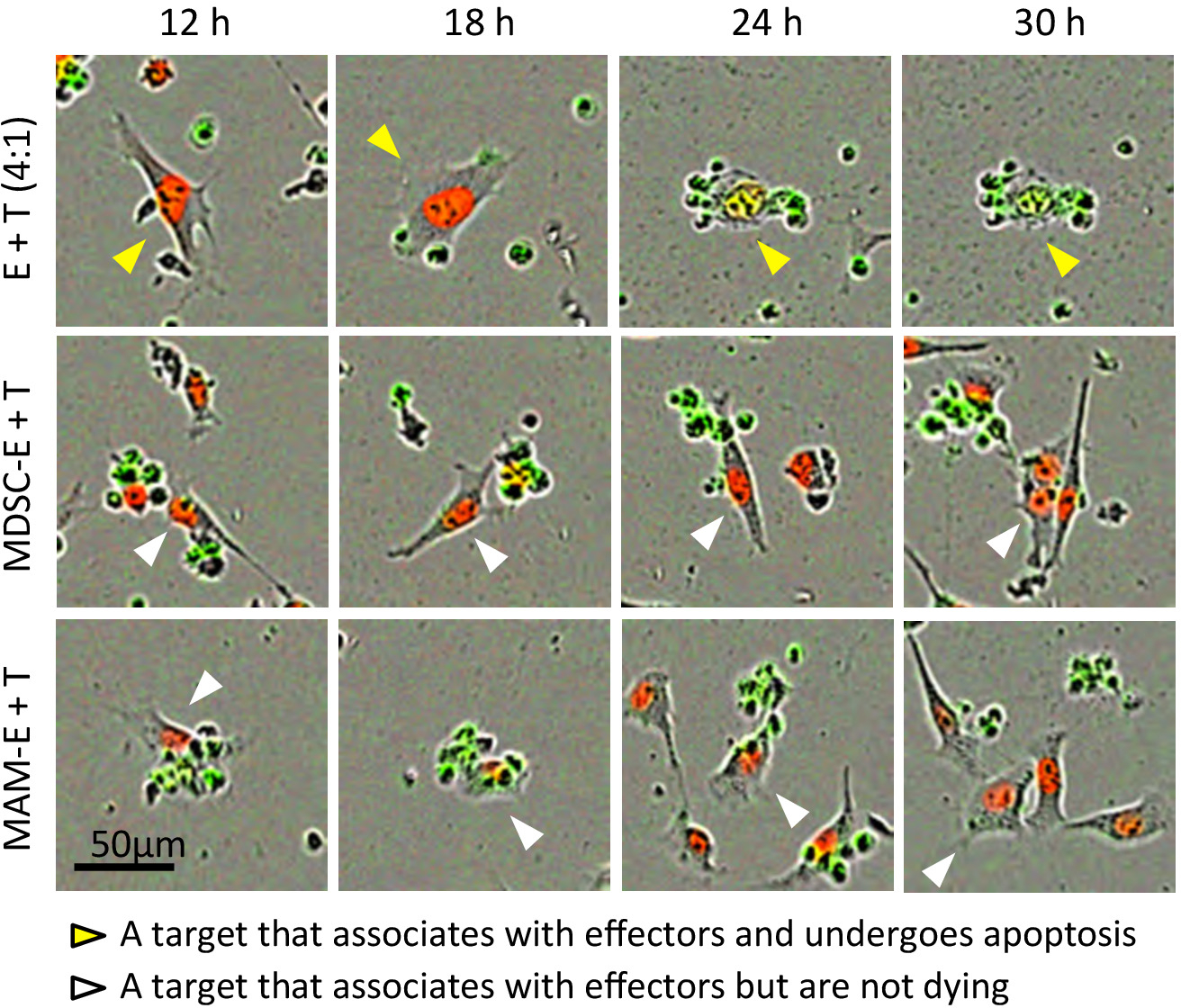{kind=link}

图 4: 凋亡癌细胞的检测.在成像后18小时从延时电影中提取的代表性场。显示了复合图像 (左; 相位对比度、红色和绿色通道) 和红色通道中没有 (中间) 或带有 (右) 红绿色重叠掩码 (黄色) 的图像。右列中的黄点代表凋亡癌细胞。请点击这里查看此图的较大版本.
{kind=link}

图 5:cd8+ t 细胞诱导的癌细胞凋亡。(a) 在不同效应与目标比率下, 用效应因子 c8+ t 细胞培养的凋亡癌细胞数量 (e:t) (b) 目标细胞群的凋亡分数。数据是指±sd. 曲线下的平均面积 (auc) 也显示。采用未配对 t 检验与韦尔奇修正的方法对 auc 进行了分析。* 与 e:tcet0x:1 相比, P<0.0001。请点击这里查看此图的较大版本.
{kind=link}

图 6: 肿瘤浸润髓质细胞对 cd8+ t 细胞细胞毒性的影响。(a) 用 c8+ t 细胞培养的凋亡癌细胞 (靶点: t) 的数量 (效应因子: e), 4:1 之比为4:1。cd8+ t 细胞是在没有 (黑圈) 或存在单核细胞-骨髓源性抑制细胞 (mdc-e: 蓝囊) 或转移相关巨噬细胞 (mam-e: 红圈) 的情况下进行的。数据是目标细胞群的±sd. (b) 凋亡分数。数据是手段±sdd. 平均 auc 也显示。采用未配对 t 检验与韦尔奇修正的方法对 auc 进行了分析。* P<0.0001 仅与 t 相比, #P<0.0001 与 e + t 相比, 请点击这里查看此图的更大版本.
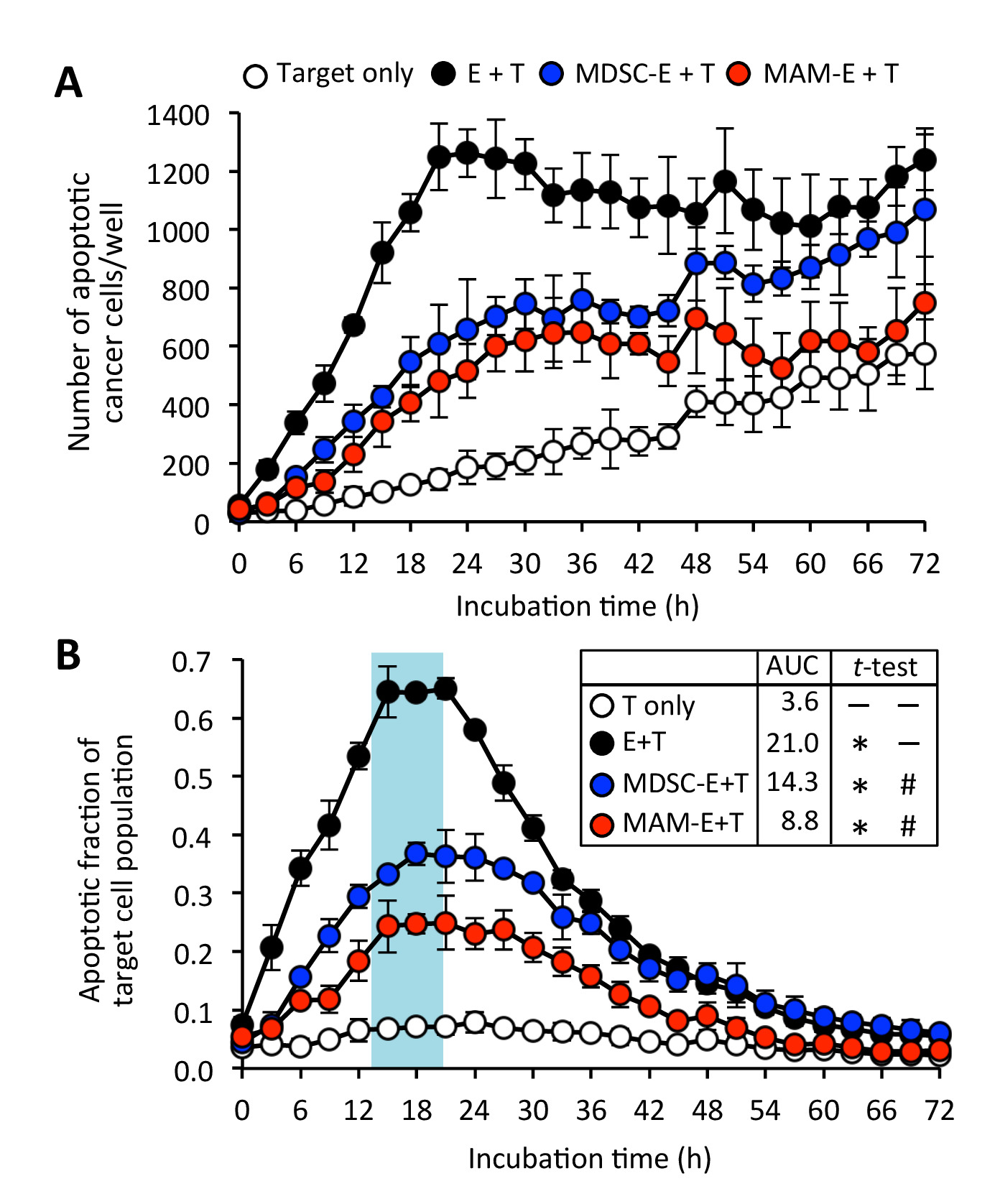{kind=link}
补充图1。从转移性肺中分离抑制细胞的称重策略。(a) 具有代表性的点图, 用于分离单核细胞骨髓源性抑制细胞 (m-mdcs) 和与转移有关的巨噬细胞 (max)。ly6c 电平区分 um (ly6c低) 和 m-mdcs (ly6c高) 的阈值是基于常驻肺泡巨噬细胞 (rmac) 的阈值。(b) 已分类的 m-mdsc (cd45+ly6g-cd11b + Ly6G 高) 和 mam(cd45+ly6g-cd11b + Ly6G低) 的纯度.请点击此处下载此文件.
补充图2。有代表性的有丝分裂靶细胞的图像。目标 e0771-g _ nlr 细胞单核培养的代表性延时电影中的史迪尔斯。顶部: 合成图像, 包括来自相位对比度、红色和绿色通道的图像。底部: 带有红色/绿色重叠掩码的复合图像 (红色和绿色通道)。请点击此处下载此文件.
补充图3。肿瘤浸润髓系细胞对 cd8+ t 细胞增殖的影响。(a) 代表性直方图, 显示 cfse 荧光标记在 cd8+ t 细胞中的稀释。如协议3所述, 对天真脾 cd8+ t 细胞进行了分离, 并在37°c 下标记了 5μm cfse 15分钟。标记 t 细胞是在 il-2 和抗 cdncd28 激活抗体的情况下培养的, 如第4议定书所述。4天后, 流式细胞仪检测 t 细胞中的绿色荧光。(b) 按上述方法计算的 cd8+ t 细胞的划分指数17。数据是指±sem. * P<0.01 与对照相比, #P<0.05 与α-cdcd28 ab 相比. 请点击这里下载此文件.
补充电影1。图3和图4的电影;E + T.请点击此处下载此文件.
补充电影2。图3和图4的电影;效果器 (e)。请点击此处下载此文件.
补充电影3。图3和图4的电影;目标 (t)。请点击此处下载此文件.
补充电影4。图3和图4的电影;MDSC-E + T.请点击此处下载此文件.
补充电影5。图3和图4的电影;MAM-E + T.请点击此处下载此文件.
补充电影6。图3和图4的电影;mam-e + t (扩散).请点击此处下载此文件.
讨论
该方法基于两个独立的共培养步骤: 将 cd8 + t细胞与潜在的抑制细胞共同培养, 以及将 "预处理" cd8+ t 细胞与靶向肿瘤细胞共同培养 (图 1)。第一个共培养步骤与 cd8+ t 细胞增殖检测非常相似, 该检测通常用于确定抑制细胞对 cd8+ t 细胞功能的影响。然而, t 细胞增殖并不总是与它们的细胞毒性相关。例如, 我们发现, 在 ccdg28 激活抗体的情况下, 与 m-mdsc 或 max 共培养增加而不是减少 cd8+ t 细胞的增殖 (补充图 3), 而这些预先设定的 cd8+t 细胞对靶向癌细胞的细胞毒性降低 (图 4,图 5,图 6)。这些结果突出了功能活性评价的重要性, 证明了目标癌细胞凋亡, 提供了这种 cd8+ t 细胞毒性检测。
在本试验中, 我们已经确定 cd8+ t 细胞需要大约15小时的共培养, 以诱导最大凋亡的 e0771-g 小鼠乳腺肿瘤细胞 (图 5)。这种延迟可能是由于效应细胞与目标的初始接触与伴随的免疫突触形成之间的滞后, 以及通过激活 caspase-3 来测量的诱导目标中的凋亡信号所需的时间 (补充电影 1)).我们还发现, 24小时后, 凋亡肿瘤细胞的数量达到了一个高原, 这可能是由于 t 细胞消除了目标, 或者死细胞的荧光信号丢失。这种识别细胞凋亡峰值时间的能力是此检测的主要优势之一, 因为确定最佳时间点对于不同条件之间的适当比较非常重要。例如, 在我们的例子中, 对照 cd8+ t 细胞和 mdsc/mam 教育的 cd8 + t 细胞在15-18小时时的细胞毒性差异要大得多, 而不是 72小时 (图 5), 因此使用72小时潜伏期的端点实验会产生误导的结果。
这种方法还可以实现实时效应与目标细胞相互作用的可视化, 从而更深入地了解 cd8+ t 细胞预培养的 cd8 + t 细胞有限的细胞毒性机制。例如, 我们观察到 cd8+ t 细胞与 m-mdsc 或 mum 预先孵育时遇到并与目标肿瘤细胞相互作用, 但并不总是诱导细胞凋亡 (补充电影 4, 补充电影 5, 补充电影 6)。虽然我们没有量化这一事件, 它将是可行和有趣的量化和比较的比例, 以及它们的相互作用时间与细胞凋亡诱导。另一个主要优点是, 这种方法需要少量的细胞 (例如, 每个井 1 x10 3 和效应单元 4 x10 3) 。事实上, 如果需要, 该协议可以进一步小型化, 适用于384孔板格式。因此, 该检测方法适用于高通量筛选和细胞数量有限的实验, 例如使用从体内或体内样品中提取的珍贵细胞进行体外检测。
另一方面, 目前检测的一个局限性是, 在某些情况下存在大量的死亡效应细胞。为了提高目标癌细胞凋亡与效应细胞 cd8+ t 细胞凋亡的准确性, 对靶细胞的细胞核进行标记, 并应用细胞核大小限制 (排除效应细胞) 进行数据分析。(图 2)。然而, 在某些情况下, (绿色) 凋亡 cd8+ t 细胞的集合体覆盖到非凋亡靶向癌细胞上, 这可能会混淆结果。在与目标肿瘤细胞共培养之前, 如果有足够数量的效应细胞, 则可以通过在效应细胞上使用死细胞去除柱来缓解这一限制。对于更复杂的显微镜系统, 也可以通过标记效应 cd8+ t 细胞与不同于目标细胞核和荧光性 caspase-3 基板的荧光体来减少假阳性信号。
到目前为止, 该方案已被用于研究 cd8+ t 细胞的抗原非特异性活化。虽然 mdsc 和 tam 在肿瘤微环境中通过抗原非特异性机制抑制 t 细胞功能, 但外周淋巴组织中的 mdsc 以抗原特异性的方式抑制 t 细胞的反应18.为了研究这种细胞类型的免疫抑制功能, 常用的方法是利用 ot-1 转基因小鼠 cd8+ t 细胞进行体外增殖检测。在本试验中, ot-1 t 细胞 (表达卵白蛋白 (ova) 特异性 t 细胞受体) 与抑制 mdsc 在 ova 肽存在的情况下共同培养, 这适用于我们的细胞毒性试验中的第一次培养 (即 t细胞的激活)。是否存在抑制剂)。操纵靶向癌细胞表达 ova 也是可行的, 它可以诱导 ot-1 t 细胞杀死抗原特异性癌细胞。因此, 该检测还将有助于研究 mam/mdcs 介导的抗原特异性 t 细胞激活抑制。也可以应用该检测方法来研究人体细胞, 因为人类 cd3 和 cd28 的激活抗体在商业上是可以获得的, 并建立了从临床样本中分离人体 tam 的协议 19.
总的来说, 这种检测是相当多才多艺, 可用于检测其他免疫细胞类型的细胞毒性。目前, 在我们的实验室, 它正在扩大, 以检查抗原依赖的细胞毒性在各种条件下, 也正在开发的高通量筛选。
披露声明
作者没有什么可透露的。
致谢
这项工作得到了 wellcome 信托基金 (101067 z/13z (jwp) 的赠款. 109657/z z (tk), 615kit/j22738 (tk), uk, 和 mrc (mrm/n022556, uk)。noc 和 dds 感谢国家表型筛查中心基因组学发现计划和英国癌症研究组织 (noc) 的支持
材料
| Name | Company | Catalog Number | Comments |
| 0.05% Trypsin EDTA (1X) | Gibco | 25300-054 | |
| 12-Well Cell Culture Plate | Freiner Bio-One | 665-180 | |
| 1x PBS | Gibco | 14190-094 | |
| 2-mercaptethanol | Sigma | M6250-10ML | |
| 5mL Plystyrene Round-Bottom Tube | FALCON | 352054 | |
| 96-Well Cell Culture Plate (Round Bottom ) | Costar | 3799 | Co-culture of CD8+T cells with sorted myeloid cells |
| 96-well plate (Flat bottom) | Nunc | 165305 | Co-culture of CD8+T cells with target cells for cytotoxicity assay |
| AF647 anti-mouse F4/80 Antibody | BIO-RAD | MCA497A647 | Clone: CIA3-1, Lot#: 1707, 2 μL/1x10^6 cells |
| AlexaFluor700 anti-mouse CD8 Antibody | Biolegend | 100730 | Clone: 53-6.7, Lot#: B205738, 0.5 μL/1x10^6 cells |
| anti-mouse CD28 Antibody | Biolegend | 102111 | Activation of isolated CD8+ T cells, Clone: 37.51, Lot#: B256340 |
| anti-mouse CD3e Antibody | Biolegend | 100314 | Activation of isolated CD8+ T cells, Clone: 145-2C11, Lot#: B233720 |
| APC anti-mouse CD3 Antibody | Biolegend | 100236 | Clone: 17A2, Lot#: B198730, 0.5 μL/1x10^6 cells |
| APC/Cy7 anti-mouse Ly6C Antibody | Biolegend | 128026 | Clone: HK1.4, Lot#: B248351, 1 μL/1x10^6 cells |
| Bovine Serum Albmin | Sigma | A1470-100G | |
| Cell Strainer (100μm Nylon) | FALCON | 352360 | To smash the spleen |
| Cell Strainer (40μm Nylon) | FALCON | 352340 | To filter the lung digestion |
| DAPI | Biolegend | 422801 | |
| Dulbecco′s Modified Eagle′s Medium | Gibco | 41966-029 | |
| EasySep Mouse CD8+ T Cell Isolation Kit | StemCell Technologies | 19853 | |
| Fetal Bovine Serum | Gibco | 10270-106 | |
| FITC anti-mouse CD4 Antibody | Biolegend | 100406 | Clone: GK1.5, Lot#: B179194, 0.5 μL/1x10^6 cells |
| Geltrex Ready-to-Use | Gibco | A1596-01 | Coating the 96-well plates for cytotoxicity assay |
| IncuCyte NucLight Red Lentivirus Reagent | Essen BioScience | 4476 | Lenti viral particules encoding mKate2 |
| IncuCyte ZOOM | Essen BioScience | Detector (fluorescence microscope) | |
| IncuCyte ZOOM 2018A | Essen BioScience | Analysis software | |
| L-Glutamine (100X) | Gibco | A2916801 | |
| Lung Dissociation Kit | Miltenyi | 130-095-927 | Preparation of single cell suspension from the tumor-bearing lung |
| MycoAlert Mycoplasma Detection Kit | Lonza | LT07-318 | |
| Non-essential amino acid (100X) | Gibco | 11140-035 | |
| NucView488 | Biotium | 10403 | Fluoregenic caspase-3 substrate |
| PE anti-mouse Ly6G Antibody | Biolegend | 127607 | Clone: 1A8, Lot#: B258704, 0.5 μL/1x10^6 cells |
| PE/Cy7 anti-mouse CD11b Antibody | Biolegend | 101216 | Clone: M1/70, Lot#: B249268, 0.5 μL/1x10^6 cells |
| Pen Strep | Gibco | 15140-122 | Penicillin Streptomycin for primary culture of cells |
| PerCP/Cy5.5 anti-mouse CD45 Antibody | Biolegend | 103132 | Clone: 30-F11, Lot#: B249564, 0.5 μL/1x10^6 cells |
| Polybrene (Hexadimethrine bromide) | Sigma | H9268 | |
| Puromycin | Gibco | A11138-03 | |
| RBC Lysis Buffer (10X) | Biolegend | 420301 | |
| Recombinant murine IL-2 | Peprotech | 212-12 | Activation of isolated CD8+ T cells |
| Sodium pyruvate (100X) | Gibco | 11360-070 | |
| TruStain fcX (anti-mouse CD16/32) Antibody | Biolegend | 101320 | |
| : Nikon 10X objective (resolution 1.22 µm) : Green channel excitation: 440 - 480 nm : Green channel emission: 504 - 544 nm : Red channel excitation: 565-605 nm : Red channel emission: 625 - 705 nm |
参考文献
- Barry, M., Bleackley, R. C. Cytotoxic T lymphocytes: all roads lead to death. Nature Reviews in Immunology. 2, 401-409 (2002).
- Durgeau, A., Virk, Y., Corgnac, S., Mami-Chouaib, F. Recent advances in targeting CD8 T-cell immunity for more effective cancer immunotherapy. Frontiers in Immunology. 9. , 14 (2018).
- Tanaka, A., Sakaguchi, S. Regulatory T cells in cancer immunotherapy. Cell Research. 27, 109-118 (2017).
- Fleming, V., et al. Targeting myeloid-derived suppressor cells to bypass tumor-induced immunosuppression. Frontiers in Immunology. 9, 398 (2018).
- Cassetta, L., Kitamura, T. Macrophage targeting: opening new possibilities for cancer immunotherapy. Immunology. 155, 285-293 (2018).
- Chahroudi, A., Silvestri, G., Feinberg, M. B. Measuring T cell-mediated cytotoxicity using fluorogenic caspase substrates. Methods. 31, 120-126 (2003).
- Zaritskaya, L., Shurin, M. R., Sayers, T. J., Malyguine, A. M. New flow cytometric assays for monitoring cell-mediated cytotoxicity. Expert Review of Vaccines. 9, 601-616 (2010).
- Azimi, M., et al. Identification, isolation, and functional assay of regulatory T Cells. Immunological Investigation. 45, 584-602 (2016).
- Bruger, A. M., et al. How to measure the immunosuppressive activity of MDSC: assays, problems and potential solutions. Cancer Immunology Immunotherapy. , (2018).
- Qian, B., et al. A distinct macrophage population mediates metastatic breast cancer cell extravasation, establishment and growth. PLoS One. 4, e6562 (2009).
- Kitamura, T., et al. Monocytes differentiate to immune suppressive precursors of metastasis-associated macrophages in mouse models of metastatic breast cancer. Frontiers in Immunology. 8, 2004 (2018).
- Bronte, V., et al. Recommendations for myeloid-derived suppressor cell nomenclature and characterization standards. Nature Communications. 7, 12150 (2016).
- Kitamura, T., et al. CCL2-induced chemokine cascade promotes breast cancer metastasis by enhancing retention of metastasis-associated macrophages. Journal of Experimental Medicine. 212, 1043-1059 (2015).
- Ewens, A., Mihich, E., Ehrke, M. J. Distant metastasis from subcutaneously grown E0771 medullary breast adenocarcinoma. AnticancerResearch. 25, 3905-3915 (2005).
- Choppa, P. C., et al. Multiplex PCR for the detection of Mycoplasma fermentans, M. hominisandM. penetransin cell cultures and blood samples of patients with chronic fatigue syndrome. Molecular and Cellular Probes. 12, 301-308 (1998).
- Strober, W. Trypan blue exclusion test of cell viability. Current protocols in immunology. , (2001).
- Koyanagi, M., Kawakabe, S., Arimura, Y. A comparative study of colorimetric cell proliferation assays in immune cells. Cytotechnology. 68, 1489-1498 (2016).
- Gabrilovich, D. I., Nagaraj, S. Myeloid-derived-suppressor cells as regulators of the immune system. Nature Reviews in Immunology. 9, 162-174 (2009).
- Cassetta, L., et al. Isolation of mouse and human tumor-associated macrophages. Advances in Experimental Medicine and Biology. 899. 899, 211-229 (2016).
转载和许可
请求许可使用此 JoVE 文章的文本或图形
请求许可This article has been published
Video Coming Soon
版权所属 © 2025 MyJoVE 公司版权所有,本公司不涉及任何医疗业务和医疗服务。