Для просмотра этого контента требуется подписка на Jove Войдите в систему или начните бесплатную пробную версию.
Method Article
Респирометрия высокого разрешения для оценки биоэнергетики в клетках и тканях с использованием камерных и пластинчатых респирометров
В этой статье
Резюме
Оценка окислительного фосфорилирования с помощью респирометров высокого разрешения стала неотъемлемой частью функционального анализа митохондрий и клеточного энергетического обмена. Здесь мы представляем протоколы анализа клеточного энергетического метаболизма с использованием камерных и микропластинчатых респирометров высокого разрешения и обсуждаем ключевые преимущества каждого устройства.
Аннотация
Респирометрия высокого разрешения (HRR) позволяет контролировать окислительное фосфорилирование в режиме реального времени для анализа отдельных клеточных энергетических состояний и оценки дыхательных комплексов с использованием диверсифицированных протоколов титрования субстрат-разъединитель-ингибитор (SUIT). Здесь демонстрируется использование двух устройств респирометрии высокого разрешения, а также представлена базовая коллекция протоколов, применимых для анализа культивируемых клеток, волокон скелетной и сердечной мышц, а также мягких тканей, таких как мозг и печень. Протоколы для культивируемых клеток и тканей предусмотрены для камерного респирометра и культивируемых клеток для респирометра на основе микропластин, оба охватывают стандартные протоколы дыхания. Для сравнения, разработанные CRISPR клетки HEK293 с дефицитом митохондриальной трансляции, вызывающей множественный дефицит дыхательной системы, используются с обоими устройствами для демонстрации клеточных дефектов дыхания. Оба респирометра позволяют комплексно измерять клеточное дыхание с их соответствующими техническими достоинствами и пригодностью в зависимости от исследуемого вопроса исследования и модели.
Введение
Митохондрии выполняют ключевое обеспечение энергией и представляют собой компартментированную органеллу, способствующую существенным клеточным биоэнергетическим и метаболическим процессам, таким как анаболизм нуклеотидов, липидов и аминокислот, биогенез железо-серного кластера и участвуют в передаче сигналов, таких как контролируемая гибель клеток 1,2,3 . Митохондриальная биоэнергетика посредством окислительного фосфорилирования способствует практически всем клеточным процессам внутри клетки, и, следовательно, митохондриальные дисфункции первичного или вторичного происхождения связаны с широким спектром заболеваний 4,5. Митохондриальная дисфункция включает в себя не только изменения в структуре или плотности митохондрий, но и в качестве и регуляции дыхательной системы6. Этот качественный элемент включает в себя управление субстратом, характеристики связи, посттрансляционные модификации, динамику крист и дыхательные суперкомплексы 7,8. Поэтому точный анализ митохондриальной биоэнергетики для экспериментальных и диагностических подходов к оценке энергетического обмена клетки важен для здоровья и болезней.
Митохондриальное окислительное фосфорилирование (OXPHOS) представляет собой последовательность реакций в дыхательной системе или системе переноса электронов (ETS) для генерации клеточной энергии через аденозинтрифосфат (АТФ)9. Мультиферментативная стадия для использования энергии от потока электронов через комплексы I и II к комплексу IV генерирует электрохимический протонный градиент через внутреннюю митохондриальную мембрану, впоследствии используемый для фосфорилирования аденозиндифосфата (АДФ) до АТФ через комплекс V (F1FO АТФ-синтаза) (Рисунок 1А).
Во-первых, два электронных носителя образуются во время трикарбоксового цикла (TCA), гликолиза и пируватного окисления: никотинамидадениндинуклеотид (NADH) и дигидрофлавинадениндинуклеотид (FADH2). NADH окисляется в комплексе I (NADH дегидрогеназа), в ходе которого два электрона переносятся в коэнзим Q (хинон восстанавливается до хинола), в то время как протоны закачиваются в межмембранное пространство (IMS). Во-вторых, комплекс II (сукцинатдегидрогеназа) окисляет FADH2 и подает электроны в коэнзим Q без перекачки протонов. В-третьих, в комплексе III (цитохром c оксидоредуктаза) электроны из коэнзима Q переносятся в цитохром c, в то время как протоны закачиваются в IMS. В-четвертых, цитохром c переносит электроны в комплекс IV (цитохром c оксидаза), конечный комплекс для перекачки протонов, и где кислород функционирует как акцептор электронов для ассимиляции протонов, в конечном итоге образуя воду. Именно этот кислород потребляют митохондрии, который можно измерить с помощью оксиграфа. Наконец, протоны, полученные из комплекса I, комплекса III и комплекса IV, используются для вращения комплекса V, тем самым генерируя АТФ9.
Важно отметить, что перенос электронов происходит не только линейным образом, иначе обозначаемым как цепь переноса электронов. Вместо этого электроны могут быть перенесены в пул коэнзима Q через несколько дыхательных путей и облегчают конвергентный поток электронов. NADH-субстраты и сукцинат, например, могут входить через комплекс I и комплекс II соответственно. Электроны от окисления жирных кислот могут быть пожертвованы через электрон-переносящий флавопротеиновый комплекс. Действительно, всесторонний анализ OXPHOS требует целостного подхода с соответствующими топливными субстратами (рисунок 1A).

Рисунок 1: Митохондриальное окислительное фосфорилирование и специфические протоколы субстрата и ингибитора. (А) Митохондрии и схема системы переноса электронов (CI-CIV) и митохондриальная F1F0 АТФ-синтаза (CV). Все структуры из PDB. На рисунках изображены только субстраты и ингибиторы, описанные в этом исследовании). (B) Образец следа потока кислорода в неповрежденных ячейках HEK293 с использованием стандартного протокола в устройстве mHRR. (C) Образец следа потока кислорода в неповрежденных ячейках HEK293 с использованием стандартного протокола в устройстве cHRR. (D) Образец следа потока кислорода в пермеабилизированных фибробластах человека от здорового донора с соответствующим протоколом SUIT. Сокращения: 1 = Рутинное дыхание неповрежденных клеток; 2 = состояние 2; 3 = государство 3(I); 4 = состояние 3(I) с cytC; 5 = состояние 3 (I+II); 6 = Утечка(ОМ); 7 = емкость ETS; 8 = S(ROT); 9 = ROX; 10 = TMPD; 11 = Az. ROT = Ротенон, AM = Антимицин, АТФ = Аденозинтрифосфат, Az = Азид, OM = Олигомицин, FCCP = Карбонилцианид-трифтор-метоксифенилгидразон; Asc = Аскорбат, TMPD = N,N,N',N'-тетраметил-p-фенилендиамин, Succ = Сукцинат, M = Малат, P = Пируват, ADP = Аденозиндифосфат, NAD = Никотинамиддениндинуклеотид, IMS = Межмембранное пространство, FAD = Флавинадениндинуклеотид. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}
Анализ емкости митохондриального OXPHOS с использованием HRR стал инструментальным биохимическим методом диагностической ценности не только для первичных митохондриальных дефектов10,11, но и для всех других областей биологии, таких как рак и старение12 лет. HRR позволяет определить клеточное дыхание путем анализа митохондриальной емкости OXPHOS, которая непосредственно отражает индивидуальную или комбинированную недостаточность митохондриального дыхательного комплекса и косвенно связана с клеточной дисфункцией и измененным энергетическим метаболизмом9. Методологически измерения дыхания выполняются с использованием клеток, тканей или изолированных митохондрий 11,13,14, с замороженным материалом, только частично подходящим15,16. Показано, что замороженная ткань имеет неповрежденную ETS с сохраненной суперкомплексной стабильностью15. Таким образом, в отличие от традиционных промежуточных продуктов TCA, соответствующие субстраты непосредственно подаются в ETS. Однако связь между ETS и синтезом АТФ теряется, поскольку целостность мембраны нарушается из-за повреждения замораживанием (образование кристаллов льда).
Эксперименты по дыханию обычно проводятся при физиологической температуре 37 °C для эндотерм в непроницаемых или пермеабилизированных клетках или тканях. В то время как первый рассматривает цитозольный метаболический контекст, последний обеспечивает энергетический вклад отдельных комплексов OXPHOS и АТФазы путем добавления специфических субстратов (и ингибиторов). Последовательность и вариация субстратов и ингибиторов привели к разработке разнообразного массива протоколовSUIT 17 и анализов18 для решения различных научных вопросов функции OXPHOS (рассмотрено в разделе12). Основной протокол клеточного дыхания оценивает четыре различных состояния: i) рутинное дыхание - дыхание в соответствующей дыхательной среде без какого-либо добавления субстратов или ингибиторов, потребляющих только эндогенные субстраты. Это состояние может выявить общий OXPHOS или вторично-индуцированные дефекты дыхания, вызванные, например, измененными профилями метаболитов. Далее добавление ингибитора АТФазы олигомицина выявляет проницаемость внутренней митохондриальной мембраны к протонам, определяемую как ii) утечка дыхания. Последующее титрование протонофора, такого как разъединитель карбонилцианида-трифтор-метоксифенилгидразон (FCCP), позволяет определить состояние, при котором емкость ETS максимальна в режиме открыто-трансмембранного протонного контура, определяемом как iii) несвязанное дыхание. Важно отметить, что несвязанное состояние может также возникать при экспериментальных вмешательствах из-за чрезмерного механического повреждения митохондриальных мембран. И наоборот, несвязанное состояние относится к дыхательному разъединению внутренним механизмом, который физиологически контролируется. Наконец, полное ингибирование ИТС путем добавления комплекса III ингибитора антимицина и комплекса I ингибитора ротенона определяет остаточное потребление кислорода (ROX) от немитохондриальных процессов, потребляющих кислород (Рисунок 1A-C).
Митохондриальная биоэнергетика состоит из пяти различных состояний дыхания19,20. Дыхание в состоянии 1 не имеет каких-либо дополнительных субстратов или АДФ, за исключением того, что эндогенно доступно. После добавления АДФ, но все же, без субстратов, достигается дыхание состояния 2. Когда добавляются подложки, позволяющие переносить электроны и синтез АТФ, достигается дыхание состояния 3. В этом состоянии емкость OXPHOS может быть определена при насыщенных концентрациях АДФ, неорганических фосфатов, кислорода, NADH- и сукцинат-связанных субстратов. Состояние дыхания 4 или дыхание LEAK может быть определено как состояние без АДФ или химически ингибированных АТФ-синтаз при наличии достаточного количества субстратов. Наконец, когда весь кислород обеднен (бескислородный) в условиях закрытой камеры, наблюдается дыхание в состоянии 5.
Существует несколько методов оценки клеточных энергетических состояний14 с двумя устройствами, доминирующими в текущей оценке OXPHOS в реальном времени путем анализа потребления кислорода, измеренного как функция снижения кислорода с течением времени в системе с закрытой камерой с различной применимостью в зависимости от экспериментальной модели и исследовательского вопроса: респирометр высокого разрешения Oroboros 2k и внеклеточный анализатор потока Seahorse XF. Оба прибора регистрируют скорость потребления кислорода как снижение пикомолей (пмоль) кислорода (О2) в секунду как абсолютное значение в камере или микропластине скважины. Удельное потребление кислорода на массу получают путем нормализации соответствующего потребления кислорода в определенном буферном рецепте на количество клеток (миллионы), вес ткани (мг) или количество белка.
O2k (Oroboros Instruments) представляет собой закрытую двухкамерную систему, оснащенную полярографическим датчиком кислорода (сокращенно камерный респирометр высокого разрешения: cHRR). Каждая экспериментальная камера вмещает 2 мл жидкости, которая поддерживается однородной магнитными мешалками. Полярографический датчик кислорода использует амперометрический подход для измерения кислорода: он содержит катод золота, анод хлорида серебра / серебра, а между раствором KCI создается электрохимическая ячейка, на которую подается напряжение (0,8 В). Кислород из пробирной среды диффундирует через 25-мкм фторированную этиленпропиленовую мембрану (O 2-проницаемую) и подвергается восстановлению на катоде, производя перекись водорода. На аноде серебро окисляется перекисью водорода, генерируя электрический ток. Этот электрический ток (ампер) линейно связан с парциальным давлением кислорода. Парциальное давление кислорода и коэффициент растворимости кислорода пробирной среды используются для расчета концентрации кислорода. Поскольку парциальное давление кислорода зависит от экспериментальной температуры, а полярографические измерения чувствительны к температуре, колебания температуры нуждаются в точном (±0,002 °C) регулировании нагревательным блоком Пельтье. Температура может контролироваться в диапазоне 4 °C и 47 °C.
Внеклеточный анализатор потока Seahorse XF (Agilent) представляет собой пластинчатую систему с 24- или 96-луночным форматом микропластин, в которой три флуоресцентных электрода измеряют потребление кислорода с течением времени в каждой скважине (сокращенно респирометр высокого разрешения на основе микропластин: mHRR). Максимум четыре порта в картридже для анализа доступны для автоматического впрыска во время анализа. Анализ содержит несколько циклов, каждый из которых состоит из трех фаз: 1) смешивание, 2) ожидание и 3) измерение. Во время фазы измерения датчики опускаются в микропластинку, создавая временно закрытую камеру, содержащую объем 7-10 мкл для измерения излучаемого света. Этот свет излучается встроенными в полимер флуорофорами на кончике сенсорных зондов, которые ощущают O2 на основе закалки фосфоресценции. Интенсивность флуоресцентного сигнала пропорциональна O2 и зависит от температуры датчика и пробирной среды. Поэтому точная оценка кислорода требует относительного подхода с фоновой скважиной без какой-либо выборки. Восстановление концентрации кислорода происходит во время фазы перемешивания, когда датчик перемещается вверх и вниз, чтобы смешать объем над временной камерой. Каждый цикл вычисляет одну норму потребления кислорода. Температура может контролироваться в диапазоне 16 °C и 42 °C.
HRR является золотым стандартом для оценки клеточной биоэнергетики при первичных и митохондриальных заболеваниях и общем клеточном метаболизме. В этом исследовании представлены основные протоколы HRR для оценки функции OXPHOS в клетках и тканях.

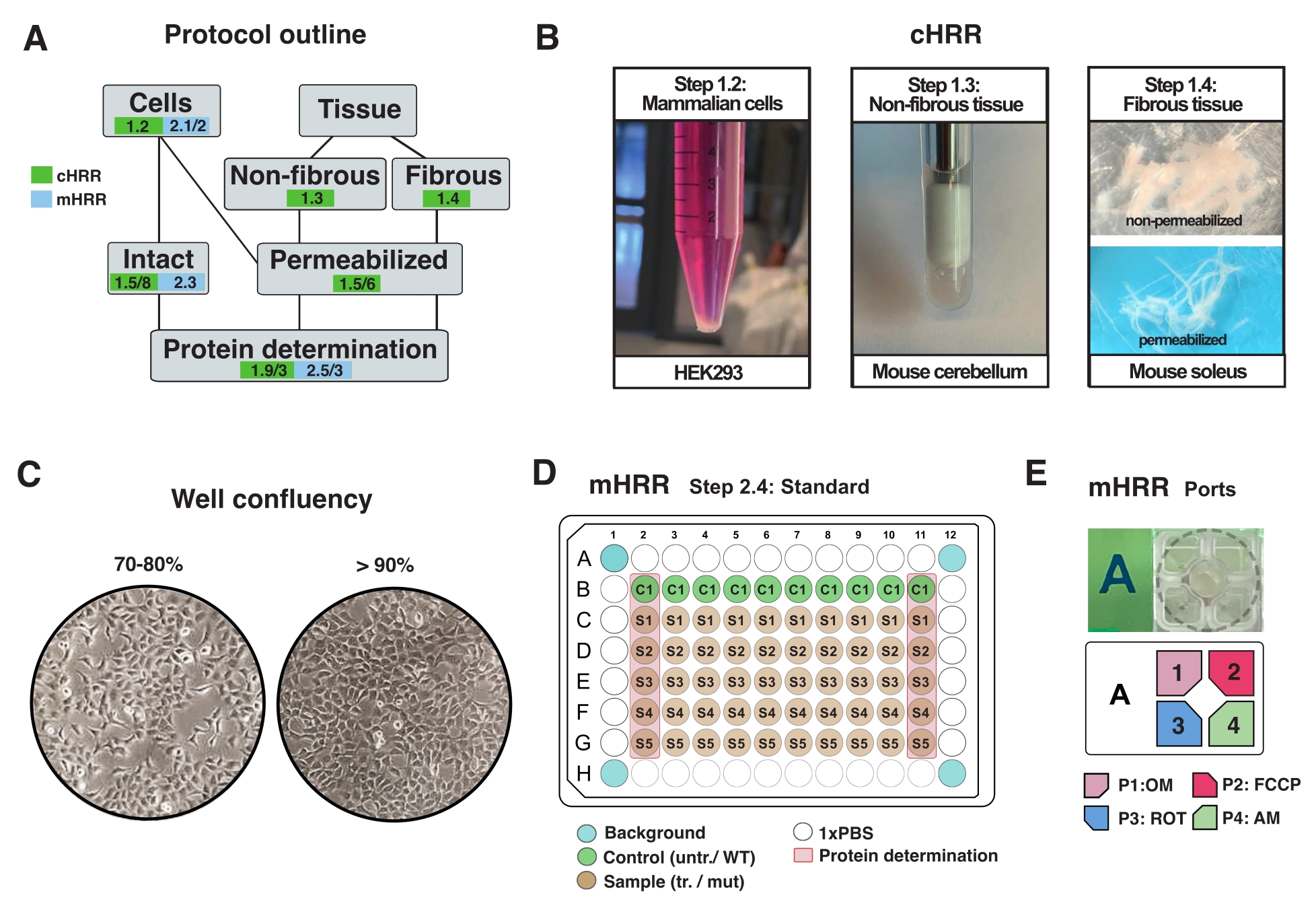
Рисунок 2: Рабочий процесс для клеточных и тканевых препаратов для cHRR и клеточной подготовки к mHRR респирометрии. (A) Краткое описание предоставленных протоколов. (B) Клетки млекопитающих (этап 1.2): гранулы HEK293, равные 3 x 106 клеткам (левая панель). Неволокнистая ткань (этап 1.3): Получение лизата мозжечка мышей в 2 мл тефлонового гончара (средняя панель). Сапонин-индуцированная пермеабилизация скелетных мышц (шаг 1.4) правая панель) для cHRR-респирометрии. (C) Стандартная схема посева микропластин (этап 2.4) и проверка слияния для анализа эукариотических клеток (HEK293) для респирометрии mHRR. (Д, Д) Схема загрузки инжекторного порта для респирометрии mHRR (шаг 2.4). Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}
протокол
Все эксперименты на животных проводятся в соответствии с Национальным советом по обзору экспериментов на животных и Региональным государственным административным агентством Южной Финляндии. В этом исследовании использовались самцы мышей C57BL/6JOlaHsd (4-6 месяцев). Согласие на использование клеточных линий человека было получено от институционального комитета по этике Хельсинкского университета.
1. Респирометрия высокого разрешения: камерный респирометр (cHRR)
ПРИМЕЧАНИЕ: Эксперименты в этом разделе протокола проводились с использованием Oroboros O2k-Core: Oxygraph-2k (Таблица материалов)
- Калибровка датчиков кислорода
- Предварительно запускают дыхательные аппараты при 37 °C в 2,1 мл митохондриальной дыхательной среды (MiR05, таблица 1, коэффициент растворимости: 0,92) в течение >45 мин и выполняют кислородную калибровку, как описано21. Продолжайте, если изменение базового уровня находится в пределах ± 4 пмоль/с.
ПРИМЕЧАНИЕ: Большие колебания фонового сигнала могут указывать на необходимость поддержания мембраны датчика или следов ингибиторов, оставшихся в камере от предыдущих экспериментов. Инструментальная коррекция фонового потока кислорода рекомендуется перед серией экспериментов25. - Записывайте значения калибровки кислорода для мониторинга производительности мембраны датчика с течением времени.
ПРИМЕЧАНИЕ: Это показывает функцию датчика, стабильность сигнал-шум и когда требуется обслуживание мембраны датчика. В зависимости от давления окружающей среды, в MiR05 солюбилизируется 180-200 мкмоль кислорода. - Удалите всю жидкость в камере перед добавлением любого образца в дыхательную среду.
ПРИМЕЧАНИЕ: Регулярно оценивайте объем дыхательных камер ровно в 2 мл.
- Предварительно запускают дыхательные аппараты при 37 °C в 2,1 мл митохондриальной дыхательной среды (MiR05, таблица 1, коэффициент растворимости: 0,92) в течение >45 мин и выполняют кислородную калибровку, как описано21. Продолжайте, если изменение базового уровня находится в пределах ± 4 пмоль/с.
- Подготовка клеток к респирометрии высокого разрешения
- Культивируйте клетки HEK293 в чашках диаметром 10см2 в среде Dulbecco's Modified Eagle's (DMEM) с высоким содержанием глюкозы, дополненной 10% термоинактивированной фетальной бычьей сывороткой (FBS), GlutaMax, заменимыми аминокислотами и Na-пируватом22 и уридином23 для поддержки OXPHOS-дефектного метаболизма в инкубаторе при 37 °C при 5% CO2.
ПРИМЕЧАНИЕ: Любой тип эукариотической клетки может быть культивирован. Для большинства типов клеток культивирование чашки размером 10см2 приводит к достаточному количеству клеток (обычно >3 х 106 клеток). Регулярно проверяйте микоплазменную инфекцию, чтобы избежать воздействия на клеточный метаболизм и дыхание. - Растите клетки без превышения 90% конфюсации (рисунок 2С).
ПРИМЕЧАНИЕ: Клетки со слиянием >90% могут проявлять зависящее от роста ингибирующее действие на дыхание (если оно не синхронизировано или постмитотическое). - Промыть клетки 1x PBS, отсоединить 1 мл теплого 0,25% трипсина, деактивировать трипсин, добавив теплый DMEM (5 мл / 10 см2 пластины) и подсчитать клетки с помощью гемоцитометра.
- Осторожно центрифугируют клеточный раствор, равный 2,5 х 106 ячеек при 300 х г в течение 5 мин, удаляют супернатант полностью и повторно суспендируют в 2,5 мл теплого MiR05 (1 х 106 клеток/мл) (рисунок 2А).
- Для суспензионных ячеек подсчитайте и удалите раствор, равный 2,5 х 106 ячеек, гранулируют и продолжают, как указано на этапе 1.2.4.
- Запустите протокол SUIT для оптимизации пермеабилизации (шаг 1.6), пермеабилизированной клетки или ткани (шаг 1.5) или интактных клеток (шаг 1.7)
ПРИМЕЧАНИЕ: Для получения стабильных результатов рекомендуется поддерживать постоянную концентрацию клеток (например, 1 x 106 клеток/мл). Хотя дыхание не зависит от плотности клеток в респирометре24, субстраты и ингибиторы находятся в сопоставимой концентрации на протяжении всех экспериментов, если количество клеток остается постоянным.
- Культивируйте клетки HEK293 в чашках диаметром 10см2 в среде Dulbecco's Modified Eagle's (DMEM) с высоким содержанием глюкозы, дополненной 10% термоинактивированной фетальной бычьей сывороткой (FBS), GlutaMax, заменимыми аминокислотами и Na-пируватом22 и уридином23 для поддержки OXPHOS-дефектного метаболизма в инкубаторе при 37 °C при 5% CO2.
- Подготовка нефиброзной ткани (например, мозга, печени) к респирометрии высокого разрешения
- Иссекните однородный кусочек ткани, весом 30-40 мг, или используйте весь орган (в данном случае мозжечок мыши).
ПРИМЕЧАНИЕ: Если ткань не используется сразу, держите в 2 мл ледяного MiR05, обеспечивая сохранение до 2 ч для большинства тканей. Индивидуальное время хранения тканей должно оцениваться во временных рядах. - Смочите ткань насухо фильтровальной бумагой Whatman (осторожно: мягкие ткани имеют тенденцию прилипать).
- Поместите кусок ткани 30-40 мг в охлажденный льдом гомогенизатор политетрафторэтиленового гончара Elvehjem.
- Добавьте соответствующее количество MiR05 для получения 20 мг/мл для поддержания соотношения ткани к буферу. Держите общее количество >1,5 мл и <2 мл, чтобы избежать недостаточного или избыточного количества жидкости для соответствующей механической пермеабилизации.
- Вставьте пестик, медленно лизируйте ткань, осторожно втягивая пестик, избегая при этом образования вакуума, вызывающего чрезмерное повреждение тканей.
- Выполните в общей сложности 7 ударов (1 удар, определяемый как один удар вверх и вниз) до лизирования (видимого в виде мутной жидкости без крупных обломков) (рисунок 2B).
ПРИМЕЧАНИЕ: Количество ударов для соответствующего лизиса должно быть проверено для каждой ткани путем оценки целостности внешней митохондриальной мембраны через цитохромный С-ответ (этап 1.5.11). Трудноизвлекаемая соединительная ткань или части сосудов могут остаться. - Декантируйте лизированную ткань в центрифужную трубку объемом 15 мл.
- Промыть внутреннюю часть гончара равным количеством MiR05, используемым на стадии лизирования (например, 1,5 мл), и добавить в 15 мл пробирку, содержащую теперь 3-4 мл MiR05 при 10 мг/мл тканевого лизата.
- Добавьте 2 мл обычного MiR05 на камеру до температуры до 37 °C.
- Медленно вращайте трубку для равномерного распределения перед пипеткой 500 мкл (что равно 5 мг) каждого лизата на камеру, чтобы свести к минимуму напряжение от холода до 37 °C.
- Подождите >3 мин, пока содержимое камеры нагреется до 37 °C, прежде чем закрывать камеру. Удалите лишнюю жидкость сверху пробки (количество на камеру после закрытия: 4 мг).
- Запустите протокол SUIT для стандартной пермеабилизации (шаг 1.5).
- Иссекните однородный кусочек ткани, весом 30-40 мг, или используйте весь орган (в данном случае мозжечок мыши).
- Подготовка фиброзной ткани (скелетной мышцы, сердечной мышцы) к респирометрии высокого разрешения
- Извлеките твердую ткань, удалите соединительную ткань и жир из мышц с помощью острых щипцов в 2 мл ледяного BIOPS (таблица 2) под рассеченным микроскопом.
- Отделите пучки волокон (~4 мг) вдоль продольной оси острыми щипцами. Выдавливайте волокна достаточно, чтобы получить сетчатую структуру (рисунок 2B).
ПРИМЕЧАНИЕ: О правильном механическом разделении и пермеабилизации волокон свидетельствует потеря красного пигмента миоглобина и повышенная полупрозрачность. - Промыть и пермеабилизировать пучок волокон в сапонине (50 мкг/мл в BIOPS, приготовленном в свежем виде) в течение 20 мин при 4 °C (волокна становятся полупрозрачными, что указывает на полную пермеабилизацию, рисунок 2B).
- Промывайте волокна дважды в MiR05 в течение 5 мин на одну стирку при 4 °C.
- Промокните фильтровальной бумагой и взвесьте перед добавлением в камеру, заполненную 2,1 мл MiR05.
- Введите пробки без полного закрытия, затем насытите камеры 2 мл чистого O2 с помощью шприца объемом 20 мл и закройте камеры, скручивая пробки во вращающемся движении. Поддерживайте концентрациюO2 в пределах 300-500 мкМ во время эксперимента, чтобы избежать ограничения диффузии кислорода.
- Протокол оценки рутинного дыхания в клетках или тканях
- Добавить образец в камеру, как указано на этапах 1.5.2-1.5.3.
- Добавьте 2,3 мл теплой клеточной суспензии MiR05 (стандартный вход: 1 x 106 клеток/мл, как на этапе 1,2 или 2 мг ткани/мл, как на этапе 1.3)
- Скелетная и сердечная мышцы (шаг 1.4): Добавьте ~ 4 мг сапонин-пермеабилизированных волокон к предварительно расплавленным 2,3 мл теплого MiR05 с учетом шагов 1.4.4-1.4.6
- Рабочие камеры при 37 °C и скорости перемешивания 700 об/мин. Подождите >3 мин, чтобы среда могла дегазировать и закрыть камеры, покрутив пробку во вращающемся движении. Стабилизация блока Пельтье указывает на достижение заданной температуры.
- (НЕОБЯЗАТЕЛЬНО) Измените скорость мешалки на 300 об/мин, чтобы оставшиеся пузырьки могли выйти через капилляр пробки.
- Аспирируйте любую лишнюю жидкость поверх пробки. Подождите 10 минут, пока не будет достигнут стабильный сигнал потока кислорода с любым типом образца, чтобы записать рутинное /состояние дыхания 1, рисунок 1B).
- Для измерения дыхания в пермеабилизированных клетках и тканях перейдите к шагу 1.6. Для интактных ячеек с шагом 1.8.
- Протокол анализа OXPHOS в пермеабилизированных клетках или тканях
- Используйте лизированный (пермеабилизированный) образец ткани или пермеабилизируйте клетки, добавляя 1 мкл дигитонина (запас дигитонина 8,1 мМ в диметилсульфоксиде (ДМСО)) для конечной концентрации 5 мкг/мл для пермеабилизации клеток. Поток будет падать и должен стабилизироваться через >5 мин.
ВНИМАНИЕ: Дигитонин остро токсичен для дыхательных путей, при контакте с кожей или при проглатывании.
ПРИМЕЧАНИЕ: Впрыскивание всех химических веществ осуществляется с помощью прецизионных стеклянных шприцев. Используйте шприцы только для указанных химических веществ, чтобы избежать перекрестного загрязнения и тщательно промыть в воде и EtOH после использования. Заблокированные шприцы могут потребовать ультразвука в теплом ddH2Oили чистящей проволоки для вытеснения любых химических засоров. Всегда втягивайте излишки соответствующего раствора в шприц, чтобы избежать попадания воздуха в камеры. Осмотрите внутреннюю часть камер на предмет введения воздуха после каждого впрыска. Записывайте каждый шаг до тех пор, пока поток не достигнет плато. - Добавить в быстрой последовательности: 5 мкл 0,4 М малата (М) для конечной концентрации 1 мМ, 5 мкл 2,0 М пирувата (Р; приготовленного в свежем виде), для конечной концентрации 5 мМ, 4 мкл 2,5 М глутамата (G) для конечной концентрации 5 мМ.
- После того, как предыдущий поток стабилизировался, добавляют 5 мкл (10 мкл для мышечной ткани) 0,5 М аденозиндифосфата (АДФ, аликвоты, хранящиеся при -80 °C) для конечной концентрации 1,25 мМ.
ПРИМЕЧАНИЕ: Ткани, такие как мышцы, могут нуждаться в другой концентрации для достижения насыщения. - Добавьте 5 мкл 4 мМ цитохрома C (cytC) для конечной концентрации 10 мкМ.
ПРИМЕЧАНИЕ: Необязательно для ячеек для оценки качества пермеабилизации. - Добавьте 16 мкл 1,25 М сукцината (S) для конечной концентрации 10 мМ. (НЕОБЯЗАТЕЛЬНО) Добавьте 3 мкл 0,5 М АДФ для конечной концентрации 2 мМ для контроля насыщенности концентрации АДФ.
- Для клеток и неволокнистой ткани добавляют 2 мкл 1 мг/мл олигомицина (ОМ) для конечной концентрации 1 мкг/мл.
ВНИМАНИЕ: Все используемые ингибиторы ETS являются высокотоксичными.
ПРИМЕЧАНИЕ: Олигомицин может потребовать титрования для оптимальной концентрации, поскольку он может подавлять способность ETS и опускается для мышечной ткани. Реоксигенат здесь, когда мышечная ткань анализируется и если O2 ниже 300 мкМ. - Титруйте FCCP из запаса 2 мМ, добавляйте 0,6 мкл с последующими шагами 0,2 мкл до тех пор, пока увеличение дыхания и дыхания не будет максимально разъединено (теоретически: несвязано).
- Добавьте 1 мкл 1 мМ ротенона (ROT) для конечной концентрации 0,5 мкМ. Добавьте 2 мкл запаса антимицина (AM) 1 мг/мл для конечной концентрации 1 мкг/мл.
- Повторное насыщение камер для достижения аналогичного уровня кислорода (~150 мкМ) во всех камерах путем медленного подъема плунжера в скручивающем движении.
- Добавьте 5 мкл аскорбата 0,8 М для конечной концентрации 2 мМ, за которым сразу же следует 5 мкл 0,2 М N,N,N',N'-тетраметил--фенилендиамин (TMPD) для конечной концентрации 0,5 мМ для оценки активности комплекса IV (необязательно).
- Добавьте 5 мкл 4 М азида для конечной концентрации 10 мМ сразу же после достижения пика потокаO2 с TMPD. Продолжайте работу в течение >5 мин для анализа автоокисления TMPD для комплексного расчета базового уровня IV.
- Пересчитайте ячейки, чтобы подтвердить количество ячеек перед запуском, и продолжите шаг 1.9.
ПРИМЕЧАНИЕ: Дигитонин-пермеабилизация (только для клеток) должна быть титрована в пробных экспериментах, чтобы достичь максимального потока и не влиять на целостность митохондриальной мембраны (см. шаг 1.7). Пермеабилизированные образцы (особенно мышечной ткани) с увеличением скорости дыхания >10% после добавления цитохрома с следует исключить из дальнейшего анализа из-за повреждения наружной митохондриальной мембраны. Ожидается кратковременное падение потока после добавления растворенных EtOH химических веществ.
- Используйте лизированный (пермеабилизированный) образец ткани или пермеабилизируйте клетки, добавляя 1 мкл дигитонина (запас дигитонина 8,1 мМ в диметилсульфоксиде (ДМСО)) для конечной концентрации 5 мкг/мл для пермеабилизации клеток. Поток будет падать и должен стабилизироваться через >5 мин.
- Протокол определения оптимальных условий пермеабилизации клеток
- Добавьте ячейки, как описано на шагах 1.2 и 1.5.2.
- Возьмите 10 мкл 10 мг/мл дигитонина и добавьте 10 мкл ДМСО для разбавления до 5 мг/мл.
- Добавить 1 мкл ротенона (запас 1 мМ). Добавьте 10 мкл сукцината (запас 2 мМ) и 5 мкл АДФ (запас 0,5 млн).
- Титруйте 1 мкл дигитонина (2,5 мг на шаг) многократно до тех пор, пока дыхание не увеличится дальше и не станет максимальным.
ПРИМЕЧАНИЕ: Снижение дыхания указывает на чрезмерную концентрацию дигитонина.
- Протокол анализа OXPHOS в интактных клетках
- После обычного дыхания (стадия 1.6.1-1.6.6) добавляют 2 мкл олигомицина 0,01 мМ для конечной концентрации 10 нМ.
- Титруйте FCCP из запаса 2 мМ, добавляйте 0,6 мкл с последующими шагами 0,2 мкл до тех пор, пока дальнейшее увеличение дыхания и дыхания не будет максимально разъединено (теоретически: несвязано)
- Добавьте 1 мкл 1 мМ ротенона для конечной концентрации 0,5 мкМ. Добавьте 2 мкл 1 мг/мл антимицинового сырья для конечной концентрации 1 мкг/мл.
- Повторное осаждение камеры до того же уровня кислорода (~150 мкМ), медленно поднимая плунжер в скручивающем движении.
- Добавьте 5 мкл 0,8 М аскорбата для конечной концентрации 2 мМ. Немедленно добавляют 5 мкл 0,2 М TMPD для конечной концентрации 0,5 мМ для оценки активности комплекса IV.
ПРИМЕЧАНИЕ: Подготовьте свежую партию перед любым большим набором экспериментов, так как TMPD склонен к автоокислению. Активность может снижаться с течением времени при хранении при температуре -20 °C. - Добавьте 5 мкл 4 М азида для конечной концентрации 10 мМ сразу же после достижения пика потокаO2 с TMPD. Продолжайте работу в течение >5 мин для анализа автоокисления TMPD для сложного расчета базового уровня IV.
- Пересчитайте ячейки, чтобы подтвердить количество ячеек перед запуском, и перейдите к шагу 1.9.
- Коллекция образцов после запуска
- Соберите ровно 1 мл MiR05-суспензии из каждой камеры (с включенными мешалками) в трубку 1,5 мл 1,5 мл.
- Центрифуга при 1000 х г для пермеабилизированных клеток или при 20 000 х г для тканевого лизата. Удалите супернатант и заморозьте гранулы при -80 °C для дальнейшей обработки (раздел 3).
- Анализ протоколов SUIT
- Анализ потока кислорода (пмоль/с, нормализованный для ввода) на каждом плато после добавления субстрата или ингибитора (Рисунок 1С и Рисунок 3А). Экспортируйте значения в электронную таблицу.
- Вычтите значение остаточного потребления кислорода (ROX, рисунок 1C и рисунок 3C) из всех значений каждого экспериментального запуска. Вычтите азидное остаточное дыхание из TMPD для получения сложного внутривенного дыхания.
- График абсолютных значений, нормализованных для клеточного (рисунок 3A, B) или тканевого ввода (рисунок 5A,B). Рассчитайте коэффициенты управления потоком (шаг 1.11) или нормализуйте их до поступления белка (рисунок 3C).
- Расчет коэффициента регулирования потока
- Получить индекс дыхательной функции и управления муфтой с помощью коэффициентов регулирования потока (FCR)9,26.
ПРИМЕЧАНИЕ: Это позволяет оценить внутреннее качество митохондрий, независимо от количества митохондрий. Кроме того, коэффициенты регулирования потока (FCR) сопоставимы в пределах одних и тех же клеточных линий, что позволяет контролировать качество реагентов (соответствующие FCR получаются с помощью указанных пронумерованных эталонных значений на рисунке 1B-D и рисунке 3C). - Рассчитайте коэффициент управления дыханием для связи OXPHOS с УТЕЧКОЙ с помощью уравнения 1.
Уравнение 1: FCRADP = 5/6 = Состояние 3 / Состояние 4 - Вычисление FCR для оценки NADH-зависимого дыхания с помощью уравнения 2
Уравнение 2: Состояние FCR3 (I) = 3/5 = Состояние 3 (I) / Состояние 3 (I+II) - Вычислите FCR для оценки сукцинат-зависимого дыхания с помощью уравнения 3.
Уравнение 3: состояние FCR3 (II) = 8/7 =S rot /емкость ETS - Вычислите FCR для оценки, связанного с несвязанным с помощью уравнения 4.
Уравнение 4: FCRсвязанный/несвязанный = 5/7 = Состояние 3 (I+II) /Емкость ETS - Чтобы проверить целостность внешней мембраны митохондрий, используйте уравнение 5.
Уравнение 5: % повреждения наружной мембраны митохондрий = 3/4 = Состояние 3 (I) / Состояние 3 (I) с цитой c
- Получить индекс дыхательной функции и управления муфтой с помощью коэффициентов регулирования потока (FCR)9,26.
2. Респирометрия высокого разрешения: микропластинчатый респирометр (mHRR)
ПРИМЕЧАНИЕ: Эксперименты в этом разделе протокола проводились с использованием анализатора внеклеточного потока Seahorse XFe96 (Таблица материалов)
- Клеточная культура
- Культивируйте клетки любого типа. Адгезивы (например, коллаген, ламинин) могут быть использованы для облегчения прикрепления клеток. Здесь клетки HEK293 культивируются, как и раньше (шаг 1.3).
- За день до эксперимента отделяют клетки и переносят их в назначенную mHRR 96-луночную микропластину для получения идеальной сливаемости в день эксперимента (80%-100%) (рисунок 2С).
ПРИМЕЧАНИЕ: Для mHRR плотность микропластинчатых ячеек имеет решающее значение. Индивидуальные свойства роста клеточных линий или методы лечения, влияющие на рост, должны быть учтены, чтобы обеспечить сопоставимое слияние в день эксперимента.
- Подготовка клеток к респирометрии высокого разрешения
- Заготавливайте и повторно суспендируйте клетки в достаточном объеме перед посевом
ПРИМЕЧАНИЕ: Рекомендуется сеять клетки из того же разведения для реплик. - Посев клеток в соответствии со скоростью роста отдельных клеточных линий или свойствами роста под обработкой.
ПРИМЕЧАНИЕ: Оптимизируйте на стандартной 96-луночной микропластине и экстраполируйте плотность ячейки на 96-луночную микропластинку, специфичную для анализа. В этой установке 7 x 104 клеток HEK293 WT были посеяны на скважину 96-лунки. Первая и последняя колонны 96-луночной пластины используются для определения белка (рисунок 2С). Четырехугольные колодцы не должны содержать никаких ячеек и используются для экспериментальной коррекции фона. В идеале, скважины, расположенные близко к краям, пусты, чтобы свести к минимуму эффект кромки (например, клетки показывают измененные темпы роста, вызванные температурными эффектами) (рисунок 2C, D).
- Заготавливайте и повторно суспендируйте клетки в достаточном объеме перед посевом
- Подготовка сенсорных пластин, загрузка ингибиторов
- В день анализа дополните 38,8 мл среды 0,4 мл 1 М глюкозы, 0,4 мл 200 мМ глутамина и 0,4 мл 100 мМ Na-пирувата.
ПРИМЕЧАНИЕ: дыхание mHRR требует специализированной небуферизованной среды DMEM при рН 7,4. В целом, 40 мл должно хватить на один эксперимент с одной 96-луночной микропластиной. - Нагрейте среду для анализа дыхания до 37 °C и замените среду клеточной культуры на среду для анализа дыхания, промыв дважды по 80 мкл на лунку.
- Установите пластину с ячейками в инкубатор при температуре 37 °C без CO2 за 60 мин до анализа.
ПРИМЕЧАНИЕ: Этот шаг необходим для дегазации пластины, так как CO2 может влиять на результаты дыхания, а сыворотка в среде может производить пузырьки во время анализа. - Предваряемые ингибиторы аликвот для OM, FCCP, ROT и AM до 37 °C и вынимают сенсорную пластину из инкубатора.
- Разбавляют OM, FCCP, ROT и AM в 3 мл пробирной среды до конечной концентрации в скважине 1,5 мкМ, 1,125 мкМ и 1 мкМ соответственно. Заполните отдельные порты, как показано на рисунке 2E.
ПРИМЕЧАНИЕ: Для заполнения картриджа датчика рекомендуется использовать многоканальную пипетку. Поскольку для нагнетания соединений используется воздух под давлением, все порты должны быть заполнены равным количеством объема жидкости всякий раз, когда порт заполняется соединением. ROT и AM могут быть объединены в одном порту. Ингибиторы могут быть растворены в EtOH или DMSO. - Проверьте порты впрыска и проверьте равномерность загрузки для каждого порта.
ПРИМЕЧАНИЕ: Все порты имеют отверстие в нижней части для инъекции. Следует соблюдать осторожность при перемещении сенсорной пластины. Пузырьки воздуха можно удалить с помощью иглы.
- В день анализа дополните 38,8 мл среды 0,4 мл 1 М глюкозы, 0,4 мл 200 мМ глутамина и 0,4 мл 100 мМ Na-пирувата.
- Протокол оценки кислорода в интактных клетках
- За день до анализа выполните шаги 2.4.2-2.4.7.
- Аликвот 20 мл калибрантного раствора в коническую трубку объемом 50 мл.
- Откройте набор для анализа внеклеточного потока и удалите содержимое.
- Поместите перевернутый картридж датчика рядом с полезной пластиной. Пипетка 200 мкл калибрантного раствора в каждую скважину полезной пластины.
- Прикрепите картридж датчика к вспомогательной пластине, обращая внимание, что все датчики погружены в воду.
- Установите пластину в инкубатор при температуре 37 °C без CO2 на ночь или минимум 12 ч. Убедитесь, что влажность внутри инкубатора достаточна для предотвращения испарения калибранта.
- Включите систему на основе микропластин и компьютер, чтобы быть готовыми к использованию на следующий день (машине требуется минимум 3 часа, чтобы уравновесить до 37 ° C перед проведением анализа).
ПРИМЕЧАНИЕ: Для обеспечения стабильности сигнала увеличьте точки измерения до 6 вместо 3 циклов измерения на состояние дыхания. Каждый цикл состоит из 3 минут смешивания и 3 минут измерения. - В день анализа XF выполните шаги 2.4.9-2.4.20.
- Проверьте слияние пластины клеточной культуры, морфологию клеток и то, что фоновые колодцы пусты.
- Промыть клетки подготовленной дыхательной средой, как указано в шагах 2.4.11-2.4.12.
- Удалите все, кроме 20 мкл питательной среды из каждой лунки. Удаляют 55 мкл, если культуральная среда была 80 мкл из-за испарения в течение ночи (приблизительно 5 мкл).
- Дважды промыть клетки 90 мкл пробирной среды. Наконец, добавьте 100 мкл пробирной среды. Конечный объем должен составлять 120 мкл.
ПРИМЕЧАНИЕ: На этом этапе рекомендуется использовать многоканальную пипетку, чтобы обеспечить применение одной и той же процедуры стирки к каждому экспериментальному состоянию (зависит от установки пластины). При аспирации наклоните пластину под углом 45° и поместите наконечники пипетки в угол колодцев для аспирации и закачки жидкостей. Крайне важно соблюдать осторожность во время промывки, так как некоторые клетки могут легко отсоединиться от нижней части пластины для культивирования клеток. - Установите пластину в инкубатор при температуре 37 °C без CO2 за 60 минут до анализа.
- Извлеките гидратированную пластину картриджа датчика из инкубатора безCO2.
- Откажитесь от старого раствора калибранта и замените его свежим раствором калибранта, предварительно расплавленным до 37 °C.
- Приготовьте ингибиторы и пробирную среду (3 мл на ингибитор в общей сложности 12 мл пробирной среды) и используйте резервуар для пипетки для загрузки ингибитора в порты.
- Откройте программное обеспечение и запустите предварительно разработанный или новый шаблон. Заполните карту пластин, отрегулируйте титрование и циклы измерения, а затем нажмите клавишу Пуск , чтобы начать калибровку оптических датчиков.
- Снимите крышку с заряженного картриджа и поместите ее в слот, который автоматически выскальзывает из устройства, убедившись, что маркировка в правом нижнем углу пластины совпадает с треугольником в правом нижнем углу слота.
- Нажмите кнопку Продолжить , чтобы выполнить автоматическую калибровку продолжительностью около 20 минут.
- После калибровки снимите опорную пластину, содержащую калибрант.
- Снимите крышку с микропластины, содержащей ячейки, и поместите пластину в прорезь по запросу устройства. Нажмите кнопку Продолжить , чтобы начать запуск.
- Коллекция образцов после запуска
- Выньте пластину из машины, осторожно удалите оставшиеся пробирные среды, не нарушая ячейки, и заморозьте всю пластину при -80 °C для дальнейшей обработки (раздел 3).
3. Определение белка с помощью анализа бицинхониновой кислоты (BCA assay)
- Приготовьте разбавленный бычий сывороточный альбумин (BSA) в буфере, используемом для экстракции белка и совместимом с BCA: 2 мг/мл, 1,5 мг/мл, 1 мг/мл, 0,5 мг/мл, 0,25 мг/мл и 0 мг/мл для стандартной кривой в дубликатах.
- Экстрагируйте белки путем повторного использования в соответствующем буфере лизиса (например, RIPA) с 20 мкл на лунку для mHRR или 100 мкл на гранулу, содержащуюся в пробирке 1,5 мл для cHRR.
- Инкубировать пластину mHRR или пробирку объемом 1,5 мл, содержащую белковые лизаты, в течение 30 мин на льду.
- Центрифугируют пробирку объемом 1,5 мл, содержащую лизат белка при 4 °C при 20 000 х г в течение 20 мин, и перенесут полученный супернатант в новую чистую трубку объемом 1,5 мл.
- Используйте 10 мкл на образец в дубликатах и эталонах в микротитерной пластине. Добавьте 200 мкл рабочего реагента BCA и инкубируйте в течение >15 мин.
- Считайте пластину в стандартном спектрофотометре на длине волны 562 нм и рассчитайте концентрации белка с помощью стандартной кривой BSA.
- Нормализуйте результаты дыхания до концентрации белка.
ПРИМЕЧАНИЕ: Нормализация количества белка позволяет подтвердить плотность посева клеток или ввод влажного веса. Экстрагированные белки пригодны для последующего иммуноблоттинга против субъединиц ETS, например, но не полностью представляют нативный образец (например, потеря участков фосфорилирования).
Результаты
Здесь мы предоставляем протоколы для определения митохондриальной биоэнергетики в эукариотических клетках, неволокнистой ткани (например, мозжечке) и фиброзной ткани (например, скелетных мышцах). Для эукариотических клеток HEK293 с CRISPR-инженерным нокаутом двух различных белков, связанн?...
Обсуждение
Традиционно митохондриальная биоэнергетика изучалась с помощью кислородных электродов типа Кларка. Однако отсутствие разрешения и пропускной способности оправдывало технический прогресс. На сегодняшний день O2k (называемый cHRR) и анализатор потока Seahorse XF96 (называемый mHRR) широко распро...
Раскрытие информации
Отсутствие конфликта интересов для раскрытия.
Благодарности
Эта работа была поддержана финансированием Академии Финляндии (C.B.J), Фонда Магнуса Эрнрута (C.B.J)) и докторской стипендии Интегрированной высшей школы наук о жизни (R.A.).
Материалы
| Name | Company | Catalog Number | Comments |
| 2 mL Potter-Elvehjem Glass/PTFE Tissue Grinder/Homogenizer | Omni International | 07-358029 | |
| 95% O2, 5% CO2 medical gas mixture | Potter for tissue grinding | ||
| ADP | Sigma | A 4386 | |
| Antimycin A | Sigma | A 8674 | Chemical |
| Ascorbate | Merck | PHR1279-1G | Chemical, dissolve in ethanol |
| BSA (fatty accid free) | Sigma | A 6003 | Chemical |
| CaCO3 | Sigma | C 4830 | Chemical |
| Cytochrome c | Sigma | C 7752 | Chemical |
| Digitonin | Sigma | D 5628 | Chemical |
| Dithiothreitol | Sigma | D 0632 | Chemical, dissolve in DMSO |
| D-Sucrose | Roth | 4621.1 | Chemical |
| Dulbecco’s modified Eagle’s medium (High glucose) | Fisher Scientific | 41965-039 | Chemical |
| Dulbecco’s modified Eagle’s medium (No Glucose) | Fisher Scientific | A14430-01 | |
| EGTA | Sigma | E 4378 | |
| Etomoxir | Sigma | E1905 | Chemical |
| Falcon 15 ml Conical Centrifuge Tubes | Fisher Scientific | AM12500 | Chemical |
| Falcon 50 ml Conical Centrifuge Tubes | Fisher Scientific | AM12501 | |
| FCCP | Sigma | C 2920 | |
| Glucose | Sigma | G7021 | Chemical, dissolve in ethanol |
| Glutamate | Sigma | G 1626 | Chemical |
| GlutaMax (100x) (200 nM L-alanyl-L-glutamine dipeptide) | Fisher Scientific | 35050061 | Chemical |
| HEK293 cells | ATTC | CRL-1573 | |
| Hemocytometer | Fisher Scientific | 0267151B | Instrument for cell counting |
| Hepes | Sigma | H 7523 | Chemical |
| Imidazole | Fluka | 56750 | Chemical |
| KCl | Merck | 1.04936 | Chemical |
| L-carnitine | Sigma | C0283 | Chemical |
| Malate | Sigma | M 1000 | Chemical |
| MES hydrate | Sigma | M8250 | Chemical |
| MgCl2 | Sigma | M 9272 | Chemical |
| Na2ATP | Sigma | A 2383 | Chemical |
| Na2Phosphocreatine | Sigma | P 7936 | Chemical |
| Na-pyruvate (100 mM) (100x) | Fisher Scientific | 11360070 | |
| NEAA (Non-essential amino acids) 100x | Fisher Scientific | 11140035 | |
| Normal FBS (10x) | Fisher Scientific | 10500064 | |
| O2k-Core: Oxygraph-2k | Oroboros Instruments | 10000-02 | High-resolution respirometry instrument |
| O2k-Titration Set | Oroboros Instruments | 20820-03 | Hamilton syringes for chemical injections |
| Oligomycin | Sigma | O 4876 | Chemical, dissolve in ethanol |
| Palmitoylcarnitine | Sigma | P 4509 | Chemical |
| Penicillin-Streptomycin | Fisher Scientific | 15140122 | |
| Pierce BCA Protein Assay Kit | Fisher Scientific | 23227 | |
| Pyruvate | Sigma | P 2256 | Chemical |
| RIPA-Buffer | Fisher Scientific | 89900 | Chemical |
| Rotenone | Sigma | R 8875 | Chemical, dissolve in ethanol |
| Saponin | Sigma | S7900 | Chemical |
Seahorse XF DMEM assay medium pack, pH 7.4 | Agilent, Santa Clara, CA | 103680-100 | |
| Seahorse XFe96 Extracellular Flux Analyzer | Agilent, Santa Clara, CA | High-throughput respirometry instrument | |
| Seahorse XFe96 FluxPak | Agilent, Santa Clara, CA | Includes assay plates, cartridges, loading guides for transferring compounds to the assay cartridge, and calibrant solution. | |
| Small scissors | Fisher Scientific | 08-951-20 | |
| Sodium azide | Sigma | S2002 | Chemical |
| Succinate | Sigma | S 2378 | Chemical |
| Taurine | Sigma | T 8691 | Chemical |
| TMPD | Sigma | T 3134 | Chemical |
| Trypan Blue solution | Merck | 72-57-1 | Chemical |
| Trypsin 0.25% EDTA | Fisher Scientific | 25200056 | |
| Two thin-edged forceps | Fisher Scientific | 12-000-122 | |
| Uridine stock (500x) | Sigma | U3750 | Chemical |
Ссылки
- McBride, H. M., Neuspiel, M., Wasiak, S. Mitochondria: More than just a powerhouse. Current Biology. 16 (14), 551-560 (2006).
- Mehta, M. M., Weinberg, S. E., Chandel, N. S. Mitochondrial control of immunity. Beyond ATP. Nature Reviews Immunology. 17 (10), 608-620 (2017).
- Spinelli, J. B., Haigis, M. C. The multifaceted contributions of mitochondria to cellular metabolism. Nature Cell Biology. 20 (7), 745-754 (2018).
- Gnaiger, E. Capacity of oxidative phosphorylation in human skeletal muscle. New perspectives of mitochondrial physiology. International Journal of Biochemistry and Cell Biology. 41 (10), 1837-1845 (2009).
- Gorman, G. S., et al. Mitochondrial diseases. Nature Reviews Disease Primers. 2, 1-23 (2016).
- Boushel, R., Gnaiger, E., Schjerling, P., Skovbro, M., Kraunsøe, R., Dela, F. Patients with type 2 diabetes have normal mitochondrial function in skeletal muscle. Diabetologia. 50 (4), 790-796 (2007).
- Cogliati, S., et al. Mitochondrial cristae shape determines respiratory chain supercomplexes assembly and respiratory efficiency. Cell. 155 (1), 160-171 (2013).
- Kühlbrandt, W. Structure and function of mitochondrial membrane protein complexes. BMC Biology. 13, 1-11 (2015).
- Gnaiger, E. Mitochondrial pathways and Respiratory control. An introduction to OXPHOS analysis. Bioenergetics communications. 5th ed. , (2020).
- Jackson, C. B., et al. Mutations in SDHD lead to autosomal recessive encephalomyopathy and isolated mitochondrial complex II deficiency. Journal of Medical Genetics. 51 (3), 170-175 (2014).
- Pesta, D., Gnaiger, E. High-resolution respirometry: OXPHOS protocols for human cells and permeabilized fibers from small biopsies of human muscle. Methods in Molecular Biology. 810, 25-58 (2012).
- Horan, M. P., Pichaud, N., Ballard, J. W. O. Review: Quantifying mitochondrial dysfunction in complex diseases of aging. Journals of Gerontology - Series A Biological Sciences and Medical Sciences. 67 (10), 1022-1035 (2012).
- Doerrier, C., Garcia-Souza, L. F., Krumschnabel, G., Wohlfarter, Y., Mészáros, A. T., Gnaiger, E. High-resolution fluorespirometry and oxphos protocols for human cells, permeabilized fibers from small biopsies of muscle, and isolated mitochondria. Methods in Molecular Biology. 1782, 31-70 (2018).
- Zhang, J., et al. Measuring energy metabolism in cultured cells, including human pluripotent stem cells and differentiated cells. Nature Protocols. 7 (6), 1068-1085 (2012).
- García-Roche, M., Casal, A., Carriquiry, M., Radi, R., Quijano, C., Cassina, A. Respiratory analysis of coupled mitochondria in cryopreserved liver biopsies. Redox Biology. 17, 207-212 (2018).
- Acin-Perez, R., et al. A novel approach to measure mitochondrial respiration in frozen biological samples. The EMBO Journal. 39 (13), 1-18 (2020).
- Cell metabolism assay kits. Seahorse assay kits and media Available from: https://www.agilent.com/en/product/cell-analysis/real-time-cell-metabolic-analysis/xf-assay-lits-reagents-cell-assay-media (2021)
- Chance, B., Williams, G. R. A method for the localization of sites for oxidative phosphorylation. Nature. 176 (4475), 250-254 (1955).
- Gnaiger, E., et al. Mitochondrial respiratory states and rates. MitoFit Preprint Arch. , (2019).
- Gnaiger, E. O2k-procedures: SOP O2k quality control 1: Polarographic oxygen sensors and accuracy of calibration Section Page. Oroboros. 03 (18), 1-21 (2020).
- Robinson, B. H., Petrova-Benedict, R., Buncic, J. R., Wallace, D. C. Nonviability of cells with oxidative defects in galactose medium: A screening test for affected patient fibroblasts. Biochemical Medicine and Metabolic Biology. 48 (2), 122-126 (1992).
- King, M. P., Attardi, G. Human cells lacking mtDNA: Repopulation with exogenous mitochondria by complementation. Science. 246 (4929), 500-503 (1989).
- Makrecka-Kuka, M., Krumschnabel, G., Gnaiger, E. High-resolution respirometry for simultaneous measurement of oxygen and hydrogen peroxide fluxes in permeabilized cells, tissue homogenate and isolated mitochondria. Biomolecules. 5 (3), 1319-1338 (2015).
- Fasching, M., Gnaiger, E. O2k quality control 2: Instrumental oxygen background correction and accuracy of oxygen flux. Mitochondrial Physiology Network. 14 (06), 1-14 (2016).
- Gnaiger, E., Lassnig, B., Kuznetsov, A., Rieger, G., Margreiter, R. Excess capacity of cytochrome c oxidase. Journal of Experimental Biology. 1139, 1129-1139 (1998).
- Gnaiger, E., et al. Mitochondria in the Cold. Life in the Cold. , 431-442 (2000).
- Fontana-Ayoub, M., Fasching, E., Gnaiger, Selected media and chemicals for respirometry with mitochondrial preparations. Mitochondrial Physiology Network. 02 (17), 1-9 (2014).
- Gerencser, A. A., et al. Quantitative microplate-based respirometry with correction for oxygen diffusion. Analytical Chemistry. 81 (16), 6868-6878 (2009).
- Krumschnabel, G., Eigentler, A., Fasching, M., Gnaiger, E. Use of safranin for the assessment of mitochondrial membrane potential by high-resolution respirometry and fluorometry. Methods in Enzymology. 542, 163-181 (2014).
- Nászai, A., Terhes, E., Kaszaki, J., Boros, M., Juhász, L. Ca(2+)N it be measured? Detection of extramitochondrial calcium movement with high-resolution fluorespirometry. Scientific Reports. 9 (1), 1-13 (2019).
- Pajak, B., et al. 2-Deoxy-d-Glucose and its analogs: From diagnostic to therapeutic agents. International Journal of Molecular Sciences. 21 (1), 234 (2019).
- Mercier-Letondal, P., Marton, C., Godet, Y., Galaine, J. Validation of a method evaluating T cell metabolic potential in compliance with ICH Q2 (R1). Journal of Translational Medicine. 19 (1), 1-15 (2021).
- Sauerbeck, A., et al. Analysis of regional brain mitochondrial bioenergetics and susceptibility to mitochondrial inhibition utilizing a microplate based system. Journal of Neuroscience Methods. 198 (1), 36-43 (2011).
- Jackman, M. R., Willis, W. T. Characteristics of mitochondria isolated from type I and type IIb skeletal muscle. American Journal of Physiology - Cell Physiology. 270 (2), 673-678 (1996).
- Ponsot, E., et al. Mitochondrial tissue specificity of substrates utilization in rat cardiac and skeletal muscles. Journal of Cellular Physiology. 203 (3), 479-486 (2005).
- Schönfeld, P., Reiser, G. Why does brain metabolism not favor burning of fatty acids to provide energy-Reflections on disadvantages of the use of free fatty acids as fuel for brain. Journal of Cerebral Blood Flow and Metabolism. 33 (10), 1493-1499 (2013).
- Calderon-Dominguez, M., Mir, J. F., Fucho, R., Weber, M., Serra, D., Herrero, L. Fatty acid metabolism and the basis of brown adipose tissue function. Adipocyte. 5 (2), 98-118 (2016).
- Divakaruni, A. S., Rogers, G. W., Murphy, A. N. Measuring mitochondrial function in permeabilized cells using the seahorse XF analyzer or a clark-type oxygen electrode. Current Protocols in Toxicology. 2014, 1-16 (2014).
- Iuso, A., Repp, B., Biagosch, C., Terrile, C., Prokisch, H. Assessing mitochondrial bioenergetics in isolated mitochondria from various mouse tissues using Seahorse XF96 analyzer. Methods in Molecular Biology. 1567, 217-230 (2017).
- Rogers, G. W., et al. High throughput microplate respiratory measurements using minimal quantities of isolated mitochondria. PLoS ONE. 6 (7), 21746 (2011).
- Jordá, A., Zaragozá, R., Portolés, M., Báguena-Cervellera, R., Renau-Piqueras, J. Long-term high-protein diet induces biochemical and ultrastructural changes in rat liver mitochondria. Archives of Biochemistry and Biophysics. 265 (2), 241-248 (1988).
- Jackson, C. B., Gallati, S., Schaller, A. QPCR-based mitochondrial DNA quantification: Influence of template DNA fragmentation on accuracy. Biochemical and Biophysical Research Communications. 423 (3), 441-447 (2012).
- Hirsch, H. M. Tissue autoxidation inhibitors: II. The presence of inhibitor in intact cells; Assay of liver and hepatoma effect on radio-oxidations. Cancer Research. 16 (11), 1076-1082 (1956).
- Picard, M., et al. Mitochondrial structure and function are disrupted by standard Isolation methods. PLoS ONE. 6 (3), 18317 (2011).
- Tanumihardja, E., Slaats, R. H., Van Der Meer, A. D., Passier, R., Olthuis, W., Van Den Berg, A. Measuring both pH and O2 with a single On-Chip sensor in cultures of human pluripotent stem cell-derived cardiomyocytes to track induced changes in cellular metabolism. ACS Sensors. 6 (1), 267-274 (2021).
- Harms, F., Stolker, R. J., Mik, E. Cutaneous respirometry as novel technique to monitor mitochondrial function: A feasibility study in healthy volunteers. PLoS ONE. 11 (7), 159544 (2016).
- Levitsky, Y., et al. Micro-respirometry of whole cells and isolated mitochondria. RSC Advances. 9 (57), 33257-33267 (2019).
Перепечатки и разрешения
Запросить разрешение на использование текста или рисунков этого JoVE статьи
Запросить разрешениеThis article has been published
Video Coming Soon
Авторские права © 2025 MyJoVE Corporation. Все права защищены