
Method Article
Zebra balığı epilepsi modelinde davranışsal ve fizyolojik analiz
Bu Makalede
Özet
Burada, DEPDC5 geninin geçici inhibisyonundan kaynaklanan bir zebra balığı epilepsi modelinin geliştirilmesi ve karakterizasyonu için bir protokol sunuyoruz.
Özet
Epilepsi, dünya çapında yaklaşık 50 milyon insanı etkileyen en yaygın nörolojik bozukluklardan birini temsil eder. Genetik araştırmalardaki son gelişmeler, bu bozukluğun heterojen doğasını vurgulayan çeşitli epilepsi formlarına karışan geniş bir gen spektrumu ortaya çıkarmıştır. Uygun hayvan modelleri, epilepsiye karışan genetik mutasyonların tetiklediği patolojik mekanizmaların araştırılması ve özel, hedefe yönelik tedavilerin geliştirilmesi için gereklidir. Son yıllarda zebra balığı, yeni anti-epileptik terapötikleri tanımlamak için hem genetik manipülasyon hem de pentylenetetrazol (PTZ) gibi bilinen epileptojenik ilaçlara maruz kalma ile epilepsilerin modellenimi için değerli bir omurgalı organizma olarak ortaya çıkmıştır. MTOR regülatörü DEPDC5'teki zararlı mutasyonlar, çeşitli odak epilepsileri formları ile ilişkilendirilmiştir ve zebra balığı ortologunun devrilmesi, spontan nöbet benzeri ataklarla ilişkili hiperaktiviteye, ayrıca gelişmiş elektrografik aktiviteye ve karakteristik dönüş tekerleği yüzmesine neden olur. Burada, DEPDC5 fonksiyon kaybı modelinin oluşturulmasında yer alan yöntemi tanımladık ve motor aktivitesini gübreleme sonrası (hpf) 28 ve 48 saat olarak değerlendirme protokolünü ve zebra balığı optik tekerleğindeki saha aktivitesini kaydetmek için bir yöntemi gösterdik. Epileptojenik ilaç PTZ'nin zaman içinde nöronal aktivite üzerindeki etkisinin bir örneği de sağlanmaktadır.
Giriş
Küçük boyutu, oviparöz gelişimi ve gelişimin erken aşamalarında şeffaflığı nedeniyle zebra balığı, kardiyovasküler, kanser veya nörolojik bozukluklar gibi çeşitli insan hastalıklarını modellemek için değerli bir omurgalı organizma olarak ortaya çıkmıştır1,2. Zebra balığı, organ mimarisinin ve genetik kodun yüksek korunması da dahil olmak üzere bir omurgalının avantajlarını, daha basit model organizmaların küçük boyutu ve genetik manipülasyon kolaylığı ile birleştirir, bu nedenle hem temel çalışmaları hem de çeviri uygulamalarını kolaylaştırır. Özellikle, hücresel süreçlerin davranış ve floresan belirteçlerinin yüksek verimli otomatik taranmasının alamet edilmesi, zebra balığını epilepsi araştırmaları için özellikle çekici bir model haline getirmiştir. Bu, epilepsi 3 ,4,5 ve daha yakın zamanda, bu modellerdekimyasal ekranlardan elde edilen umut verici terapötiklerin raporlarını içeren kimyasal kaynaklı ve / veya genetik modelleri içeren yayın sayısının son on yılında yüksek bir artışla gösterilmiştir6,7,8.
DEPDC5, mTOR signaling9'un negatif regülatörü olan GATOR1 kompleksinin bir üyesidir. DEPDC5 geninde mutasyonlar ilk olarak 2013 yılında otozomal dominant fokal epilepsi10,11'den muzdarip probandlarda keşfedilmiştir ve o zamandan beri fokal epileptik bulgular ve fokal kortikal displazi12ile ilişkili bir dizi klinik durumda bildirilmiştir. Bildirilen mutasyonların büyük çoğunluğunun gen12'ninfonksiyon kaybına neden olduğu tahmin edilmektedir ve bu, saçma aracılı mRNA çürümesi12,13tarafından hedeflenen bir dizi DEPDC5 mutasyona uğramış transkript için resmi olarak gösterilmiştir. Buna karşılık, antisense morfolüino oligonükleotidler (ABO' lar) kullanılarak zebra balıklarındaki gen ortoologunun devrilmesi, hiperaktivite, dönüş tekerleği benzeri yüzme, spontan nöbetler ve gelişmiş nöronal aktivite14 , 15 , 16,17,18dahil olmak üzere bu organizmadaki epileptik modellerde yaygın olan bir dizi özellik ile sonuçlanır. İlginçtir ki, mTOR sinyalinin bir inhibitörü olan rapamycin ile tedavi, bu modelin davranışsal özelliklerini tersine çevirdi18 DEPDC5 fonksiyon kaybının mTOR yolunun yanlış düzenlenmesi nedeniyle epilepsiyi tetikleyebileceği hipotezini destekleyen9,19.
Morfotin modifikasyon taşıyan antisense oligonükleotidler kullanılarak in vivo gen ekspresyonunun geçici olarak devrilmesi, si / shRNA tabanlı tekniklerle eşit olarak, belirli genlerin rolünü incelemek için paha biçilmez bir araç olmuştur. Son zamanlarda, AMO tabanlı stratejiler de klinik uygulamalar buldu, ilk AMO tedavisi 2016'da Duchenne kas atrofisi tedavisi için FDA onayını aldı20. Zebra balıklarında akut AMO tabanlı gen devirme fenotipinin her zaman constitutive knock-out modelleri21ile ilişkili olmadığı bildirilirken, bu en azından bazı durumlarda konsitülatif genetik değişikliklerle ortaya çıkan telafi edici mekanizmalara bağlı olabilir22. Bununla birlikte, AMO kaynaklı fenotipin özgüllüğü konusu, bu teknolojiyi kullanan çalışmalarda özenle ele alınması gereken tartışmasız birendişedir 23. AMO tabanlı devirme fenotipinin özgüllüğünü sağlamak için birkaç anahtar kontrol gereklidir. Bunlar arasında, gen devrilmesi için etkili en düşük dozda AMO'nun seçilmesine izin veren ve fazla miktarda genetik materyalin piyasaya sürülmesi nedeniyle genel toksisiteyi önleyen bir doz-yanıt eğrisi bulunur. Genomdaki belirli bir bölgeyi hedeflemeyen bir Uyumsuz AMO kullanımı da uygun bir doz oluşturmak ve belirli bir fenotip tanımlamak için gereklidir. Splice engelleyici AMO gibi aynı genin farklı bir bölgesini hedefleyen ikinci bir AMO, fenotipin hedef genin devrilmesinden kaynaklandığını doğrulamak için gereklidir. AMO tarafından hedef alınamayan zebra balığı geninin insan ortologu veya kodon modifiye edilmiş bir versiyonu olan genin cDNA'sıyla devirilen fenotipin kurtarılması, fenotip özgüllüğü lehine güçlü bir argüman sağlar. Fonksiyon kaybı mutasyonları içeren aynı cDNA ile kurtarma eksikliği (erken durdurma kodonlarının tanıtılması gibi) bu yönde başka bir kanıttır.
Burada, zebra balığı DEPDC5 fonksiyon kaybı modeli ve 28 ve 48 saat sonrası döllenme (hpf) davranışsal fenotipleme protokolü oluşturmak için bir yöntem sunuyoruz. 28 hpf'de DEPDC5 fonksiyon kaybı, koroyon içindeki embriyoların gelişmiş bobinleme ve seğirme hareketleriyle kanıtlandıkça genel hiperaktiviteye neden olur. Bu aşamada embriyo başına genel aktiviteyi ölçmek için otomatik hareket algılama sistemi kullanılabilir. 48 hpf'de zebra balığı dokunmaya yanıt olarak stereotipli kaçış yüzmesi sergiliyor. DEPDC5'inküçültülmüş ifadesine sahip zebra balıklarında, yüzme yörüngesi kontrollerden önemli ölçüde daha işkenceli, balıklar bu organizmadaki diğer bildirilen epilepsi modellerine benzer bir "mantar vidası" veya "dönüş tekerleği" gibi bir desen sergiliyor3,4. Zebra balığı larvalarındaki optik tekekumda döllenme sonrası 4-6 gün (dpf) arasında elektrofizyolojik kayıtlar elde edildi ve DEPDC5 devrilen hayvanlarda nöronal aktivitede temel bir artış gösterdi. Bu modelin avantajı, farklı zaman noktalarında çeşitli fenotipik özellikler sunmasıdır, bu da geliştirme sırasında ilaç tedavilerinin etkinliğini izlemede ve değerlendirmede yararlı olabilir.
Protokol
Deneysel prosedürler Ulusal ve Kurumsal Etik Kurullar tarafından onaylanmıştır.
1. Zebra Balığı Embriyosunda DEPDC5 Geninin Geçici Devrilmesi
- Araçların hazırlanması:
- Silikon elastomer kaplı enjeksiyon Petri kapları hazırlayın: Kitin tabanını ve kürleyici maddesini (bkz. Malzeme Tablosu)10:1 oranında karıştırın. 35 mm Petri kabını karışımla yarıya kadar doldurun. Kullanmadan önce silikonun sertleşmesi için bekleyin (bu birkaç gün sürebilir).
- Antisense morfinolino oligonükleotidlerin 1.2 mmol/L stok çözeltilerini hazırlayın (AMO; bkz. Malzeme Tablosu). 1,2 mmol/L stok çözeltisi elde etmek için 300 nmole lyophilized AMO'ya 250 μL steril su ekleyin. Tamamen çözünmesi için şişeleri 65 °C'de 5 dakika ısıtın. Kısaca Girdap. Tüp kapağını plastik bir filmle kapatın (bkz. Malzeme Tablosu).
- Kontrol kurtarma deneyleri için, pCS2 omurgasında klonlanmış DEPDC5'in insan cDNA'sını içeren bir ifade plazmidi hazırlayın (bkz. Malzeme Tablosu)veya benzer bir zebra balığı uyumlu ifade plazmidi. Negatif kontrol olarak, cDNA'da erken durma kodon (p.Arg487*) neden olan bir mutasyon ortaya konmuştır.
- Embriyo suyu hazırlayın: Ters ozmoz suyunda 0,06 g/L akvaryum tuzu (bkz. Malzeme Tablosu)+ 0,5 mg/L metilen mavisi.
- Enjeksiyon günü, bir çekme kullanarak mikroenjeksiyon borosilikat cam iğneler hazırlayın (bkz. Malzeme Tablosu). İğne çekmede uygun sıcaklık ayarlarını yapın. Yaklaşık 1 cm uzunluğunda ince uçlu iki ~5 cm kılcal damar oluşturmak için 10 cm uzunluğunda, 1/0,5 OD/ID mm borosilikat cam kılcal damar kullanın.
- İğnelerin ucu çok iyiyse, çözeltinin atılmasını önler, mikroskop altında forseps kullanarak konik ucun en ucunu kırın.
- Enjeksiyondan hemen önce, ABO'ların çalışma çözümlerini hazırlayın. Sonuçların tekrarlanabilirliğini sağlamak için her zaman yeni bir çözüm hazırlayın. AMO stok şişelerini 65 °C'de 5 dakika ısıtın. Hızlı Yeşil boya (%0,02 son konsantrasyon, bkz. Malzeme Tablosu)ve sudaki çalışma konsantrasyonunda seyreltilmiş AMO içeren 5 μL enjeksiyon numunesi hazırlayın.
- Bir doz yanıt eğrisi kullanarak her gen için AMO'nun çalışma konsantrasyonunu ampirik olarak belirleyin. Çalışma konsantrasyonu, AMO'nun brüt morfolojik kusurlar gibi genel toksisiteye neden olmadan geni devirmede etkili olduğu bir konsantrasyonu temsil eder. Tipik olarak, AMO çalışma konsantrasyonları 0,2 mmol / L ila 1 mmol / L aralığında olacaktır (bu çalışma için etkili konsantrasyon olarak 0,4 mmol / L belirlenmiştir18). Kontrol Uyumsuz morfotinini etkili AMO ile aynı konsantrasyonda enjekte edin.
- Damlacıkları tüplerin dibine getirmek için tüpleri ve santrifüjleri kısaca vorteks edin.
- Kurtarma deneyleri için, çalışma konsantrasyonunda seyreltilmiş AMO ve suda seyreltilmiş cDNA ekspresyon plazmidi ile ampirik olarak belirlenecek nihai konsantrasyona kadar 5 μL enjeksiyon örneği hazırlayın. DEPDC5 ve negatif kontrol plazmidi ekspresyasyonu için fenotipik kurtarma için 100 ng/μL etkili oldu.
- Embriyo hazırlığı:
- Mikroenjeksiyondan bir gün önce zebra balığı çiftleşme tanklarını kurun. Enjeksiyon sabahı, yumurtlamayı etkinleştirmek için bölücüleri çıkarın. Yumurtaları ince bir elek kullanarak embriyo suyu ile doldurulmuş 100 mm Petri kaplarında toplayın. Yumurtalar tek hücre aşamasındayken, toplamadan 20-30 dakika içinde enjekte edin.
- Plastik pastör pipet ile 60-80 yumurta seçin ve enjeksiyon için silikon kaplı Petri kabına yerleştirin. Silikon yüzey, enjeksiyonlar sırasında yumurtaların kaymasını önleyecektir. Embriyo suyunun çoğunu çıkarın, yumurtaları yarıya kadar kaplayacak kadar bırakın.
- Mikroinjections:
- Bir cam iğneyi enjeksiyon çözeltisi ile doldurun. İğneyi enjeksiyon çözeltisini içeren tüplerden birine dikey olarak yerleştirin ve iğnenin alt ucunun çözeltiye dokunmasını sağlayın. Renkli enjeksiyon çözeltisi kapillarite ile yükselene ve iğnenin ucunda görünene kadar birkaç dakika bekleyin.
- Doldurulmuş iğneyi mikroinjektörün enjekte sapına monte edin (bkz. Malzeme Tablosu).
- Hava kompresörini açın ve ~2 nL enjeksiyon hacmi oluşturmak için basınç ayarını ayarlayın.
- Enjekte edilen çözeltinin hacmini hesaplamak için, bir mikrotom kaydırağına bir damla mineral yağ yerleştirin. Ayarlanan basınç ve zaman parametrelerini kullanarak boya içeren çözeltiyi enjekte edin. Enjekte edilen akışkan kürenin çapını ölçün ve Hacim=4/3*π*(d/2)3formülünü kullanarak toplam hacmi hesaplayın, burada d=enjekte edilen bolus'un ölçülen çapı.
- 4X büyütmeli bir diseksiyon dürbün mikroskobu kullanarak, koroyon ve yumurta sarısı içinden geçerek ve çözeltiyi doğrudan hücre içine yansıtarak yumurtaları tek hücre aşamasında enjekte edin.
- Enjekte edilen embriyoları embriyo suyu ile 100 mm Petri kabında toplayın, kabı etiketleyin ve 28 °C'de kuluçkaya yatırın.
- Embriyoların gelişme hızı termosensitif olduğundan, inkübatör sıcaklığının zamanla stabil olduğundan emin olun. Örneğin, büyüme daha yüksek sıcaklıklarda hızlandırılacaktır ve gelişim aşaması fenotip24'üdüzgün bir şekilde değerlendirmek için kritik öneme sahiptir.
- Enjeksiyondan 6-8 saat sonra yumurtaların kalitesini kontrol edin ve plastik bir Pasteur pipet kullanarak ölü ve kısırlaştırılmamış embriyoları çıkarın.
- Ertesi sabah, her tabaktaki ölü embriyoları plastik bir Pasteur pipeti ile sayın ve çıkarın.
2. Davranış Analizi
- 28 hpf'de küresel faaliyet analizi:
- Testi mikroenjeksiyondan sonraki günün öğleden sonra yapın (28 hpf), embriyo gelişimi çok hızlı olduğu için geçerli istatistiksel analiz yapmak için testin yapıldığı günün saatinin deneyler üzerinde tutarlı olmasını sağlayın.
- 35 mm'lik bir kabı (test kabı) embriyo suyu ile doldurun ve teste başlamadan önce en az 15 dakika boyunca inkübatörde (28 °C) ısınmasını bırakın.
- Test kabının altına boyutuna göre kesilmiş plastik bir ağ ızgarası (1,2x1,2 mm) yerleştirin.
- Başka bir deneycinin embriyoların test sırasını rastgele hale vermesini ve test edilecek koşulların adlarını maskelemesi.
- Fenotipin özgüllüğünü sağlamak için mortalite oranının koşullar arasında ve enjekte edilmemiş embriyolarla karşılaştırıldığında değişmemesine dikkat edin. Ölü embriyoların her koşuldaki yüzdesi% 10-13'ü geçemez18.
- Plastik pasteur pipet kullanarak plastik ağ üzerinde hala korozyon içinde 10-12 embriyo yerleştirin. Test kabını embriyoları su altında tutacak kadar embriyo suyu ile doldurun, ancak yüzmeyen. Gerekirse, embriyoları ızgaraya yerleştirmek için plastik bir uç kullanarak dikkatli bir şekilde hareket ettinin.
- Diseksiyon mikroskobuna bağlı bir video kamera (bkz. Malzeme Tablosu)kullanarak, spontan bobinleme aktivitesini belirli bir süre boyunca kaydedin (10-20 dakikalık videolar genellikle nicelik için temsili aktivite patlamaları örnekleri elde etmek için yeterlidir)
- Embriyoları kendi çanaklarına geri koyun ve inkübatöre geri koyun. Deneyi her durum için gerektiği kadar embriyo ile tekrarlayın (%90 güç analizi ile belirlenir).
- Toplam spontan hareketi analiz etmek için zebralab sistemi kullanın (bkz. Malzeme Tablosu). Aktivite nicelleştirme modülini kullanarak, kaydedilen videoyu yükleyin ve her embriyonun etrafındaki izleme arenalarını uygun şekilde tasarlayın. Donma ve patlama eşiğini sırasıyla 10 ve 50 olarak ayarlayın.
- Tanımlanan arenaların her birinin içindeki toplam etkinliği ölçen otomatik video analizini çalıştırın, ardından veri kümesini elektronik tablo olarak kurtarın ve analizi bir veri çözümleme yazılımı kullanarak gerçekleştirin.
- Touch-Evoked-Escape-Response (TEER) 48 hpf'de:
- Testi enjeksiyondan iki gün sonra (döllenmeden 48 saat sonra) sabah gerçekleştirin.
- Testlerden en az 2 saat önce, ince kümesleri kullanarak embriyoları kaynatın. Azmonun ve davranış testinin günün saatinin deneyler üzerinde tutarlı olduğundan emin olun.
- 130 mm'lik bir kabı (test kabı) embriyo suyu ile doldurun ve teste başlamadan önce en az 15 dakika boyunca inkübatörde (28 °C) ısınmasını bırakın.
- Ölü ve morfolojik olarak deforme olmuş larvaları sayın ve çıkarın. Her koşul için sayıları kaydedin.
- Başka bir deneycinin sırayı rastgeleleştirmesini ve test edilecek koşulların adlarını kodlamasını sağlamak.
- Tüm test çanağının görüş alanında olduğundan emin olmak için kamerayı test kabının üzerine monte edin (bkz. Malzeme Tablosu). Görüş alanına bir cetvel yerleştirmek, mesafe için dahili bir kalibrasyon sağlar.
- Plastik bir Pasteur pipet ile, test kabının ortasına bir embriyo koyun ve 30 fps'lik bir alım oranı kullanarak kayda başlayın.
- İnce bir plastik uçla, titreşen bir hareketle embriyonun kuyruğuna hafifçe dokunun.
- Larva hareketini sonlandırdığında kaydı durdurun.
- Embriyoyu test kabından çıkarın ve embriyo suyuyla dolu yeni bir tabağa yerleştirin. Testi her durum için gerektiği kadar embriyo ile tekrarlayın (%90 güç analizi ile belirlenir).
- Embriyoları orijinal çanaklarına geri koyun ve inkübatöre geri koyun.
- Yüzme davranışının parametrelerini analiz etmek için, kaydedilen videoyu ImageJ analiz yazılımına yükleyin. ImageJ'in Manuel takip eklentisini indirip yükleyin (bkz. Malzeme Tablosu). Araçlar |'nı seçerek eklentiyi başlatın Eklenti | Menüde Manuel İzleme.
- İletişim penceresinde, görüntünün kalibre edilmiş ölçeğini tanıtın. Kamera alanına cetvel dahil etmek cm'nin piksellere dönüştürülmesini kolaylaştırır.
- Parça ekle'yi seçin ve ilk karedeki zebra balığı larvasının görüntüsüne tıklayarak yörünge izlemeyi başlatın. Çerçeveler, izleme eklenen her noktayla otomatik olarak ilerler.
- Yüzme bölümünün sonuna kadar hareketi izlemeye devam edin.
- İzleme penceresinde parçayı sonlandır'ı seçin, X-Y koordinatlarını alın ve toplam mesafeyi, hızı ve dönüş açısını hesaplayın.
3. Elektrofizyolojik Analiz
- Reaktif ve takım hazırlama:
- Embriyo suyunda %1 agarose hazırlayın (bkz. bölüm 1.1.4). Sıvı agaroseyu mikrosantrifüj tüplerinde aliquot ve agarose'un sertleşmesini önlemek için bunları 42 ° C'de bir ısıtma bloğunda tutun.
- Kayıt çözümünü hazırlayın (mmol/L):NaCl 134, KCl 2.9, CaCl2 2.1, MgCl2 1.2, glikoz 10, HEPES 10, pH 7.8.
- 1,5-2 μm uç açıklığı olan borosilikat cam mikropipetleri çekin (5-6 Mm2·kg·s−3· Bir−2 direnci) parlatılmamış.
- Zebra balığı larvalarının elektrofizyoloji için hazırlanması:
- Balığı cam tabanlı bir Petri kabına yerleştirin (bkz. Malzeme Masası)ve balığın kapak kaymasına mümkün olduğunca yakın getirilmesini sağlamak için fazla hücre dışı ortamı çıkarın.
- Plastik bir Pasteur pipet kullanarak, larvaların üzerine ve etrafına ılık sıvı agarose ekleyin. Balığı örtmek için yeterli agarose kullanın. Agarose sertleşirken, balığı yemeğin ortasında düz bir pozisyonda, ventral tarafta aşağı yönlendirmek için ince tokmaklar kullanın.
- Nöromüsküler iletimi engellemek için 10 μM Pancuronium bromür içeren kayıt çözeltisinin 2 mL'lik kısmınıekleyin (bkz. Malzeme Tablosu). Felçleyicinin eklenmesi, kayıtlar sırasında küçük hareketler nedeniyle eserleri ortadan kaldırmak için gereklidir.
- Elektrofizyolojik kayıt
- Mikropipette kayıt çözeltisi ile doldurun.
- Gerilim kelepçesi konfigürasyonunda yama kelepçesi amplifikatörü (bkz. Malzeme Tablası)ile, doğru değerini doğrulamak için banyoda elektrot direncini ölçün.
- 20x'lik bir hedef kullanarak, larva başını merkezi görüş alanına yerleştirin ve optik tectum içinde beyindeki kayıt konumuna ulaşmak için mikropipleti küçümser.
- Yama kelepçesi amplifikatörini akım kelepçesine geçirin ve tutma akımını 0 mA'ya sabitleyin.
- 1 kHz'lik düşük geçiş filtresi, 1 kHz'lik bir alım hızı ve 10 dijital kazanç kullanarak, temel etkinlik seviyelerini belirlemek için 60 dakika boyunca spontan aktivite kaydedin.
- 1 saat temel kayıttan sonra, 20 mmol / L PTZ son konsantrasyon için banyoya 200 μL pentylenetetrazol (PTZ, bkz. Malzeme Tablosu)çözeltisi 300 mmol / L ekleyin.
- PTZ'deki nöronal aktiviteyi 120 dakika daha kaydedin.
- Depolarizasyon olayı belirleme
- Alan kayıt olayları çok yavaş dinamiklere sahiptir (ilgi alanları 0.005-0.2 s-1aralığındadır). Bu nedenle, takma ad vermekten kaçınmak için sinyali düşük bir geçişle filtreleyin (Butterworth 5. sıra LPF 100 s-1'de). Kaydedilen voltaj verilerini alım kare hızından (bu durumda 1 ks-1)250 s-1'e (RAW SIGNAL) kadar alt örnekle.
- Her depolarizasyon olayının zaman damgalarını tanımlamak için, kaydedilen sinyalin yüksek geçişli filtrelenmiş bir sürümü olan algılama sinyali kullanın (Butterworth 1st order HPF at 0.01 s-1).
- Düşük frekanslı bileşenleri ortadan kaldırarak, depolarizasyon olaylarının tespiti basit bir eşikleme yöntemi kullanılarak gerçekleştirilebilir. Gürültü giderme ve olay algılama için sabit bir eşik kullanın (bu çalışma için 0,3 mV kullanılmıştır).
- Depolarizasyon olayını, 4 s'den küçük zaman aralıklarında meydana gelen bir dizi eşik geçişi ile karakterize edin. Ardışık eşik geçiş dizilerinden belirlendiği gibi depolarizasyon olayının başlangıcını ve sonunu hesaplayın. 40 ms'den kısa olaylar gürültü olarak atılabilir.
- Düşük geçiş filtrelemesinin olayın zirvesindeki etkisi nedeniyle hataları ortadan kaldırmak için filtrelenmemiş (RAW SIGNAL) olayların genliğini hesapleyin. Filtrelenmiş sinyalde belirlenen zaman damgalarını kullanarak ham sinyalden depolarizasyon dalgasını seçin. Genliği, ham sinyalden seçilen dalga adayının maksimum ve minimum değerleri arasındaki fark olarak ölçün.
NOT: Adım 3.4 — Depolarizasyon olay belirleme — gerçekleştirmek ve Şekil 1'i elde etmek için komut dosyası dosyaları bu makaleye ekli Ek Dosya olarak sağlanmıştır.
Sonuçlar
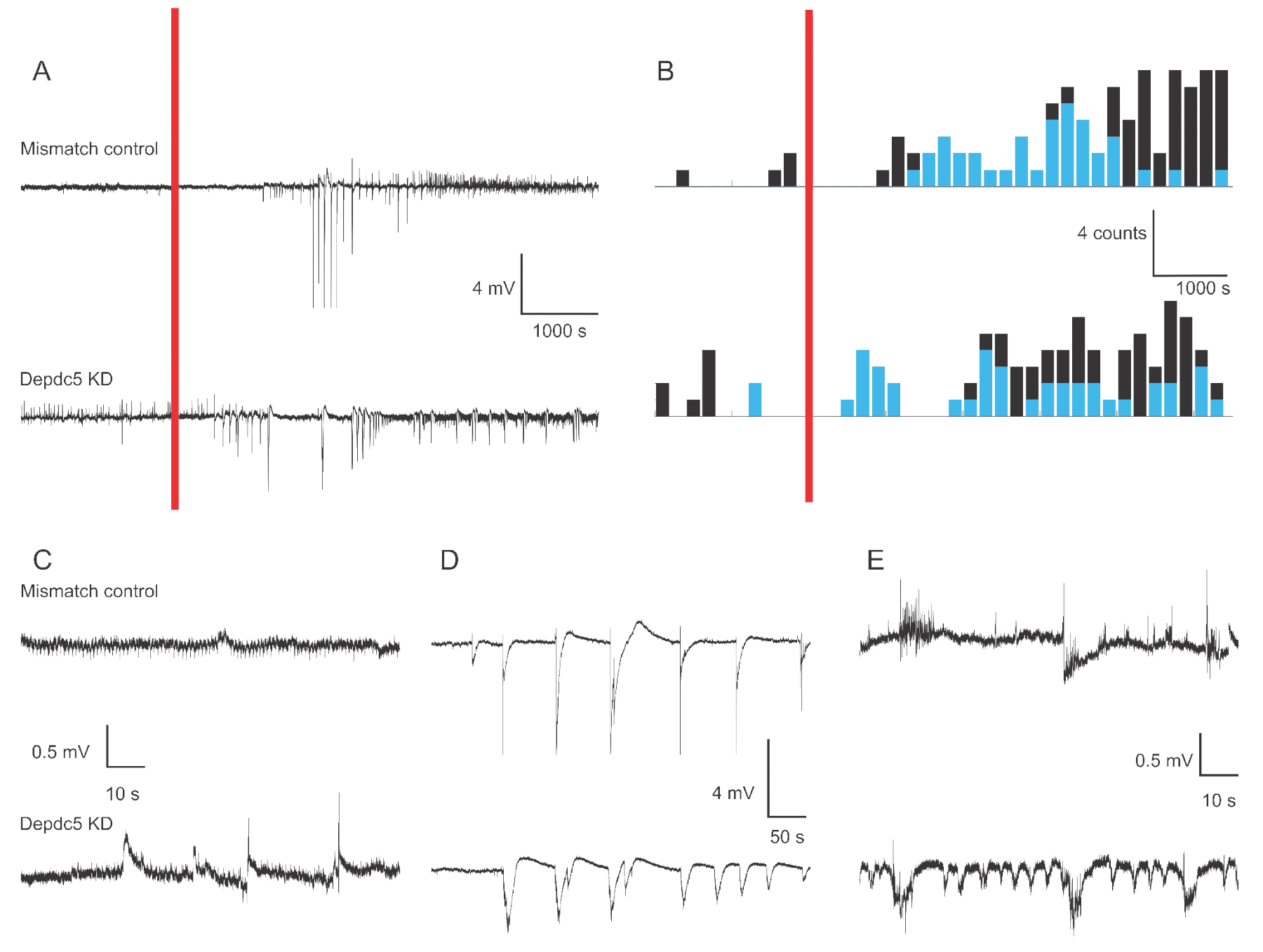
Şekil 1, iki genetik durumda 4-6 dpf zebra balığı larva hücre dışı alan kayıtlarının temsili voltaj izlerini göstermektedir: Uyumsuzluk kontrolü ve DEPDC5 devirme. Kaydın temel döneminde, DEPDC5 devirme spontan olayların daha yüksek bir oluşumunu gösterirken, Uyuşmazlık denetimi çok az dalgalanma görüntüler. Bu aktivite kalıpları, depdc5fonksiyon kaybı nedeniyle nöronal aktivitedeki önemli artışın temsilcisidir Daha önce bildirdiğimiz gibi18. PTZ uygulamasından sonra, hem Uyuşmazlık denetimi hem de DEPDC5 devirme, depolarizasyon olaylarının sayısının arttığını gösterir. PTZ uygulamasından sonraki ilk dönemde (10 – 60 dk), hem Uyuşmazlık kontrolünde hem de olayların çoğunun yüksek genlikte (>1 mV) olduğu DEPDC5 devirmede dakikada 0,8 olay oranı gözlenir. İkinci yanıt döneminde (PTZ uygulamasından sonra 60 – 120 dk), depolarizasyon olaylarının oranı dakikada yaklaşık 1 olaya çıkar ve olayların çoğu düşük genliktedir (≤1 mV).

Şekil 1: Zebra balığı larva beynindeki alan kayıtlarının örnek izleri. (A) Uyumsuz kontrol larvası ve DEPDC5 Knock-down için 180 dakikalık kayda genel bakış. önce spontan taban çizgisi aktivitesi kaydedildi, daha sonra banyoda PTZ uygulandı (kırmızı çubuk). (B) Uyumsuzluk kontrolü ve DEPDC5 devirme için depolarizasyon olaylarının peri-uyaran zaman histogramları. Olaylar yüksek genlik (>1 mV - mavi) ve düşük genlik (≤1 mV - siyah) olarak sınıflandırıldı. (C-E) Kaydın farklı dönemlerine ait örnek izler: (C) spontan aktivite, (D) PTZ uygulamasından sonraki ilk dönemde yüksek genlik olayları, (E) PTZ uygulamasından sonraki ikinci dönemde düşük genlik olayları. Bu şekilleri elde etmek için komut dosyası dosyalarının Tamamlayıcı Dosyaolarak sağlandığını unutmayın. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
{kind=link}
Tamamlayıcı Dosya: Adım 3.4 için komut dosyası dosyaları. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
Tartışmalar
Epilepsi, genetik dizileme teknolojilerinin ortaya çıkmasıyla aydınlatılmaya başlayan çok çeşitli etiyolojilere sahip karmaşık bir nörolojik hastalıktır25,26,27. Çok yönlü hayvan modelleri, genetik olarak bağlantılı epilepsilerin patolojik mekanizmaları hakkında hem içgörüler hem de bu durumun farklı formları için hedefli tedaviler sağlayacak verimli bir çeviri stratejisi için gereklidir. Zebra balığı modelleri epilepsinin başlıca özelliklerinin yeniden üretilmesinde ve anti-epileptik ilaç taraması için güvenilir okumalar sağlamada çok etkili olmuştur5,28. Spontan nöbetler genetiği değiştirilmiş zebra balıklarında tespit edilebilir15,29,30,31 ve bu modellerde nörofizyolojik analiz28 epileptik benzeri davranışın nöronal temelini doğrulamıştır32,33. Küçük boyutlu zebra balığı larvaları, potansiyel terapötiklerin hızlı bir şekilde algılanmasını sağlayan spontan yüzme gibi basit davranışların otomatik olarak algılanmasını kullanarak 96 kuyu formatında kimyasal ekranlara elverişlidir.
Burada sunulan DEPDC5 devirme modeli, geliştirme sırasında gen ekspresyonunun engellenmesi için zebra balığı embriyosuna AMO enjekte edilmesiyle elde edilir. Bu model, larva gelişiminin farklı zaman noktalarında, kimyasal veya genetik tarama protokolü sırasında terapi verimliliğinin göstergeleri olarak kullanılabilecek birkaç kilit taşı fenotipik özellik sunar. AMO aracılı gen devirme, özellikle ilgi çekici bir genin ekspresyonını hedeflediği için kimyasal olarak indüklenen nöbet modellerine göre avantajlar gösteren güçlü bir tekniktir, böylece genetik bir mutasyonun tetiklediği alttaki patojenik mekanizmaların tanımlanmasına izin verir. Yine de ilaç taramaları için güçlü araçlar olan kimyasal indükleyiciler, her zaman incelenen genetik mutasyonla ilgili olmayabilecek birden fazla hücresel yoldan hareket edebilir. AMO enjeksiyonu, deneyci tarafından ustalaşınca kendi başına basit bir teknik olsa da, bir dizi sınırlama da sunar. Enjeksiyonlar tek hücreli embriyoda yapılmalıdır; elimizde, daha sonraki aşamalarda enjeksiyonlar fenotipin değişkenliğini büyük ölçüde artırdı. Bu, enjeksiyon için mevcut süreyi sınırlar; bu nedenle, bir zaman dizisinde enjeksiyon için yumurta üretme stratejisi yararlıdır. Rutin olarak 15-20 dakika aralıklarla açtığımız 4-5 haç kullanıyoruz, bir sonrakini elde etmeden önce bir debriyajın enjeksiyonuna izin veriyoruz. Ayrıca, kalıplaşmış davranışlar gelişimin ilk günlerinde hızla geliştiği için, fenotipin farklı deneyler arasındaki aynı zaman noktalarında değerlendirilmesine özen gösterilmelidir. Aşırı miktarda enjekte edilmesine bağlı genel toksisite spesifik fenotipi maskeleyeceğinden, APO'ların hacmi ve konsantrasyonu da dikkatlice kontrol edilmelidir. Girişte sunulan farklı kontroller, doğru enjeksiyon dozunu ve ilgili fenotipi belirlemek için gereklidir.
Larva zebra balığı beyninin alan kayıtları, farklı beyin bozukluklarında rol oynayan genetik mutasyonların küresel nöronal aktivite üzerindeki zararlı etkilerini araştırmak için yararlı bir araçtır34. Bu deneysel koşullar altında görülen depolarizasyon olayları, ilaçların farklı epileptik koşullardaki elektrofizyolojik etkilerini değerlendirmek için belirlenmiş bir yöntemdir15,35. Bununla birlikte, bu etkilerin değerlendirilmesi çoğunlukla nicel olarak değil niteliksel olarak yapılmıştır ve analizde bir aktör olarak öznel bir gözlemciye sahip olmak. Burada, depolarizasyonların oranını, genliklerini ve sürelerini objektif olarak ölçebilen ve bu parametrelerin zaman boyunca veya farklı genetik veya farmakolojik müdahalelerle ilerlemesini değerlendirebilen otomatik bir algılama stratejisi geliştiriyoruz.
Burada sunulan temsili sonuçlar, EPILEPTIFORM benzeri elektrografik aktiviteyi tanıtmak için PTZ uygulamasından önce ve sonra, 4-6 dpf zebra balığında uyumsuzluk kontrolüne kıyasla DEPDC5 devirme genetik modelinin beklenen saha aktivitesini göstermektedir. Daha önce, DEPDC5 devirme durumunun bazal aktivitesinde önemli bir artış gösterdik18. Burada, bu iki koşulun kimyasal bir epileptikform aktivite indükleyicisi olan PTZ'ye verdiği yanıtın, nispeten düşük frekanslı, yüksek genlikli depolarizasyon olaylarından başlayarak ve daha yüksek frekanslı, daha düşük genlik depolarizasyon olayları ile devam eden zaman içinde benzer bir yörüngeye sahip olduğunu gösteriyoruz. Alan kayıt olayları yavaş dinamiklere sahiptir (ilgi alanları 0.005-0.2 s-1aralığındadır), bu nedenle bu protokolde ilgi çekici olayları yalıtmak için hem düşük geçişli hem de yüksek geçişli filtreler kullanılır. Düşük frekans gürültüsü ortadan kaldırıldıktan sonra, depolarizasyon olaylarının algılanması basit bir eşik kullanılarak gerçekleştirilir. Sinyalin istatistikleri depolarizasyon olaylarının varlığından büyük ölçüde etkilendiğinden, bu eşiği belirlemek için toplam sinyalin standart sapmasını kullanamadık. Veri kümeleri arasında standart sapmanın değerinin değişkenliği, gözlemlenen kayıt gürültüsü seviyelerinden daha büyüktü. Bu nedenle, izlerin görsel incelemesinden sonra, farklı depolarizasyon aktivitesi seviyelerinin neden olduğu önyargıyı önlemek için 0,3 mV eşik sabit bir değer kullandık.
Açıklanan protokol, zebra balığı modellerinde epileptiform benzeri fenotipleri karakterize etmek için optik tectumdaki depolarizasyon olaylarının otomatik olarak algılanması ile birlikte hücre dışı akım kelepçe gerilimi kaydı aracılığıyla motor davranışını ve nöronal alan aktivitesini değerlendirmek için standartlaştırılmış ve basit bir yöntem sağlar.
Açıklamalar
Yazarların açıklayacak bir şeyi yok.
Teşekkürler
Nörofizyoloji deneylerinin yapıldığı ICM elektrofizyoloji platformu personeline teşekkür ederiz. Teknik yardım için Anca Marian'a da teşekkür ederiz. SC Trambolin Hibesi #21488 tarafından desteklendi. EK, AFM Grant #18469 ve ERC Consolidator Grant (ALS-Networks) tarafından desteklendi. HC, Fondation pour la Recherche Médicale (PLP20141031462) ve ARSLA'dan doktora ödülleri ile desteklendi. AD ve RM için bu çalışma Romanya Bilimsel Araştırma ve İnovasyon Ulusal Otoritesi CNCS-UEFISCDI'den (proje numaraları PN-III-P4-ID-PCE-2016-0010, PN-III-P2-2.1-PED-2016-0007, ve COFUND-NEURON-NMDAR-PSY), Avrupa Birliği'nin Horizon 2020 araştırma ve yenilik programı tarafından verilen bir hibe - 668863-SyBil-AA hibe anlaşması ve ABD Hükümeti tarafından finanse edilen bir Ulusal Bilim Vakfı hibesi NSF-IOS-1656830.
Malzemeler
| Name | Company | Catalog Number | Comments |
| Agarose | Sigma-Aldrich, France | A9539 | |
| Aquarium salt | Instant Ocean, Blacksburg, VA | SS15-10 | |
| Borosilicate glass with filament | Sutter Instruments | BF100-50-10 | OD: 1.5mm, ID: 0.5 mm |
| CaCl2 | Sigma-Aldrich, France | C1016 | |
| Depdc5-atg antisense morpholino | GeneTools, OR, USA | N/A | sequence 5’- TGCCTTCATGGTGACCGTCATTTTA -3’ |
| Depdc5-mis antisense morpholino | GeneTools, OR, USA | N/A | sequence 5’- TGCgTTgATcGTGACCcTgATTTTA -3’ |
| Depdc5-splice antisense morpholino | GeneTools, OR, USA | N/A | sequence 5’- ACATTCCTGTTTCACCATAGATGAT -3’ |
| Digitizer | Molecular Devices, CA, USA | Digidata 1550 | |
| Fast Green Dye | Sigma-Aldrich, France | F7258 | Stock solution of 0.2% |
| Glass-bottom petri dishes | Ibidi, Germany | 81218 | |
| Glucose | Sigma-Aldrich, France | 68270 | |
| Grasshopper 2 camera | FLIR, BC, Canada | GRAS-03K2M-C | formerly Point Grey Research |
| HEPES | Sigma-Aldrich, France | H3375 | |
| Human wild-type DEPDC5 cDNA | Dharmacon, France | NM_001242897.1 | Accession: BC144291 Clone ID 905 |
| ImageJ software | NIH, USA | N/A | |
| KCl | Sigma-Aldrich, France | P9333 | |
| Matlab software | MathWorks, MA, USA | N/A | |
| MgCl2 | Sigma-Aldrich, France | M2670 | |
| NaCl | Sigma-Aldrich, France | S7653 | |
| NaOH | Sigma-Aldrich, France | 71687 | |
| Pancuronium bromide | Alomone Labs | P-130 | Stock solution of 60 mM in water |
| Parafilm | Sigma-Aldrich, France | P7793 | |
| Patch clamp amplifier | Molecular Devices, CA, USA | MultiClamp 700B | Computer-controled patch clamp amplifier |
| pClamp10 acquisition software | Molecular Devices | N/A | |
| Pentylenetetrazol (PTZ) | Sigma-Aldrich, France | P6500 | Stock solution of 300 mM (dissolved in recording solution) |
| Pipette puller | Narishige, Japan | PC-10 | |
| Pneumatic PicoPump | WPI, France | PV 820 | |
| Sylgard 184 kit | Sigma-Aldrich Intl. | 761036 | |
| Transfer plastic pipettes | Sigma-Aldrich, France | Z350605 | |
| Zebralab | Viewpoint, France | N/A |
Referanslar
- Kabashi, E., Champagne, N., Brustein, E., Drapeau, P. In the swim of things: Recent insights to neurogenetic disorders from zebrafish. Trends in Genetics. 26 (8), 373-381 (2010).
- Baxendale, S., van Eeden, F., Wilkinson, R. The Power of Zebrafish in personalised medicine. Advances in Experimental Medicine and Biology. , (2017).
- Baraban, S. C., Taylor, M. R., Castro, P. A., Baier, H. Pentylenetetrazole induced changes in zebrafish behavior, neural activity and c-fos expression. Neuroscience. , (2005).
- Cunliffe, V. T. Building a zebrafish toolkit for investigating the pathobiology of epilepsy and identifying new treatments for epileptic seizures. Journal of Neuroscience Methods. , (2016).
- Griffin, A., Krasniak, C., Baraban, S. C. Advancing epilepsy treatment through personalized genetic zebrafish models. Progress in Brain Research. , (2016).
- Griffin, A., Hamling, K. R., Knupp, K., Hong, S. G., Lee, L. P., Baraban, S. C. Clemizole and modulators of serotonin signalling suppress seizures in Dravet syndrome. Brain. , (2017).
- Orellana-Paucar, A. M., et al. Insights from zebrafish and mouse models on the activity and safety of ar-turmerone as a potential drug candidate for the treatment of epilepsy. PLoS ONE. , (2013).
- Baxendale, S., et al. Identification of compounds with anti-convulsant properties in a zebrafish model of epileptic seizures. Disease Models & Mechanisms. , (2012).
- Bar-Peled, L., et al. A tumor suppressor complex with GAP activity for the Rag GTPases that signal amino acid sufficiency to mTORC1. Science. 340 (6136), 1100-1106 (2013).
- Ishida, S., et al. Mutations of DEPDC5 cause autosomal dominant focal epilepsies. Nature Genetics. , (2013).
- Dibbens, L. M., et al. Mutations in DEPDC5 cause familial focal epilepsy with variable foci. Nature Genetics. , (2013).
- Baulac, S., Weckhuysen, S. . DEPDC5-Related Epilepsy. GeneReviews®. , (1993).
- Picard, F., et al. DEPDC5 mutations in families presenting as autosomal dominant nocturnal frontal lobe epilepsy. Neurology. , (2014).
- Teng, Y., et al. Knockdown of zebrafish lgi1a results in abnormal development, brain defects and a seizure-like behavioral phenotype. Human Molecular Genetics. , (2010).
- Baraban, S. C., Dinday, M. T., Hortopan, G. A. Drug screening in Scn1a zebrafish mutant identifies clemizole as a potential Dravet syndrome treatment. Nature Communications. , (2013).
- Suls, A., et al. De novo loss-of-function mutations in CHD2 cause a fever-sensitive myoclonic epileptic encephalopathy sharing features with dravet syndrome. American Journal of Human Genetics. , (2013).
- Grone, B. P., et al. Epilepsy, behavioral abnormalities, and physiological comorbidities in syntaxin-binding protein 1 (STXBP1) mutant zebrafish. PLoS ONE. , (2016).
- de Calbiac, H., et al. DEPDC5 knockdown causes mTOR-dependent motor hyperactivity in zebrafish. Annals of Clinical and Translational Neurology. , (2018).
- Panchaud, N., Péli-Gulli, M. P., De Virgilio, C. Amino acid deprivation inhibits TORC1 through a GTPase-activating protein complex for the Rag family GTPase Gtr1. Science Signaling. , (2013).
- Lim, K. R. Q., Maruyama, R., Yokota, T. Eteplirsen in the treatment of Duchenne muscular dystrophy. Drug Design, Development and Therapy. , (2017).
- Kok, F. O., et al. Reverse genetic screening reveals poor correlation between morpholino-induced and mutant phenotypes in zebrafish. Developmental Cell. , (2015).
- Rossi, A., et al. Genetic compensation induced by deleterious mutations but not gene knockdowns. Nature. , (2015).
- Stainier, D. Y. R., et al. Guidelines for morpholino use in zebrafish. PLoS Genetics. , (2017).
- Kimmel, C. B., Ballard, W. W., Kimmel, S. R., Ullmann, B., Schilling, T. F. Stages of embryonic development of the zebrafish. Developmental dynamics an official public. 203 (3), 253-310 (1995).
- Møller, R. S., Dahl, H. A., Helbig, I. The contribution of next generation sequencing to epilepsy genetics. Expert Review of Molecular Diagnostics. , (2015).
- Allen, A. S., et al. De novo mutations in epileptic encephalopathies. Nature. , (2013).
- Dunn, P., et al. Next generation sequencing methods for diagnosis of epilepsy syndromes. Frontiers in Genetics. , (2018).
- Grone, B. P., Baraban, S. C. Animal models in epilepsy research: Legacies and new directions. Nature Neuroscience. , (2015).
- Zhang, Y., et al. Pharmacological characterization of an antisense knockdown zebrafish model of Dravet syndrome: Inhibition of epileptic seizures by the serotonin agonist fenfluramine. PLoS ONE. , (2015).
- Swaminathan, A., et al. Non-canonical mTOR-Independent Role of DEPDC5 in Regulating GABAergic Network Development. Current Biology. , (2018).
- Samarut, &. #. 2. 0. 1. ;., et al. γ-Aminobutyric acid receptor alpha 1 subunit loss of function causes genetic generalized epilepsy by impairing inhibitory network neurodevelopment. Epilepsia. , 2061-2074 (2018).
- Turrini, L., et al. Optical mapping of neuronal activity during seizures in zebrafish. Scientific Reports. , (2017).
- Rosch, R. E., Hunter, P. R., Baldeweg, T., Friston, K. J., Meyer, M. P. Calcium imaging and dynamic causal modelling reveal brain-wide changes in effective connectivity and synaptic dynamics during epileptic seizures. PLoS Computational Biology. , (2018).
- Baraban, S. C. Forebrain Electrophysiological Recording in Larval Zebrafish. Journal of Visualized Experiments. , (2013).
- Afrikanova, T., et al. Validation of the Zebrafish Pentylenetetrazol Seizure Model: Locomotor versus Electrographic Responses to Antiepileptic Drugs. PLoS ONE. , (2013).
Yeniden Basımlar ve İzinler
Bu JoVE makalesinin metnini veya resimlerini yeniden kullanma izni talebi
Izin talebiDaha Fazla Makale Keşfet
This article has been published
Video Coming Soon
JoVE Hakkında
Telif Hakkı © 2020 MyJove Corporation. Tüm hakları saklıdır