
Method Article
Поведенческий и физиологический анализ в модели эпилепсии рыбок данио
В этой статье
Резюме
Здесь мы представляем протокол разработки и характеристики модели эпилепсии рыбок данио, возникающую в результате преходящего ингибирования гена DEPDC5.
Аннотация
Эпилепсия представляет собой одно из наиболее распространенных неврологических расстройств, затрагивающих, по оценкам, 50 миллионов человек во всем мире. Недавние достижения в области генетических исследований выявили большой спектр генов, причастных к различным формам эпилепсии, подчеркивая гетерогенную природу этого расстройства. Соответствующие животные модели необходимы для исследования патологических механизмов, вызванных генетическими мутациями, замешанными в эпилепсии, и для разработки специализированных, целевых методов лечения. В последние годы рыбки данио стали ценным позвоночным организмом для моделирования эпилепсии с использованием как генетических манипуляций, так и воздействия известных эпилептогенных препаратов, таких как пентилентетразол (PTZ), для выявления новых противоэпилептических терапевтических средств. Вредные мутации в регуляторе mTOR DEPDC5 были связаны с различными формами фокальной эпилепсии и сбитием ортолога рыбок данио, вызывающих гиперактивность, связанную со спонтанными приступообразными эпизодами, а также повышенную электрографическую активность и характерное плавание с поворотным колесом. Здесь мы описали метод, участвующий в создании модели потери функции DEPDC5 и проиллюстрировали протокол оценки двигательной активности через 28 и 48 ч после оплодотворения (hpf), а также метод регистрации полевой активности в зрительном тектуме рыбок данио. Также приведена иллюстрация влияния эпилептогенного препарата PTZ на активность нейронов с течением времени.
Введение
Благодаря своим небольшим размерам, яйцекладу и прозрачности на ранних стадиях развития, рыбка данио стала ценным позвоночным организмом для моделирования заболеваний человека, столь же разнообразных, как сердечно-сосудистые, онкологические или неврологические расстройства1,2. Рыбка данио сочетает в себе преимущества позвоночных, в том числе высокую сохранность архитектуры органов и генетического кода, с небольшим размером и простотой генетических манипуляций с более простыми модельными организмами, что облегчает как фундаментальные исследования, так и трансляционные приложения. В частности, его поддаться высокопроизводительному автоматизированному скринингу поведения и флуоресцентным маркерам клеточных процессов сделала рыбок данио особенно привлекательной моделью для исследований эпилепсии. Это было продемонстрировано высоким увеличением в последнее десятилетие числа публикаций, включающих химически индуцированные и/или генетические модели эпилепсии3,4, 5и, в последнее время, сообщения о перспективныхтерапевтических средствах, полученных из химических экранов в этих моделях6,7,8.
DEPDC5 является членом комплекса GATOR1, отрицательного регулятора сигнализации mTOR9. Мутации в гене DEPDC5 были впервые обнаружены в 2013 году у пробандов, страдающих аутосомно-доминантными фокальными эпилепсиями10,11,и с тех пор были зарегистрированы в ряде клинических состояний, связанных с очаговыми эпилептическими проявлениями и фокальной кортикальной дисплазией12. Подавляющее большинство зарегистрированных мутаций, по прогнозам, вызывают потерю функции гена12,и это было официально продемонстрировано для ряда мутированных транскриптов DEPDC5, которые нацелены на бессмысленный опосредованный распад мРНК12,13. В согласии, сбивание гена-ортолога у рыбок данио с использованием антисмысловых морфолиноолиголидов (АМО) приводит к ряду особенностей, которые являются общими для эпилептических моделей в этом организме, включая гиперактивность, поворотно-подобное плавание, спонтанные судороги и повышенную нейрональную активность14,15,16,17,18. Интересно, что лечение рапамицином, ингибитором передачи сигналов mTOR, обратило вспять поведенческие особенности этой модели18,поддерживая гипотезу о том, что потеря функции DEPDC5 может вызвать эпилепсию из-за неправильной регуляции пути mTOR9,19.
Преходящее сбивание экспрессии генов in vivo с использованием антисмысловых олигонуклеотидов, несущих морфолинодедирование, стало бесценным инструментом для изучения роли специфических генов наравне с методами на основе si/shRNA. В последнее время стратегии, основанные на АМО, также нашли клиническое применение, причем первая АМО-терапия получила одобрение FDA для лечения мышечной атрофии Дюшенна в 2016году 20. Хотя сообщалось, что у рыбок данио фенотип острого нокдауна генов на основе АМО не всегда коррелирует с конститутивными нокаут-моделями21,это может быть связано, по крайней мере, в некоторых случаях с компенсаторными механизмами, порожденными конститутивными генетическими модификациями22. Однако вопрос о специфичности фенотипа, индуцированного АМО, является бесспорным вопросом, который должен быть усердно рассмотрен в исследованиях с использованием этой технологии23. Чтобы обеспечить специфичность нокдаун-фенотипа на основе AMO, необходимо несколько ключевых элементов управления. К ним относится кривая доза-реакция, которая позволяет выбрать самую низкую дозу AMO, эффективную для сбивания генов, избегая общей токсичности из-за введения избытка генетического материала. Использование Mismatch AMO, который не нацелен на какую-либо конкретную область в геноме, также требуется для установления соответствующей дозы и идентификации конкретного фенотипа. Второй AMO, который нацелен на другую область того же гена, такую как AMO, блокирующий сращивание, необходим для подтверждения того, что фенотип обусловлен сносом гена-мишени. Спасение нокдауна фенотипа с кДНК гена, либо человеческого ортолога, либо кодон-модифицированной версии гена рыбки данио, которая не может быть нацелена на AMO, дает веский аргумент в пользу специфичности фенотипа. Отсутствие спасения с той же кДНК, содержащей мутации потери функции (такие как введение кодонов ранней остановки), является еще одним доказательством в этом направлении.
Здесь мы представляем метод генерации модели потери функции рыбок данио DEPDC5 и протокол поведенческого фенотипирования через 28 и 48 ч после оплодотворения (hpf). При 28 л.с. потеря функции DEPDC5 вызывает общую гиперактивность, о чем свидетельствует усиленное сворачивание и подергивание движений эмбрионов внутри хориона. Автоматизированная система обнаружения движения может быть использована на этом этапе для количественной оценки общей активности на эмбрион. При 48 л.с. рыбки данио демонстрируют стереотипное побег, плавая в ответ на прикосновение. У рыбок данио с пониженной экспрессией DEPDC5траектория плавания значительно более извилиста, чем у контрольных, рыба демонстрирует «пробковый винт» или «поворотное колесо», подобное рисунку, аналогичному другим зарегистрированным моделям эпилепсии в этом организме3,4. Электрофизиологические записи были получены в зрительном тектуме у личинок рыбок данио между 4-6 днями после оплодотворения (dpf) и показали базовое увеличение активности нейронов у животных DEPDC5. Преимущество этой модели заключается в том, что она представляет несколько фенотипических особенностей в разные моменты времени, которые могут быть полезны при мониторинге и оценке эффективности лекарственной терапии во время разработки.
протокол
Экспериментальные процедуры были одобрены Национальным и Институциональным этическими комитетами.
1. Переходный снос гена DEPDC5 в эмбрионе рыбки данио
- Подготовка инструментов:
- Приготовьте силиконовые эластомерные инъекционные чашки Петри: смешайте основу и отверждающий агент набора (см. Таблицу материалов)в соотношении 10:1. Наполните смесью чашку Петри толщиной 35 мм наполовину. Подождите, пока кремний затвердеет, прежде чем использовать его (это может занять несколько дней).
- Готовят 1,2 ммоль/л стоковых растворов антисмысловых морфолиноолиголидолидов (АМО; см. Таблицу материалов). Добавьте 250 мкл стерильной воды к 300 нмоль лиофилизированной АМО для получения 1,2 ммоль/л бульонного раствора. Для полного растворения нагревайте флаконы в течение 5 мин при 65 °C. Вихрь кратко. Запечатайте крышку трубки полиэтиленовой пленкой (см. Таблицу материалов).
- Для контрольных спасательных экспериментов подготовьте экспрессионную плазмиду, содержащую человеческую кДНК DEPDC5 (см. Таблицу материалов),клонированную в позвоночнике pCS2 или аналогичную плазмиду экспрессии, совместимую с рыбками данио. В качестве отрицательного контроля в кДНК была введена мутация, вызывающая кодон ранней остановки (p.Arg487*).
- Подготовьте эмбриональную воду: 0,06 г/л аквариумной соли (см. Таблицу материалов)в воде обратного осмоса + 0,5 мг/л метиленового синего.
- В день инъекции приготовьте микроинъекцию боросиликатных стеклянных игл с помощью съемника (см. Таблицу материалов). Установите соответствующие температурные настройки на съемнике иглы. Используйте боросиликатный стеклянный капилляр длиной 10 см, 1/0,5 OD/ID мм, чтобы создать два капилляра ~ 5 см с тонкими наконечниками длиной около 1 см.
- Если кончик иглы очень мелкий, препятствующий выбросу раствора, разбейте самый конец конического наконечника с помощью щипцов под микроскопом.
- Непосредственно перед инъекцией подготовьте рабочие растворы АМО. Всегда готовьте свежий раствор, чтобы обеспечить воспроизводимость результатов. Нагревать флаконы AMO при 65 °C в течение 5 мин. Подготовьте 5 мкл инъекционного образца, содержащего краситель Fast Green (конечная концентрация 0,02%, см. Таблицу материалов)и АМО, разбавленный при рабочей концентрации в воде.
- Определите рабочую концентрацию AMO эмпирически для каждого гена, используя кривую дозового ответа. Рабочая концентрация представляет собой концентрацию, в которой AMO эффективен в сбивающей ген, не вызывая общей токсичности, такой как грубые морфологические дефекты. Как правило, рабочие концентрации AMO будут находиться в диапазоне от 0,2 ммоль/л до 1 ммоль/л (0,4 ммоль/л было определено как эффективная концентрация для данного исследования18). Вводят контрольный морфолин несоответствия в той же концентрации, что и эффективный АМО.
- Вихрьте трубки и центрифугу ненадолго, чтобы довести капли до дна трубок.
- Для спасательных экспериментов подготовьте образец инъекции 5 мкл с АМО, разбавленным в рабочей концентрации, и плазмидой экспрессии кДНК, разбавленной в воде до конечной концентрации, которая будет определена эмпирически. Для экспрессии DEPDC5 и отрицательной контрольной плазмиды 100 нг/мкл было эффективно для фенотипического спасения.
- Подготовка эмбриона:
- За день до микроинъекции установите брачные резервуары для рыбок данио. Утром после инъекции удалите разделители, чтобы обеспечить нерест. Соберите яйца в 100 мм чашки Петри, наполненные эмбриональной водой, используя тонкое сито. Вводят в течение 20-30 мин после сбора, пока яйцеклетки находятся на стадии одной клетки.
- Соберите 60-80 яиц пластиковой пипеткой Пастера и разложите их в чашке Петри с силиконовым покрытием для инъекций. Кремниевая поверхность предотвратит скольжение яиц во время инъекций. Удалите большую часть эмбриональной воды, оставив ровно настолько, чтобы покрыть яйца наполовину.
- Микроинъекции:
- Наполните стеклянную иглу инъекционным раствором. Поместите иглу вертикально в одну из трубок, содержащих инъекционный раствор, следя за тем, чтобы нижний конец иглы касался раствора. Подождите несколько минут, пока цветной раствор для инъекций поднимется на капиллярность и не будет виден на кончике иглы.
- Установите заполненную иглу на инжекторную ручку микроинжектора (см. Таблицу материалов).
- Включите воздушный компрессор и отрегулируйте настройку давления, чтобы создать объем впрыска ~ 2 нЛ.
- Чтобы рассчитать объем вводимого раствора, поместите каплю минерального масла на микротомовую горку. Вводят раствор, содержащий краситель, используя заданные параметры давления и времени. Измерьте диаметр сферы впрыскиваемой жидкости и рассчитайте общий объем по формуле Объем=4/3*π*(d/2)3, где d= измеренный диаметр впрыскиваемого болюса.
- Используя рассекающий бинокулярный микроскоп с 4-кратным увеличением, вводите яйцеклетки на стадии одной клетки, проходя через хорион и желток и проецируя раствор непосредственно внутри клетки.
- Соберите введенные эмбрионы в 100-миллиметровую чашку Петри с эмбриональной водой, пометьте чашку и высиживайте их при 28 °C.
- Убедитесь, что температура инкубатора стабильна с течением времени, так как скорость развития эмбрионов термочувствительна. Например, рост будет ускорен при более высоких температурах, и стадия развития имеет решающее значение для правильной оценки фенотипа24.
- Проверьте качество яйцеклеток через 6-8 ч после инъекции и удалите мертвые и неодобрительные эмбрионы с помощью пластиковой пипетки Пастера.
- На следующее утро подсчитайте и удалите мертвые эмбрионы в каждой посуде пластиковой пипеткой Пастера.
2. Анализ поведения
- Анализ глобальной активности при 28 л.с.:
- Проведите тест во второй половине дня после микроинъекции (28 л.с.), гарантируя, что время суток, в которое проводится тест, соответствует экспериментам для выполнения достоверного статистического анализа, поскольку развитие эмбрионов происходит очень быстро.
- Наполните 35-миллиметровую посуду (тестовую посуду) эмбриональной водой и дайте ей разогреться в инкубаторе (28 °C) в течение не менее 15 минут до начала теста.
- Поместите пластиковую сетку (1,2х1,2 мм), разрезаемую по размеру, на дно тестовой посуды.
- Попрошу другого экспериментатора рандомизировать порядок тестирования эмбрионов и замаскировать названия условий, которые будут тестироваться.
- Убедитесь, что уровень смертности не меняется между состояниями и по сравнению с неинъекционными эмбрионами, чтобы обеспечить специфичность фенотипа. Процент умерших эмбрионов при всех условиях не должен превышать 10-13%18.
- Поместите 10-12 эмбрионов еще в их хорион на пластиковую сетку с помощью пластиковой пипетки Пастера. Наполните тестовую чашку достаточным количеством эмбриональной воды, чтобы эмбрионы были погружены, но не плавали. При необходимости осторожно перемещайте эмбрионы, используя пластиковый наконечник, чтобы расположить их на сетке.
- Используя видеокамеру (см. Таблицу материалов),прикрепленную к рассеченному микроскопу, записывайте спонтанную активность накатки в течение определенного периода времени (10-20-минутных видео обычно достаточно для получения репрезентативных образцов всплесков активности для количественной оценки)
- Верните эмбрионы в соответствующую посуду и поместите их обратно в инкубатор. Повторите эксперимент с таким количеством эмбрионов, сколько необходимо для каждого состояния (как определено анализом мощности 90%).
- Для анализа общего спонтанного движения используйте систему ZebraLab (см. Таблицу материалов). Используя модуль количественной оценки активности, загрузите записанное видео и спроектируйте арены отслеживания вокруг каждого эмбриона по мере необходимости. Установите порог заморозки и разрыва на 10 и 50 соответственно.
- Запустите автоматизированный анализ видео, который количественно оценивает общую активность внутри каждой из определенных арен, затем восстановите набор данных в виде электронной таблицы и выполните анализ с помощью программного обеспечения для анализа данных.
- Сенсорный-вызванный-escape-response (TEER) при 48 л.с.:
- Проводят тест утром через два дня после инъекции (через 48 ч после оплодотворения).
- По крайней мере, за 2 ч до тестирования дехорионировать эмбрионы с помощью тонких щипцов. Убедитесь, что время суток для дехорионации и поведенческого теста согласовано по сравнению с экспериментами.
- Наполните 130-миллиметровую посуду (тестовую посуду) эмбриональной водой и дайте ей нагреться в инкубаторе (28 °C) в течение не менее 15 минут до начала теста.
- Подсчитайте и удалите мертвых и морфологически деформированных личинок. Запишите числа для каждого условия.
- Порубаку другому экспериментатору рандомизировать порядок и кодифицировать названия тестируемых условий.
- Установите камеру (см. Таблицу материалов)над тестовой тарелкой, убедившись, что вся тестовая тарелка находится в поле зрения. Размещение линейки в поле зрения обеспечивает внутреннюю калибровку расстояния.
- С помощью пластиковой пипетки Пастера поместите эмбрион в центр тестовой тарелки и начните запись, используя скорость сбора 30 кадров в секунду.
- Тонким пластиковым наконечником слегка коснитесь хвоста эмбриона мелькающим движением.
- Остановите запись, когда личинка прекратит свое движение.
- Извлеките эмбрион из тестовой посуды и поместите его в новую посуду, наполненную эмбриональной водой. Повторите тест с таким количеством эмбрионов, сколько необходимо для каждого состояния (как определено 90% анализом мощности).
- Верните эмбрионы в исходное блюдо и поместите их обратно в инкубатор.
- Чтобы проанализировать параметры поведения при плавании, загрузите записанное видео в программное обеспечение для анализа ImageJ. Загрузите и установите плагин Ручного отслеживания ImageJ (см. Таблицу материалов). Запустите плагин, выбрав Инструменты | | плагина Ручное отслеживание в меню.
- В диалоговом окне введите калиброванную шкалу изображения. Включение линейки в поле камеры облегчает преобразование см в пиксели.
- Выберите Добавить трек и начните трассировку траектории, нажав на изображение личинки рыбки данио в первом кадре. Кадры автоматически продвигаются вместе с каждой точкой, добавленной в трассировку.
- Продолжайте отслеживать движение до конца эпизода плавания.
- Выберите Конец трека в окне отслеживания, получите координаты X-Y и рассчитайте общее расстояние, скорость и угол поворота.
3. Электрофизиологический анализ
- Подготовка реагентов и инструментов:
- Приготовьте 1% агарозы в эмбриональной воде (см. раздел 1.1.4). Аликвотирует жидкую агарозу в микроцентрифужных трубках и хранит их на нагревательном блоке при 42 °C, чтобы предотвратить затвердевание агарозы.
- Готовят регистрирующий раствор (в ммоль/л): NaCl 134, KCl 2,9, CaCl2 2,1, MgCl2 1,2, глюкоза 10, HEPES 10, рН 7,8.
- Вытягиваем микропипеты из боросиликатного стекла с отверстием наконечника 1,5-2 мкм (5-6 мм2·кг·с−3· Сопротивление−2) неполированное.
- Подготовка личинок рыбок данио к электрофизиологии:
- Поместите рыбу в чашку Петри со стеклянным дном (см. Таблицу материалов)и удалите лишние внеклеточные среды, чтобы убедиться, что рыба поднесена как можно ближе к крышке.
- Используя пластиковую пипетку Пастера, добавьте теплую жидкую агарозу на личинку и вокруг нее. Используйте достаточно агарозы, чтобы покрыть рыбу. Пока агароза затвердевает, используйте тонкие щипцы, чтобы сориентировать рыбу в прямом положении, вентральной стороной вниз, в центре блюда.
- Добавьте 2 мл регистрируемого раствора, содержащего 10 мкМ бромида панкурония (см. Таблицу материалов), чтобыблокировать нервно-мышечную передачу. Добавление паралича необходимо для устранения артефактов из-за небольших движений во время записи.
- Электрофизиологический учет
- Заполните микропипетку записывающим раствором.
- С помощью усилителя патч-зажима (см. Таблицу материалов)в конфигурации зажима напряжения измерьте сопротивление электрода в ванне, чтобы подтвердить его правильное значение.
- Используя 20-кратный объектив, расположите голову личинки в центральном поле зрения и опустите микропипетку, чтобы достичь записывающего положения в мозге, в пределах оптического тектума.
- Переключите усилитель патч-зажима на токовый зажим и зафиксируйте удерживающие токи до 0 мА.
- Используя фильтр нижних частот 1 кГц, скорость сбора 1 кГц и цифровой коэффициент усиления 10, регистрируют спонтанную активность в течение 60 мин для определения базовых уровней активности.
- После 1 ч исходной регистрации добавляют в ванну 200 мкл пентиленететразола (PTZ, см. Таблицу материалов)раствор 300 ммоль/л для получения окончательной концентрации 20 ммоль/л PTZ.
- Записывайте активность нейронов в PTZ еще 120 мин.
- Определение события деполяризации
- Полевая запись событий имеет очень медленную динамику (интересуемые частоты находятся в диапазоне 0,005-0,2с-1). Поэтому фильтруйте сигнал с нижними частотами (Butterworth 5-го порядка LPF при 100 с-1),чтобы избежать сглаживания. Подвыборка записанных данных о напряжении от частоты кадров сбора (в данном случае 1 ks-1)до 250 с-1 (RAW SIGNAL).
- Чтобы определить временные метки для каждого события деполяризации, используйте СИГНАЛ ОБНАРУЖЕНИЯ, который представляет собой отфильтрованную версию записанного сигнала высоких частот (BUTTERworth 1-го порядка HPF при 0,01 с-1).
- Исключая низкочастотные компоненты, обнаружение событий деполяризации может быть выполнено с использованием простого метода порогового значения. Используйте фиксированный порог для устранения шума и обнаружения событий (для этого исследования использовалось 0,3 мВ).
- Охарактеризуйте событие деполяризации серией пересечений пороговых значений, которые происходят через промежутки времени менее 4 с. Вычислите начало и конец события деполяризации, определенные из последовательных последовательностей пересечений пороговых значений. События, которые короче 40 мс, могут быть отброшены как шум.
- Вычислите амплитуду событий в нефильтрованном (RAW SIGNAL) для устранения ошибок, обусловленных влиянием фильтрации нижних частот на пик события. Выберите вейвлет деполяризации из необработанного сигнала, используя временные метки, определенные в отфильтрованном сигнале. Измерьте амплитуду как разницу между максимальным и минимальным значениями вейвлета, выбранного из необработанного сигнала.
ПРИМЕЧАНИЕ: Файлы скриптов для выполнения шага 3.4 — определение события деполяризации — и для получения рисунка 1 представлены в виде дополнительного файла, прилагаемого к этой статье.
Результаты
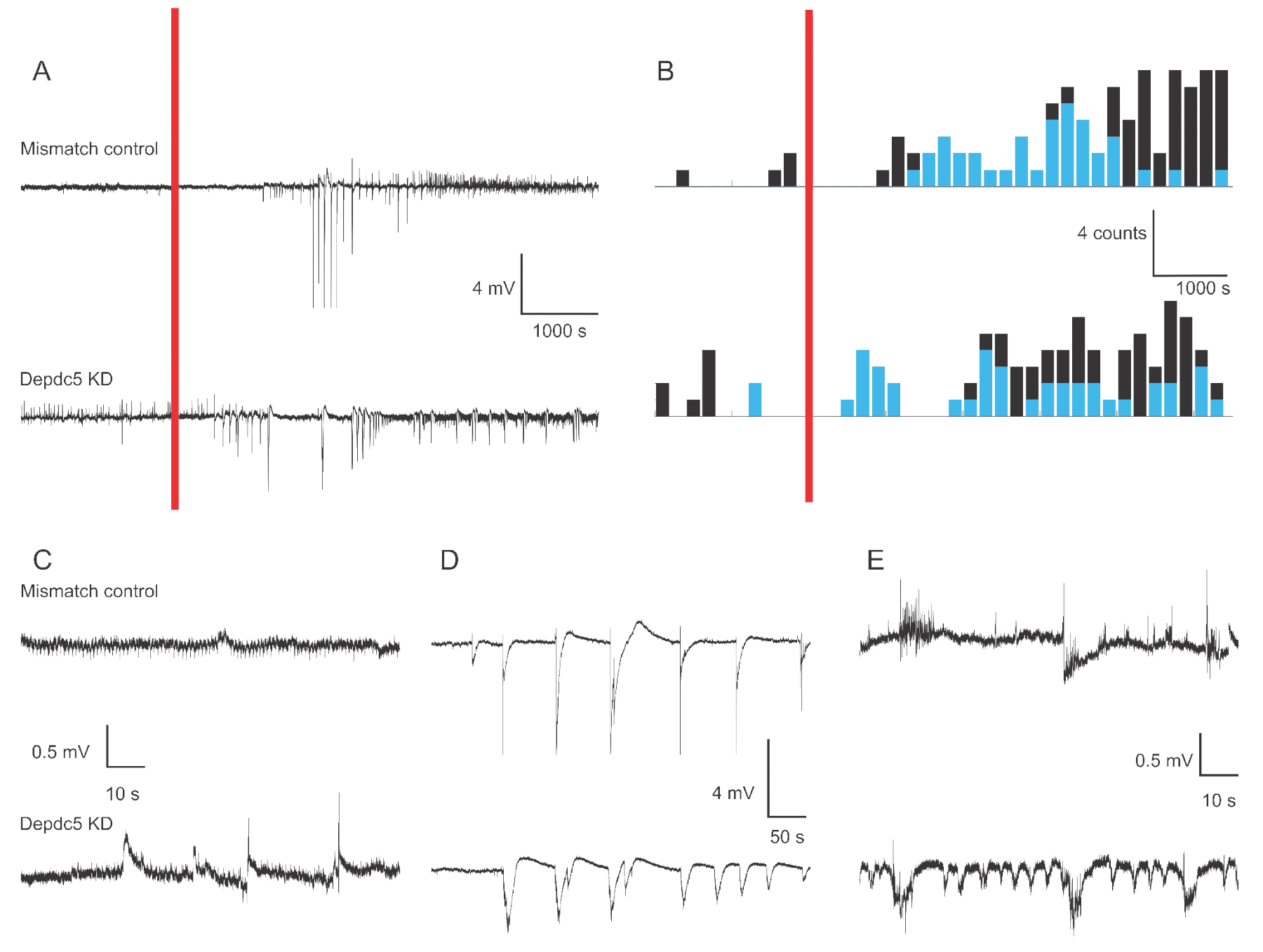
На рисунке 1 показаны репрезентативные следы напряжения 4-6 dpf записей внеклеточного поля личинок рыбок данио в случае двух генетических состояний: контроль несоответствия и депой DEPDC5. В базовом периоде записи декдаун DEPDC5 показывает более высокую распространенность спонтанных событий, в то время как контроль несоответствия отображает очень мало колебаний. Эти паттерны активности являются репрезентативными для значительного увеличения активности нейронов из-за потери функции DEPDC5,как мы ранее сообщали18. После применения PTZ как контроль несоответствия, так и сбитие DEPDC5 показывают увеличение числа событий деполяризации. В течение первого периода после применения PTZ (10 – 60 мин) наблюдается скорость 0,8 событий в минуту как в контроле несоответствия, так и в детонации DEPDC5, где большинство событий имеют высокую амплитуду (>1 мВ). В течение последнего периода отклика (60 – 120 мин после применения PTZ) скорость событий деполяризации увеличивается примерно до 1 события в минуту, и большинство событий имеют низкую амплитуду (≤1 мВ).

Рисунок 1:Пример следов полевых записей в мозге личинок рыбок данио. (A) Обзор 180-минутной записи для контрольной личинки mismatch и DEPDC5 Knock-down. Сначала регистрировалась спонтанная исходная активность, затем В ванне применяли PTZ (красная полоса). (B) Пери-стимульные временные гистограммы событий деполяризации для контроля несоответствия и детонации DEPDC5. События были классифицированы как высокая амплитуда (>1 мВ - синий) и низкая амплитуда (≤1 мВ - черный). (С-Е) Примеры следов различных периодов записи:(C)спонтанная активность,(D)События высокой амплитуды в течение первого периода после применения PTZ,(E)События низкой амплитуды в течение последнего периода после применения PTZ. Обратите внимание, что файлы скриптов для получения этих рисунков предоставляются в виде дополнительного файла. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}
Дополнительный файл: файлы сценариев для шага 3.4. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
Обсуждение
Эпилепсия является сложным неврологическим заболеванием, отличающимся широким спектром этиологий, которые начинают выясняться с появлением технологий генетического секвенирования25,26,27. Универсальные животные модели необходимы для эффективной трансляционной стратегии, которая даст как понимание патологических механизмов генетически связанных эпилепсий, так и таргетную терапию для различных форм этого состояния. Модели рыбок данио были очень эффективны в воспроизведении основных признаков эпилепсии и обеспечении надежных показаний для скрининга противоэпилептических препаратов5,28. Спонтанные судороги могут быть обнаружены у генетически модифицированных рыбок данио15,29,30,31 и нейрофизиологический анализ в этих моделях28 подтвердил нейронную основу эпилептикоподобного поведения32,33. Малогабаритные личинки рыбок данио поддаются химическим экранам в формате 96 скважин с использованием автоматического обнаружения простого поведения, такого как спонтанное плавание, что позволяет быстро обнаруживать потенциальные терапевтические средства.
Представленная здесь модель детона DEPDC5 получена путем инъекции AMO в эмбрион рыбки данио для блокирования экспрессии генов во время развития. Эта модель представляет несколько ключевых фенотипических особенностей в разные временные точки развития личинок, которые могут быть использованы в качестве показателей эффективности терапии во время протокола химического или генетического скрининга. Нокаут гена, опосредованный АМО, является мощным методом, демонстрирующим преимущества перед химически индуцированными моделями судорог, поскольку он специально нацелен на экспрессию интересующего гена, что позволяет идентифицировать основные патогенные механизмы, вызванные генетической мутацией. Химические индуцеры, которые, тем не менее, являются мощными инструментами для скрининга лекарств, могут действовать через несколько клеточных путей, которые не всегда могут быть связаны с изучаемой генетической мутацией. Хотя инъекция АМО сама по себе является простой техникой, когда она осваивается экспериментатором, она также представляет ряд ограничений. Инъекции должны быть выполнены на стадии одной клетки эмбриона; в наших руках инъекции на более поздних стадиях значительно увеличивались вариабельность фенотипа. Это ограничивает время, доступное для инъекций; поэтому полезна стратегия генерации яйцеклеток для инъекций во временной последовательности. Мы обычно используем 4-5 крестов, которые мы открываем с интервалом 15-20 минут, позволяя впрыскивать одно сцепление перед получением следующего. Кроме того, необходимо позаботиться о том, чтобы оценить фенотип в одних и тех же точках между различными экспериментами, поскольку стереотипное поведение быстро развивается в течение первых дней развития. Объем и концентрация АМО также должны тщательно контролироваться, так как общая токсичность из-за инъекций чрезмерных количеств будет маскировать конкретный фенотип. Различные контрольные элементы, представленные во введении, необходимы для определения правильной дозы инъекции и соответствующего фенотипа.
Полевые записи мозга личинок рыбок данио являются полезным инструментом для исследования вредных последствий генетических мутаций, участвующих в различных расстройствах мозга, на глобальную активность нейронов34. События деполяризации, наблюдаемые в этих экспериментальных условиях, являются устоявшимся методом оценки электрофизиологического воздействия лекарственных средств в различных эпилептических условиях15,35. Однако оценка этих эффектов в основном проводилась качественно, а не количественно, и в качестве субъекта анализа был установлен субъективный наблюдатель. Здесь мы разрабатываем автоматическую стратегию обнаружения, которая может объективно количественно оценить скорость деполяризации, их амплитуду и продолжительность, а также может оценить прогресс этих параметров во времени или с различными генетическими или фармакологическими вмешательствами.
Репрезентативные результаты, представленные здесь, показывают ожидаемую полевую активность генетической модели DEPDC5 по сравнению с контролем несоответствия у рыбок данио 4-6 dpf до и после применения PTZ для введения эпилептиформной электрографической активности. Ранее мы показали значительное увеличение базальной активности депонирования DEPDC5 при условии18. Здесь мы показываем, что реакция этих двух условий на PTZ, химический индуктор эпилептиформной активности, имеет аналогичную траекторию во времени, начиная с периода относительно низкой частоты, событий деполяризации с высокой амплитудой и продолжая периодом событий деполяризации с более высокой частотой, более низкой амплитудой. События полевой записи имеют медленную динамику (интересуемые частоты находятся в диапазоне 0,005-0,2с-1),поэтому для изоляции интересующих событий используются как фильтры нижних, так и высоких частот. После устранения низкочастотного шума обнаружение событий деполяризации осуществляется с помощью простого порога. Поскольку на статистику сигнала сильно влияет наличие событий деполяризации, мы не могли использовать стандартное отклонение общего сигнала для определения этого порога. Изменчивость значения стандартного отклонения между наборами данных была больше, чем наблюдаемые уровни регистрируемого шума. Поэтому после визуального осмотра следов мы использовали фиксированное значение порога 0,3 мВ, чтобы избежать смещения, вызванного разными уровнями деполяризационной активности.
Описанный протокол обеспечивает стандартизированный и простой метод оценки двигательного поведения и активности нейронного поля посредством регистрации напряжения внеклеточного тока в сочетании с автоматическим обнаружением событий деполяризации в зрительном тектуме для характеристики эпилептиформоподобных фенотипов в моделях рыбок данио.
Раскрытие информации
Авторам нечего раскрывать.
Благодарности
Мы хотели бы поблагодарить сотрудников электрофизиологической платформы ICM, где проводились нейрофизиологические эксперименты. Мы также благодарим Anca Marian за техническую помощь. SC был поддержан грантом на батуте No 21488. EK был поддержан грантом AFM No 18469 и грантом ERC Consolidator Grant (ALS-Networks). HC был поддержан наградами PhD от Фонда медицинских наук (PLP20141031462) и ARSLA. Для AD и RM эта работа была поддержана тремя грантами от Национального управления по научным исследованиям и инновациям Румынии, CNCS-UEFISCDI (номера проектов PN-III-P4-ID-PCE-2016-0010, PN-III-P2-2.1-PED-2016-0007 и COFUND-NEURON-NMDAR-PSY), грантом исследовательской и инновационной программы Европейского Союза Horizon 2020 - грантовое соглашение No 668863-SyBil-AA и грантом Национального научного фонда NSF-IOS-1656830, финансируемым правительством США.
Материалы
| Name | Company | Catalog Number | Comments |
| Agarose | Sigma-Aldrich, France | A9539 | |
| Aquarium salt | Instant Ocean, Blacksburg, VA | SS15-10 | |
| Borosilicate glass with filament | Sutter Instruments | BF100-50-10 | OD: 1.5mm, ID: 0.5 mm |
| CaCl2 | Sigma-Aldrich, France | C1016 | |
| Depdc5-atg antisense morpholino | GeneTools, OR, USA | N/A | sequence 5’- TGCCTTCATGGTGACCGTCATTTTA -3’ |
| Depdc5-mis antisense morpholino | GeneTools, OR, USA | N/A | sequence 5’- TGCgTTgATcGTGACCcTgATTTTA -3’ |
| Depdc5-splice antisense morpholino | GeneTools, OR, USA | N/A | sequence 5’- ACATTCCTGTTTCACCATAGATGAT -3’ |
| Digitizer | Molecular Devices, CA, USA | Digidata 1550 | |
| Fast Green Dye | Sigma-Aldrich, France | F7258 | Stock solution of 0.2% |
| Glass-bottom petri dishes | Ibidi, Germany | 81218 | |
| Glucose | Sigma-Aldrich, France | 68270 | |
| Grasshopper 2 camera | FLIR, BC, Canada | GRAS-03K2M-C | formerly Point Grey Research |
| HEPES | Sigma-Aldrich, France | H3375 | |
| Human wild-type DEPDC5 cDNA | Dharmacon, France | NM_001242897.1 | Accession: BC144291 Clone ID 905 |
| ImageJ software | NIH, USA | N/A | |
| KCl | Sigma-Aldrich, France | P9333 | |
| Matlab software | MathWorks, MA, USA | N/A | |
| MgCl2 | Sigma-Aldrich, France | M2670 | |
| NaCl | Sigma-Aldrich, France | S7653 | |
| NaOH | Sigma-Aldrich, France | 71687 | |
| Pancuronium bromide | Alomone Labs | P-130 | Stock solution of 60 mM in water |
| Parafilm | Sigma-Aldrich, France | P7793 | |
| Patch clamp amplifier | Molecular Devices, CA, USA | MultiClamp 700B | Computer-controled patch clamp amplifier |
| pClamp10 acquisition software | Molecular Devices | N/A | |
| Pentylenetetrazol (PTZ) | Sigma-Aldrich, France | P6500 | Stock solution of 300 mM (dissolved in recording solution) |
| Pipette puller | Narishige, Japan | PC-10 | |
| Pneumatic PicoPump | WPI, France | PV 820 | |
| Sylgard 184 kit | Sigma-Aldrich Intl. | 761036 | |
| Transfer plastic pipettes | Sigma-Aldrich, France | Z350605 | |
| Zebralab | Viewpoint, France | N/A |
Ссылки
- Kabashi, E., Champagne, N., Brustein, E., Drapeau, P. In the swim of things: Recent insights to neurogenetic disorders from zebrafish. Trends in Genetics. 26 (8), 373-381 (2010).
- Baxendale, S., van Eeden, F., Wilkinson, R. The Power of Zebrafish in personalised medicine. Advances in Experimental Medicine and Biology. , (2017).
- Baraban, S. C., Taylor, M. R., Castro, P. A., Baier, H. Pentylenetetrazole induced changes in zebrafish behavior, neural activity and c-fos expression. Neuroscience. , (2005).
- Cunliffe, V. T. Building a zebrafish toolkit for investigating the pathobiology of epilepsy and identifying new treatments for epileptic seizures. Journal of Neuroscience Methods. , (2016).
- Griffin, A., Krasniak, C., Baraban, S. C. Advancing epilepsy treatment through personalized genetic zebrafish models. Progress in Brain Research. , (2016).
- Griffin, A., Hamling, K. R., Knupp, K., Hong, S. G., Lee, L. P., Baraban, S. C. Clemizole and modulators of serotonin signalling suppress seizures in Dravet syndrome. Brain. , (2017).
- Orellana-Paucar, A. M., et al. Insights from zebrafish and mouse models on the activity and safety of ar-turmerone as a potential drug candidate for the treatment of epilepsy. PLoS ONE. , (2013).
- Baxendale, S., et al. Identification of compounds with anti-convulsant properties in a zebrafish model of epileptic seizures. Disease Models & Mechanisms. , (2012).
- Bar-Peled, L., et al. A tumor suppressor complex with GAP activity for the Rag GTPases that signal amino acid sufficiency to mTORC1. Science. 340 (6136), 1100-1106 (2013).
- Ishida, S., et al. Mutations of DEPDC5 cause autosomal dominant focal epilepsies. Nature Genetics. , (2013).
- Dibbens, L. M., et al. Mutations in DEPDC5 cause familial focal epilepsy with variable foci. Nature Genetics. , (2013).
- Baulac, S., Weckhuysen, S. . DEPDC5-Related Epilepsy. GeneReviews®. , (1993).
- Picard, F., et al. DEPDC5 mutations in families presenting as autosomal dominant nocturnal frontal lobe epilepsy. Neurology. , (2014).
- Teng, Y., et al. Knockdown of zebrafish lgi1a results in abnormal development, brain defects and a seizure-like behavioral phenotype. Human Molecular Genetics. , (2010).
- Baraban, S. C., Dinday, M. T., Hortopan, G. A. Drug screening in Scn1a zebrafish mutant identifies clemizole as a potential Dravet syndrome treatment. Nature Communications. , (2013).
- Suls, A., et al. De novo loss-of-function mutations in CHD2 cause a fever-sensitive myoclonic epileptic encephalopathy sharing features with dravet syndrome. American Journal of Human Genetics. , (2013).
- Grone, B. P., et al. Epilepsy, behavioral abnormalities, and physiological comorbidities in syntaxin-binding protein 1 (STXBP1) mutant zebrafish. PLoS ONE. , (2016).
- de Calbiac, H., et al. DEPDC5 knockdown causes mTOR-dependent motor hyperactivity in zebrafish. Annals of Clinical and Translational Neurology. , (2018).
- Panchaud, N., Péli-Gulli, M. P., De Virgilio, C. Amino acid deprivation inhibits TORC1 through a GTPase-activating protein complex for the Rag family GTPase Gtr1. Science Signaling. , (2013).
- Lim, K. R. Q., Maruyama, R., Yokota, T. Eteplirsen in the treatment of Duchenne muscular dystrophy. Drug Design, Development and Therapy. , (2017).
- Kok, F. O., et al. Reverse genetic screening reveals poor correlation between morpholino-induced and mutant phenotypes in zebrafish. Developmental Cell. , (2015).
- Rossi, A., et al. Genetic compensation induced by deleterious mutations but not gene knockdowns. Nature. , (2015).
- Stainier, D. Y. R., et al. Guidelines for morpholino use in zebrafish. PLoS Genetics. , (2017).
- Kimmel, C. B., Ballard, W. W., Kimmel, S. R., Ullmann, B., Schilling, T. F. Stages of embryonic development of the zebrafish. Developmental dynamics an official public. 203 (3), 253-310 (1995).
- Møller, R. S., Dahl, H. A., Helbig, I. The contribution of next generation sequencing to epilepsy genetics. Expert Review of Molecular Diagnostics. , (2015).
- Allen, A. S., et al. De novo mutations in epileptic encephalopathies. Nature. , (2013).
- Dunn, P., et al. Next generation sequencing methods for diagnosis of epilepsy syndromes. Frontiers in Genetics. , (2018).
- Grone, B. P., Baraban, S. C. Animal models in epilepsy research: Legacies and new directions. Nature Neuroscience. , (2015).
- Zhang, Y., et al. Pharmacological characterization of an antisense knockdown zebrafish model of Dravet syndrome: Inhibition of epileptic seizures by the serotonin agonist fenfluramine. PLoS ONE. , (2015).
- Swaminathan, A., et al. Non-canonical mTOR-Independent Role of DEPDC5 in Regulating GABAergic Network Development. Current Biology. , (2018).
- Samarut, &. #. 2. 0. 1. ;., et al. γ-Aminobutyric acid receptor alpha 1 subunit loss of function causes genetic generalized epilepsy by impairing inhibitory network neurodevelopment. Epilepsia. , 2061-2074 (2018).
- Turrini, L., et al. Optical mapping of neuronal activity during seizures in zebrafish. Scientific Reports. , (2017).
- Rosch, R. E., Hunter, P. R., Baldeweg, T., Friston, K. J., Meyer, M. P. Calcium imaging and dynamic causal modelling reveal brain-wide changes in effective connectivity and synaptic dynamics during epileptic seizures. PLoS Computational Biology. , (2018).
- Baraban, S. C. Forebrain Electrophysiological Recording in Larval Zebrafish. Journal of Visualized Experiments. , (2013).
- Afrikanova, T., et al. Validation of the Zebrafish Pentylenetetrazol Seizure Model: Locomotor versus Electrographic Responses to Antiepileptic Drugs. PLoS ONE. , (2013).
Перепечатки и разрешения
Запросить разрешение на использование текста или рисунков этого JoVE статьи
Запросить разрешениеThis article has been published
Video Coming Soon
Авторские права © 2025 MyJoVE Corporation. Все права защищены