
Method Article
Verhaltens- und physiologische Analyse in einem Zebrafischmodell der Epilepsie
In diesem Artikel
Zusammenfassung
Hier stellen wir ein Protokoll für die Entwicklung und Charakterisierung eines Zebrafischmodells der Epilepsie vor, das aus der vorübergehenden Hemmung des DEPDC5-Gens resultieren.
Zusammenfassung
Epilepsie stellt eine der häufigsten neurologischen Erkrankungen dar und betrifft schätzungsweise 50 Millionen Menschen weltweit. Jüngste Fortschritte in der Genforschung haben ein großes Spektrum von Genen aufgedeckt, die an verschiedenen Formen der Epilepsie beteiligt sind, was die heterogene Natur dieser Störung unterstreicht. Geeignete Tiermodelle sind unerlässlich, um die pathologischen Mechanismen zu untersuchen, die durch genetische Mutationen ausgelöst werden, die mit Epilepsie in Einleiden sind, und um spezialisierte, zielgerichtete Therapien zu entwickeln. In den letzten Jahren hat sich Zebrafisch zu einem wertvollen Wirbeltierorganismus für die Modellierung von Epilepsien entwickelt, wobei sowohl genetische Manipulation als auch die Exposition gegenüber bekannten epileptogenen Medikamenten wie Pentylenetetrazol (PTZ) verwendet wurden, um neuartige antiepileptische Therapeutika zu identifizieren. Schädliche Mutationen im mTOR-Regulator DEPDC5 wurden mit verschiedenen Formen fokaler Epilepsien in Verbindung gebracht und der Knock-Down des Zebrafisch-Orthologs verursacht Hyperaktivität im Zusammenhang mit spontanen anfallsähnlichen Episoden sowie eine erhöhte elektrografische Aktivität und charakteristisches Drehradschwimmen. Hier beschrieben wir die Methode zur Generierung des DEPDC5-Funktionsverlustmodells und veranschaulichen das Protokoll zur Beurteilung der motorischen Aktivität bei 28 und 48 h nach der Befruchtung (hpf) sowie eine Methode zur Aufzeichnung der Feldaktivität im zebrafischen Optischen Tectum. Eine Illustration der Wirkung des epileptogenen Medikaments PTZ auf die neuronale Aktivität im Laufe der Zeit wird ebenfalls bereitgestellt.
Einleitung
Aufgrund seiner geringen Größe, oviparen Entwicklung und Transparenz in frühen Entwicklungsstadien hat sich Zebrafisch zu einem wertvollen Wirbeltierorganismus für die Modellierung menschlicher Krankheiten wie Herz-Kreislauf-, Krebs- oder neurologische Erkrankungen1,2entwickelt. Zebrafisch kombiniert die Vorteile eines Wirbeltiers, einschließlich der hohen Erhaltung der Organarchitektur und des genetischen Codes, mit der geringen Größe und Leichtigkeit der genetischen Manipulation einfacherer Modellorganismen und ermöglicht so sowohl grundlegende Studien als auch translationale Anwendungen. Insbesondere seine Zugänglichkeit zum automatisierten Hochdurchsatz-Screening von Verhalten und fluoreszierenden Markern zellulärer Prozesse hat Zebrafische zu einem besonders attraktiven Modell für die Epilepsieforschung gemacht. Dies wurde durch einen hohen Anstieg der Anzahl von Publikationen mit chemisch induzierten und/oder genetischen Modellen der Epilepsie 3 , 4,5und in jüngerer Zeit Berichte übervielversprechendeTherapeutika aus chemischen Screens in diesen Modellen 6,7,8gezeigt.
DEPDC5 ist ein Mitglied des GATOR1-Komplexes, einem negativen Regulator der mTOR-Signalisierung9. Mutationen im DEPDC5-Gen wurden erstmals 2013 bei Probanden entdeckt, die an autosomal-dominanten fokalen Epilepsien10,11leiden, und wurden seitdem in einer Reihe von klinischen Zuständen berichtet, die mit fokalen epileptischen Manifestationen und fokaler kortikaler Dysplasie assoziiert sind12. Es wird vorhergesagt, dass die große Mehrheit der berichteten Mutationen den Funktionsverlust des Gens12verursacht, und dies wurde formal für eine Reihe von DEPDC5-mutierten Transkripten nachgewiesen, die durch unsinnigen mRNA-Zerfall12,13angegriffen werden. In Übereinstimmung führt der Knock-down des Genorthologs bei Zebrafischen unter Verwendung von Antisense-Morpholino-Oligonukleotiden (AMOs) zu einer Reihe von Merkmalen, die epileptischen Modellen in diesem Organismus gemeinsam sind, einschließlich Hyperaktivität, drehradartiges Schwimmen, spontane Anfälle und erhöhte neuronale Aktivität14,15,16,17,18. Interessanterweise kehrte die Behandlung mit Rapamycin, einem Inhibitor der mTOR-Signalgebung, die Verhaltensmerkmale dieses Modells18um und unterstützte die Hypothese, dass DEPDC5-Funktionsverlust Epilepsie aufgrund einer Fehlregulation des mTOR-Signalwegs auslösen kann9,19.
Der vorübergehende Abbau der Genexpression in vivo unter Verwendung von Antisense-Oligonukleotiden, die die Morpholino-Modifikation tragen, war ein unschätzbares Werkzeug für die Untersuchung der Rolle bestimmter Gene, vergleichbar mit si / shRNA-basierten Techniken. In jüngster Zeit haben AMO-basierte Strategien auch klinische Anwendungen gefunden, wobei eine erste AMO-Therapie 2016 die FDA-Zulassung zur Behandlung der Duchenne-Muskelatrophie erhielt20. Während berichtet wurde, dass bei Zebrafischen der Phänotyp des akuten AMO-basierten Gen-Knock-downs nicht immer mit den konstitutiven Knock-out-Modellen21korreliert, kann dies zumindest in einigen Fällen auf kompensatorische Mechanismen zurückzuführen sein, die durch konstitutive genetische Modifikationen hervorgerufen werden22. Die Frage der Spezifität des AMO-induzierten Phänotyps ist jedoch ein unbestreitbares Anliegen, das in Studien mit dieser Technologie sorgfältig angegangen werden muss23. Um die Spezifität des AMO-basierten Knock-Down-Phänotyps zu gewährleisten, sind mehrere Schlüsselkontrollen notwendig. Dazu gehört eine Dosis-Wirkungs-Kurve, die die Auswahl der niedrigsten Dosis von AMO ermöglicht, die für den Gen-Knock-down wirksam ist, wodurch eine Gesamttoxizität aufgrund der Einführung eines Überschusses an genetischem Material vermieden wird. Die Verwendung eines Mismatch-AMO, das nicht auf eine bestimmte Region im Genom abzielt, ist auch für die Festlegung einer geeigneten Dosis und die Identifizierung eines bestimmten Phänotyps erforderlich. Eine zweite AMO, die auf eine andere Region desselben Gens abzielt, z. B. eine spleißblockierende AMO, ist notwendig, um zu bestätigen, dass der Phänotyp auf den Knock-down des Zielgens zurückzuführen ist. Die Rettung des Knock-Down-Phänotyps mit der cDNA des Gens, entweder dem menschlichen Ortholog oder einer Codon-modifizierten Version des Zebrafisch-Gens, die von der AMO nicht angegriffen werden kann, liefert ein starkes Argument für die Phänotypspezifität. Mangelnde Rettung mit der gleichen cDNA, die Funktionsverlustmutationen enthält (wie die Einführung eines frühen Stopp-Codons), ist ein weiterer Beweis in diese Richtung.
Hier stellen wir eine Methode zur Generierung eines Zebrafisch-DEPDC5-Funktionsverlustmodells und das Protokoll für die Verhaltensphänotypisierung bei 28 und 48 h nach der Befruchtung (hpf) vor. Bei 28 HPF verursacht der Funktionsverlust von DEPDC5 eine allgemeine Hyperaktivität, wie durch verstärkte Wickel- und Zuckungsbewegungen der Embryonen im Chorion belegt wird. Ein automatisiertes Bewegungserkennungssystem kann in diesem Stadium verwendet werden, um die Gesamtaktivität pro Embryo zu quantifizieren. Mit 48 PS zeigen Zebrafische stereotypes Fluchtschwimmen als Reaktion auf Berührung. Bei Zebrafischen mit herunterregulierter Expression von DEPDC5ist die Schwimmbahn deutlich gewundener als bei Kontrollen, wobei die Fische ein "Korkschrauben"- oder "Drehrad"-ähnliches Muster aufweisen, ähnlich wie bei anderen berichteten Epilepsiemodellen in diesem Organismus3,4. Elektrophysiologische Aufzeichnungen wurden im optischen Tectum bei Zebrafischlarven zwischen 4-6 Tagen nach der Befruchtung (dpf) erhalten und zeigen einen Baseline-Anstieg der neuronalen Aktivität bei den DEPDC5-Knock-Down-Tieren. Der Vorteil dieses Modells besteht darin, dass es mehrere phänotypische Merkmale zu verschiedenen Zeitpunkten aufweist, die bei der Überwachung und Bewertung der Wirksamkeit von medikamentösen Therapien während der Entwicklung nützlich sein können.
Protokoll
Die experimentellen Verfahren wurden von den nationalen und institutionellen Ethikkommissionen genehmigt.
1. Transienter Knock-down des DEPDC5-Gens im Zebrafisch-Embryo
- Vorbereitung der Werkzeuge:
- Silikonelastomer-beschichtete Injektions-Petrischalen zubereiten: Mischen Sie die Basis und das Härter des Kits (siehe Materialtabelle)im Verhältnis 10:1. Füllen Sie eine 35 mm Petrischale zur Hälfte mit der Mischung. Warten Sie, bis das Silizium aushärtet, bevor Sie es verwenden (dies kann mehrere Tage dauern).
- Herstellung von 1,2 mmol/L Stammlösungen von Antisense-Morpholino-Oligonukleotiden (AMO; siehe Materialtabelle). 250 μL steriles Wasser werden zu 300 nmole lyophilisiertem AMO hinzugefügt, um eine 1,2 mmol/L Stammlösung zu erhalten. Zur vollständigen Auflösung die Fläschchen 5 min bei 65 °C erhitzen. Vortex kurz. Versiegeln Sie die Rohrkappe mit einer Kunststofffolie (siehe Materialtabelle).
- Für die Kontrollrettungsexperimente bereiten Sie ein Expressionsplasmid vor, das die menschliche cDNA von DEPDC5 (siehe Materialtabelle)enthält, die im pCS2-Backbone oder einem ähnlichen Zebrafisch-kompatiblen Expressionsplasmid geklont ist. Als Negativkontrolle wurde eine Mutation, die ein frühes Stopp-Codon (p.Arg487*) verursacht, in die cDNA eingeführt.
- Embryonenwasser vorbereiten: 0,06 g/L Aquariensalz (siehe Materialtabelle)in Umkehrosmosewasser + 0,5 mg/L Methylenblau.
- Am Tag der Injektion bereiten Sie Mikroinjektions-Borosilikatglasnadeln mit einem Puller vor (siehe Materialtabelle). Stellen Sie geeignete Temperatureinstellungen am Nadelzieher ein. Verwenden Sie eine 10 cm lange, 1/0,5 OD/ID mm Borosilikatglaskapillare, um zwei ~5 cm Kapillaren mit dünnen Spitzen mit einer Länge von ca. 1 cm zu erzeugen.
- Wenn die Spitze der Nadeln sehr fein ist und den Auswurf der Lösung verhindert, brechen Sie das Ende der sich verjüngten Spitze mit einer Handette unter einem Mikroskop.
- Bereiten Sie kurz vor der Injektion die Arbeitslösungen von AMOs vor. Bereiten Sie immer eine frische Lösung vor, um die Reproduzierbarkeit der Ergebnisse zu gewährleisten. Die AMO-Stammfläschchen 5 min lang bei 65 °C erhitzen. Bereiten Sie eine 5 μL Injektionsprobe vor, die Fast Green Farbstoff (0,02% Endkonzentration, siehe Materialtabelle)und die AMO in der Arbeitskonzentration in Wasser verdünnt enthält.
- Bestimmen Sie die Arbeitskonzentration des AMO empirisch für jedes Gen anhand einer Dosis-Wirkungs-Kurve. Die Arbeitskonzentration stellt eine Konzentration dar, bei der die AMO das Gen wirksam niederschlägt, ohne allgemeine Toxizität wie grobe morphologische Defekte zu verursachen. Typischerweise liegen die AMO-Arbeitskonzentrationen in einem Bereich von 0,2 mmol/L bis 1 mmol/L (0,4 mmol/L wurden als effektive Konzentration für diese Studie bestimmt18). Injizieren Sie das Kontroll-Mismatch-Morpholino in der gleichen Konzentration wie das effektive AMO.
- Wirbeln Sie die Röhrchen und zentrifugieren Sie kurz, um die Tröpfchen auf den Boden der Röhrchen zu bringen.
- Für Rettungsexperimente bereiten Sie eine 5 μL Injektionsprobe vor, wobei das AMO in der Arbeitskonzentration verdünnt und das cDNA-Expressionsplasmid in Wasser auf eine empirisch zu bestimmende Endkonzentration verdünnt wird. Für die Expression von DEPDC5 und dem Negativkontrollplasmid waren 100 ng/μL für die phänotypische Rettung wirksam.
- Vorbereitung des Embryos:
- Richten Sie am Tag vor der Mikroinjektion die Zebrafisch-Paarungsbecken ein. Entfernen Sie am Morgen der Injektion die Trennwände, um das Laichen zu ermöglichen. Sammeln Sie die Eier in 100 mm Petrischalen, die mit Embryonenwasser gefüllt sind, mit einem feinen Sieb. Injizieren Sie innerhalb von 20-30 Minuten nach der Entnahme, während sich die Eier im Einzellstadium befinden.
- 60-80 Eier mit einer Pasteur-Pipette aus Kunststoff pflücken und in der silikonbeschichteten Petrischale zur Injektion anrichten. Die Silikonoberfläche verhindert, dass die Eier während der Injektionen gleiten. Entfernen Sie den größten Teil des Embryowassers und lassen Sie gerade genug, um die Eier auf halbem Weg zu bedecken.
- Mikroinjektionen:
- Füllen Sie eine Glasnadel mit Injektionslösung. Legen Sie die Nadel vertikal in eines der Röhrchen, die die Injektionslösung enthalten, und stellen Sie sicher, dass das untere Ende der Nadel die Lösung berührt. Warten Sie einige Minuten, bis die farbige Injektionslösung durch Kapillarität aufsteigt und an der Spitze der Nadel sichtbar ist.
- Montieren Sie die gefüllte Nadel am Injektionsgriff des Mikroinjektors (siehe Materialtabelle).
- Schalten Sie den Luftkompressor ein und stellen Sie die Druckeinstellung ein, um ein Einspritzvolumen von ~ 2 nL zu erzeugen.
- Um das Volumen der injizierten Lösung zu berechnen, legen Sie einen Tropfen Mineralöl auf einen Mikrotom-Objektträger. Injizieren Sie die farbstoffhaltige Lösung mit den eingestellten Druck- und Zeitparametern. Messen Sie den Durchmesser der injizierten Flüssigkeitskugel und berechnen Sie das Gesamtvolumen mit der Formel Volume=4/3*π*(d/2)3, wobei d=der gemessene Durchmesser des injizierten Bolus.
- Mit einem binokularen Seziermikroskop mit einer 4-fachen Vergrößerung injizieren Sie die Eier im Einzelzellstadium, indem Sie durch das Chorion und das Eigelb gehen und die Lösung direkt in die Zelle projizieren.
- Die injizierten Embryonen in einer 100 mm Petrischale mit Embryonenwasser sammeln, die Schale etikettieren und bei 28 °C inkubieren.
- Stellen Sie sicher, dass die Inkubatortemperatur im Laufe der Zeit stabil ist, da die Entwicklungsrate der Embryonen thermosensitiv ist. Zum Beispiel würde das Wachstum bei höheren Temperaturen beschleunigt werden und das Entwicklungsstadium ist entscheidend für die richtige Beurteilung des Phänotyps24.
- Überprüfen Sie die Qualität der Eier 6-8 h nach der Injektion und entfernen Sie tote und unbefruchtete Embryonen mit einer Pasteur-Pipette aus Kunststoff.
- Am nächsten Morgen zählen und entfernen Sie tote Embryonen in jeder Schüssel mit einer Pasteur-Pipette aus Kunststoff.
2. Verhaltensanalyse
- Globale Aktivitätsanalyse bei 28 HPF:
- Führen Sie den Test am Nachmittag des Tages nach der Mikroinjektion (28 HPF) durch und stellen Sie sicher, dass die Tageszeit, zu der der Test durchgeführt wird, im Gegensatz zu Experimenten konsistent ist, um eine gültige statistische Analyse durchzuführen, da die Entwicklung der Embryonen sehr schnell ist.
- Füllen Sie eine 35-mm-Schale (Testschale) mit Embryowasser und lassen Sie sie vor Beginn des Tests mindestens 15 Minuten im Inkubator (28 °C) aufwärmen.
- Legen Sie ein auf Maß zugeschnittenes Kunststoffgitter (1,2 x 1,2 mm) auf den Boden der Testschale.
- Lassen Sie einen anderen Experimentator die Reihenfolge der Tests der Embryonen randomisieren und die Namen der zu testenden Bedingungen maskieren.
- Stellen Sie sicher, dass sich die Sterblichkeitsrate zwischen den Bedingungen und im Vergleich zu nicht injizierten Embryonen nicht ändert, um die Spezifität des Phänotyps sicherzustellen. Der Prozentsatz der toten Embryonen darf unter allen Bedingungen 10-13%18nicht überschreiten.
- Legen Sie 10-12 Embryonen, die sich noch in ihrem Chorion befindet, mit einer Pasteur-Pipette aus Kunststoff auf das Kunststoffnetz. Füllen Sie die Testschale mit genügend Embryowasser, um die Embryonen unter Wasser zu halten, aber nicht schwimmend. Bewegen Sie die Embryonen bei Bedarf vorsichtig mit einer Kunststoffspitze, um sie auf dem Gitter zu positionieren.
- Zeichnen Sie mit einer Videokamera (siehe Materialtabelle),die an einem Präpariermikroskop befestigt ist, die spontane Wickelaktivität für eine definierte Zeit auf (10-20 min lange Videos reichen in der Regel aus, um repräsentative Proben von Aktivitätsausbrüchen für die Quantifizierung zu erhalten)
- Bringen Sie die Embryonen in ihre jeweilige Schale zurück und legen Sie sie wieder in den Inkubator. Wiederholen Sie das Experiment mit so vielen Embryonen, wie für jede Bedingung benötigt werden (wie durch eine 90% ige Leistungsanalyse bestimmt).
- Um die gesamte spontane Bewegung zu analysieren, verwenden Sie ein ZebraLab-System (siehe Materialtabelle). Laden Sie mit dem Aktivitätsquantifizierungsmodul das aufgezeichnete Video hoch und entwerfen Sie die Tracking-Arenen um jeden Embryo herum. Legen Sie den Freeze- und Burst-Schwellenwert auf 10 bzw. 50 fest.
- Führen Sie die automatisierte Videoanalyse durch, die die Gesamtaktivität in jeder der definierten Arenen quantifiziert, stellen Sie dann den Datensatz als Tabellenkalkulation wieder her und führen Sie die Analyse mit einer Datenanalysesoftware durch.
- Touch-Evoked-Escape-Response (TEER) bei 48 PSF:
- Führen Sie den Test morgens zwei Tage nach der Injektion durch (48 h nach der Befruchtung).
- Mindestens 2 h vor dem Test die Embryonen mit einer feinen Zette enthorionieren. Stellen Sie sicher, dass die Tageszeit für die Dehorionierung und den Verhaltenstest im Laufe der Experimente konsistent ist.
- Füllen Sie eine 130-mm-Schale (Testschale) mit Embryowasser und lassen Sie sie vor Beginn des Tests mindestens 15 Minuten im Inkubator (28 °C) aufwärmen.
- Zählen und entfernen Sie tote und morphologisch deformierte Larven. Notieren Sie die Zahlen für jede Bedingung.
- Lassen Sie einen anderen Experimentator die Reihenfolge randomisieren und die Namen der zu testenden Bedingungen kodifizieren.
- Montieren Sie die Kamera (siehe Materialtabelle)über der Testschale und stellen Sie sicher, dass sich die gesamte Testschale im Sichtfeld befindet. Das Platzieren eines Lineals innerhalb des Sichtfelds ermöglicht eine interne Kalibrierung für die Entfernung.
- Legen Sie mit einer Pasteur-Pipette aus Kunststoff einen Embryo in die Mitte der Testschale und beginnen Sie die Aufnahme mit einer Erfassungsrate von 30 fps.
- Berühren Sie mit einer feinen Plastikspitze leicht den Schwanz des Embryos mit einer flackernden Bewegung.
- Stoppen Sie die Aufnahme, wenn die Larve ihre Bewegung beendet hat.
- Entfernen Sie den Embryo aus der Testschale und legen Sie ihn in eine neue Schüssel, die mit Embryonenwasser gefüllt ist. Wiederholen Sie den Test mit so vielen Embryonen, wie für jede Bedingung erforderlich sind (wie durch eine 90% ige Leistungsanalyse bestimmt).
- Bringen Sie die Embryonen in ihre ursprüngliche Schale zurück und legen Sie sie wieder in den Inkubator.
- Um die Parameter des Schwimmverhaltens zu analysieren, laden Sie das aufgezeichnete Video in die ImageJ-Analysesoftware. Laden Sie das Manuelle Tracking-Plugin von ImageJ herunter und installieren Sie es (siehe Materialtabelle). Starten Sie das Plugin, indem Sie Tools | Plugin-| Manuelles Tracking im Menü.
- Geben Sie im Dialogfenster den kalibrierten Maßstab des Bildes ein. Die Aufnahme eines Lineals in das Kamerafeld erleichtert die Konvertierung von cm in Pixel.
- Wählen Sie Spur hinzufügen und starten Sie die Trajektorienverfolgung, indem Sie im ersten Bild auf das Bild der Zebrafischlarve klicken. Die Frames werden automatisch mit jedem Punkt gerechnet, der der Ablaufverfolgung hinzugefügt wird.
- Verfolgen Sie die Bewegung bis zum Ende der Schwimmepisode weiter.
- Wählen Sie Spur im Tracking-Fenster beenden, rufen Sie die X-Y-Koordinaten ab und berechnen Sie die Gesamtentfernung, Geschwindigkeit und den Wendewinkel.
3. Elektrophysiologische Analyse
- Reagenz- und Werkzeugvorbereitung:
- 1% Agarose in Embryowasser zubereiten (siehe Abschnitt 1.1.4). Aliquotieren Sie die flüssige Agarose in Mikrozentrifugenröhrchen und halten Sie diese auf einem Heizblock bei 42 °C, um ein Aushärten der Agarose zu verhindern.
- Bereiten Sie die Aufzeichnungslösung (in mmol/L) vor: NaCl 134, KCl 2.9, CaCl2 2.1, MgCl2 1.2, Glucose 10, HEPES 10, pH 7.8.
- Ziehen Sie Borosilikatglas-Mikropipetten mit einer Spitzenöffnung von 1,5-2 μm (5-6 Mm2·kg·s−3· A−2 Widerstand) unpoliert.
- Vorbereitung von Zebrafischlarven für die Elektrophysiologie:
- Legen Sie den Fisch in eine Petrischale mit Glasboden (siehe Materialtabelle)und entfernen Sie überschüssige extrazelluläre Medien, um sicherzustellen, dass der Fisch so nah wie möglich an den Deckzettel gebracht wird.
- Fügen Sie mit einer Pasteur-Pipette aus Kunststoff warme flüssige Agarose auf und um die Larve hinzu. Verwenden Sie gerade genug Agarose, um den Fisch zu bedecken. Während die Agarose aushärtet, verwenden Sie eine feine Zette, um den Fisch in einer geraden Position, ventral mit der Seite nach unten, in der Mitte des Gerichts zu orientieren.
- 2 ml der Aufzeichnungslösung, die 10 μM Pancuroniumbromid enthält (siehe Materialtabelle),um die neuromuskuläre Übertragung zu blockieren. Die Zugabe des Lähmungsmittels ist notwendig, um Artefakte aufgrund kleiner Bewegungen während der Aufnahmen zu eliminieren.
- Elektrophysiologische Aufzeichnung
- Füllen Sie die Mikropipette mit Aufnahmelösung.
- Messen Sie mit dem Patch-Clamp-Verstärker (siehe Materialtabelle)in Spannungsklemmenkonfiguration den Elektrodenwiderstand im Bad, um seinen korrekten Wert zu bestätigen.
- Positionieren Sie mit einem 20-fachen Objektiv den Kopf der Larve im zentralen Sichtfeld und senken Sie die Mikropipette, um die Aufnahmeposition im Gehirn innerhalb des optischen Tectums zu erreichen.
- Schalten Sie den Patch-Clamp-Verstärker auf Stromzange um und fixieren Sie den Haltestrom auf 0 mA.
- Mit einem Tiefpassfilter von 1 kHz, einer Erfassungsrate von 1 kHz und einer digitalen Verstärkung von 10 zeichnen Sie spontane Aktivität für 60 Minuten auf, um das Basisaktivitätsniveau zu bestimmen.
- Nach 1 h Basislinienaufzeichnung wird 200 μL Pentylentetrazol (PTZ, siehe Materialtabelle)Lösung 300 mmol/L in das Bad für eine Endkonzentration von 20 mmol/L PTZ geben.
- Zeichnen Sie die neuronale Aktivität in PTZ für weitere 120 Minuten auf.
- Bestimmung des Depolarisationsereignisses
- Feldaufzeichnungsereignisse haben eine sehr langsame Dynamik (frequenzen von Interesse liegen im Bereich von 0,005-0,2 s-1). Filtern Sie daher das Signal mit einem Tiefpass (Butterworth 5. Ordnung LPF bei 100 s-1),um Aliasing zu vermeiden. Subsample die aufgezeichneten Spannungsdaten von der Erfassungsbildrate (in diesem Fall 1 ks-1) bis hinunter zu 250 s-1 (RAW SIGNAL).
- Um die Zeitstempel für jedes Depolarisationsereignis zu identifizieren, verwenden Sie ein DETECTION SIGNAL, eine hochpassgefilterte Version des aufgezeichneten Signals (Butterworth HPF 1. Ordnung bei 0,01 s-1).
- Durch die Eliminierung der niederfrequenten Komponenten kann die Detektion von Depolarisationsereignissen mit einer einfachen Schwellenwertmethode durchgeführt werden. Verwenden Sie einen festen Schwellenwert für die Geräuschbeseitigung und Ereigniserkennung (0,3 mV wurden für diese Studie verwendet).
- Charakterisieren Sie das Depolarisationsereignis durch eine Reihe von Schwellenwertüberschreitungen, die in Zeitintervallen kleiner als 4 s auftreten. Berechnen Sie den Beginn und das Ende des Depolarisationsereignisses, wie sie aus aufeinanderfolgenden Sequenzen von Schwellenwertüberschreitungen ermittelt wurden. Ereignisse, die kürzer als 40 ms sind, können als Rauschen verworfen werden.
- Berechnen Sie die Amplitude der Ereignisse im ungefilterten (RAW-SIGNAL), um Fehler aufgrund der Auswirkungen der Tiefpassfilterung auf den Höhepunkt des Ereignisses zu eliminieren. Wählen Sie das Depolarisations-Wavelet aus dem Rohsignal mit den im gefilterten Signal ermittelten Zeitstempeln aus. Messen Sie die Amplitude als Differenz zwischen den Maximal- und Minimalwerten des aus dem Rohsignal ausgewählten Wavelets.
HINWEIS: Die Skriptdateien zum Ausführen von Schritt 3.4 – Bestimmung des Depolarisationsereignisses – und zum Abrufen von Abbildung 1 werden als Ergänzende Datei im Anhang zu diesem Artikel bereitgestellt.
Ergebnisse
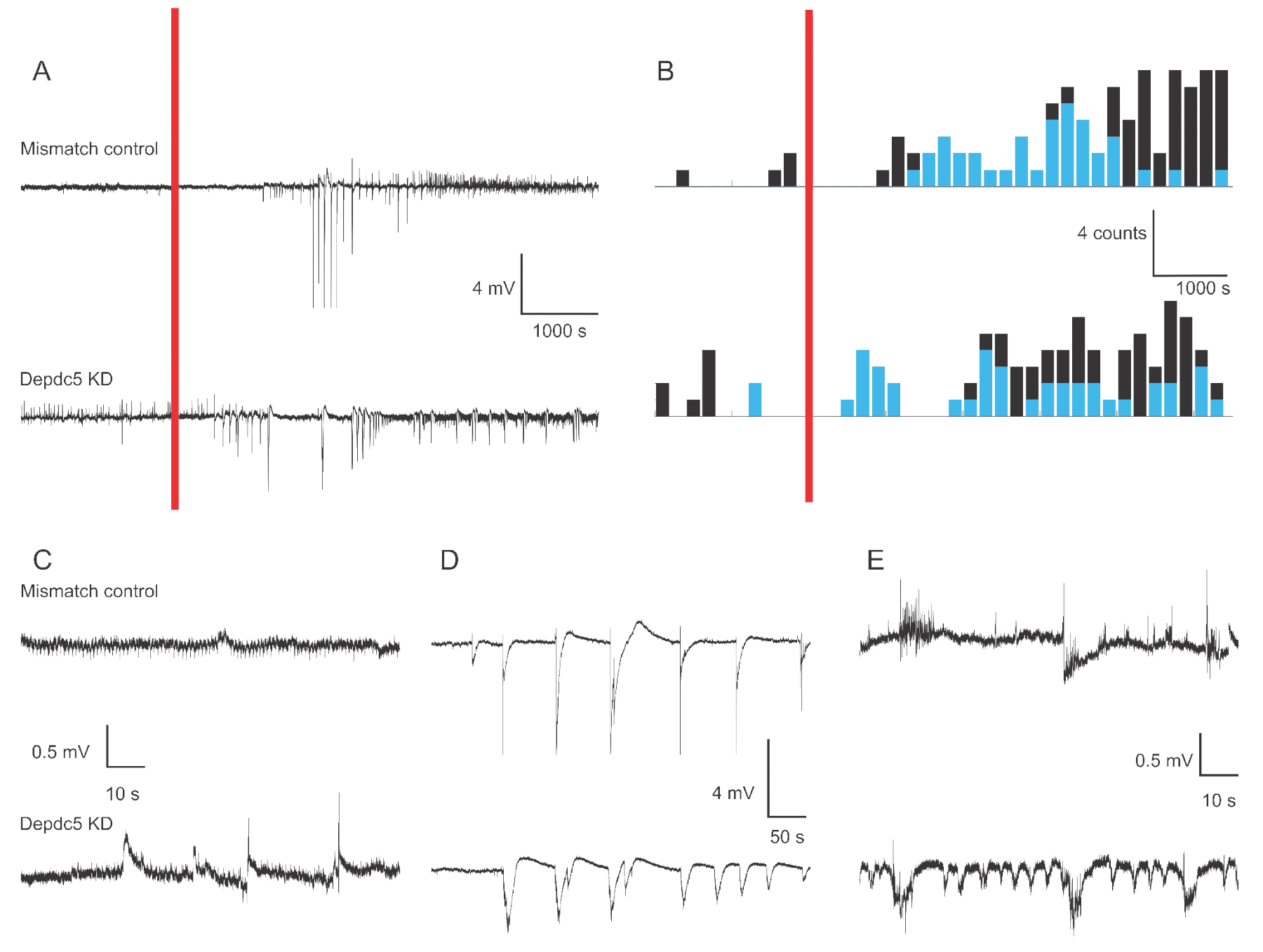
Abbildung 1 zeigt repräsentative Spannungsspuren von 4-6 dpf Zebrafischlarven extrazellulären Feldaufnahmen bei zwei genetischen Bedingungen: Mismatch Control und DEPDC5 Knock-Down. Im Basiszeitraum der Aufzeichnung zeigt der DEPDC5-Knock-Down ein höheres Auftreten spontaner Ereignisse, während das Mismatch-Steuerelement nur sehr wenige Schwankungen anzeigt. Diese Aktivitätsmuster sind repräsentativ für den signifikanten Anstieg der neuronalen Aktivität aufgrund des Funktionsverlusts von DEPDC5, wie wir bereits berichtet haben18. Nach der PTZ-Anwendung zeigen sowohl die Mismatch-Steuerung als auch der DEPDC5-Knock-Down eine erhöhte Anzahl von Depolarisationsereignissen. Während der ersten Periode nach der PTZ-Anwendung (10 – 60 min) wird eine Rate von 0,8 Ereignissen pro Minute sowohl bei der Mismatch-Kontrolle als auch beim DEPDC5-Knock-Down beobachtet, wobei die Mehrheit der Ereignisse eine hohe Amplitude (>1 mV) aufwiesen. Während der letzteren Ansprechphase (60 – 120 min nach PTZ-Anwendung) steigt die Rate der Depolarisationsereignisse auf etwa 1 Ereignis pro Minute an, und die Mehrheit der Ereignisse hat eine geringe Amplitude (≤1 mV).

Abbildung 1: Beispiel Spuren von Feldaufnahmen im Gehirn der Zebrafischlarven. (A) Übersicht über 180-minütige Aufzeichnung für eine Mismatch-Kontrolllarve und einen DEPDC5 Knock-down. Zuerst wurde die spontane Baseline-Aktivität aufgezeichnet, dann wurde PTZ im Bad (roter Balken) angewendet. (B) Peristimulationszeit histogramme der Depolarisationsereignisse für Mismatch-Kontrolle und DEPDC5-Knock-Down. Die Ereignisse wurden als hohe Amplitude (>1 mV - blau) und niedrige Amplitude (≤1 mV - schwarz) klassifiziert. (C-E) Beispielspuren der verschiedenen Perioden der Aufzeichnung: (C) spontane Aktivität, (D) Ereignisse mit hoher Amplitude während der ersten Periode nach PTZ-Anwendung, (E) Ereignisse mit niedriger Amplitude während der letzten Periode nach PTZ-Anwendung. Beachten Sie, dass die Skriptdateien zum Abrufen dieser Zahlen als Zusatzdateibereitgestellt werden. Bitte klicken Sie hier, um eine größere Version dieser Abbildung anzuzeigen.
{kind=link}
Zusatzdatei: Skriptdateien für Schritt 3.4. Bitte klicken Sie hier, um eine größere Version dieser Abbildung anzuzeigen.
Diskussion
Epilepsie ist eine komplexe neurologische Erkrankung mit einer breiten Palette von Ätiologien, die mit dem Aufkommen genetischer Sequenzierungstechnologien aufgeklärt werden25,26,27. Vielseitige Tiermodelle sind essentiell für eine effiziente translationale Strategie, die sowohl Einblicke in die pathologischen Mechanismen genetisch bedingter Epilepsien als auch gezielte Therapien für die verschiedenen Formen dieser Erkrankung liefert. Zebrafischmodelle waren sehr effektiv bei der Reproduktion von Hauptmerkmalen der Epilepsie und lieferten zuverlässige Auslesungen für das Screening von Antiepileptika5,28. Spontane Anfälle können bei genetisch veränderten Zebrafischen15,29,30,31 nachgewiesen werden und die neurophysiologische Analyse in diesen Modellen28 hat die neuronale Grundlage des epileptischen Verhaltensbestätigt 32,33. Kleine Zebrafischlarven sind für chemische Screens im 96-Well-Format durch automatisierte Erkennung von einfachem Verhalten wie spontanem Schwimmen geeignet, was eine schnelle Erkennung potenzieller Therapeutika ermöglicht.
Das hier vorgestellte DEPDC5-Knock-Down-Modell wird durch Injektion von AMO in den Zebrafischembryo erhalten, um die Genexpression während der Entwicklung zu blockieren. Dieses Modell präsentiert mehrere phänotypische Schlüsselmerkmale während verschiedener Zeitpunkte der Larvenentwicklung, die als Indikatoren für die Therapieeffizienz während eines chemischen oder genetischen Screening-Protokolls verwendet werden können. Der AMO-vermittelte Gen-Knock-down ist eine leistungsfähige Technik, die Vorteile gegenüber chemisch induzierten Anfallsmodellen aufweist, da sie spezifisch auf die Expression eines interessierenden Gens abzielt und so die Identifizierung der zugrunde liegenden pathogenen Mechanismen ermöglicht, die durch eine genetische Mutation ausgelöst werden. Chemische Induktoren, die dennoch potente Werkzeuge für Arzneimittel-Screenings sind, können über mehrere zelluläre Wege wirken, die für die untersuchte genetische Mutation möglicherweise nicht immer relevant sind. Während die AMO-Injektion an sich eine einfache Technik ist, wenn sie vom Experimentator beherrscht wird, stellt sie auch eine Reihe von Einschränkungen dar. Die Injektionen müssen im Einzellstadium des Embryos durchgeführt werden; in unseren Händen erhöhten Injektionen in späteren Stadien die Variabilität des Phänotyps erheblich. Dies begrenzt die für die Injektion verfügbare Zeit; Daher ist eine Strategie zur Erzeugung von Eiern zur Injektion in einer Zeitsequenz nützlich. Wir verwenden routinemäßig 4-5 Kreuze, die wir in Abständen von 15-20 Minuten öffnen, so dass eine Kupplung einspritzt werden kann, bevor die nächste erhalten wird. Darüber hinaus muss darauf geachtet werden, den Phänotyp gleichzeitig zwischen verschiedenen Experimenten zu bewerten, da sich stereotype Verhaltensweisen in den ersten Tagen der Entwicklung schnell entwickeln. Das Volumen und die Konzentration von AMOs müssen ebenfalls sorgfältig kontrolliert werden, da die allgemeine Toxizität aufgrund übermäßiger Mengen den spezifischen Phänotyp maskiert. Die verschiedenen Kontrollen, die in der Einleitung vorgestellt werden, sind für die Bestimmung der richtigen Injektionsdosis und des entsprechenden Phänotyps unerlässlich.
Feldaufnahmen des Larvenhirnfischgehirns sind ein nützliches Werkzeug, um die schädlichen Auswirkungen genetischer Mutationen, die an verschiedenen Hirnerkrankungen beteiligt sind, auf die globale neuronale Aktivität zu untersuchen34. Depolarisationsereignisse, die unter diesen experimentellen Bedingungen beobachtet werden, sind eine etablierte Methode zur Beurteilung der elektrophysiologischen Wirkungen von Arzneimitteln unter verschiedenen epileptischenErkrankungen 15,35. Die Bewertung dieser Effekte erfolgte jedoch meist qualitativ und nicht quantitativ und mit einem subjektiven Beobachter als Akteur in der Analyse. Hier entwickeln wir eine automatische Nachweisstrategie, die die Rate der Depolarisationen, ihre Amplitude und Dauer objektiv quantifizieren und den Fortschritt dieser Parameter im Laufe der Zeit oder mit verschiedenen genetischen oder pharmakologischen Interventionen bewerten kann.
Die hier vorgestellten repräsentativen Ergebnisse zeigen die erwartete Feldaktivität des DEPDC5-Knock-Down-Genmodells im Vergleich zu einer Mismatch-Kontrolle bei 4-6 dpf Zebrafischen vor und nach der Anwendung von PTZ zur Einführung epileptiformer elektrografischer Aktivität. Zuvor haben wir eine signifikante Zunahme der Basalaktivität der DEPDC5-Knockdown-Bedingung 18gezeigt. Hier zeigen wir, dass die Reaktion dieser beiden Bedingungen auf PTZ, einen chemischen epileptiformen Aktivitätsinduktor, eine ähnliche Zeitbahn hat, beginnend mit einer Periode relativ niederfrequenter Depolarisationsereignisse mit hoher Amplitude und einer Periode höherfrequenter Depolarisationsereignisse mit niedrigerer Amplitude. Feldaufzeichnungsereignisse haben eine langsame Dynamik (frequenzen von Interesse liegen im Bereich von 0,005-0,2 s-1),daher werden in diesem Protokoll sowohl Tiefpass- als auch Hochpassfilter verwendet, um die interessierenden Ereignisse zu isolieren. Nach Beseitigung des niederfrequenten Rauschens erfolgt die Erkennung von Depolarisationsereignissen mit einem einfachen Schwellenwert. Da die Statistik des Signals stark durch das Vorhandensein von Depolarisationsereignissen beeinflusst wird, konnten wir die Standardabweichung des Gesamtsignals nicht verwenden, um diesen Schwellenwert zu bestimmen. Die Variabilität des Wertes der Standardabweichung über Datensätze hinweg war größer als die beobachteten Aufzeichnungsgeräusche. Daher haben wir nach der visuellen Inspektion der Spuren einen festen Wert des Schwellenwerts von 0,3 mV verwendet, um die durch unterschiedliche Depolarisationsaktivitäten induzierte Verzerrung zu vermeiden.
Das beschriebene Protokoll bietet eine standardisierte und einfache Methode zur Bewertung des motorischen Verhaltens und der neuronalen Feldaktivität über extrazelluläre Stromzangenspannungsaufzeichnung gekoppelt mit automatischer Erkennung von Depolarisationsereignissen im optischen Tectum, um epileptiforme Phänotypen in Zebrafischmodellen zu charakterisieren.
Offenlegungen
Die Autoren haben nichts preiszugeben.
Danksagungen
Wir danken den Mitarbeitern der ICM-Elektrophysiologie-Plattform, auf der die neurophysiologischen Experimente durchgeführt wurden. Wir danken auch Anca Marian für die technische Hilfe. SC wurde durch den Trampoline Grant #21488 unterstützt. EK wurde durch den AFM Grant #18469 und den ERC Consolidator Grant (ALS-Networks) unterstützt. HC wurde durch PhD-Preise der Fondation pour la Recherche Médicale (PLP20141031462) und ARSLA unterstützt. Für AD und RM wurde diese Arbeit durch drei Zuschüsse der rumänischen Nationalen Behörde für wissenschaftliche Forschung und Innovation, CNCS-UEFISCDI (Projektnummern PN-III-P4-ID-PCE-2016-0010, PN-III-P2-2.1-PED-2016-0007 und COFUND-NEURON-NMDAR-PSY), einen Zuschuss des Forschungs- und Innovationsprogramms Horizon 2020 der Europäischen Union – Finanzhilfevereinbarung Nr. 668863-SyBil-AA und einen von der US-Regierung finanzierten Zuschuss der National Science Foundation NSF-IOS-1656830 unterstützt.
Materialien
| Name | Company | Catalog Number | Comments |
| Agarose | Sigma-Aldrich, France | A9539 | |
| Aquarium salt | Instant Ocean, Blacksburg, VA | SS15-10 | |
| Borosilicate glass with filament | Sutter Instruments | BF100-50-10 | OD: 1.5mm, ID: 0.5 mm |
| CaCl2 | Sigma-Aldrich, France | C1016 | |
| Depdc5-atg antisense morpholino | GeneTools, OR, USA | N/A | sequence 5’- TGCCTTCATGGTGACCGTCATTTTA -3’ |
| Depdc5-mis antisense morpholino | GeneTools, OR, USA | N/A | sequence 5’- TGCgTTgATcGTGACCcTgATTTTA -3’ |
| Depdc5-splice antisense morpholino | GeneTools, OR, USA | N/A | sequence 5’- ACATTCCTGTTTCACCATAGATGAT -3’ |
| Digitizer | Molecular Devices, CA, USA | Digidata 1550 | |
| Fast Green Dye | Sigma-Aldrich, France | F7258 | Stock solution of 0.2% |
| Glass-bottom petri dishes | Ibidi, Germany | 81218 | |
| Glucose | Sigma-Aldrich, France | 68270 | |
| Grasshopper 2 camera | FLIR, BC, Canada | GRAS-03K2M-C | formerly Point Grey Research |
| HEPES | Sigma-Aldrich, France | H3375 | |
| Human wild-type DEPDC5 cDNA | Dharmacon, France | NM_001242897.1 | Accession: BC144291 Clone ID 905 |
| ImageJ software | NIH, USA | N/A | |
| KCl | Sigma-Aldrich, France | P9333 | |
| Matlab software | MathWorks, MA, USA | N/A | |
| MgCl2 | Sigma-Aldrich, France | M2670 | |
| NaCl | Sigma-Aldrich, France | S7653 | |
| NaOH | Sigma-Aldrich, France | 71687 | |
| Pancuronium bromide | Alomone Labs | P-130 | Stock solution of 60 mM in water |
| Parafilm | Sigma-Aldrich, France | P7793 | |
| Patch clamp amplifier | Molecular Devices, CA, USA | MultiClamp 700B | Computer-controled patch clamp amplifier |
| pClamp10 acquisition software | Molecular Devices | N/A | |
| Pentylenetetrazol (PTZ) | Sigma-Aldrich, France | P6500 | Stock solution of 300 mM (dissolved in recording solution) |
| Pipette puller | Narishige, Japan | PC-10 | |
| Pneumatic PicoPump | WPI, France | PV 820 | |
| Sylgard 184 kit | Sigma-Aldrich Intl. | 761036 | |
| Transfer plastic pipettes | Sigma-Aldrich, France | Z350605 | |
| Zebralab | Viewpoint, France | N/A |
Referenzen
- Kabashi, E., Champagne, N., Brustein, E., Drapeau, P. In the swim of things: Recent insights to neurogenetic disorders from zebrafish. Trends in Genetics. 26 (8), 373-381 (2010).
- Baxendale, S., van Eeden, F., Wilkinson, R. The Power of Zebrafish in personalised medicine. Advances in Experimental Medicine and Biology. , (2017).
- Baraban, S. C., Taylor, M. R., Castro, P. A., Baier, H. Pentylenetetrazole induced changes in zebrafish behavior, neural activity and c-fos expression. Neuroscience. , (2005).
- Cunliffe, V. T. Building a zebrafish toolkit for investigating the pathobiology of epilepsy and identifying new treatments for epileptic seizures. Journal of Neuroscience Methods. , (2016).
- Griffin, A., Krasniak, C., Baraban, S. C. Advancing epilepsy treatment through personalized genetic zebrafish models. Progress in Brain Research. , (2016).
- Griffin, A., Hamling, K. R., Knupp, K., Hong, S. G., Lee, L. P., Baraban, S. C. Clemizole and modulators of serotonin signalling suppress seizures in Dravet syndrome. Brain. , (2017).
- Orellana-Paucar, A. M., et al. Insights from zebrafish and mouse models on the activity and safety of ar-turmerone as a potential drug candidate for the treatment of epilepsy. PLoS ONE. , (2013).
- Baxendale, S., et al. Identification of compounds with anti-convulsant properties in a zebrafish model of epileptic seizures. Disease Models & Mechanisms. , (2012).
- Bar-Peled, L., et al. A tumor suppressor complex with GAP activity for the Rag GTPases that signal amino acid sufficiency to mTORC1. Science. 340 (6136), 1100-1106 (2013).
- Ishida, S., et al. Mutations of DEPDC5 cause autosomal dominant focal epilepsies. Nature Genetics. , (2013).
- Dibbens, L. M., et al. Mutations in DEPDC5 cause familial focal epilepsy with variable foci. Nature Genetics. , (2013).
- Baulac, S., Weckhuysen, S. . DEPDC5-Related Epilepsy. GeneReviews®. , (1993).
- Picard, F., et al. DEPDC5 mutations in families presenting as autosomal dominant nocturnal frontal lobe epilepsy. Neurology. , (2014).
- Teng, Y., et al. Knockdown of zebrafish lgi1a results in abnormal development, brain defects and a seizure-like behavioral phenotype. Human Molecular Genetics. , (2010).
- Baraban, S. C., Dinday, M. T., Hortopan, G. A. Drug screening in Scn1a zebrafish mutant identifies clemizole as a potential Dravet syndrome treatment. Nature Communications. , (2013).
- Suls, A., et al. De novo loss-of-function mutations in CHD2 cause a fever-sensitive myoclonic epileptic encephalopathy sharing features with dravet syndrome. American Journal of Human Genetics. , (2013).
- Grone, B. P., et al. Epilepsy, behavioral abnormalities, and physiological comorbidities in syntaxin-binding protein 1 (STXBP1) mutant zebrafish. PLoS ONE. , (2016).
- de Calbiac, H., et al. DEPDC5 knockdown causes mTOR-dependent motor hyperactivity in zebrafish. Annals of Clinical and Translational Neurology. , (2018).
- Panchaud, N., Péli-Gulli, M. P., De Virgilio, C. Amino acid deprivation inhibits TORC1 through a GTPase-activating protein complex for the Rag family GTPase Gtr1. Science Signaling. , (2013).
- Lim, K. R. Q., Maruyama, R., Yokota, T. Eteplirsen in the treatment of Duchenne muscular dystrophy. Drug Design, Development and Therapy. , (2017).
- Kok, F. O., et al. Reverse genetic screening reveals poor correlation between morpholino-induced and mutant phenotypes in zebrafish. Developmental Cell. , (2015).
- Rossi, A., et al. Genetic compensation induced by deleterious mutations but not gene knockdowns. Nature. , (2015).
- Stainier, D. Y. R., et al. Guidelines for morpholino use in zebrafish. PLoS Genetics. , (2017).
- Kimmel, C. B., Ballard, W. W., Kimmel, S. R., Ullmann, B., Schilling, T. F. Stages of embryonic development of the zebrafish. Developmental dynamics an official public. 203 (3), 253-310 (1995).
- Møller, R. S., Dahl, H. A., Helbig, I. The contribution of next generation sequencing to epilepsy genetics. Expert Review of Molecular Diagnostics. , (2015).
- Allen, A. S., et al. De novo mutations in epileptic encephalopathies. Nature. , (2013).
- Dunn, P., et al. Next generation sequencing methods for diagnosis of epilepsy syndromes. Frontiers in Genetics. , (2018).
- Grone, B. P., Baraban, S. C. Animal models in epilepsy research: Legacies and new directions. Nature Neuroscience. , (2015).
- Zhang, Y., et al. Pharmacological characterization of an antisense knockdown zebrafish model of Dravet syndrome: Inhibition of epileptic seizures by the serotonin agonist fenfluramine. PLoS ONE. , (2015).
- Swaminathan, A., et al. Non-canonical mTOR-Independent Role of DEPDC5 in Regulating GABAergic Network Development. Current Biology. , (2018).
- Samarut, &. #. 2. 0. 1. ;., et al. γ-Aminobutyric acid receptor alpha 1 subunit loss of function causes genetic generalized epilepsy by impairing inhibitory network neurodevelopment. Epilepsia. , 2061-2074 (2018).
- Turrini, L., et al. Optical mapping of neuronal activity during seizures in zebrafish. Scientific Reports. , (2017).
- Rosch, R. E., Hunter, P. R., Baldeweg, T., Friston, K. J., Meyer, M. P. Calcium imaging and dynamic causal modelling reveal brain-wide changes in effective connectivity and synaptic dynamics during epileptic seizures. PLoS Computational Biology. , (2018).
- Baraban, S. C. Forebrain Electrophysiological Recording in Larval Zebrafish. Journal of Visualized Experiments. , (2013).
- Afrikanova, T., et al. Validation of the Zebrafish Pentylenetetrazol Seizure Model: Locomotor versus Electrographic Responses to Antiepileptic Drugs. PLoS ONE. , (2013).
Nachdrucke und Genehmigungen
Genehmigung beantragen, um den Text oder die Abbildungen dieses JoVE-Artikels zu verwenden
Genehmigung beantragenWeitere Artikel entdecken
This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. Alle Rechte vorbehalten