
Method Article
Análise comportamental e fisiológica em um modelo de zebrafish de epilepsia
Neste Artigo
Resumo
Aqui, apresentamos um protocolo para o desenvolvimento e a caracterização de um modelo de zebrafish de epilepsia resultante da inibição transitória do gene DEPDC5.
Resumo
A epilepsia representa uma das doenças neurológicas mais comuns, afetando cerca de 50 milhões de pessoas em todo o mundo. Avanços recentes na pesquisa genética descobriram um grande espectro de genes implicados em várias formas de epilepsia, destacando a natureza heterogênea deste transtorno. Modelos animais adequados são essenciais para investigar os mecanismos patológicos desencadeados por mutações genéticas implicadas na epilepsia e para o desenvolvimento de terapias especializadas e direcionadas. Nos últimos anos, o zebrafish emergiu como um valioso organismo vertebrado para modelar epilepsias, com o uso de manipulação genética e exposição a drogas epiléptogênicas conhecidas, como pentilenotetrazol (PTZ), para identificar novas terapêuticas antiepilépticas. Mutações deletérios no regulador mTOR DEPDC5 têm sido associadas a várias formas de epilepsias focais e a derrubada da ortopedia de zebrafish causa hiperatividade associada a episódios semelhantes a convulsões espontâneas, bem como atividade eletrográfica aprimorada e natação característica da roda. Aqui, descrevemos o método envolvido na geração do modelo de perda de função DEPDC5 e ilustramos o protocolo de avaliação da atividade motora em 28 e 48 horas de fertilização pós-fertilização (hpf), bem como um método para registro da atividade de campo no tectum óptico zebrafish. Uma ilustração do efeito da droga epieptogênica PTZ sobre a atividade neuronal ao longo do tempo também é fornecida.
Introdução
Devido ao seu pequeno tamanho, desenvolvimento oviparas e transparência nos estágios iniciais do desenvolvimento, o zebrafish emergiu como um valioso organismo vertebrado para modelar doenças humanas tão diversas quanto doenças cardiovasculares, cancerígenas ou neurológicas1,2. O zebrafish combina as vantagens de um vertebrado, incluindo a alta conservação da arquitetura de órgãos e código genético, com o pequeno tamanho e facilidade de manipulação genética de organismos modelos mais simples, facilitando, assim, tanto estudos fundamentais quanto aplicações translacionais. Em particular, sua comodidade à triagem automatizada de comportamento e marcadores fluorescentes de processos celulares tornou o zebrafish um modelo particularmente atraente para a pesquisa de epilepsia. Isso foi demonstrado por um alto aumento na última década do número de publicações com modelos quimicamente induzidos e/ou genéticos de epilepsia3,4,5 e, mais recentemente, relatos de terapêuticas promissoras obtidas a partir de telas químicas nesses modelos6,7,8.
O DEPDC5 é membro do complexo GATOR1, um regulador negativo da sinalização mTOR9. Mutações no gene DEPDC5 foram descobertas pela primeira vez em 2013 em probandes que sofrem de epilepsias focais autossômicas dominantes10,11, e desde então foram relatadas em uma série de condições clínicas associadas a manifestações epilépticas focais e displasia cortical12. Prevê-se que a grande maioria das mutações relatadas causem a perda de função do gene12, e isso foi formalmente demonstrado para uma série de transcrições mutadas DEPDC5 que são alvo de decadência mRNA mediada sem sentido12,13. De acordo com, a derrubada da ortopedia genética em zebrafish usando oligonucleotídeos de morfolino antiscenos (AMOs) resulta em uma série de características comuns a modelos epilépticos neste organismo, incluindo hiperatividade, natação em forma de roda, convulsões espontâneas e atividade neuronal aprimorada14,15,16,17,18. Curiosamente, o tratamento com rapamicina, um inibidor da sinalização mTOR, inverteu as características comportamentais deste modelo18,apoiando a hipótese de que a perda de função de DEPDC5 pode desencadear a epilepsia devido a uma má regulação da via mTOR9,19.
A derrubada transitória da expressão genética in vivo usando oligonucleotídeos antissentidos que carregam a modificação da morfolino tem sido uma ferramenta inestimável para estudar o papel de genes específicos, em comparação com as técnicas baseadas em si/shRNA. Recentemente, estratégias baseadas em AMO também encontraram aplicações clínicas, com uma primeira terapia AMO recebendo a aprovação da FDA para o tratamento da atrofia muscular de Duchenne em 201620. Embora tenha sido relatado que, em zebrafish, o fenótipo de knock-down genético agudo baseado em AMO nem sempre se correlaciona com os modelos de knock-out constitutivos21, isso pode ser devido, pelo menos em alguns casos, a mecanismos compensatórios gerados por modificações genéticas constitutivas22. No entanto, a questão da especificidade do fenótipo induzido pela AMO é uma preocupação indiscutível que deve ser diligentemente abordada em estudos utilizando essa tecnologia23. Para garantir a especificidade do fenótipo de knock-down baseado em AMO, vários controles-chave são necessários. Estes incluem uma curva dose-resposta que permite a seleção da menor dose de AMO eficaz para knock-down genético, evitando a toxicidade geral devido à introdução de um excesso de material genético. O uso de um AMO incompatível que não visa nenhuma região específica no genoma também é necessário para estabelecer uma dose apropriada e na identificação de um fenótipo específico. Um segundo AMO que tem como alvo uma região diferente do mesmo gene, como uma AMO de bloqueio de emendas, é necessário para confirmar que o fenótipo é devido à derrubada do gene alvo. O resgate do fenótipo derrubado com o cDNA do gene, seja o ortopedia humano ou uma versão modificada por codon do gene zebrafish que não pode ser alvo da AMO, fornece um forte argumento a favor da especificidade do fenótipo. A falta de resgate com o mesmo cDNA contendo mutações de perda de função (como a introdução de um códons de parada precoce) é mais uma prova nesta direção.
Aqui, apresentamos um método para gerar um modelo de perda de função de zebrafish DEPDC5 e o protocolo para fenotipagem comportamental em 28 e 48 h pós fertilização (hpf). Com 28 cvf, a perda de função DEPDC5 causa hiperatividade geral, como evidenciado pelo aumento dos movimentos de enrolamento e contração dos embriões dentro do acorde. Um sistema automatizado de detecção de movimento pode ser usado nesta fase para quantificar a atividade global por embrião. Com 48 hpf, zebrafish exibem fuga estereotipada natação em resposta ao toque. Em zebrafish com expressão de baixa regulagem do DEPDC5, a trajetória de natação é significativamente mais tortuosa do que nos controles, o peixe exibindo um "parafuso de cortiça" ou "roda-de-giro" como padrão, semelhante a outros modelos de epilepsia relatados neste organismo3,4. Os registros eletrofisiológicos foram obtidos no tecto óptico em larvas de zebrafish entre 4-6 dias após a fertilização (dpf) e mostram um aumento da atividade neuronal nos animais de knock-down DEPDC5. A vantagem deste modelo é que ele apresenta várias características fenotípicas em diferentes momentos, o que pode ser útil no monitoramento e avaliação da eficácia das terapias medicamentosas durante o desenvolvimento.
Protocolo
Os procedimentos experimentais foram aprovados pelos Comitês de Ética Nacional e Institucional.
1. Derrubada transitória do gene DEPDC5 em embrião de zebrafish
- Elaboração de ferramentas:
- Prepare pratos de Petri revestidos de elastômero de silício: Misture a base e o agente de cura do kit (ver Tabela de Materiais) em uma proporção de 10:1. Encha uma placa de Petri de 35 mm no meio do caminho com a mistura. Aguarde o silício endurecer antes de usá-lo (isso pode levar vários dias).
- Preparar soluções de estoque de 1,2 mmol/L de oligonucleotídeos de morfolino antissense (AMO; ver Tabela de Materiais). Adicione 250 μL de água estéril a 300 nmole liophilized AMO para obter uma solução de estoque de 1,2 mmol/L. Para dissolução completa, aqueça os frascos por 5 min a 65 °C. Vórtice brevemente. Sele a tampa do tubo com uma película plástica (ver Tabela de Materiais).
- Para os experimentos de resgate de controle, prepare uma expressão plasmid contendo o cDNA humano de DEPDC5 (ver Tabela de Materiais) clonado na espinha dorsal pCS2 ou um plasmídeo de expressão compatível com zebrafish semelhante. Como um controle negativo, uma mutação causando um códon de parada precoce (p.Arg487*) foi introduzida no cDNA.
- Prepare a água do embrião: 0,06 g/L de sal de aquário (ver Tabela de Materiais) em água de osmose reversa + 0,5 mg/L azul de metileno.
- No dia da injeção, prepare agulhas de vidro borossilicação de microinjeção usando um puxador (ver Tabela de Materiais). Estabeleça as configurações de temperatura apropriadas no puxador de agulhas. Use um capilar de vidro borossilicato de vidro de 10 cm de comprimento, 1/0,5 OD/ID mm para gerar dois capilares de ~5 cm com pontas finas com um comprimento de aproximadamente 1 cm.
- Se a ponta das agulhas estiver muito fina, impedindo a ejeção da solução, quebre a extremidade da ponta afilada usando fórceps sob um microscópio.
- Pouco antes da injeção, prepare as soluções de trabalho dos AMOs. Prepare sempre uma nova solução para garantir a reprodutibilidade dos resultados. Aqueça os frascos amo a 65 °C por 5 min. Prepare uma amostra de injeção de 5 μL contendo corante Fast Green (concentração final de 0,02%, consulte Tabela de Materiais) e a AMO diluída na concentração de trabalho na água.
- Determine a concentração de trabalho da AMO empiricamente para cada gene usando uma curva de resposta de dose. A concentração de trabalho representa uma concentração onde a AMO é eficaz na derrubada do gene sem causar toxicidade geral, como defeitos morfológicos brutos. Normalmente, as concentrações de trabalho amo estarão em uma faixa de 0,2 mmol/L a 1 mmol/L (0,4 mmol/L foi determinada como a concentração efetiva para este estudo18). Injete o controle Incompatível morfolino na mesma concentração que o AMO eficaz.
- Vórtice os tubos e centrífugas brevemente para trazer as gotículas para o fundo dos tubos.
- Para experimentos de resgate, prepare uma amostra de injeção de 5 μL com a AMO diluída na concentração de trabalho e a expressão cDNA plasmid diluída na água para uma concentração final a ser determinada empiricamente. Para a expressão de DEPDC5 e o plasmídeo de controle negativo, 100 ng/μL foi eficaz para o resgate fenotípico.
- Preparação do embrião:
- Um dia antes da microinjeção, configure os tanques de acasalamento de zebrafish. Na manhã da injeção, remova os divisores para permitir a desova. Colete os ovos em pratos petri de 100 mm cheios de água embrionária usando uma peneira fina. Injete dentro de 20-30 minutos da coleta, enquanto os ovos estão em um estágio celular.
- Escolha 60-80 ovos com uma pipeta pasteur de plástico e organize-os na placa de Petri revestida de silício para injeção. A superfície de silício evitará que os ovos deslizem durante as injeções. Remova a maior parte da água do embrião, deixando o suficiente para cobrir os ovos no meio do caminho.
- Microinjeções:
- Encha uma agulha de vidro com solução de injeção. Coloque a agulha verticalmente em um dos tubos contendo a solução de injeção, garantindo que a extremidade inferior da agulha esteja tocando a solução. Aguarde vários minutos até que a solução de injeção colorida suba pela capilaridade e seja visível na ponta da agulha.
- Monte a agulha cheia na alça de injeção do microinjetor (ver Tabela de Materiais).
- Ligue o compressor de ar e ajuste a configuração de pressão para gerar um volume de injeção de ~2 nL.
- Para calcular o volume da solução injetada, coloque uma gota de óleo mineral em um slide de microtome. Injete a solução contendo corantes usando os parâmetros de pressão e tempo definidos. Meça o diâmetro da esfera do fluido injetado e calcule o volume total utilizando a fórmula Volume=4/3*π*(d/2)3, onde d=o diâmetro medido do bolus injetado.
- Usando um microscópio binóculo dissecando com uma ampliação 4X, injete os ovos no estágio celular único passando pelo acorde e pela gema, e projetando a solução diretamente dentro da célula.
- Colete os embriões injetados em uma placa de Petri de 100 mm com água de embrião, rotule a placa e incuba-os a 28 °C.
- Certifique-se de que a temperatura da incubadora é estável ao longo do tempo, pois a taxa de desenvolvimento dos embriões é termosensível. Por exemplo, o crescimento seria acelerado a temperaturas mais altas e o estágio de desenvolvimento é fundamental para avaliar adequadamente o fenótipo24.
- Verifique a qualidade dos ovos 6-8 h após a injeção e remova embriões mortos e não fertilizados usando uma pipeta pasteur de plástico.
- Na manhã seguinte, conte e remova embriões mortos em cada prato com uma pipeta pasteur de plástico.
2. Análise de Comportamento
- Análise de atividade global a 28 hpf:
- Realizar o teste na tarde do dia após a microinjeção (28 hpf), garantindo que a hora do dia em que o teste é realizado seja consistente sobre experimentos para realizar análise estatística válida, pois o desenvolvimento de embriões é muito rápido.
- Encha um prato de 35 mm (prato de teste) com água de embrião e deixe aquecer na incubadora (28 °C) por pelo menos 15 minutos antes de iniciar o teste.
- Coloque uma grade de malha plástica (1,2x1,2 mm) cortada em tamanho, na parte inferior da placa de teste.
- Que outro experimentador randomize a ordem de testes dos embriões e mascarar os nomes das condições a serem testadas.
- Certifique-se de que a taxa de mortalidade não muda entre as condições e em comparação com embriões não injetados para garantir a especificidade do fenótipo. O percentual de embriões mortos em todas as condições não deve exceder 10-13%18.
- Coloque 10-12 embriões ainda dentro de seu acorde na malha plástica usando uma pipeta pasteur de plástico. Encha o prato de teste com água de embrião suficiente para manter os embriões submersos, mas não flutuando. Se necessário, mova os embriões com cuidado usando uma ponta plástica para posicioná-los na grade.
- Usando uma câmera de vídeo (ver Tabela de Materiais) anexada a um microscópio de dissecção, registo da atividade de enrolamento espontâneo por um período de tempo definido (vídeos longos de 10 a 20 minutos geralmente são suficientes para obter amostras representativas de rajadas de atividade para a quantificação)
- Devolva os embriões ao seu respectivo prato e coloque-os de volta na incubadora. Repita o experimento com o máximo de embriões necessários para cada condição (conforme determinado por uma análise de energia de 90%).
- Para analisar o movimento espontâneo total, utilize um sistema ZebraLab (ver Tabela de Materiais). Usando o módulo de quantificação de atividade, carregue o vídeo gravado e projete as arenas de rastreamento ao redor de cada embrião conforme apropriado. Defina o limiar de congelamento e estouro para 10 e 50, respectivamente.
- Execute a análise de vídeo automatizada, que quantifica a atividade total dentro de cada uma das arenas definidas, em seguida, recupere o conjunto de dados como uma planilha e realize a análise usando um software de análise de dados.
- Touch-Evoked-Escape-Response (TEER) a 48 hpf:
- Realizar o teste pela manhã dois dias após a injeção (48h após a fertilização).
- Pelo menos 2 h antes do teste, descorionato os embriões usando fórceps finos. Certifique-se de que a hora do dia para a descorção e o teste de comportamento é consistente em relação aos experimentos.
- Encha um prato de 130 mm (prato de teste) com água de embrião e deixe aquecer na incubadora (28 °C) por pelo menos 15 minutos antes de iniciar o teste.
- Conte e remova larvas mortas e morfologicamente deformadas. Registo os números de cada condição.
- Peça a outro experimentador para randomizar a ordem e codificar os nomes das condições a serem testadas.
- Monte a câmera (ver Tabela de Materiais) sobre a prato de teste certificando-se de que toda a prato de teste está dentro do campo de visão. Colocar uma régua dentro do campo de visão fornece uma calibração interna para a distância.
- Com uma pipeta pasteur de plástico, coloque um embrião no centro da folha de teste e comece a gravação usando uma taxa de aquisição de 30 fps.
- Com uma ponta plástica fina, toque ligeiramente a cauda do embrião com um movimento de movimento.
- Pare a gravação quando a larva terminar seu movimento.
- Retire o embrião do prato de teste e coloque-o em um novo prato cheio de água embrionária. Repita o teste com o máximo de embriões necessários para cada condição (conforme determinado por uma análise de energia de 90%).
- Devolva os embriões ao prato original e coloque-os de volta na incubadora.
- Para analisar os parâmetros do comportamento da natação, carregue o vídeo gravado para o software de análise ImageJ. Baixe e instale o plugin de rastreamento manual do ImageJ (ver Tabela de Materiais). Inicie o plugin escolhendo ferramentas | | plugin Acompanhamento Manual no menu.
- Na janela de diálogo, introduza a escala calibrada da imagem. Incluir uma régua no campo da câmera facilita a conversão de cm para pixels.
- Selecione Adicionar faixa e iniciar o traçado da trajetória clicando na imagem da larva de zebrafish no primeiro quadro. Os quadros avançam automaticamente com cada ponto adicionado ao traço.
- Continue rastreando o movimento até o final do episódio de natação.
- Selecione A faixa final na janela de rastreamento, recupere as coordenadas X-Y e calcule a distância total, velocidade e ângulo de giro.
3. Análise Eletrofisiológica
- Reagente e preparação de ferramentas:
- Prepare 1% de agarose em água de embrião (ver seção 1.1.4). Alíquotar o líquido surgiu em tubos de microcentrifuuge e mantê-los em um bloco de aquecimento a 42 °C para evitar que a agarose endureça.
- Prepare a solução de gravação (em mmol/L): NaCl 134, KCl 2.9, CaCl2 2.1, MgCl2 1.2, glicose 10, HEPES 10, pH 7.8.
- Puxe micropipettos de vidro borossilicatos com uma abertura de ponta de 1,5-2 μm (5-6 Mm2·kg·s−3· Umaresistência −2) não polida.
- Preparação de larvas de zebrafish para eletrofisiologia:
- Coloque o peixe em uma placa de Petri de fundo de vidro (ver Tabela de Materiais) e remova o excesso de mídia extracelular para garantir que o peixe seja trazido o mais perto possível do deslizamento da tampa.
- Usando uma pipeta pasteur de plástico, adicione líquido quente em e ao redor da larva. Use apenas agarose suficiente para cobrir o peixe. Enquanto a agarose endurece, use fórceps finos para orientar o peixe em uma posição reta, lado ventral para baixo, no centro do prato.
- Adicione 2 mL da solução de gravação contendo brometo de pancurônio de 10 μM (ver Tabela de Materiais) para bloquear a transmissão neuromuscular. A adição do paralisador é necessária para eliminar artefatos devido a pequenos movimentos durante as gravações.
- Gravação eletrofisiológica
- Encha a micropipette com solução de gravação.
- Com o amplificador de grampo de remendo (ver Tabela de Materiais) na configuração do grampo de tensão, meça a resistência ao eletrodo no banho para confirmar seu valor correto.
- Usando um objetivo de 20x, posicione a cabeça da larva no campo de visão central e baixe a micropipette para alcançar a posição de gravação no cérebro, dentro do tectum óptico.
- Troque o amplificador do grampo de remendo para o grampo de corrente e fixe a corrente de retenção para 0 mA.
- Usando um filtro de baixa passagem de 1 kHz, uma taxa de aquisição de 1 kHz e um ganho digital de 10, registre atividade espontânea por 60 minutos para determinar os níveis de atividade na linha de base.
- Após 1 h de gravação de linha de base, adicione 200 μL pentiltetrazol (PTZ, ver Tabela de Materiais) solução 300 mmol/L ao banho para uma concentração final de 20 mmol/L PTZ.
- Registo a atividade neuronal em PTZ por mais 120 min.
- Determinação do evento de despolarização
- Os eventos de gravação de campo têm dinâmicas muito lentas (frequências de interesse estão na faixa de 0,005-0,2 s-1). Portanto, filtre o sinal com um passe baixo (Butterworth 5ª ordem LPF a 100 s-1) a fim de evitar o aliasing. Subsample os dados de tensão registrados da taxa de quadros de aquisição (neste caso, 1 ks-1) até 250 s-1 (RAW SIGNAL).
- Para identificar os carimbos de tempo para cada evento de despolarização, use um DETECTION SIGNAL, que é uma versão filtrada de alto passe do sinal gravado (Butterworth 1ª ordem HPF em 0,01 s-1).
- Eliminando os componentes de baixa frequência, a detecção de eventos de despolarização pode ser realizada usando um método de limiar simples. Utilize um limiar fixo para eliminação de ruído e detecção de eventos (0,3 mV foi utilizado para este estudo).
- Caracterize o evento de despolarização por uma série de cruzamentos de limiares que ocorrem em intervalos de tempo menores que 4 s. Calcule o início e o fim do evento de despolarização conforme determinado a partir de sequências consecutivas de cruzamentos limiares. Eventos com menos de 40 ms podem ser descartados como ruído.
- Calcule a amplitude dos eventos no RAW SIGNAL não filtrado para eliminar erros devido ao efeito da filtragem de passe baixo no pico do evento. Selecione a onda de despolarização do sinal bruto usando os carimbos de tempo determinados no sinal filtrado. Meça a amplitude como a diferença entre os valores máximo e mínimo da vaga selecionada a partir do sinal bruto.
NOTA: Os arquivos de script para executar a etapa 3.4 — determinação do evento de despolarização — e para obter a Figura 1 são fornecidos como Arquivo Suplementar anexado a este artigo.
Resultados
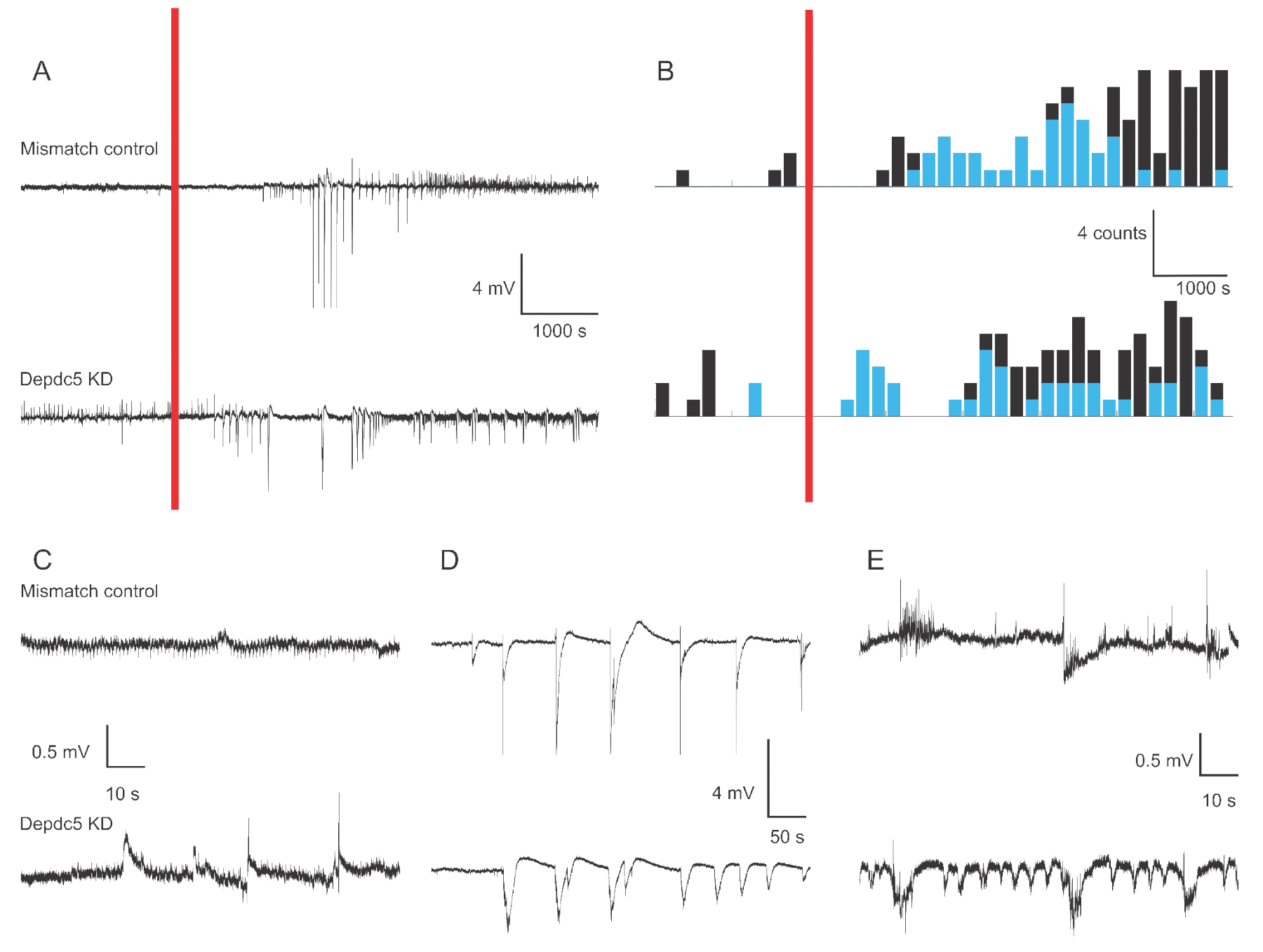
A Figura 1 mostra traços representativos de tensão de 4-6 dpf zebrafish larva extracelular no caso de duas condições genéticas: controle de incompatibilidade e knock-down DEPDC5. No período de base da gravação, o knock-down DEPDC5 mostra maior ocorrência de eventos espontâneos, enquanto o controle de incompatibilidade apresenta muito poucas flutuações. Esses padrões de atividade são representativos do aumento significativo da atividade neuronal devido à perda de função do DEPDC5,como já noticiamos anteriormente18. Após a aplicação ptz, tanto o controle de incompatibilidade quanto o knock-down DEPDC5 mostram um número crescente de eventos de despolarização. Durante o primeiro período após a aplicação de PTZ (10 – 60 min), observa-se uma taxa de 0,8 eventos por min tanto no controle de incompatibilidade quanto no knock-down de DEPDC5, onde a maioria dos eventos são de alta amplitude (>1 mV). Durante este último período de resposta (60 – 120 min após a aplicação do PTZ), a taxa de eventos de despolarização aumenta para cerca de 1 evento por minuto, e a maioria dos eventos são de baixa amplitude (≤1 mV).

Figura 1: Exemplos de gravações de campo no cérebro de larvas de zebrafish. (A) Visão geral da gravação de 180 min para uma larva de controle de incompatibilidade e um Knock-down DEPDC5. Primeiro, foi registrada atividade de base espontânea, depois o PTZ foi aplicado no banho (barra vermelha). (B) Histogramas de tempo de peri-estímulo dos eventos de despolarização para controle de incompatibilidade e knock-down DEPDC5. Os eventos foram classificados como alta amplitude (>1 mV - azul) e baixa amplitude (≤1 mV - preto). (C-E) Exemplos de traços dos diferentes períodos da gravação: (C) atividade espontânea, (D) Eventos de alta amplitude durante o primeiro período após a aplicação ptz, (E) Eventos de baixa amplitude durante este último período após a aplicação ptz. Observe que os arquivos de script para obter esses números são fornecidos como Arquivo Suplementar. Clique aqui para ver uma versão maior desta figura.
{kind=link}
Arquivo suplementar: Arquivos de script para a etapa 3.4. Clique aqui para ver uma versão maior desta figura.
Discussão
A epilepsia é uma doença neurológica complexa, com uma ampla gama de etiologias que começam a ser elucidadas com o advento das tecnologias de sequenciamento genético25,26,27. Modelos animais versáteis são essenciais para uma estratégia translacional eficiente que produzirá tanto insights sobre os mecanismos patológicos de epilepsias geneticamente ligadas, bem como terapias direcionadas para as formas distintas dessa condição. Os modelos de zebrafish têm sido muito eficazes na reprodução das principais características da epilepsia e no fornecimento de leituras confiáveis para o rastreamento de medicamentos antiepilépticos5,28. Convulsões espontâneas podem ser detectadas em zebrafish geneticamentemodificados 15,29,30,31 e análise neurofisiológica nesses modelos28 confirmou a base neuronal do comportamento epiléptico32,33. Larvas de zebrafish de pequeno porte são favoráveis a telas químicas em formato de 96 poços usando detecção automatizada de comportamentos simples, como a natação espontânea, que permite a detecção rápida de potenciais terapêuticos.
O modelo de knock-down DEPDC5 apresentado aqui é obtido pela injeção de AMO no embrião de zebrafish para bloquear a expressão genética durante o desenvolvimento. Este modelo apresenta várias características fenotípicas de pedra-chave durante diferentes pontos de tempo do desenvolvimento larval, que podem ser usados como indicadores de eficiência terapêutica durante um protocolo de triagem química ou genética. O knock-down genético mediado pela AMO é uma técnica poderosa, exibindo vantagens sobre modelos de convulsão induzidos quimicamente, pois visa especificamente a expressão de um gene de interesse, permitindo assim a identificação dos mecanismos patogênicos subjacentes desencadeados por uma mutação genética. Os indutores químicos, que são, no entanto, ferramentas potentes para rastreamento de drogas, podem atuar através de múltiplas vias celulares que podem nem sempre ser relevantes para a mutação genética em estudo. Embora a injeção AMO seja em si uma técnica simples quando dominada pelo experimentador, ela também apresenta uma série de limitações. As injeções devem ser realizadas no embrião de um estágio celular; em nossas mãos, as injeções em estágios posteriores aumentaram muito a variabilidade do fenótipo. Isso limita o tempo disponível para injeção; portanto, uma estratégia de geração de ovos para injeção em uma sequência de tempo é útil. Usamos rotineiramente 4-5 cruzes que abrimos em intervalos de 15-20 minutos, permitindo a injeção de uma embreagem antes de obter a próxima. Além disso, deve-se tomar cuidado para avaliar o fenótipo ao mesmo tempo entre diferentes experimentos, à medida que comportamentos estereotipados evoluem rapidamente durante os primeiros dias de desenvolvimento. O volume e a concentração de AMOs também devem ser cuidadosamente controlados, pois a toxicidade geral devido à injeção de quantidades excessivas mascarará o fenótipo específico. Os diferentes controles apresentados na introdução são essenciais para determinar a dose de injeção certa e o fenótipo correspondente.
As gravações de campo do cérebro de zebrafish larval são uma ferramenta útil para investigar os efeitos deletérios das mutações genéticas envolvidas em diferentes distúrbios cerebrais na atividade neuronal global34. Eventos de despolarização vistos nessas condições experimentais são um método estabelecido para avaliar efeitos eletrofisiológicos de drogas em diferentes condições epilépticas15,35. No entanto, a avaliação desses efeitos tem sido feita em sua maioria qualitativamente e não quantitativamente, e tendo um observador subjetivo como ator na análise. Aqui, desenvolvemos uma estratégia de detecção automática que pode quantificar objetivamente a taxa de despolarizações, sua amplitude e duração, e pode avaliar o progresso desses parâmetros ao longo do tempo, ou com diferentes intervenções genéticas ou farmacológicas.
Os resultados representativos aqui apresentados mostram a atividade de campo esperada do modelo genético de knock-down DEPDC5 em comparação com um controle de incompatibilidade em 4-6 dpf zebrafish, antes e depois da aplicação do PTZ para introduzir atividade eletrográfica semelhante à epipeso. Anteriormente, mostramos um aumento significativo na atividade basal da condição de knockdown DEPDC5 18. Aqui, mostramos que a resposta dessas duas condições ao PTZ, um indutor de atividade epileptiforme química, tem trajetória semelhante no tempo, começando com um período de frequência relativamente baixa, eventos de despolarização de alta amplitude e continuando com um período de maior frequência, eventos de despolarização de menor amplitude. Os eventos de gravação de campo têm dinâmica lenta (as frequências de interesse estão na faixa de 0,005-0,2 s-1), portanto, tanto filtros de baixo e alto passe são usados neste protocolo para isolar os eventos de interesse. Após a eliminação do ruído de baixa frequência, a detecção de eventos de despolarização é realizada usando um limiar simples. Uma vez que as estatísticas do sinal são muito afetadas pela presença de eventos de despolarização, não poderíamos usar o desvio padrão do sinal total para determinar esse limiar. A variabilidade do valor do desvio padrão entre os conjuntos de dados foi maior do que os níveis de ruído de gravação observados. Portanto, após a inspeção visual dos traços, utilizou-se um valor fixo do limiar de 0,3 mV, a fim de evitar o viés induzido por diferentes níveis de atividade de despolarização.
O protocolo descrito fornece um método padronizado e simples para avaliar o comportamento motor e a atividade do campo neuronal, por meio do registro de tensão de corrente extracelular, juntamente com a detecção automática de eventos de despolarização no tectum óptico, para caracterizar fenótipos epitipos epilépticos em modelos de zebrafish.
Divulgações
Os autores não têm nada a revelar.
Agradecimentos
Gostaríamos de agradecer ao pessoal da plataforma de eletrofisiologia do ICM onde foram realizados os experimentos de neurofisiologia. Agradecemos também a Anca Marian pela ajuda técnica. SC foi apoiado pelo Trampolim Grant #21488. O EK foi apoiado pelo AFM Grant #18469 e pela ERC Consolidator Grant (ALS-Networks). O HC foi apoiado por prêmios de doutorado da Fondation pour la Recherche Médicale (PLP20141031462) e arsla. Para AD e RM, este trabalho foi apoiado por três bolsas da Autoridade Nacional Romena de Pesquisa Científica e Inovação, CNCS-UEFISCDI (números do projeto PN-III-P4-ID-PCE-2016-0010, PN-III-P2-2.1-PED-2016-0007, e COFUND-NEURON-NMDAR-PSY), uma subvenção do programa de pesquisa e inovação Horizon 2020 da União Europeia – acordo de subvenção nº 668863-SyBil-AA, e uma bolsa da Fundação Nacional de Ciência nSF-IOS-1656830 financiada pelo Governo dos EUA.
Materiais
| Name | Company | Catalog Number | Comments |
| Agarose | Sigma-Aldrich, France | A9539 | |
| Aquarium salt | Instant Ocean, Blacksburg, VA | SS15-10 | |
| Borosilicate glass with filament | Sutter Instruments | BF100-50-10 | OD: 1.5mm, ID: 0.5 mm |
| CaCl2 | Sigma-Aldrich, France | C1016 | |
| Depdc5-atg antisense morpholino | GeneTools, OR, USA | N/A | sequence 5’- TGCCTTCATGGTGACCGTCATTTTA -3’ |
| Depdc5-mis antisense morpholino | GeneTools, OR, USA | N/A | sequence 5’- TGCgTTgATcGTGACCcTgATTTTA -3’ |
| Depdc5-splice antisense morpholino | GeneTools, OR, USA | N/A | sequence 5’- ACATTCCTGTTTCACCATAGATGAT -3’ |
| Digitizer | Molecular Devices, CA, USA | Digidata 1550 | |
| Fast Green Dye | Sigma-Aldrich, France | F7258 | Stock solution of 0.2% |
| Glass-bottom petri dishes | Ibidi, Germany | 81218 | |
| Glucose | Sigma-Aldrich, France | 68270 | |
| Grasshopper 2 camera | FLIR, BC, Canada | GRAS-03K2M-C | formerly Point Grey Research |
| HEPES | Sigma-Aldrich, France | H3375 | |
| Human wild-type DEPDC5 cDNA | Dharmacon, France | NM_001242897.1 | Accession: BC144291 Clone ID 905 |
| ImageJ software | NIH, USA | N/A | |
| KCl | Sigma-Aldrich, France | P9333 | |
| Matlab software | MathWorks, MA, USA | N/A | |
| MgCl2 | Sigma-Aldrich, France | M2670 | |
| NaCl | Sigma-Aldrich, France | S7653 | |
| NaOH | Sigma-Aldrich, France | 71687 | |
| Pancuronium bromide | Alomone Labs | P-130 | Stock solution of 60 mM in water |
| Parafilm | Sigma-Aldrich, France | P7793 | |
| Patch clamp amplifier | Molecular Devices, CA, USA | MultiClamp 700B | Computer-controled patch clamp amplifier |
| pClamp10 acquisition software | Molecular Devices | N/A | |
| Pentylenetetrazol (PTZ) | Sigma-Aldrich, France | P6500 | Stock solution of 300 mM (dissolved in recording solution) |
| Pipette puller | Narishige, Japan | PC-10 | |
| Pneumatic PicoPump | WPI, France | PV 820 | |
| Sylgard 184 kit | Sigma-Aldrich Intl. | 761036 | |
| Transfer plastic pipettes | Sigma-Aldrich, France | Z350605 | |
| Zebralab | Viewpoint, France | N/A |
Referências
- Kabashi, E., Champagne, N., Brustein, E., Drapeau, P. In the swim of things: Recent insights to neurogenetic disorders from zebrafish. Trends in Genetics. 26 (8), 373-381 (2010).
- Baxendale, S., van Eeden, F., Wilkinson, R. The Power of Zebrafish in personalised medicine. Advances in Experimental Medicine and Biology. , (2017).
- Baraban, S. C., Taylor, M. R., Castro, P. A., Baier, H. Pentylenetetrazole induced changes in zebrafish behavior, neural activity and c-fos expression. Neuroscience. , (2005).
- Cunliffe, V. T. Building a zebrafish toolkit for investigating the pathobiology of epilepsy and identifying new treatments for epileptic seizures. Journal of Neuroscience Methods. , (2016).
- Griffin, A., Krasniak, C., Baraban, S. C. Advancing epilepsy treatment through personalized genetic zebrafish models. Progress in Brain Research. , (2016).
- Griffin, A., Hamling, K. R., Knupp, K., Hong, S. G., Lee, L. P., Baraban, S. C. Clemizole and modulators of serotonin signalling suppress seizures in Dravet syndrome. Brain. , (2017).
- Orellana-Paucar, A. M., et al. Insights from zebrafish and mouse models on the activity and safety of ar-turmerone as a potential drug candidate for the treatment of epilepsy. PLoS ONE. , (2013).
- Baxendale, S., et al. Identification of compounds with anti-convulsant properties in a zebrafish model of epileptic seizures. Disease Models & Mechanisms. , (2012).
- Bar-Peled, L., et al. A tumor suppressor complex with GAP activity for the Rag GTPases that signal amino acid sufficiency to mTORC1. Science. 340 (6136), 1100-1106 (2013).
- Ishida, S., et al. Mutations of DEPDC5 cause autosomal dominant focal epilepsies. Nature Genetics. , (2013).
- Dibbens, L. M., et al. Mutations in DEPDC5 cause familial focal epilepsy with variable foci. Nature Genetics. , (2013).
- Baulac, S., Weckhuysen, S. . DEPDC5-Related Epilepsy. GeneReviews®. , (1993).
- Picard, F., et al. DEPDC5 mutations in families presenting as autosomal dominant nocturnal frontal lobe epilepsy. Neurology. , (2014).
- Teng, Y., et al. Knockdown of zebrafish lgi1a results in abnormal development, brain defects and a seizure-like behavioral phenotype. Human Molecular Genetics. , (2010).
- Baraban, S. C., Dinday, M. T., Hortopan, G. A. Drug screening in Scn1a zebrafish mutant identifies clemizole as a potential Dravet syndrome treatment. Nature Communications. , (2013).
- Suls, A., et al. De novo loss-of-function mutations in CHD2 cause a fever-sensitive myoclonic epileptic encephalopathy sharing features with dravet syndrome. American Journal of Human Genetics. , (2013).
- Grone, B. P., et al. Epilepsy, behavioral abnormalities, and physiological comorbidities in syntaxin-binding protein 1 (STXBP1) mutant zebrafish. PLoS ONE. , (2016).
- de Calbiac, H., et al. DEPDC5 knockdown causes mTOR-dependent motor hyperactivity in zebrafish. Annals of Clinical and Translational Neurology. , (2018).
- Panchaud, N., Péli-Gulli, M. P., De Virgilio, C. Amino acid deprivation inhibits TORC1 through a GTPase-activating protein complex for the Rag family GTPase Gtr1. Science Signaling. , (2013).
- Lim, K. R. Q., Maruyama, R., Yokota, T. Eteplirsen in the treatment of Duchenne muscular dystrophy. Drug Design, Development and Therapy. , (2017).
- Kok, F. O., et al. Reverse genetic screening reveals poor correlation between morpholino-induced and mutant phenotypes in zebrafish. Developmental Cell. , (2015).
- Rossi, A., et al. Genetic compensation induced by deleterious mutations but not gene knockdowns. Nature. , (2015).
- Stainier, D. Y. R., et al. Guidelines for morpholino use in zebrafish. PLoS Genetics. , (2017).
- Kimmel, C. B., Ballard, W. W., Kimmel, S. R., Ullmann, B., Schilling, T. F. Stages of embryonic development of the zebrafish. Developmental dynamics an official public. 203 (3), 253-310 (1995).
- Møller, R. S., Dahl, H. A., Helbig, I. The contribution of next generation sequencing to epilepsy genetics. Expert Review of Molecular Diagnostics. , (2015).
- Allen, A. S., et al. De novo mutations in epileptic encephalopathies. Nature. , (2013).
- Dunn, P., et al. Next generation sequencing methods for diagnosis of epilepsy syndromes. Frontiers in Genetics. , (2018).
- Grone, B. P., Baraban, S. C. Animal models in epilepsy research: Legacies and new directions. Nature Neuroscience. , (2015).
- Zhang, Y., et al. Pharmacological characterization of an antisense knockdown zebrafish model of Dravet syndrome: Inhibition of epileptic seizures by the serotonin agonist fenfluramine. PLoS ONE. , (2015).
- Swaminathan, A., et al. Non-canonical mTOR-Independent Role of DEPDC5 in Regulating GABAergic Network Development. Current Biology. , (2018).
- Samarut, &. #. 2. 0. 1. ;., et al. γ-Aminobutyric acid receptor alpha 1 subunit loss of function causes genetic generalized epilepsy by impairing inhibitory network neurodevelopment. Epilepsia. , 2061-2074 (2018).
- Turrini, L., et al. Optical mapping of neuronal activity during seizures in zebrafish. Scientific Reports. , (2017).
- Rosch, R. E., Hunter, P. R., Baldeweg, T., Friston, K. J., Meyer, M. P. Calcium imaging and dynamic causal modelling reveal brain-wide changes in effective connectivity and synaptic dynamics during epileptic seizures. PLoS Computational Biology. , (2018).
- Baraban, S. C. Forebrain Electrophysiological Recording in Larval Zebrafish. Journal of Visualized Experiments. , (2013).
- Afrikanova, T., et al. Validation of the Zebrafish Pentylenetetrazol Seizure Model: Locomotor versus Electrographic Responses to Antiepileptic Drugs. PLoS ONE. , (2013).
Reimpressões e Permissões
Solicitar permissão para reutilizar o texto ou figuras deste artigo JoVE
Solicitar PermissãoExplore Mais Artigos
This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. Todos os direitos reservados