
Method Article
Analisi comportamentale e fisiologica in un modello di zebrafish di epilessia
In questo articolo
Riepilogo
Qui presentiamo un protocollo per lo sviluppo e la caratterizzazione di un modello di epilessia zebrafish derivante dall'inibizione transitoria del gene DEPDC5.
Abstract
L'epilessia rappresenta uno dei disturbi neurologici più comuni, che colpisce circa 50 milioni di persone in tutto il mondo. I recenti progressi nella ricerca genetica hanno scoperto un ampio spettro di geni implicati in varie forme di epilessia, evidenziando la natura eterogenea di questo disturbo. Modelli animali appropriati sono essenziali per studiare i meccanismi patologici innescati da mutazioni genetiche implicate nell'epilessia e per sviluppare terapie specializzate e mirate. Negli ultimi anni, il pesce zebra è emerso come un prezioso organismo vertebrato per modellare le epilessie, con l'uso sia della manipolazione genetica che dell'esposizione a farmaci epiletrogeni noti, come il pentilenetetrazolo (PTZ), per identificare nuove terapie antiepilettiche. Mutazioni deleterio nel regolatore mTOR DEPDC5 sono state associate a varie forme di epilessia focale e l'abbattimento dell'ortologo zebrafish causa iperattività associata a episodi spontanei simili a convulsioni, nonché una maggiore attività elettrografica e il caratteristico nuoto della ruota girevole. Qui, abbiamo descritto il metodo coinvolto nella generazione del modello di perdita di funzione DEPDC5 e illustrato il protocollo per la valutazione dell'attività motoria a 28 e 48 ore dopo la fecondazione (hpf), nonché un metodo per registrare l'attività sul campo nel tectum ottico del pesce zebra. Viene inoltre fornita un'illustrazione dell'effetto del farmaco epileptogenico PTZ sull'attività neuronale nel tempo.
Introduzione
Grazie alle sue piccole dimensioni, allo sviluppo oviparo e alla trasparenza nelle prime fasi dello sviluppo, il pesce zebra è emerso come un prezioso organismo vertebrato per modellare malattie umane diverse come disturbi cardiovascolari, tumorali o neurologici1,2. Zebrafish combina i vantaggi di un vertebrato, tra cui l'elevata conservazione dell'architettura degli organi e del codice genetico, con le piccole dimensioni e la facilità di manipolazione genetica di organismi modello più semplici, facilitando così sia gli studi fondamentali che le applicazioni traslazionali. In particolare, la sua suscettibilità allo screening automatizzato ad alto rendimento del comportamento e dei marcatori fluorescenti dei processi cellulari ha reso il pesce zebra un modello particolarmente attraente per la ricerca sull'epilessia. Ciò è stato dimostrato da un elevato aumento nell'ultimo decennio del numero di pubblicazioni con modelli di epilessia indotti chimicamente e/o genetici3,4,5 e, più recentemente, segnalazioni di terapie promettenti ottenute da screening chimici in questi modelli6,7,8.
DEPDC5 è un membro del complesso GATOR1, un regolatore negativo della segnalazione mTOR9. Le mutazioni nel gene DEPDC5 sono state scoperte per la prima volta nel 2013 in probande affette da epilessie focali autosomiche dominanti10,11,e da allora sono state riportate in una serie di condizioni cliniche associate a manifestazioni epilettiche focali e displasia corticale focale12. Si prevede che la grande maggioranza delle mutazioni riportate causi la perdita di funzione del gene12e questo è stato formalmente dimostrato per un certo numero di trascritti mutati DEPDC5 che sono presi di mira da decadimento dell'mRNA mediato senza senso12,13. D'accordo, l'abbattimento dell'ortologo genico nel pesce zebra usando oligonucleotidi morfolino antisenso (AMO) si traduce in una serie di caratteristiche comuni ai modelli epilettici in questo organismo, tra cui iperattività, nuoto a ruota girare, convulsioni spontanee e attività neuronale potenziata14,15,16,17,18. È interessante notare che il trattamento con rapamicina, un inibitore della segnalazione mTOR, ha invertito le caratteristiche comportamentali di questo modello18, sostenendo l'ipotesi che la perdita di funzione DEPDC5 possa innescare l'epilessia a causa di una cattiva regolamentazione della via mTOR9,19.
L'abbattimento transitorio dell'espressione genica in vivo utilizzando oligonucleotidi antisenso portatori della modificazione del morfolino è stato uno strumento inestimabile per studiare il ruolo di geni specifici, alla pari con le tecniche basate su si/shRNA. Recentemente, le strategie basate su AMO hanno anche trovato applicazioni cliniche, con una prima terapia AMO che ha ricevuto l'approvazione della FDA per il trattamento dell'atrofia muscolare di Duchenne nel 201620. Mentre è stato riportato che nel pesce zebra il fenotipo del knock-down genico acuto basato sull'AMO non è sempre correlato con i modelli costitutivi knock-out21, ciò può essere dovuto almeno in alcuni casi a meccanismi compensatori generati da modificazioni genetiche costitutive22. Tuttavia, la questione della specificità del fenotipo indotto da AMO è una preoccupazione indiscutibile che deve essere affrontata diligentemente negli studi che utilizzano questa tecnologia23. Al fine di garantire la specificità del fenotipo knock-down basato su AMO, sono necessari diversi controlli chiave. Questi includono una curva dose-risposta che consente la selezione della dose più bassa di AMO efficace per l'abbattimento genico, evitando la tossicità complessiva dovuta all'introduzione di un eccesso di materiale genetico. L'uso di un AMO Mismatch che non prende di mira una particolare regione del genoma è necessario anche per stabilire una dose appropriata e per identificare un fenotipo specifico. Un secondo AMO che prende di mira una regione diversa dello stesso gene, come un AMO che blocca la giunzione, è necessario per confermare che il fenotipo è dovuto all'abbattimento del gene bersaglio. Il salvataggio del fenotipo knock-down con il cDNA del gene, sia l'ortologo umano che una versione modificata dal codone del gene zebrafish che non può essere presa di mira dall'AMO, fornisce un forte argomento a favore della specificità del fenotipo. La mancanza di salvataggio con lo stesso cDNA contenente mutazioni di perdita di funzione (come l'introduzione di un codone di arresto precoce) è un'ulteriore prova in questa direzione.
Qui, presentiamo un metodo per generare un modello di perdita di funzione DEPDC5 di zebrafish e il protocollo per la fenotipizzazione comportamentale a 28 e 48 ore dopo la fecondazione (hpf). A 28 hpf, la perdita di funzione di DEPDC5 causa iperattività complessiva, come evidenziato da movimenti di avvolgimento e contrazioni potenziati degli embrioni all'interno del corion. Un sistema automatizzato di rilevamento del movimento può essere utilizzato in questa fase per quantificare l'attività complessiva per embrione. A 48 hpf, i pesci zebra mostrano una fuga stereotipata che nuota in risposta al tocco. Nel pesce zebra con espressione downregulata di DEPDC5, la traiettoria di nuoto è significativamente più tortuosa che nei controlli, il pesce esibisce un modello simile a "vite di sughero" o "ruota girevole", simile ad altri modelli di epilessia riportati in questo organismo3,4. Le registrazioni elettrofisiologiche sono state ottenute nel tectum ottico nelle larve di zebrafish tra 4-6 giorni dopo la fecondazione (dpf) e mostrano un aumento al basale dell'attività neuronale negli animali abbattuti DEPDC5. Il vantaggio di questo modello è che presenta diverse caratteristiche fenotipiche in diversi punti temporali, che possono essere utili nel monitoraggio e nella valutazione dell'efficacia delle terapie farmacologiche durante lo sviluppo.
Protocollo
Le procedure sperimentali sono state approvate dai Comitati Etici Nazionali ed Istituzionali.
1. Knock-down transitorio del gene DEPDC5 nell'embrione di zebrafish
- Preparazione degli strumenti:
- Preparare piastre di Petri per iniezione rivestite di elastomero di silicio: mescolare la base e l'agente polimerizzante del kit (vedere Tabella dei materiali)in un rapporto 10: 1. Riempire una capsula di Petri da 35 mm a metà strada con la miscela. Attendere che il silicio si indurisca prima di usarlo (questo può richiedere diversi giorni).
- Preparare soluzioni stock da 1,2 mmol/L di oligonucleotidi morfolino antisenso (AMO; vedi Tabella dei materiali). Aggiungere 250 μL di acqua sterile a 300 nmole AMO liofilizzati per ottenere una soluzione stock da 1,2 mmol/L. Per una completa dissoluzione, riscaldare i flaconcini per 5 minuti a 65 °C. Vortice brevemente. Sigillare il tappo del tubo con un film plastico (vedi Tabella dei materiali).
- Per gli esperimenti di salvataggio di controllo, preparare un plasmide di espressione contenente il cDNA umano di DEPDC5 (vedi Tabella dei materiali)clonato nella spina dorsale pCS2 o un plasmide di espressione compatibile con zebrafish simile. Come controllo negativo, una mutazione che causa un codone di arresto precoce (p.Arg487*) è stata introdotta nel cDNA.
- Preparare l'acqua dell'embrione: 0,06 g/L di sale per acquari (vedi Tabella dei materiali)in acqua ad osmosi inversa + 0,5 mg/L di blu di metilene.
- Il giorno dell'iniezione, preparare aghi di vetro borosilicato per microiniezione utilizzando un estrattore (vedi Tabella dei materiali). Stabilire impostazioni di temperatura appropriate sull'estrattore dell'ago. Utilizzare un capillare di vetro borosilicato lungo 10 cm, 1/0,5 OD/ID mm per generare due capillari di ~ 5 cm con punte sottili con una lunghezza di circa 1 cm.
- Se la punta degli aghi è molto fine, impedendo l'espulsione della soluzione, rompere l'estremità della punta affusolata usando una pinna al microscopio.
- Poco prima dell'iniezione, preparare le soluzioni di lavoro degli AMO. Preparare sempre una soluzione fresca per garantire la riproducibilità dei risultati. Riscaldare i flaconcini di AMO a 65 °C per 5 minuti. Preparare un campione di iniezione da 5 μL contenente colorante Fast Green (concentrazione finale dello 0,02%, vedere Tabella dei materiali)e l'AMO diluito alla concentrazione di lavoro in acqua.
- Determinare empiricamente la concentrazione di lavoro dell'AMO per ciascun gene utilizzando una curva di risposta alla dose. La concentrazione di lavoro rappresenta una concentrazione in cui l'AMO è efficace nell'abbattere il gene senza causare tossicità generale, come gravi difetti morfologici. Tipicamente, le concentrazioni di lavoro AMO saranno in un intervallo da 0,2 mmol / L a 1 mmol / L (0,4 mmol / L è stato determinato come concentrazione efficace per questo studio18). Iniettare il morfolino mismatch di controllo alla stessa concentrazione dell'AMO efficace.
- Vortice i tubi e centrifuga brevemente per portare le goccioline sul fondo dei tubi.
- Per gli esperimenti di salvataggio, preparare un campione di iniezione da 5 μL con l'AMO diluito alla concentrazione di lavoro e il plasmide di espressione del cDNA diluito in acqua fino a una concentrazione finale da determinare empiricamente. Per l'espressione di DEPDC5 e del plasmide di controllo negativo, 100 ng/μL è risultato efficace per il salvataggio fenotipico.
- Preparazione dell'embrione:
- Il giorno prima della microiniezione, allestire le vasche di accoppiamento del pesce zebra. La mattina dell'iniezione, rimuovere i divisori per consentire la deposizione delle uova. Raccogliere le uova in piastre di Petri da 100 mm riempite con acqua embrionale usando un setaccio fine. Iniettare entro 20-30 minuti dalla raccolta, mentre le uova sono allo stadio di una cellula.
- Raccogli 60-80 uova con una pipetta pasteur di plastica e disponile nella capsula di Petri rivestita di silicio per l'iniezione. La superficie in silicio impedirà alle uova di scivolare durante le iniezioni. Rimuovere la maggior parte dell'acqua dell'embrione, lasciando quel tanto che basta per coprire le uova a metà strada.
- Microiniezioni:
- Riempire un ago di vetro con soluzione per iniezione. Posizionare l'ago verticalmente in uno dei tubi contenenti la soluzione iniettabile, assicurandosi che l'estremità inferiore dell'ago tocchi la soluzione. Attendere alcuni minuti fino a quando la soluzione per iniezione colorata aumenta per capillarità ed è visibile sulla punta dell'ago.
- Montare l'ago riempito sul manico di iniezione del microiniettore (vedere Tabella dei materiali).
- Accendere il compressore d'aria e regolare l'impostazione della pressione per generare un volume di iniezione di ~ 2 nL.
- Per calcolare il volume della soluzione iniettata, posizionare una goccia di olio minerale su un vetrino microtome. Iniettare la soluzione contenente colorante utilizzando i parametri di pressione e tempo impostati. Misurare il diametro della sfera fluida iniettata e calcolare il volume totale utilizzando la formula Volume=4/3*π*(d/2)3, dove d=il diametro misurato del bolo iniettato.
- Utilizzando un microscopio binoculare sezionante con un ingrandimento 4X, iniettare le uova allo stadio della singola cellula passando attraverso il corion e il tuorlo e proiettando la soluzione direttamente all'interno della cellula.
- Raccogliere gli embrioni iniettati in una capsula di Petri da 100 mm con acqua embrionale, etichettare il piatto e incubarli a 28 °C.
- Assicurarsi che la temperatura dell'incubatrice sia stabile nel tempo, poiché il tasso di sviluppo degli embrioni è termosensibile. Ad esempio, la crescita verrebbe accelerata a temperature più elevate e la fase di sviluppo è fondamentale per valutare correttamente il fenotipo24.
- Controllare la qualità delle uova 6-8 ore dopo l'iniezione e rimuovere gli embrioni morti e non fecondati utilizzando una pipetta pasteur di plastica.
- La mattina dopo, conta e rimuovi gli embrioni morti in ogni piatto con una pipetta di plastica Pasteur.
2. Analisi del comportamento
- Analisi dell'attività globale a 28 hpf:
- Condurre il test il pomeriggio del giorno dopo la microiniezione (28 hpf), assicurandosi che l'ora del giorno in cui viene condotto il test sia coerente rispetto agli esperimenti per eseguire analisi statistiche valide poiché lo sviluppo degli embrioni è molto rapido.
- Riempire un piatto da 35 mm (piatto di prova) con acqua embrionale e lasciarlo riscaldare nell'incubatrice (28 °C) per almeno 15 minuti prima di iniziare il test.
- Posizionare una griglia di rete di plastica (1,2x1,2 mm) tagliata a misura, sul fondo della parabola di prova.
- Avere un altro sperimentatore randomizzare l'ordine di test degli embrioni e mascherare i nomi delle condizioni da testare.
- Assicurarsi che il tasso di mortalità non cambi tra le condizioni e rispetto agli embrioni non iniettati per garantire la specificità del fenotipo. La percentuale di embrioni morti in tutte le condizioni non deve superare il 10-13%18.
- Posiziona 10-12 embrioni ancora all'interno del loro coro sulla rete di plastica usando una pipetta pasteur di plastica. Riempi il piatto di prova con abbastanza acqua embrionale per mantenere gli embrioni sommersi ma non galleggianti. Se necessario, spostare gli embrioni con cura utilizzando una punta di plastica per posizionarli sulla griglia.
- Utilizzando una videocamera (vedi Tabella dei materiali)collegata a un microscopio di dissezione, registrare l'attività di avvolgimento spontaneo per un periodo di tempo definito (i video lunghi 10-20 minuti sono solitamente sufficienti per ottenere campioni rappresentativi di esplosioni di attività per la quantificazione)
- Riportare gli embrioni nel rispettivo piatto e rimetterli nell'incubatrice. Ripeti l'esperimento con tutti gli embrioni necessari per ogni condizione (come determinato da un'analisi di potenza del 90%).
- Per analizzare il movimento spontaneo totale, utilizzate un sistema ZebraLab (vedi Tabella dei materiali). Utilizzando il modulo di quantificazione dell'attività, carica il video registrato e progetta le arene di tracciamento intorno a ciascun embrione come appropriato. Impostare la soglia di congelamento e burst rispettivamente su 10 e 50.
- Esegui l'analisi video automatizzata, che quantifica l'attività totale all'interno di ciascuna delle arene definite, quindi recupera il set di dati come foglio di calcolo ed esegui l'analisi utilizzando un software di analisi dei dati.
- Touch-Evoked-Escape-Response (TEER) a 48 hpf:
- Condurre il test al mattino due giorni dopo l'iniezione (48 ore dopo la fecondazione).
- Almeno 2 ore prima del test, decioionare gli embrioni usando una pinci forza fine. Assicurarsi che l'ora del giorno per la dechorionation e il test comportamentale sia coerente rispetto agli esperimenti.
- Riempire un piatto da 130 mm (piatto di prova) con acqua embrionale e lasciarlo riscaldare nell'incubatrice (28 °C) per almeno 15 minuti prima di iniziare il test.
- Contare e rimuovere le larve morte e morfologicamente deformate. Registrare i numeri per ogni condizione.
- Fai in modo che un altro sperimentatore randomizzi l'ordine e codifichi i nomi delle condizioni da testare.
- Montare la telecamera (vedere Tabella dei materiali)sulla parabola di prova assicurandosi che l'intera parabola di prova rientri nel campo visivo. Posizionare un righello all'interno del campo visivo fornisce una calibrazione interna per la distanza.
- Con una pipetta Pasteur di plastica, metti un embrione al centro della parabola di prova e inizia la registrazione utilizzando una velocità di acquisizione di 30 fps.
- Con una punta di plastica fine, tocca leggermente la coda dell'embrione con un movimento di scorrimento.
- Interrompere la registrazione quando la larva ha terminato il suo movimento.
- Rimuovere l'embrione dal piatto di prova e metterlo in un nuovo piatto pieno di acqua embrionale. Ripetere il test con tutti gli embrioni necessari per ogni condizione (come determinato da un'analisi di potenza del 90%).
- Riportare gli embrioni al loro piatto originale e rimetterli nell'incubatrice.
- Per analizzare i parametri del comportamento di nuoto, caricare il video registrato sul software di analisi ImageJ. Scarica e installa il plugin Manual Tracking di ImageJ (vedi Tabella dei Materiali). Avvia il plugin scegliendo Strumenti | | dei plugin Tracciamento manuale nel menu.
- Nella finestra di dialogo, introdurre la scala calibrata dell'immagine. L'inclusione di un righello nel campo della fotocamera facilita la conversione di cm in pixel.
- Seleziona Aggiungi traccia e inizia il tracciamento della traiettoria facendo clic sull'immagine della larva del pesce zebra nel primo fotogramma. I fotogrammi avanzano automaticamente con ogni punto aggiunto alla traccia.
- Continua a tracciare il movimento fino alla fine dell'episodio di nuoto.
- Selezionate Termina traccia nella finestra di tracciamento, recuperate le coordinate X-Y e calcolate la distanza totale, la velocità e l'angolo di sterzata.
3. Analisi elettrofisiologica
- Preparazione di reagenti e utensili:
- Preparare l'1% di acarosio in acqua embrionale (vedere paragrafo 1.1.4). Aliquotare l'agarosio liquido in tubi di microcentrifuga e tenerli su un blocco riscaldante a 42 °C per evitare che l'agarosi si indurisca.
- Preparare la soluzione di registrazione (in mmol/L): NaCl 134, KCl 2.9, CaCl2 2.1, MgCl2 1.2, glucosio 10, HEPES 10, pH 7.8.
- Tirare micropipette di vetro borosilicato con apertura della punta di 1,5-2 μm (5-6 Mm2·kg·s−3· A−2 resistenza) non lucidato.
- Preparazione di larve di zebrafish per elettrofisiologia:
- Posizionare il pesce in una capsula di Petri con fondo di vetro (vedi Tabella dei materiali)e rimuovere i mezzi extracellulari in eccesso per garantire che il pesce sia portato il più vicino possibile allo scivolo di copertura.
- Usando una pipetta pasteur di plastica, aggiungi agarose liquido caldo su e intorno alla larva. Usa abbastanza agarose per coprire il pesce. Mentre l'agarose si indurisce, usa una pinna fine per orientare il pesce in posizione diritta, lato ventrale verso il basso, al centro del piatto.
- Aggiungere 2 mL della soluzione di registrazione contenente 10 μM di bromuro di pancuronio (vedere Tabella dei materiali)per bloccare la trasmissione neuromuscolare. L'aggiunta del paralizzatore è necessaria per eliminare gli artefatti dovuti a piccoli movimenti durante le registrazioni.
- Registrazione elettrofisiologica
- Riempire la micropipetta con una soluzione di registrazione.
- Con l'amplificatore patch clamp (vedi Tabella dei materiali)in configurazione voltage clamp, misurare la resistenza dell'elettrodo nel bagno per confermarne il valore corretto.
- Usando un obiettivo 20x, posiziona la testa della larva nel campo visivo centrale e abbassa la micropipetta per raggiungere la posizione di registrazione nel cervello, all'interno del tectum ottico.
- Commutare l'amplificatore patch clamp sul morsetto di corrente e fissare la corrente di tenuta a 0 mA.
- Utilizzando un filtro passa-basso di 1 kHz, una velocità di acquisizione di 1 kHz e un guadagno digitale di 10, registrare l'attività spontanea per 60 minuti per determinare i livelli di attività di base.
- Dopo 1 ora di registrazione al basale, aggiungere 200 μL di pentilenetetrazolo (PTZ, vedi Tabella dei materiali)soluzione 300 mmol/L al bagno per una concentrazione finale di 20 mmol/L PTZ.
- Registrare l'attività neuronale in PTZ per altri 120 minuti.
- Determinazione dell'evento di depolarizzazione
- Gli eventi di field recording hanno dinamiche molto lente (le frequenze di interesse sono nell'intervallo 0,005-0,2 s-1). Pertanto, filtrare il segnale con un passaggio basso (Butterworth 5 ° ordine LPF a 100 s-1) al fine di evitare l'aliasing. Sottocampionare i dati di tensione registrati dal frame rate di acquisizione (in questo caso, 1 ks-1) fino a 250 s-1 (RAW SIGNAL).
- Per identificare i timestamp per ogni evento di depolarizzazione, utilizzare un SEGNALE DI RILEVAMENTO, che è una versione filtrata passa-alto del segnale registrato (HpF di 1° ordine di Butterworth a 0,01 s-1).
- Eliminando i componenti a bassa frequenza, è possibile eseguire il rilevamento di eventi di depolarizzazione utilizzando un semplice metodo di soglia. Utilizzare una soglia fissa per l'eliminazione del rumore e il rilevamento degli eventi (per questo studio sono stati utilizzati 0,3 mV).
- Caratterizzare l'evento di depolarizzazione da una serie di attraversamenti di soglie che si verificano a intervalli di tempo inferiori a 4 s. Calcola l'inizio e la fine dell'evento di depolarizzazione come determinato da sequenze consecutive di attraversamenti di soglia. Gli eventi di durata inferiore a 40 ms possono essere eliminati come rumore.
- Calcola l'ampiezza degli eventi nel segnale non filtrato (RAW SIGNAL) per eliminare gli errori dovuti all'effetto del filtro passa-basso sul picco dell'evento. Selezionare la wavelet di depolarizzazione dal segnale raw utilizzando i timestamp determinati nel segnale filtrato. Misurare l'ampiezza come differenza tra i valori massimi e minimi della wavelet selezionata dal segnale raw.
NOTA BENE: I file di script per eseguire il passaggio 3.4 — Determinazione dell'evento di depolarizzazione — e per ottenere la Figura 1 sono forniti come file supplementare allegato a questo articolo.
Risultati
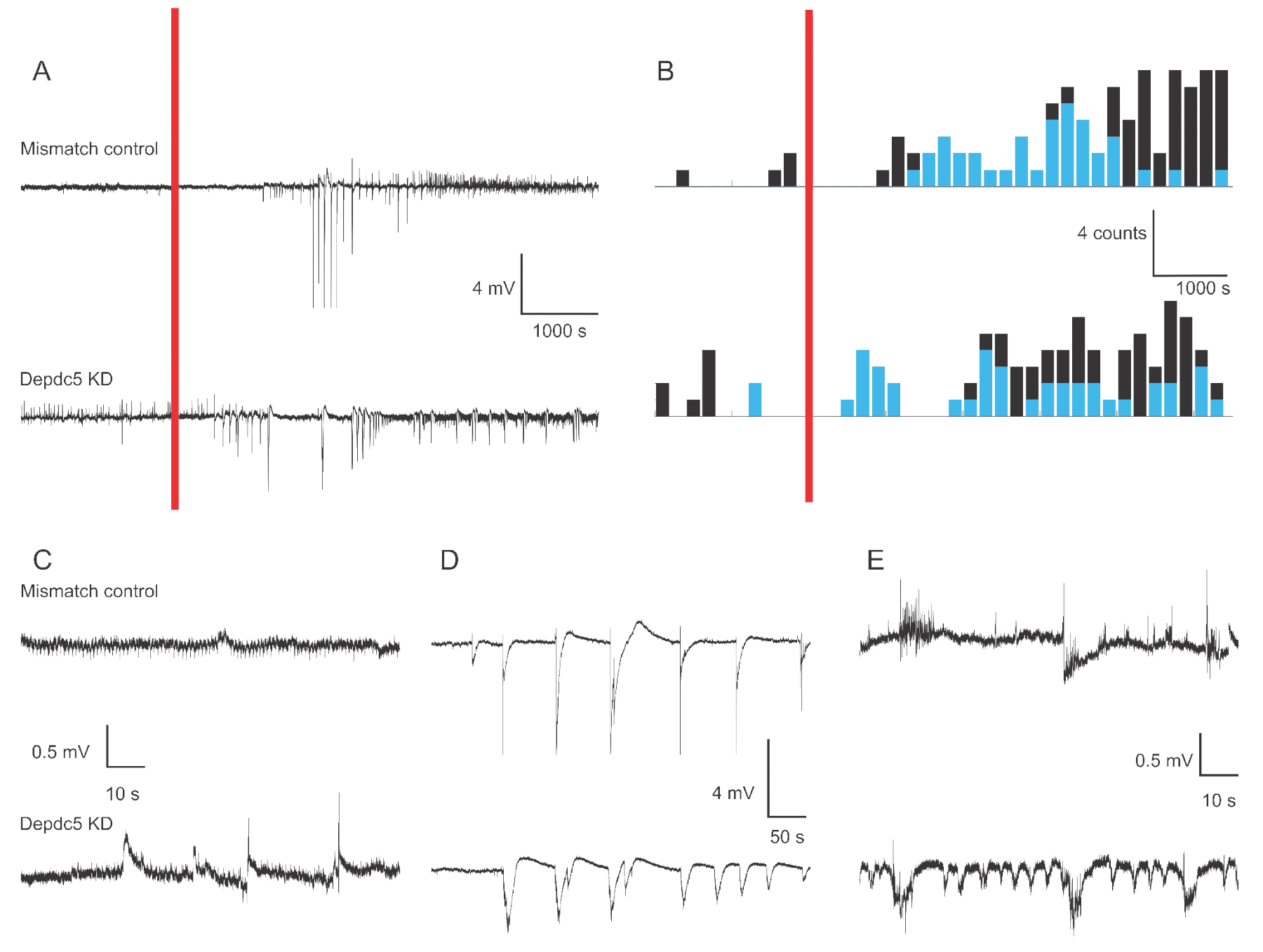
La Figura 1 mostra tracce di tensione rappresentative di registrazioni extracellulari del campo extracellulare della larva di zebrafish 4-6 dpf nel caso di due condizioni genetiche: controllo del mismatch e knock-down DEPDC5. Nel periodo di riferimento della registrazione, il knock-down DEPDC5 mostra una maggiore occorrenza di eventi spontanei, mentre il controllo Mismatch mostra pochissime fluttuazioni. Questi modelli di attività sono rappresentativi del significativo aumento dell'attività neuronale dovuto alla perdita di funzione di DEPDC5, come abbiamo precedentemente riportato18. Dopo l'applicazione PTZ, sia il controllo Mismatch che il knock-down DEPDC5 mostrano un numero maggiore di eventi di depolarizzazione. Durante il primo periodo dopo l'applicazione PTZ (10 – 60 min), si osserva una velocità di 0,8 eventi al minuto sia nel controllo Mismatch che nel knock-down DEPDC5, dove la maggior parte degli eventi sono di elevata ampiezza (>1 mV). Durante quest'ultimo periodo di risposta (60 – 120 minuti dopo l'applicazione PTZ), il tasso di eventi di depolarizzazione aumenta a circa 1 evento al minuto e la maggior parte degli eventi sono di bassa ampiezza (≤1 mV).

Figura 1: Tracce di esempio di registrazioni sul campo nel cervello delle larve di zebrafish. (A) Panoramica della registrazione di 180 minuti per una larva di controllo Mismatch e un DEPDC5 Knock-down. In primo luogo, è stata registrata l'attività basale spontanea, quindi la PTZ è stata applicata nel bagno (barra rossa). (B)Istogrammi temporali peristimoli degli eventi di depolarizzazione per il controllo del Mismatch e il knock-down DEPDC5. Gli eventi sono stati classificati come alta ampiezza (>1 mV - blu) e bassa ampiezza (≤1 mV - nero). (C-E) Esempi di tracce dei diversi periodi della registrazione: (C) attività spontanea, (D) Eventi di ampiezza elevata durante il primo periodo dopo l'applicazione PTZ, (E) Eventi di bassa ampiezza durante quest'ultimo periodo dopo l'applicazione PTZ. Si noti che i file di script per ottenere queste cifre sono forniti come file supplementare. Fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}
File supplementare: file di script per il passaggio 3.4. Fare clic qui per visualizzare una versione più grande di questa figura.
Discussione
L'epilessia è una malattia neurologica complessa, caratterizzata da una vasta gamma di eziologie che stanno iniziando a essere chiarite con l'avvento delle tecnologie di sequenziamento genetico25,26,27. Modelli animali versatili sono essenziali per una strategia traslazionale efficiente che produrrà sia approfondimenti sui meccanismi patologici delle epilessie geneticamente collegate, sia terapie mirate per le forme distinte di questa condizione. I modelli di Zebrafish sono stati molto efficaci nel riprodurre le principali caratteristiche dell'epilessia e nel fornire leggi affidabili per lo screening farmacologico antiepilettico5,28. Le convulsioni spontanee possono essere rilevate nel pesce zebra geneticamente modificato15,29,30,31 e l'analisi neurofisiologica in questi modelli28 ha confermato la base neuronale del comportamento epilettico-simile32,33. Le larve di zebrafish di piccole dimensioni sono suscettibili di schermi chimici in formato 96 pozzettati utilizzando il rilevamento automatico di comportamenti semplici, come il nuoto spontaneo, che consente un rapido rilevamento di potenziali terapie.
Il modello di knock-down DEPDC5 qui presentato è ottenuto mediante iniezione di AMO nell'embrione di zebrafish per bloccare l'espressione genica durante lo sviluppo. Questo modello presenta diverse caratteristiche fenotipiche chiave di volta durante diversi punti temporali dello sviluppo larvale, che possono essere utilizzate come indicatori dell'efficienza della terapia durante un protocollo di screening chimico o genetico. Il knock-down del gene mediato dall'AMO è una tecnica potente, che mostra vantaggi rispetto ai modelli di convulsioni indotte chimicamente, in quanto si rivolge specificamente all'espressione di un gene di interesse, consentendo così l'identificazione dei meccanismi patogenetici sottostanti innescati da una mutazione genetica. Gli induttori chimici, che sono tuttavia potenti strumenti per gli screening farmacologici, possono agire attraverso molteplici percorsi cellulari che potrebbero non essere sempre rilevanti per la mutazione genetica in studio. Mentre l'iniezione di AMO è di per sé una tecnica semplice quando padroneggiata dallo sperimentatore, presenta anche una serie di limitazioni. Le iniezioni devono essere eseguite all'embrione a una cellula; nelle nostre mani, le iniezioni nelle fasi successive hanno aumentato notevolmente la variabilità del fenotipo. Questo limita il tempo disponibile per l'iniezione; pertanto, è utile una strategia di generazione di uova per iniezione in una sequenza temporale. Usiamo abitualmente 4-5 croci che apriamo a intervalli di 15-20 minuti, consentendo l'iniezione di una frizione prima di ottenere quella successiva. Inoltre, bisogna fare attenzione a valutare il fenotipo allo stesso tempo tra diversi esperimenti, poiché i comportamenti stereotipati si evolvono rapidamente durante i primi giorni di sviluppo. Anche il volume e la concentrazione di AMO devono essere attentamente controllati, poiché la tossicità generale dovuta all'iniezione di quantità eccessive maschererà il fenotipo specifico. I diversi controlli presentati nell'introduzione sono essenziali per determinare la giusta dose di iniezione e il fenotipo corrispondente.
Le registrazioni sul campo del cervello larvale del pesce zebra sono uno strumento utile per indagare gli effetti deleteri delle mutazioni genetiche coinvolte in diversi disturbi cerebrali sull'attività neuronale globale34. Gli eventi di depolarizzazione osservati in queste condizioni sperimentali sono un metodo consolidato per valutare gli effetti elettrofisiologici dei farmaci in diverse condizioni epilettiche15,35. Tuttavia, la valutazione di questi effetti è stata per lo più fatta qualitativamente piuttosto che quantitativamente, e avendo un osservatore soggettivo come attore nell'analisi. Qui, sviluppiamo una strategia di rilevamento automatico in grado di quantificare oggettivamente il tasso di depolarizzazioni, la loro ampiezza e durata, e può valutare l'avanzamento di questi parametri nel tempo, o con diversi interventi genetici o farmacologici.
I risultati rappresentativi qui presentati mostrano l'attività sul campo prevista del modello genetico di knock-down DEPDC5 rispetto a un controllo Mismatch in 4-6 dpf zebrafish, prima e dopo l'applicazione di PTZ per introdurre attività elettrografica epileptiforme. In precedenza, abbiamo mostrato un aumento significativo dell'attività basale della condizione di knockdown DEPDC5 18. Qui, mostriamo che la risposta di queste due condizioni al PTZ, un induttore di attività epilettiforme chimica, ha una traiettoria simile nel tempo, iniziando con un periodo di eventi di depolarizzazione a frequenza relativamente bassa, alta ampiezza e continuando con un periodo di eventi di depolarizzazione a frequenza più alta, a bassa ampiezza. Gli eventi di registrazione sul campo hanno dinamiche lente (le frequenze di interesse sono nell'intervallo 0,005-0,2 s-1), quindi sia i filtri passa-basso che quelli passa-alto sono utilizzati in questo protocollo per isolare gli eventi di interesse. Dopo aver eliminato il rumore a bassa frequenza, il rilevamento degli eventi di depolarizzazione viene eseguito utilizzando una semplice soglia. Poiché le statistiche del segnale sono fortemente influenzate dalla presenza di eventi di depolarizzazione, non è stato possibile utilizzare la deviazione standard del segnale totale per determinare questa soglia. La variabilità del valore della deviazione standard tra i set di dati era maggiore dei livelli di rumore di registrazione osservati. Pertanto, dopo l'ispezione visiva delle tracce, abbiamo utilizzato un valore fisso della soglia di 0,3 mV, al fine di evitare la polarizzazione indotta da diversi livelli di attività di depolarizzazione.
Il protocollo descritto fornisce un metodo standardizzato e semplice per valutare il comportamento motorio e l'attività del campo neuronale, tramite la registrazione della tensione del morsetto di corrente extracellulare accoppiata con il rilevamento automatico di eventi di depolarizzazione nel tectum ottico, per caratterizzare fenotipi epilettiformi simili in modelli zebrafish.
Divulgazioni
Gli autori non hanno nulla da rivelare.
Riconoscimenti
Ringraziamo lo staff della piattaforma di elettrofisiologia ICM dove sono stati eseguiti gli esperimenti di neurofisiologia. Ringraziamo anche Anca Marian per l'aiuto tecnico. SC è stato supportato dal Trampoline Grant #21488. EK è stato supportato dall'AFM Grant #18469 e dall'ERC Consolidator Grant (ALS-Networks). HC è stato sostenuto da premi di dottorato della Fondation pour la Recherche Médicale (PLP20141031462) e ARSLA. Per AD e RM, questo lavoro è stato supportato da tre sovvenzioni dell'Autorità nazionale rumena per la ricerca scientifica e l'innovazione, CNCS-UEFISCDI (numeri di progetto PN-III-P4-ID-PCE-2016-0010, PN-III-P2-2.1-PED-2016-0007 e COFUND-NEURON-NMDAR-PSY), una sovvenzione del programma di ricerca e innovazione Horizon 2020 dell'Unione europea - convenzione di sovvenzione n. 668863-SyBil-AA e una sovvenzione della National Science Foundation NSF-IOS-1656830 finanziata dal governo degli Stati Uniti.
Materiali
| Name | Company | Catalog Number | Comments |
| Agarose | Sigma-Aldrich, France | A9539 | |
| Aquarium salt | Instant Ocean, Blacksburg, VA | SS15-10 | |
| Borosilicate glass with filament | Sutter Instruments | BF100-50-10 | OD: 1.5mm, ID: 0.5 mm |
| CaCl2 | Sigma-Aldrich, France | C1016 | |
| Depdc5-atg antisense morpholino | GeneTools, OR, USA | N/A | sequence 5’- TGCCTTCATGGTGACCGTCATTTTA -3’ |
| Depdc5-mis antisense morpholino | GeneTools, OR, USA | N/A | sequence 5’- TGCgTTgATcGTGACCcTgATTTTA -3’ |
| Depdc5-splice antisense morpholino | GeneTools, OR, USA | N/A | sequence 5’- ACATTCCTGTTTCACCATAGATGAT -3’ |
| Digitizer | Molecular Devices, CA, USA | Digidata 1550 | |
| Fast Green Dye | Sigma-Aldrich, France | F7258 | Stock solution of 0.2% |
| Glass-bottom petri dishes | Ibidi, Germany | 81218 | |
| Glucose | Sigma-Aldrich, France | 68270 | |
| Grasshopper 2 camera | FLIR, BC, Canada | GRAS-03K2M-C | formerly Point Grey Research |
| HEPES | Sigma-Aldrich, France | H3375 | |
| Human wild-type DEPDC5 cDNA | Dharmacon, France | NM_001242897.1 | Accession: BC144291 Clone ID 905 |
| ImageJ software | NIH, USA | N/A | |
| KCl | Sigma-Aldrich, France | P9333 | |
| Matlab software | MathWorks, MA, USA | N/A | |
| MgCl2 | Sigma-Aldrich, France | M2670 | |
| NaCl | Sigma-Aldrich, France | S7653 | |
| NaOH | Sigma-Aldrich, France | 71687 | |
| Pancuronium bromide | Alomone Labs | P-130 | Stock solution of 60 mM in water |
| Parafilm | Sigma-Aldrich, France | P7793 | |
| Patch clamp amplifier | Molecular Devices, CA, USA | MultiClamp 700B | Computer-controled patch clamp amplifier |
| pClamp10 acquisition software | Molecular Devices | N/A | |
| Pentylenetetrazol (PTZ) | Sigma-Aldrich, France | P6500 | Stock solution of 300 mM (dissolved in recording solution) |
| Pipette puller | Narishige, Japan | PC-10 | |
| Pneumatic PicoPump | WPI, France | PV 820 | |
| Sylgard 184 kit | Sigma-Aldrich Intl. | 761036 | |
| Transfer plastic pipettes | Sigma-Aldrich, France | Z350605 | |
| Zebralab | Viewpoint, France | N/A |
Riferimenti
- Kabashi, E., Champagne, N., Brustein, E., Drapeau, P. In the swim of things: Recent insights to neurogenetic disorders from zebrafish. Trends in Genetics. 26 (8), 373-381 (2010).
- Baxendale, S., van Eeden, F., Wilkinson, R. The Power of Zebrafish in personalised medicine. Advances in Experimental Medicine and Biology. , (2017).
- Baraban, S. C., Taylor, M. R., Castro, P. A., Baier, H. Pentylenetetrazole induced changes in zebrafish behavior, neural activity and c-fos expression. Neuroscience. , (2005).
- Cunliffe, V. T. Building a zebrafish toolkit for investigating the pathobiology of epilepsy and identifying new treatments for epileptic seizures. Journal of Neuroscience Methods. , (2016).
- Griffin, A., Krasniak, C., Baraban, S. C. Advancing epilepsy treatment through personalized genetic zebrafish models. Progress in Brain Research. , (2016).
- Griffin, A., Hamling, K. R., Knupp, K., Hong, S. G., Lee, L. P., Baraban, S. C. Clemizole and modulators of serotonin signalling suppress seizures in Dravet syndrome. Brain. , (2017).
- Orellana-Paucar, A. M., et al. Insights from zebrafish and mouse models on the activity and safety of ar-turmerone as a potential drug candidate for the treatment of epilepsy. PLoS ONE. , (2013).
- Baxendale, S., et al. Identification of compounds with anti-convulsant properties in a zebrafish model of epileptic seizures. Disease Models & Mechanisms. , (2012).
- Bar-Peled, L., et al. A tumor suppressor complex with GAP activity for the Rag GTPases that signal amino acid sufficiency to mTORC1. Science. 340 (6136), 1100-1106 (2013).
- Ishida, S., et al. Mutations of DEPDC5 cause autosomal dominant focal epilepsies. Nature Genetics. , (2013).
- Dibbens, L. M., et al. Mutations in DEPDC5 cause familial focal epilepsy with variable foci. Nature Genetics. , (2013).
- Baulac, S., Weckhuysen, S. . DEPDC5-Related Epilepsy. GeneReviews®. , (1993).
- Picard, F., et al. DEPDC5 mutations in families presenting as autosomal dominant nocturnal frontal lobe epilepsy. Neurology. , (2014).
- Teng, Y., et al. Knockdown of zebrafish lgi1a results in abnormal development, brain defects and a seizure-like behavioral phenotype. Human Molecular Genetics. , (2010).
- Baraban, S. C., Dinday, M. T., Hortopan, G. A. Drug screening in Scn1a zebrafish mutant identifies clemizole as a potential Dravet syndrome treatment. Nature Communications. , (2013).
- Suls, A., et al. De novo loss-of-function mutations in CHD2 cause a fever-sensitive myoclonic epileptic encephalopathy sharing features with dravet syndrome. American Journal of Human Genetics. , (2013).
- Grone, B. P., et al. Epilepsy, behavioral abnormalities, and physiological comorbidities in syntaxin-binding protein 1 (STXBP1) mutant zebrafish. PLoS ONE. , (2016).
- de Calbiac, H., et al. DEPDC5 knockdown causes mTOR-dependent motor hyperactivity in zebrafish. Annals of Clinical and Translational Neurology. , (2018).
- Panchaud, N., Péli-Gulli, M. P., De Virgilio, C. Amino acid deprivation inhibits TORC1 through a GTPase-activating protein complex for the Rag family GTPase Gtr1. Science Signaling. , (2013).
- Lim, K. R. Q., Maruyama, R., Yokota, T. Eteplirsen in the treatment of Duchenne muscular dystrophy. Drug Design, Development and Therapy. , (2017).
- Kok, F. O., et al. Reverse genetic screening reveals poor correlation between morpholino-induced and mutant phenotypes in zebrafish. Developmental Cell. , (2015).
- Rossi, A., et al. Genetic compensation induced by deleterious mutations but not gene knockdowns. Nature. , (2015).
- Stainier, D. Y. R., et al. Guidelines for morpholino use in zebrafish. PLoS Genetics. , (2017).
- Kimmel, C. B., Ballard, W. W., Kimmel, S. R., Ullmann, B., Schilling, T. F. Stages of embryonic development of the zebrafish. Developmental dynamics an official public. 203 (3), 253-310 (1995).
- Møller, R. S., Dahl, H. A., Helbig, I. The contribution of next generation sequencing to epilepsy genetics. Expert Review of Molecular Diagnostics. , (2015).
- Allen, A. S., et al. De novo mutations in epileptic encephalopathies. Nature. , (2013).
- Dunn, P., et al. Next generation sequencing methods for diagnosis of epilepsy syndromes. Frontiers in Genetics. , (2018).
- Grone, B. P., Baraban, S. C. Animal models in epilepsy research: Legacies and new directions. Nature Neuroscience. , (2015).
- Zhang, Y., et al. Pharmacological characterization of an antisense knockdown zebrafish model of Dravet syndrome: Inhibition of epileptic seizures by the serotonin agonist fenfluramine. PLoS ONE. , (2015).
- Swaminathan, A., et al. Non-canonical mTOR-Independent Role of DEPDC5 in Regulating GABAergic Network Development. Current Biology. , (2018).
- Samarut, &. #. 2. 0. 1. ;., et al. γ-Aminobutyric acid receptor alpha 1 subunit loss of function causes genetic generalized epilepsy by impairing inhibitory network neurodevelopment. Epilepsia. , 2061-2074 (2018).
- Turrini, L., et al. Optical mapping of neuronal activity during seizures in zebrafish. Scientific Reports. , (2017).
- Rosch, R. E., Hunter, P. R., Baldeweg, T., Friston, K. J., Meyer, M. P. Calcium imaging and dynamic causal modelling reveal brain-wide changes in effective connectivity and synaptic dynamics during epileptic seizures. PLoS Computational Biology. , (2018).
- Baraban, S. C. Forebrain Electrophysiological Recording in Larval Zebrafish. Journal of Visualized Experiments. , (2013).
- Afrikanova, T., et al. Validation of the Zebrafish Pentylenetetrazol Seizure Model: Locomotor versus Electrographic Responses to Antiepileptic Drugs. PLoS ONE. , (2013).
Ristampe e Autorizzazioni
Richiedi autorizzazione per utilizzare il testo o le figure di questo articolo JoVE
Richiedi AutorizzazioneEsplora altri articoli
This article has been published
Video Coming Soon
Personale delle biblioteche
Copyright © 2025 MyJoVE Corporation. Tutti i diritti riservati