
Method Article
Analyse comportementale et physiologique dans un modèle d’épilepsie de poisson-zèbre
Dans cet article
Résumé
Nous présentons ici un protocole pour le développement et la caractérisation d’un modèle d’épilepsie du poisson-zèbre résultant de l’inhibition transitoire du gène DEPDC5.
Résumé
L’épilepsie représente l’un des troubles neurologiques les plus courants, touchant environ 50 millions de personnes dans le monde. Les progrès récents de la recherche génétique ont mis au jour un large spectre de gènes impliqués dans diverses formes d’épilepsie, soulignant la nature hétérogène de ce trouble. Des modèles animaux appropriés sont essentiels pour étudier les mécanismes pathologiques déclenchés par les mutations génétiques impliquées dans l’épilepsie et pour développer des thérapies spécialisées et ciblées. Au cours des dernières années, le poisson-zèbre est devenu un organisme vertébré précieux pour la modélisation des épilepsies, avec l’utilisation de manipulations génétiques et l’exposition à des médicaments épileptogènes connus, tels que le ptéylènetétrazole (PTZ), pour identifier de nouveaux traitements antiépileptiques. Des mutations délétères dans le régulateur mTOR DEPDC5 ont été associées à diverses formes d’épilepsies focales et l’élimination de l’orthologue du poisson-zèbre provoque une hyperactivité associée à des épisodes spontanés de type crise, ainsi qu’une activité électrographique accrue et une nage caractéristique sur roue de virage. Ici, nous avons décrit la méthode impliquée dans la génération du modèle de perte de fonction DEPDC5 et illustrons le protocole d’évaluation de l’activité motrice à 28 et 48 h après la fécondation (HPF), ainsi qu’une méthode d’enregistrement de l’activité sur le terrain dans le tectum optique du poisson-zèbre. Une illustration de l’effet du médicament épileptogène PTZ sur l’activité neuronale au fil du temps est également fournie.
Introduction
En raison de sa petite taille, de son développement ovipare et de sa transparence aux premiers stades de développement, le poisson-zèbre est devenu un organisme vertébré précieux pour modéliser des maladies humaines aussi diverses que les troubles cardiovasculaires, cancéreux ou neurologiques1,2. Le poisson-zèbre combine les avantages d’un vertébré, y compris la haute conservation de l’architecture des organes et du code génétique, avec la petite taille et la facilité de manipulation génétique d’organismes modèles plus simples, facilitant ainsi à la fois les études fondamentales et les applications translationnelles. En particulier, son aptitude au dépistage automatisé à haut débit du comportement et aux marqueurs fluorescents des processus cellulaires a fait du poisson-zèbre un modèle particulièrement attrayant pour la recherche sur l’épilepsie. Cela a été démontré par une forte augmentation au cours de la dernière décennie du nombre de publications présentant des modèles chimiques et /ou génétiques de l’épilepsie 3 ,4,5 et, plus récemment, des rapports de thérapies prometteuses obtenues à partir de é écrans chimiques dans ces modèles6,7,8.
DEPDC5 est membre du complexe GATOR1, un régulateur négatif de la signalisation mTOR9. Des mutations du gène DEPDC5 ont été découvertes pour la première fois en 2013 chez des probands souffrant d’épilepsies focales autosomiques dominantes10,11, et ont depuis été rapportées dans un certain nombre de conditions cliniques associées à des manifestations épileptiques focales et à une dysplasie corticale focale12. La grande majorité des mutations rapportées devraient entraîner la perte de fonction du gène12,ce qui a été formellement démontré pour un certain nombre de transcriptions mutées DEPDC5 qui sont ciblées par une désintégration de l’ARNm médiée par des absurdités12,13. En accord, l’élimination de l’orthologue du gène chez le poisson-zèbre à l’aide d’oligonucléotides morpholino antisens (AMO) entraîne un certain nombre de caractéristiques communes aux modèles épileptiques de cet organisme, notamment l’hyperactivité, la nage en forme de roue, les crises spontanées etl’activiténeuronale accrue14, 15,16,17,18. Fait intéressant, le traitement par la rapamycine, un inhibiteur de la signalisation mTOR, a inversé les caractéristiques comportementales de ce modèle18,soutenant l’hypothèse selon laquelle la perte de fonction DEPDC5 peut déclencher l’épilepsie en raison d’une mauvaise régulation de la voie mTOR9,19.
L’élimination transitoire de l’expression génique in vivo à l’aide d’oligonucléotides antisens porteurs de la modification morpholino a été un outil inestimable pour étudier le rôle de gènes spécifiques, au même titre que les techniques à base de si / shRNA. Récemment, les stratégies basées sur l’AMO ont également trouvé des applications cliniques, avec une première thérapie AMO recevant l’approbation de la FDA pour le traitement de l’atrophie musculaire de Duchenne en 201620. Bien qu’il ait été rapporté que chez le poisson-zèbre, le phénotype de l’élimination génique aiguë à base d’AMO n’est pas toujours en corrélation avec les modèles constitutifsd’élimination 21,cela peut être dû au moins dans certains cas à des mécanismes compensatoires engendrés par des modifications génétiques constitutives22. Cependant, la question de la spécificité du phénotype induit par l’AMO est une préoccupation incontestable qui doit être traitée avec diligence dans les études utilisant cette technologie23. Afin d’assurer la spécificité du phénotype knock-down basé sur AMO, plusieurs contrôles clés sont nécessaires. Ceux-ci incluent une courbe dose-réponse qui permet de sélectionner la dose la plus faible d’AMO efficace pour l’élimination des gènes, évitant ainsi la toxicité globale due à l’introduction d’un excès de matériel génétique. L’utilisation d’une AMO incompatible qui ne cible aucune région particulière du génome est également nécessaire pour établir une dose appropriée et identifier un phénotype spécifique. Une deuxième AMO qui cible une région différente du même gène, telle qu’une AMO bloquant l’épissure, est nécessaire pour confirmer que le phénotype est dû à l’élimination du gène cible. Le sauvetage du phénotype knock-down avec l’ADNc du gène, soit l’orthologue humain, soit une version modifiée par codon du gène du poisson-zèbre qui ne peut pas être ciblée par l’AMO, fournit un argument fort en faveur de la spécificité du phénotype. L’absence de secours avec le même ADNc contenant des mutations de perte de fonction (comme l’introduction d’un codons d’arrêt précoce) est une preuve supplémentaire dans cette direction.
Nous présentons ici une méthode pour générer un modèle de perte de fonction DEPDC5 du poisson zèbre et le protocole de phénotypage comportemental à 28 et 48 h après la fécondation (hpf). À 28 hpf, la perte de fonction de DEPDC5 provoque une hyperactivité globale, comme en témoigne l’amélioration des mouvements d’enroulement et de contraction des embryons dans le chorion. Un système automatisé de détection de mouvement peut être utilisé à ce stade pour quantifier l’activité globale par embryon. À 48 hpf, les poissons-zèbres présentent une évasion stéréotypée en réponse au toucher. Chez le poisson-zèbre avec une expression régulée à la baisse de DEPDC5, la trajectoire de nage est significativement plus tortueuse que chez les témoins, le poisson présentant un motif de type « tire-bouchon » ou « roue tournant », similaire à d’autres modèles d’épilepsie rapportés dans cet organisme3,4. Des enregistrements électrophysiologiques ont été obtenus dans le tectum optique chez les larves de poisson zèbre entre 4 et 6 jours après la fécondation (dpf) et montrent une augmentation initiale de l’activité neuronale chez les animaux de depDC5. L’avantage de ce modèle est qu’il présente plusieurs caractéristiques phénotypiques à différents moments temporels, ce qui peut être utile pour surveiller et évaluer l’efficacité des thérapies médicamenteuses au cours du développement.
Protocole
Les procédures expérimentales ont été approuvées par les comités d’éthique nationaux et institutionnels.
1. Knock-down transitoire du gène DEPDC5 dans l’embryon de poisson zèbre
- Préparation des outils :
- Préparer des boîtes de Petri d’injection revêtues d’élastomère de silicium: Mélanger la base et l’agent de durcissement du kit (voir tableau des matériaux)dans un rapport de 10: 1. Remplissez une boîte de Petri de 35 mm à mi-chemin avec le mélange. Attendez que le silicium durcisse avant de l’utiliser (cela peut prendre plusieurs jours).
- Préparer des solutions mères de 1,2 mmol/L d’oligonucléotides morpholino antisens (AMO; voir Tableau des matériaux). Ajouter 250 μL d’eau stérile à 300 NMole lyophilisés AMO pour obtenir une solution mère de 1,2 mmol/L. Pour une dissolution complète, chauffer les flacons pendant 5 min à 65 °C. Vortex brièvement. Scellez le bouchon du tube avec un film plastique (voir Tableau des matériaux).
- Pour les expériences de sauvetage de contrôle, préparez un plasmide d’expression contenant l’ADNc humain de DEPDC5 (voir Table des matériaux)cloné dans l’épine dorsale pCS2 ou un plasmide d’expression similaire compatible avec le poisson-zèbre. En tant que témoin négatif, une mutation provoquant un arrêt précoce du codon (p.Arg487*) a été introduite dans l’ADNc.
- Préparer l’eau embryonnaire : 0,06 g/L de sel d’aquarium (voir tableau des matériaux)dans de l’eau d’osmose inverse + 0,5 mg/L de bleu de méthylène.
- Le jour de l’injection, préparez des aiguilles en verre borosilicate de micro-injection à l’aide d’un arracheur (voir Tableau des matériaux). Établissez les réglages de température appropriés sur l’arracheur d’aiguilles. Utilisez un capillaire en verre borosilicate de 10 cm de long, 1/0,5 OD/ID mm pour générer deux capillaires d’environ 5 cm avec des pointes minces d’une longueur d’environ 1 cm.
- Si la pointe des aiguilles est très fine, empêchant l’éjection de la solution, cassez l’extrémité de la pointe effilée à l’aide d’une pince au microscope.
- Juste avant l’injection, préparez les solutions de travail des MUNITIONS. Préparez toujours une solution fraîche pour assurer la reproductibilité des résultats. Chauffer les flacons de stock AMO à 65 °C pendant 5 min. Préparer un échantillon d’injection de 5 μL contenant du colorant Fast Green (concentration finale de 0,02 %, voir tableau des matériaux)et l’AMO dilué à la concentration de travail dans l’eau.
- Déterminer empiriquement la concentration de travail de l’AMO pour chaque gène à l’aide d’une courbe dose-réponse. La concentration de travail représente une concentration où l’AMO est efficace pour faire tomber le gène sans provoquer de toxicité générale, telle que des défauts morphologiques bruts. En règle générale, les concentrations de travail de l’AMO se situeront dans une plage de 0,2 mmol/L à 1 mmol/L (0,4 mmol/L a été déterminé comme concentration efficace pour cette étude18). Injecter le morpholino de contrôle Mismatch à la même concentration que l’AMO efficace.
- Vortex les tubes et centrifuger brièvement pour amener les gouttelettes au fond des tubes.
- Pour les expériences de sauvetage, préparer un échantillon d’injection de 5 μL avec l’AMO dilué à la concentration de travail et le plasmide d’expression de l’ADNc dilué dans l’eau jusqu’à une concentration finale à déterminer empiriquement. Pour l’expression de DEPDC5 et du plasmide témoin négatif, 100 ng/μL ont été efficaces pour le sauvetage phénotypique.
- Préparation de l’embryon:
- La veille de la microinjection, installez les bassins d’accouplement du poisson-zèbre. Le matin de l’injection, retirez les séparateurs pour permettre le frai. Recueillir les œufs dans des boîtes de Petri de 100 mm remplies d’eau embryonnaire à l’aide d’un tamis fin. Injecter dans les 20-30 minutes de la collecte, tandis que les œufs sont au stade d’une cellule.
- Choisissez 60 à 80 œufs avec une pipette Pasteur en plastique et disposez-les dans la boîte de Petri recouverte de silicium pour injection. La surface en silicium empêchera les œufs de glisser pendant les injections. Retirez la majeure partie de l’eau embryonnaire, en laissant juste assez pour couvrir les œufs à mi-chemin.
- Microinjections:
- Remplissez une aiguille en verre avec une solution injectable. Placez l’aiguille verticalement dans l’un des tubes contenant la solution injectable, en vous assurant que l’extrémité inférieure de l’aiguille touche la solution. Attendez plusieurs minutes jusqu’à ce que la solution injectable colorée augmente par capillarité et soit visible à l’extrémité de l’aiguille.
- Montez l’aiguille remplie sur la poignée d’injection du micro-injecteur (voir Tableau des matériaux).
- Allumez le compresseur d’air et ajustez le réglage de la pression pour générer un volume d’injection d’environ 2 nL.
- Pour calculer le volume de la solution injectée, placez une goutte d’huile minérale sur une lame de microtome. Injecter la solution contenant du colorant en utilisant les paramètres de pression et de temps définis. Mesurer le diamètre de la sphère de fluide injecté et calculer le volume total en utilisant la formule Volume=4/3*π*(d/2)3, où d=le diamètre mesuré du bolus injecté.
- À l’aide d’un microscope binoculaire disséquant avec un grossissement 4X, injecter les œufs au stade de cellule unique en passant à travers le chorion et le jaune, et en projetant la solution directement dans la cellule.
- Recueillir les embryons injectés dans une boîte de Petri de 100 mm avec de l’eau embryonnaire, étiqueter la boîte et les incuber à 28 °C.
- Assurez-vous que la température de l’incubateur est stable dans le temps, car le taux de développement des embryons est thermosensible. Par exemple, la croissance serait accélérée à des températures plus élevées et le stade de développement est essentiel pour évaluer correctement le phénotype24.
- Vérifiez la qualité des ovules 6 à 8 h après l’injection et retirez les embryons morts et non fécondés à l’aide d’une pipette Pasteur en plastique.
- Le lendemain matin, comptez et retirez les embryons morts dans chaque plat avec une pipette Pasteur en plastique.
2. Analyse du comportement
- Analyse de l’activité globale à 28 hpf :
- Effectuez le test l’après-midi du lendemain de la microinjection (28 hpf), en vous assurant que l’heure de la journée à laquelle le test est effectué est cohérente par rapport aux expériences pour effectuer une analyse statistique valide car le développement des embryons est très rapide.
- Remplissez un plat de 35 mm (plat d’essai) avec de l’eau embryonnaire et laissez-le se réchauffer dans l’incubateur (28 °C) pendant au moins 15 min avant de commencer le test.
- Placez une grille en maille en plastique (1,2 x 1,2 mm) coupée à la taille, sur le fond du plat d’essai.
- Demandez à un autre expérimentateur de randomiser l’ordre des tests des embryons et de masquer les noms des conditions à tester.
- Assurez-vous que le taux de mortalité ne change pas entre les conditions et par rapport aux embryons non injectés pour assurer la spécificité du phénotype. Le pourcentage d’embryons morts dans toutes les conditions ne doit pas dépasser 10-13%18.
- Placez 10 à 12 embryons encore dans leur chorion sur la maille en plastique à l’aide d’une pipette Pasteur en plastique. Remplissez le plat d’essai avec suffisamment d’eau embryonnaire pour garder les embryons submergés mais pas flottants. Si nécessaire, déplacez les embryons avec soin à l’aide d’une pointe en plastique pour les positionner sur la grille.
- À l’aide d’une caméra vidéo (voir Tableau des matériaux)attachée à un microscope à dissection, enregistrez l’activité d’enroulement spontanée pendant une durée définie (des vidéos de 10 à 20 minutes suffisent généralement pour obtenir des échantillons représentatifs de sursauts d’activité pour la quantification)
- Remettez les embryons dans leur plat respectif et remettez-les dans l’incubateur. Répétez l’expérience avec autant d’embryons que nécessaire pour chaque condition (telle que déterminée par une analyse de puissance à 90%).
- Pour analyser le mouvement spontané total, utilisez un système ZebraLab (voir Tableau des matériaux). À l’aide du module de quantification de l’activité, téléchargez la vidéo enregistrée et concevez les arènes de suivi autour de chaque embryon, le cas échéant. Réglez le seuil de gel et d’éclatement sur 10 et 50, respectivement.
- Exécutez l’analyse vidéo automatisée, qui quantifie l’activité totale dans chacune des arènes définies, puis récupérez l’ensemble de données sous forme de feuille de calcul et effectuez l’analyse à l’aide d’un logiciel d’analyse de données.
- Touch-Evoked-Escape-Response (TEER) à 48 chf :
- Effectuer le test le matin deux jours après l’injection (48 h après la fécondation).
- Au moins 2 heures avant le test, déchorionate les embryons à l’aide de pinces fines. Assurez-vous que l’heure de la journée pour la déséchorionation et le test de comportement est cohérente par rapport aux expériences.
- Remplissez un plat de 130 mm (plat d’essai) avec de l’eau embryonnaire et laissez-le se réchauffer dans l’incubateur (28 °C) pendant au moins 15 minutes avant de commencer le test.
- Compter et enlever les larves mortes et morphologiquement déformées. Enregistrez les chiffres pour chaque condition.
- Demandez à un autre expérimentateur de randomiser l’ordre et de codifier les noms des conditions à tester.
- Montez la caméra (voir Tableau des matériaux)sur la parabole d’essai en vous assurant que l’ensemble de la parabole d’essai se trouve dans le champ de vision. Le placement d’une règle dans le champ de vision fournit un étalonnage interne de la distance.
- Avec une pipette Pasteur en plastique, placez un embryon au centre de la boîte d’essai et commencez l’enregistrement en utilisant un taux d’acquisition de 30 fps.
- Avec une fine pointe en plastique, touchez légèrement la queue de l’embryon avec un mouvement de scintillement.
- Arrêtez l’enregistrement lorsque la larve a terminé son mouvement.
- Retirez l’embryon du plat d’essai et placez-le dans un nouveau plat rempli d’eau embryonnaire. Répétez le test avec autant d’embryons que nécessaire pour chaque condition (tel que déterminé par une analyse de puissance à 90%).
- Remettez les embryons dans leur plat d’origine et remettez-les dans l’incubateur.
- Pour analyser les paramètres du comportement de nage, chargez la vidéo enregistrée dans le logiciel d’analyse ImageJ. Téléchargez et installez le plugin Manual Tracking d’ImageJ (voir Table of Materials). Lancez le plugin en choisissant Outils | Plugin | Suivi manuel dans le menu.
- Dans la fenêtre de dialogue, introduisez l’échelle calibrée de l’image. L’inclusion d’une règle dans le champ de la caméra facilite la conversion de cm en pixels.
- Sélectionnez Ajouter une piste et commencez le traçage de trajectoire en cliquant sur l’image de la larve de poisson zèbre dans la première image. Les images avancent automatiquement avec chaque point ajouté à la trace.
- Continuez à suivre le mouvement jusqu’à la fin de l’épisode de natation.
- Sélectionnez Fin de piste dans la fenêtre de suivi, récupérez les coordonnées X-Y et calculez la distance totale, la vitesse et l’angle de braquage.
3. Analyse électrophysiologique
- Préparation des réactifs et des outils :
- Préparer 1 % d’agarose dans de l’eau embryonnaire (voir rubrique 1.1.4). Aliquoter l’agarose liquide dans des tubes de microcentrifugation et les maintenir sur un bloc chauffant à 42 °C pour empêcher l’agarose de durcir.
- Préparer la solution d’enregistrement (en mmol/L) : NaCl 134, KCl 2,9, CaCl2 2,1, MgCl2 1,2, glucose 10, HEPES 10, pH 7,8.
- Tirez des micropipettes en verre borosilicate avec une ouverture de pointe de 1,5-2 μm (5-6 Mm2·kg·s−3· Unerésistance −2) non polie.
- Préparation des larves de poisson zèbre pour l’électrophysiologie:
- Placez le poisson dans une boîte de Petri à fond de verre (voir Tableau des matériaux)et retirez l’excès de milieu extracellulaire afin de s’assurer que le poisson est amené le plus près possible du couvercle.
- À l’aide d’une pipette Pasteur en plastique, ajoutez de l’agarose liquide chaud sur et autour de la larve. Utilisez juste assez d’agarose pour couvrir le poisson. Pendant que l’agarose durcit, utilisez une pince fine pour orienter le poisson en position droite, côté ventral vers le bas, au centre du plat.
- Ajouter 2 mL de la solution d’enregistrement contenant 10 μM de bromure de pancuronium (voir tableau des matériaux)pour bloquer la transmission neuromusculaire. L’ajout du paralyseur est nécessaire pour éliminer les artefacts dus à de petits mouvements pendant les enregistrements.
- Enregistrement électrophysiologique
- Remplissez la micropipette avec une solution d’enregistrement.
- Avec l’amplificateur à pince de patch (voir Tableau des matériaux)en configuration de pince de tension, mesurez la résistance de l’électrode dans le bain pour confirmer sa valeur correcte.
- À l’aide d’un objectif 20x, placez la tête de la larve dans le champ de vision central et abaissez la micropipette pour atteindre la position d’enregistrement dans le cerveau, dans le tectum optique.
- Basculez l’amplificateur de pince de patch sur la pince de courant et fixez le courant de maintien à 0 mA.
- À l’aide d’un filtre passe-bas de 1 kHz, d’un taux d’acquisition de 1 kHz et d’un gain numérique de 10, enregistrez l’activité spontanée pendant 60 minutes pour déterminer les niveaux d’activité de base.
- Après 1 h d’enregistrement de base, ajouter 200 μL de pentylènetétrazol (PTZ, voir Tableau des matériaux)solution 300 mmol/L au bain pour une concentration finale de 20 mmol/L PTZ.
- Enregistrez l’activité neuronale dans le PTZ pendant encore 120 minutes.
- Détermination de l’événement de dépolarisation
- Les événements d’enregistrement sur le terrain ont une dynamique très lente (les fréquences d’intérêt sont comprises entre 0,005 et 0,2 s-1). Par conséquent, filtrez le signal avec un passe-bas (Butterworth 5th order LPF à 100 s-1) afin d’éviter l’aliasing. Sous-échantillonnez les données de tension enregistrées à partir de la fréquence d’images d’acquisition (dans ce cas, 1 ks-1) jusqu’à 250 s-1 (SIGNAL RAW).
- Pour identifier les horodatages de chaque événement de dépolarisation, utilisez un SIGNAL DE DÉTECTION, qui est une version filtrée passe-haut du signal enregistré (HPF de Butterworth 1er ordre à 0,01 s-1).
- En éliminant les composants basse fréquence, la détection des événements de dépolarisation peut être effectuée à l’aide d’une méthode de seuillage simple. Utiliser un seuil fixe pour l’élimination du bruit et la détection des événements (0,3 mV a été utilisé pour cette étude).
- Caractériser l’événement de dépolarisation par une série de franchissements de seuils qui se produisent à des intervalles de temps inférieurs à 4 s. Calculez le début et la fin de l’événement de dépolarisation déterminés à partir de séquences consécutives de franchissement de seuil. Les événements de moins de 40 ms peuvent être rejetés comme du bruit.
- Calculez l’amplitude des événements dans le non filtré (SIGNAL RAW) pour éliminer les erreurs dues à l’effet du filtrage passe-bas sur le pic de l’événement. Sélectionnez l’ondelette de dépolarisation du signal brut à l’aide des horodatages déterminés dans le signal filtré. Mesurez l’amplitude comme la différence entre les valeurs maximale et minimale de l’ondelette sélectionnée dans le signal brut.
REMARQUE : Les fichiers de script pour effectuer l’étape 3.4 — Détermination de l’événement de dépolarisation — et pour obtenir la Figure 1 sont fournis en tant que fichier supplémentaire joint à cet article.
Résultats
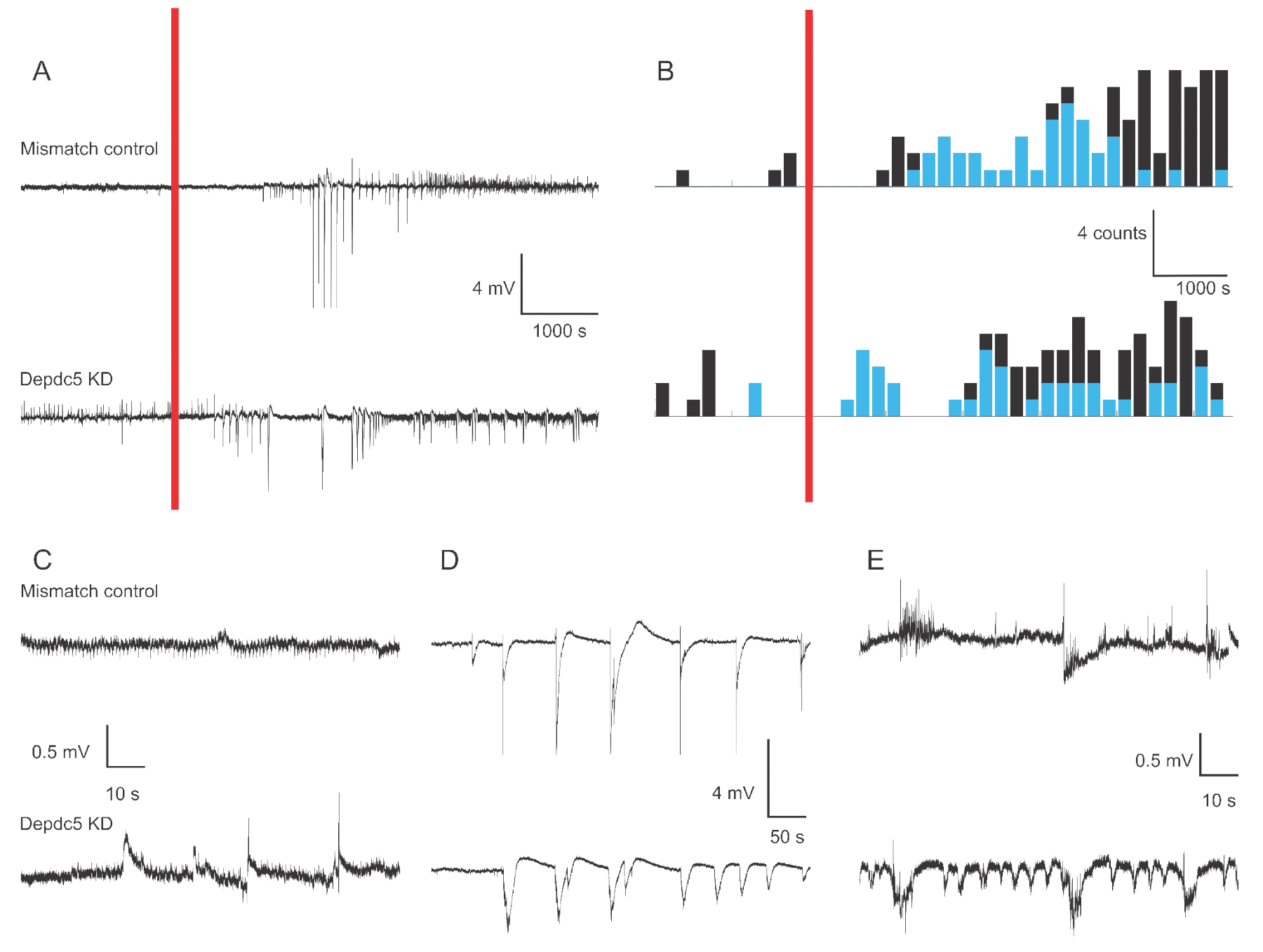
La figure 1 montre des traces de tension représentatives de 4 à 6 dpfs d’enregistrements de champ extracellulaire de larves de poisson zèbre dans le cas de deux conditions génétiques: le contrôle de l’inadéquation et l’élimination du DEPDC5. Au cours de la période de référence de l’enregistrement, le knock-down DEPDC5 montre une occurrence plus élevée d’événements spontanés, tandis que le contrôle Mismatch affiche très peu de fluctuations. Ces modèles d’activité sont représentatifs de l’augmentation significative de l’activité neuronale due à la perte de fonction de DEPDC5, comme nous l’avons déjà signalé18. Après l’application PTZ, le contrôle De l’incompatibilité et le knock-down DEPDC5 montrent un nombre accru d’événements de dépolarisation. Au cours de la première période suivant l’application de PTZ (10 à 60 min), un taux de 0,8 événement par minute est observé à la fois dans le contrôle de l’inadéquation et dans la chute de DEPDC5, où la majorité des événements sont de grande amplitude (>1 mV). Au cours de cette dernière période de réponse (60 à 120 minutes après l’application du PTZ), le taux d’événements de dépolarisation augmente à environ 1 événement par minute, et la majorité des événements sont de faible amplitude (≤1 mV).

Figure 1: Exemples de traces d’enregistrements sur le terrain dans le cerveau des larves de poisson zèbre. (A) Vue d’ensemble de 180 minutes d’enregistrement pour une larve de contrôle d’incompatibilité et un KNOCK-down DEPDC5. Tout d’abord, une activité de base spontanée a été enregistrée, puis le PTZ a été appliqué dans le bain (barre rouge). (B) Histogrammes de temps péri-stimulus des événements de dépolarisation pour le contrôle de l’inadéquation et l’élimination du DEPDC5. Les événements ont été classés comme étant de grande amplitude (>1 mV - bleu) et de faible amplitude (≤1 mV - noir). (C-E) Exemples de traces des différentes périodes de l’enregistrement: (C) activité spontanée, (D) événements de haute amplitude au cours de la première période après l’application de PTZ, (E) événements de faible amplitude au cours de la dernière période après application de PTZ. Notez que les fichiers de script pour obtenir ces chiffres sont fournis en tant que fichier supplémentaire. Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}
Fichier supplémentaire : Fichiers de script pour l’étape 3.4. Veuillez cliquer ici pour voir une version agrandie de cette figure.
Discussion
L’épilepsie est une maladie neurologique complexe, présentant un large éventail d’étiologies qui commencent à être élucidées avec l’avènement des technologies de séquençage génétique25,26,27. Des modèles animaux polyvalents sont essentiels pour une stratégie translationnelle efficace qui permettra de mieux comprendre les mécanismes pathologiques des épilepsies génétiquement liées, ainsi que des thérapies ciblées pour les formes distinctes de cette maladie. Les modèles de poisson-zèbre ont été très efficaces pour reproduire les principales caractéristiques de l’épilepsie et fournir des lectures fiables pour le dépistage des médicaments antiépileptiques5,28. Des crises spontanées peuvent être détectées chez les poissons-zèbres génétiquement modifiés15,29,30,31 et l’analyse neurophysiologique dans ces modèles28 a confirmé la base neuronale du comportement épileptique32,33. Les larves de poisson zèbre de petite taille se prêtent à des écrans chimiques au format 96 puits en utilisant la détection automatisée de comportements simples, tels que la nage spontanée, ce qui permet une détection rapide des traitements potentiels.
Le modèle de démolition DEPDC5 présenté ici est obtenu par injection d’AMO dans l’embryon de poisson zèbre pour bloquer l’expression des gènes pendant le développement. Ce modèle présente plusieurs caractéristiques phénotypiques clés de voûte à différents moments du développement larvaire, qui peuvent être utilisées comme indicateurs de l’efficacité du traitement au cours d’un protocole de dépistage chimique ou génétique. L’élimination du gène médiée par l’AMO est une technique puissante, présentant des avantages par rapport aux modèles de crises induites chimiquement, car elle cible spécifiquement l’expression d’un gène d’intérêt, permettant ainsi l’identification des mécanismes pathogènes sous-jacents déclenchés par une mutation génétique. Les inducteurs chimiques, qui sont néanmoins des outils puissants pour le dépistage des médicaments, peuvent agir par de multiples voies cellulaires qui pourraient ne pas toujours être pertinentes pour la mutation génétique à l’étude. Bien que l’injection d’AMO soit en soi une technique simple lorsqu’elle est maîtrisée par l’expérimentateur, elle présente également un certain nombre de limites. Les injections doivent être effectuées au stade cellulaire de l’embryon; dans nos mains, les injections à des stades ultérieurs ont considérablement augmenté la variabilité du phénotype. Cela limite le temps disponible pour l’injection; par conséquent, une stratégie de génération d’ovules à injecter dans une séquence temporelle est utile. Nous utilisons régulièrement 4-5 croix que nous ouvrons à des intervalles de 15-20 minutes, permettant l’injection d’un embrayage avant d’obtenir le suivant. En outre, il faut prendre soin d’évaluer le phénotype en même temps entre les différentes expériences, car les comportements stéréotypés évoluent rapidement au cours des premiers jours de développement. Le volume et la concentration des MUNITIONS doivent également être soigneusement contrôlés, car la toxicité générale due à l’injection de quantités excessives masquera le phénotype spécifique. Les différents contrôles présentés dans l’introduction sont essentiels pour déterminer la bonne dose d’injection et le phénotype correspondant.
Les enregistrements sur le terrain du cerveau larvaire du poisson-zèbre sont un outil utile pour étudier les effets délétères des mutations génétiques impliquées dans différents troubles cérébraux sur l’activité neuronale globale34. Les événements de dépolarisation observés dans ces conditions expérimentales sont une méthode établie pour évaluer les effets électrophysiologiques des médicaments dans différentes conditions épileptiques15,35. Cependant, l’évaluation de ces effets a surtout été faite qualitativement plutôt que quantitativement, et avec un observateur subjectif comme acteur dans l’analyse. Ici, nous développons une stratégie de détection automatique qui peut quantifier objectivement le taux de dépolarisations, leur amplitude et leur durée, et peut évaluer la progression de ces paramètres dans le temps, ou avec différentes interventions génétiques ou pharmacologiques.
Les résultats représentatifs présentés ici montrent l’activité de terrain attendue du modèle génétique de knock-down DEPDC5 par rapport à un contrôle d’incompatibilité chez le poisson zèbre 4-6 dpf, avant et après l’application de PTZ pour introduire une activité électrographique de type épileptiforme. Auparavant, nous avons montré une augmentation significative de l’activité basale de la condition de knockdown DEPDC5 18. Ici, nous montrons que la réponse de ces deux conditions au PTZ, un inducteur d’activité épileptiforme chimique, a une trajectoire similaire dans le temps, commençant par une période d’événements de dépolarisation de fréquence relativement basse, d’amplitude élevée et se poursuivant par une période d’événements de dépolarisation de fréquence plus élevée et d’amplitude plus faible. Les événements d’enregistrement sur le terrain ont une dynamique lente (les fréquences d’intérêt sont comprises entre 0,005 et 0,2 s-1),c’est pourquoi des filtres passe-bas et passe-haut sont utilisés dans ce protocole pour isoler les événements d’intérêt. Après avoir éliminé le bruit de basse fréquence, la détection des événements de dépolarisation est effectuée à l’aide d’un simple seuil. Étant donné que les statistiques du signal sont grandement affectées par la présence d’événements de dépolarisation, nous n’avons pas pu utiliser l’écart-type du signal total pour déterminer ce seuil. La variabilité de la valeur de l’écart-type entre les ensembles de données était supérieure aux niveaux de bruit d’enregistrement observés. Par conséquent, après inspection visuelle des traces, nous avons utilisé une valeur fixe du seuil de 0,3 mV, afin d’éviter le biais induit par différents niveaux d’activité de dépolarisation.
Le protocole décrit fournit une méthode standardisée et simple pour évaluer le comportement moteur et l’activité du champ neuronal, via l’enregistrement de la tension de la pince de courant extracellulaire couplé à la détection automatique des événements de dépolarisation dans le tectum optique, pour caractériser les phénotypes de type épileptiforme dans les modèles de poisson-zèbre.
Déclarations de divulgation
Les auteurs n’ont rien à divulguer.
Remerciements
Nous tenons à remercier le personnel de la plateforme d’électrophysiologie ICM où les expériences de neurophysiologie ont été réalisées. Nous remercions également Anca Marian pour son aide technique. SC a été soutenu par le Trampoline Grant #21488. EK a été soutenu par l’AFM Grant #18469 et ERC Consolidator Grant (ALS-Networks). HC a été soutenu par des bourses de doctorat de la Fondation pour la Recherche Médicale (PLP20141031462) et de l’ARSLA. Pour AD et RM, ce travail a été soutenu par trois subventions de l’Autorité nationale roumaine pour la recherche scientifique et l’innovation, CNCS-UEFISCDI (numéros de projet PN-III-P4-ID-PCE-2016-0010, PN-III-P2-2.1-PED-2016-0007 et COFUND-NEURON-NMDAR-PSY), une subvention du programme de recherche et d’innovation Horizon 2020 de l’Union européenne – convention de subvention n° 668863-SyBil-AA, et une subvention de la National Science Foundation NSF-IOS-1656830 financée par le gouvernement américain.
matériels
| Name | Company | Catalog Number | Comments |
| Agarose | Sigma-Aldrich, France | A9539 | |
| Aquarium salt | Instant Ocean, Blacksburg, VA | SS15-10 | |
| Borosilicate glass with filament | Sutter Instruments | BF100-50-10 | OD: 1.5mm, ID: 0.5 mm |
| CaCl2 | Sigma-Aldrich, France | C1016 | |
| Depdc5-atg antisense morpholino | GeneTools, OR, USA | N/A | sequence 5’- TGCCTTCATGGTGACCGTCATTTTA -3’ |
| Depdc5-mis antisense morpholino | GeneTools, OR, USA | N/A | sequence 5’- TGCgTTgATcGTGACCcTgATTTTA -3’ |
| Depdc5-splice antisense morpholino | GeneTools, OR, USA | N/A | sequence 5’- ACATTCCTGTTTCACCATAGATGAT -3’ |
| Digitizer | Molecular Devices, CA, USA | Digidata 1550 | |
| Fast Green Dye | Sigma-Aldrich, France | F7258 | Stock solution of 0.2% |
| Glass-bottom petri dishes | Ibidi, Germany | 81218 | |
| Glucose | Sigma-Aldrich, France | 68270 | |
| Grasshopper 2 camera | FLIR, BC, Canada | GRAS-03K2M-C | formerly Point Grey Research |
| HEPES | Sigma-Aldrich, France | H3375 | |
| Human wild-type DEPDC5 cDNA | Dharmacon, France | NM_001242897.1 | Accession: BC144291 Clone ID 905 |
| ImageJ software | NIH, USA | N/A | |
| KCl | Sigma-Aldrich, France | P9333 | |
| Matlab software | MathWorks, MA, USA | N/A | |
| MgCl2 | Sigma-Aldrich, France | M2670 | |
| NaCl | Sigma-Aldrich, France | S7653 | |
| NaOH | Sigma-Aldrich, France | 71687 | |
| Pancuronium bromide | Alomone Labs | P-130 | Stock solution of 60 mM in water |
| Parafilm | Sigma-Aldrich, France | P7793 | |
| Patch clamp amplifier | Molecular Devices, CA, USA | MultiClamp 700B | Computer-controled patch clamp amplifier |
| pClamp10 acquisition software | Molecular Devices | N/A | |
| Pentylenetetrazol (PTZ) | Sigma-Aldrich, France | P6500 | Stock solution of 300 mM (dissolved in recording solution) |
| Pipette puller | Narishige, Japan | PC-10 | |
| Pneumatic PicoPump | WPI, France | PV 820 | |
| Sylgard 184 kit | Sigma-Aldrich Intl. | 761036 | |
| Transfer plastic pipettes | Sigma-Aldrich, France | Z350605 | |
| Zebralab | Viewpoint, France | N/A |
Références
- Kabashi, E., Champagne, N., Brustein, E., Drapeau, P. In the swim of things: Recent insights to neurogenetic disorders from zebrafish. Trends in Genetics. 26 (8), 373-381 (2010).
- Baxendale, S., van Eeden, F., Wilkinson, R. The Power of Zebrafish in personalised medicine. Advances in Experimental Medicine and Biology. , (2017).
- Baraban, S. C., Taylor, M. R., Castro, P. A., Baier, H. Pentylenetetrazole induced changes in zebrafish behavior, neural activity and c-fos expression. Neuroscience. , (2005).
- Cunliffe, V. T. Building a zebrafish toolkit for investigating the pathobiology of epilepsy and identifying new treatments for epileptic seizures. Journal of Neuroscience Methods. , (2016).
- Griffin, A., Krasniak, C., Baraban, S. C. Advancing epilepsy treatment through personalized genetic zebrafish models. Progress in Brain Research. , (2016).
- Griffin, A., Hamling, K. R., Knupp, K., Hong, S. G., Lee, L. P., Baraban, S. C. Clemizole and modulators of serotonin signalling suppress seizures in Dravet syndrome. Brain. , (2017).
- Orellana-Paucar, A. M., et al. Insights from zebrafish and mouse models on the activity and safety of ar-turmerone as a potential drug candidate for the treatment of epilepsy. PLoS ONE. , (2013).
- Baxendale, S., et al. Identification of compounds with anti-convulsant properties in a zebrafish model of epileptic seizures. Disease Models & Mechanisms. , (2012).
- Bar-Peled, L., et al. A tumor suppressor complex with GAP activity for the Rag GTPases that signal amino acid sufficiency to mTORC1. Science. 340 (6136), 1100-1106 (2013).
- Ishida, S., et al. Mutations of DEPDC5 cause autosomal dominant focal epilepsies. Nature Genetics. , (2013).
- Dibbens, L. M., et al. Mutations in DEPDC5 cause familial focal epilepsy with variable foci. Nature Genetics. , (2013).
- Baulac, S., Weckhuysen, S. . DEPDC5-Related Epilepsy. GeneReviews®. , (1993).
- Picard, F., et al. DEPDC5 mutations in families presenting as autosomal dominant nocturnal frontal lobe epilepsy. Neurology. , (2014).
- Teng, Y., et al. Knockdown of zebrafish lgi1a results in abnormal development, brain defects and a seizure-like behavioral phenotype. Human Molecular Genetics. , (2010).
- Baraban, S. C., Dinday, M. T., Hortopan, G. A. Drug screening in Scn1a zebrafish mutant identifies clemizole as a potential Dravet syndrome treatment. Nature Communications. , (2013).
- Suls, A., et al. De novo loss-of-function mutations in CHD2 cause a fever-sensitive myoclonic epileptic encephalopathy sharing features with dravet syndrome. American Journal of Human Genetics. , (2013).
- Grone, B. P., et al. Epilepsy, behavioral abnormalities, and physiological comorbidities in syntaxin-binding protein 1 (STXBP1) mutant zebrafish. PLoS ONE. , (2016).
- de Calbiac, H., et al. DEPDC5 knockdown causes mTOR-dependent motor hyperactivity in zebrafish. Annals of Clinical and Translational Neurology. , (2018).
- Panchaud, N., Péli-Gulli, M. P., De Virgilio, C. Amino acid deprivation inhibits TORC1 through a GTPase-activating protein complex for the Rag family GTPase Gtr1. Science Signaling. , (2013).
- Lim, K. R. Q., Maruyama, R., Yokota, T. Eteplirsen in the treatment of Duchenne muscular dystrophy. Drug Design, Development and Therapy. , (2017).
- Kok, F. O., et al. Reverse genetic screening reveals poor correlation between morpholino-induced and mutant phenotypes in zebrafish. Developmental Cell. , (2015).
- Rossi, A., et al. Genetic compensation induced by deleterious mutations but not gene knockdowns. Nature. , (2015).
- Stainier, D. Y. R., et al. Guidelines for morpholino use in zebrafish. PLoS Genetics. , (2017).
- Kimmel, C. B., Ballard, W. W., Kimmel, S. R., Ullmann, B., Schilling, T. F. Stages of embryonic development of the zebrafish. Developmental dynamics an official public. 203 (3), 253-310 (1995).
- Møller, R. S., Dahl, H. A., Helbig, I. The contribution of next generation sequencing to epilepsy genetics. Expert Review of Molecular Diagnostics. , (2015).
- Allen, A. S., et al. De novo mutations in epileptic encephalopathies. Nature. , (2013).
- Dunn, P., et al. Next generation sequencing methods for diagnosis of epilepsy syndromes. Frontiers in Genetics. , (2018).
- Grone, B. P., Baraban, S. C. Animal models in epilepsy research: Legacies and new directions. Nature Neuroscience. , (2015).
- Zhang, Y., et al. Pharmacological characterization of an antisense knockdown zebrafish model of Dravet syndrome: Inhibition of epileptic seizures by the serotonin agonist fenfluramine. PLoS ONE. , (2015).
- Swaminathan, A., et al. Non-canonical mTOR-Independent Role of DEPDC5 in Regulating GABAergic Network Development. Current Biology. , (2018).
- Samarut, &. #. 2. 0. 1. ;., et al. γ-Aminobutyric acid receptor alpha 1 subunit loss of function causes genetic generalized epilepsy by impairing inhibitory network neurodevelopment. Epilepsia. , 2061-2074 (2018).
- Turrini, L., et al. Optical mapping of neuronal activity during seizures in zebrafish. Scientific Reports. , (2017).
- Rosch, R. E., Hunter, P. R., Baldeweg, T., Friston, K. J., Meyer, M. P. Calcium imaging and dynamic causal modelling reveal brain-wide changes in effective connectivity and synaptic dynamics during epileptic seizures. PLoS Computational Biology. , (2018).
- Baraban, S. C. Forebrain Electrophysiological Recording in Larval Zebrafish. Journal of Visualized Experiments. , (2013).
- Afrikanova, T., et al. Validation of the Zebrafish Pentylenetetrazol Seizure Model: Locomotor versus Electrographic Responses to Antiepileptic Drugs. PLoS ONE. , (2013).
Réimpressions et Autorisations
Demande d’autorisation pour utiliser le texte ou les figures de cet article JoVE
Demande d’autorisationExplorer plus d’articles
This article has been published
Video Coming Soon
À PROPOS DE JoVE
Copyright © 2025 MyJoVE Corporation. Tous droits réservés.