
Method Article
Clearing Teknolojisi ile Pirinç Sürgünlerinde Derin Floresan Gözlemi
Bu Makalede
Özet
Mevcut protokol, dokuların sert, kalın veya katmanlı doğası nedeniyle iç yapısal gözlemlere hazırlanması zor olan pirinç sürgünleri için bir temizleme tekniğini tanımlamaktadır. Bu yöntem, yetişkin pirinç bitkilerinde bile sürekli ve derin floresan gözlemlerini kolaylaştırır.
Özet
Kırılma indisi uyumsuzluklarını ortadan kaldıran ve otomatik floresan malzemeyi azaltan yakın zamanda geliştirilen temizleme teknolojisi, bitki dokularının iç yapılarını korurken üç boyutlu (3D) olarak gözlemlenmesini mümkün kılmıştır. Monokot model bir bitki ve küresel olarak önemli bir ürün olan pirinçte (Oryza sativa L.), kökler ve yapraklar gibi gözlemlenmesi nispeten kolay organlarda temizleme teknolojisi bildirilmiştir. Sürgün apikal meristemi (SAM) ve saplarında temizleme teknolojisinin uygulamaları da bildirilmiştir, ancak bu dokulardaki temizleme çözeltisinin (CS) zayıf penetrasyonu nedeniyle yalnızca sınırlı bir dereceye kadar. Bu dokulardaki temizleme çözeltilerinin sınırlı etkinliği, vasküler demetler ve epidermis geliştikçe ve SAM'ın su itici yapraklarla katmanlaşmasıyla gövdedeki dokuların otomatik floresansı, kalınlaşması ve sertleşmesine bağlanmıştır. Mevcut protokol, SAM / genç salkımdan gelişim sırasında sürgünlerin tabanına kadar gen ekspresyonunun sürekli ve 3D gözlemi için bir temizleme yaklaşımının optimizasyonunu bildirmektedir. Bir floresan protein raporlayıcısını ifade eden sabit doku örnekleri, titreşimli bir mikro dilimleyici kullanılarak bölümlere ayrıldı. Uygun bir kalınlık elde edildiğinde CS uygulandı. Özellikle merkezi dokuyu hedef alarak, CS'nin penetrasyon hızı ve homojenliği arttı ve dokuyu şeffaf hale getirmek için gereken süre azaldı. Ek olarak, kesilmiş bölümlerin temizlenmesi, tüm çekimin iç yapısının makro bir perspektiften gözlemlenmesini sağladı. Bu yöntem, temizlenmesi zor olan diğer bitki türlerinin dokularının derin görüntülenmesinde potansiyel uygulamalara sahiptir.
Giriş
Son zamanlarda geliştirilen temizleme teknolojisi, bitkilerin iç yapılarını korurken derin dokularını gözlemlemeyi mümkün kılmıştır 1,2,3. Dikot model bitki Arabidopsis'te, kırılma indisi uyumsuzluklarını ortadan kaldırmak ve otomatik floresan malzemeleri gidermek için temizleme teknolojisi kullanılarak floresan protein görüntüleme üzerine birçok çalışma yapılmıştır 4,5,6. Hücresel çözünürlük 9'da temizleme teknolojisi7,8 ve 3D görüntülemenin kullanımı, monokot model bir bitki ve küresel olarak önemli bir ürün olan pirinçte (Oryza sativa L.) bildirilmiş olmasına rağmen, bunlar gözlemlenmesi kolay olan kökler, yapraklar ve sürgün apikal meristemi (SAM) gibi nispeten ince ve yumuşak organlarla sınırlıdır.
Sürgün, vasküler bitkilerin yer üstü kısımlarını oluşturan ana organdır. Pirinçte, sürgünler aksiller tomurcuklar, yapraklar ve gövde10'dan oluşan bir dizi dikey olarak istiflenmiş "fitomer" den oluşur. Çekimin ucunda, SAM merkezdeki farklılaşmamış kök hücrelerden oluşur. Fitomerler, SAM'den türetilen hücrelerin farklılaşmasıyla oluşur. Bitkiler vejetatiften üreme fazına geçtikten sonra, pirinç sapları uzar ve SAM genç salkımlara farklılaşır10. Bu gelişimsel değişime, saplardaki çeşitli genlerin ekspresyonunda ve SAM / genç salkımlarda dalgalanmalar eşlik eder. Çeşitli dokularda hücre farklılaşmasının altında yatan mekanizmaları anlamak için, iç sürgün dokularındaki hücre morfolojisini ve gen ekspresyonunu yapısal olarak gözlemlemek önemlidir. Bununla birlikte, sürgündeki sapların (düğümler ve internodlar) derin görüntülenmesi, dokuya nüfuz etmek için temizleme çözeltilerinin verimsizliği nedeniyle bir zorluk teşkil etmektedir. Saplar, SAM'dan farklılaştıktan sonra yanal büyümeden hemen hızlı bir hacim artışına uğrar. Vasküler demetlerin kalınlaşması ve nodal vasküler anastomozun yatay kompleks bağlantısı nedeniyle pirinç düğümü dokularının sertleşmesi, pirinç sürgünlerinin yüksek iticiliğine ek olarak, CS'nin sap10'daki penetrasyonunu sınırlamaya katkıda bulunur.
Bu çalışmada pirinç filizi dokularındaki gen ekspresyonundaki değişiklikleri yapısal derin floresan tekniği kullanılarak gözlemlemek amaçlanmıştır. Bu çalışma, pirincin konfokal lazer mikroskobu kullanarak SAM / genç salkımdan tabana gen ekspresyonunu düz bir yüzey yerine 3D bir yapıda sürekli olarak gözlemlemesi için bir temizleme protokolünü optimize eder.
Protokol
1. Fiksatif çözeltinin hazırlanması
- 70 mL steril suyu bir cam şişeye aktarın ve 10 g paraformaldehit (PFA) ekleyin.
DİKKAT: PFA toksiktir; Bu nedenle, eldiven giyilmesi önerilir. - PFA çözeltisini çözmek için 2 mL 1 N NaOH ekleyin.
- PFA çözeltisini, PFA çözeltisi şeffaf olana kadar yaklaşık 1 saat boyunca 60 °C'de sürekli karıştırma (300-500 rpm) ile çözün.
- PFA çözeltisinin hacmini 100 mL'ye çıkarmak için sterilize su ekleyin.
NOT: PFA çözeltisi kullanımdan önce taze olarak hazırlanmıştır. Ayrıca, 4 ° C'de depolanan bir PFA çözeltisi yaklaşık 2 ay boyunca kullanılabilir. - Numune almadan hemen önce, 50 mL'lik fiksatif bir çözelti hazırlamak için 20 mL'lik %10 PFA'nın 20 mL'sine 25 mL 60 mM HEPES (pH 7.4), 3 mL 1 M sakkaroz ve 2 mL steril su ekleyin.
2. Takas çözeltisinin (CS) hazırlanması
- 50 mL steril suda 7.5 g sodyum deoksikolat, 5 g ksilitol ve 12.5 g üre (bkz.
NOT: Sodyum deoksikolat tozu, havadaki dağılımı önlemek için genel bir laboratuvar taslak odasında tartılmıştır. - CS'yi hazırlamak için bileşenleri (adım 2.1.) manyetik bir karıştırıcı kullanarak çözün.
NOT: Bu çalışmada şirket içi hazırlanmış takas çözümü (CS) kullanılmıştır, ancak ticari olarak temin edilebilen bir takas çözümü olan ClearSee ( bakınız Malzeme Tablosu) da kullanılabilir. - CS'yi 50 mL'lik konik bir tüpe aktarın ve karanlıkta oda sıcaklığında (15-30 ° C) saklayın.
NOT: CS 1 yıldan fazla saklanabilir.
3. Örnekleme
- Bitkilere zarar vermeden kökleri dikkatlice kesin. Kiri temizlemek için bitkileri suyla yıkayın.
NOT: Bu yöntem, LS'yi işaretlemek için üç yapraklı aşamadan (LS) dokuların gözlemlenmesini inceledi. Bu çalışmada 8-10 LS'de Nipponbare (Nip) ve Taichung 65 (T65) pirinç çeşitleri kullanılmıştır. - Eski dış yaprakları eller ve cımbızla soyun. Yaklaşık 2-3 yaprak kalmıştı.
- Hücrelerin ezilmesini önlemek için kayan bir hareket kullanarak dokuyu SAM / genç salkımından tabana tek kenarlı bir tıraş bıçağı ile kesin (Şekil 1A-C). SAM/genç salkımının konumu görünmüyorsa, daha uzun süre kesin.
- Pirinç filizinin enine kesiti oval bir şekle sahip olduğundan (Şekil 1D), ovalin bir tarafını çift kenarlı bir tıraş bıçağı ile uzun ekseni yönünde ince bir şekilde tıraş edin (Şekil 1E-F).
NOT: Bu adım, fiksatif çözeltinin numuneye nüfuz etmesini kolaylaştırmıştır. Yatay kesme yüzeyi, numunenin titreşimli bir mikro dilimleyici tepsisine yapışmasını kolaylaştırır (bkz. - SAM/genç salkımının konumunu, numunede beyazımsı bir rengin ortaya çıkmasıyla onaylayın (Şekil 1G). Fazla dokuyu kesin.
NOT: Numunenin uzunluğu, titreşimli bir mikro dilimleyici kullanılarak kesilebilecek maksimum uzunluğa ayarlanmalıdır. Numune çok uzunsa, birkaç parçaya bölün.
4. Örneklerin sabitlenmesi
- Pipet, fiksatif çözeltinin 1 mL'sini (1. adımda hazırlanan.) 1,5 mL'lik bir mikrosantrifüj tüpüne koyun ve hemen numunelere koyun.
NOT: Numune miktarının, fiksatif çözeltinin hacminin% 20'sini aşmadığından emin olun. - Bir parça parafin filmi 2,5 cm2'ye kesin. Parafin filmi toplar halinde şekillendirin ve fiksatif çözeltiden dışarı çıkmalarını önlemek için numunelerin üzerine yerleştirin.
- Numuneyi içeren tüpü bir kurutucuya yerleştirin. Ardından, kurutucunun kapağını kapatın ve kurutucunun içini yavaşça -0.095 MPa'ya kadar basınçsızlaştırmak için vakum pompasını çalıştırın.
- Kurutucuyu -0.095 MPa'da kapattıktan sonra, vakum pompasını kapatın ve oda sıcaklığında 30 dakika bekletin.
NOT: Vakum seviyesi, numunelerde kabarcıkların kademeli olarak ortaya çıktığı şekildeydi. - Kurutucuyu hafifçe açın ve yavaşça atmosferik basınca geri döndürün. Kurutucunun basıncını -0.095 MPa'ya düşürün, vakum pompasını kapatın ve karışımın oda sıcaklığında 30 dakika beklemesine izin verin.
- Kurutucuyu hafifçe açın ve yavaşça atmosferik basınca geri döndürün. Numune doğru şekilde sabitlenirse, fiksatif çözeltiye batar. Batırmadıysa, basıncı tekrar azaltın.
- Parafin filmi çıkarın, kapağı kapatın ve tüpleri gece boyunca 4 ° C'de tutun.
NOT: Numuneler fiksatif çözelti içinde en az 2 saat sabitlenmiştir. ~ 1 hafta saklanabilir.
5. Sabit numunenin kesilmesi
- Tüy bırakmayan mendiller kullanarak fazla fiksatif çözeltiyi numuneden çıkarın.
NOT: Aşırı fiksatif çözelti, numunenin tepsiye düzgün bir şekilde yapışmasını önler. - Numuneyi anında tutkalla titreşimli bir mikro dilimleyici tepsiye sabitleyin. Örnekleme sırasında tıraş edilen düz tarafı aşağı doğru bir tepsiye yerleştirin.
NOT: Pirinç filizinin yapısı nedeniyle, tabandan SAM / genç salkımına doğru kesilmelidir. - Kırpma koşullarını sabit numuneye göre ayarlayın.
NOT: Bu çalışmada, kırpma THICK, 130 μm olarak ayarlanmıştır; GENİŞLİK, 15 mm; SIKLIK,% 15; ve KESME HIZI, 32 mm. - RVS veya FWD düğmesi ve YUKARI veya AŞAĞI düğmesiyle numune konumunu ayarlayın. Tüm numuneler daldırılana kadar tepsiyi deiyonize suyla doldurun. Tepsiyi doldurduktan sonra START düğmesine basın. Kesilmiş numuneleri 1x PBS damlasıyla cam bir slayta yerleştirin.
NOT: Sert numune tıraş bıçağını yıprattığından, her kırpmadan sonra numunenin konumunu diğer tarafa kaydırın. - Hedef bölgeyi içeren numuneyi bir stereomikroskop altında kontrol ettikten sonra, kesilmiş numuneyi 1 mL 1x PBS içeren çok kuyulu bir plakaya aktarın.
NOT: Numunenin boyutuna göre kuyucuk sayısını seçin. Bu çalışmada 12 multiwell'li bir plaka kullanılmıştır.
6. Takas çözeltisi ile tedavi
- 1x PBS'yi çok kuyulu plakadan çıkarın ve taze 1x PBS ekleyin. 1 dakika bekletin.
- 1x PBS'yi kaldırın ve CS'yi ekleyin.
NOT: CS miktarı, numune hacminin beş katı olmalıdır. - Numuneyi içeren çok kuyulu plakayı bir kurutucuya yerleştirin. Ardından, kurutucunun kapağını kapatın ve kurutucunun içini yavaşça -0.09 MPa'ya kadar basınçsızlaştırmak için vakum pompasını çalıştırın.
- Kurutucuyu -0.09 MPa'da kapattıktan sonra, vakum pompasını kapatın ve oda sıcaklığında 1 saat bekletin.
- Kurutucuyu hafifçe açın ve yavaşça atmosferik basınca geri döndürün.
- CS'nin buharlaşmasını önlemek için kuyular arasındaki boşluğa deiyonize su dökün. çok kuyulu plakaları temizlemek için karanlıkta oda sıcaklığında saklayın.
NOT: Klorofil ve diğer bileşenleri dağıtmak için çok kuyulu plakayı her 1-2 günde bir hafifçe sallayın. - CS yeşile döndüğünde, yeni bir çözümle değiştirin.
7. Kimyasal boya ile hücre duvarı boyama
- Kalkoflor beyaz çözeltisini (1 mL, son konsantrasyon: temizleme çözeltisinde 1 mg / mL, bakınız Malzeme Tablosu) çok kuyulu bir plakaya pipetleyin. Numuneleri boya çözeltisine yerleştirin ve 1 saat boyunca inkübe edin.
NOT: Boyanın hacmi, CS hacminin% 10'unu geçmemelidir. - Lekeli numuneleri 1 mL CS içeren başka bir çok kuyucuklu plakada en az 1 saat boyunca yıkayın.
8. Konfokal lazer mikroskop ile gözlem
- Bir mikroskop slaytına bir damla CS yerleştirin ve işlenmiş (CS ile) örnekleri slayda aktarın.
NOT: İşlem görmüş numuneler yumuşak olduğundan, cımbızla dikkatlice tutulmalıdır. - CS'nin buharlaşmasını önlemek için numuneyi cam bir kapak kayması ile örtün.
NOT: CS kolayca çökelir. - Tedavi edilen örnekleri konfokal lazer mikroskop altında gözlemleyin.
NOT: Gözlemden sonra, tedavi edilen numuneler daha sonra daha fazla gözlem için CS'ye iade edilebilir.
Sonuçlar
İlk olarak, yetişkin pirinç bitkilerinin sert ve kalın dokularının temizleme işlemi ile ne ölçüde şeffaf hale getirilebileceği doğrulanmıştır. Salkım oluşum aşaması olan 9-10 LS'de nip ve genç salkımdan sürgünün tabanına kadar kesilen 12-16 mm uzunluğunda dokular kullanıldı. Genç salkım da dahil olmak üzere bu örnekler fiksatif çözeltiye sabitlendi ve temizleme tedavisinden 1-4 hafta önce ve sonra gözlendi (Şekil 2). Kesilmemiş numuneler yeşil kaldı ve 4 hafta sonra veya hatta temizleme çözeltisi ile 3 aylık tedaviden sonra şeffaf hale gelmedi (Şekil 2A). Tüm yapraklardan soyulmuş örneklerde, düğümler yeşil kalmasına rağmen, internodlar CS ile 1 haftalık tedaviden sonra beyaza döndü. Bu örnek 4 hafta sonra saydam hale gelmedi (Şekil 2B). CS ile tedaviden 3 ay sonra, sadece genç salkım şeffaf hale geldi. Elle kesilmiş örnekler, CS ile 1 haftalık tedaviden sonra, dış yapraklar ve vasküler demetler hariç, beyaza dönüştü. Yaprakların, genç salkımların ve internodların bazı kısımları yarı saydam hale geldi ancak 4 hafta sonra saydam hale gelmedi (Şekil 2C). 3 ay sonra, sadece genç salkım ve iç yapraklar şeffaf hale geldi. Titreşimli mikro dilimleyici ile 130 μm kalınlığa kadar kesilen numuneler, CS ile 1 haftalık işlemden sonra yarı saydam hale geldi ve yapraklar saydam hale geldi. 2 hafta sonra, örnek neredeyse saydamdı ve 4 hafta sonra değişmeden kaldı (Şekil 2D). 3 ay sonra, örnekler tamamen şeffaf hale geldi.
Daha sonra, temizleme tedavisinin 8 LS'deki T65 çekiminde gen ekspresyonu gözlemlerine izin verdiği derinlik doğrulandı. UBQpro: :NLS-sGFP-nClover3-mNeonGreen (UBQpro ::NGCN, bakınız Malzeme Tablosu) ifade edilen sürgünler kullanılmıştır. Doku örnekleri, titreşimli bir mikro dilimleyici kullanılarak 130 μm kalınlığa kesilmeden önce fiksatif çözeltiye sabitlendi. Örnekler daha sonra konfokal lazer mikroskop altında (lazer: 488 nm / %13, amaç: 20x, iğne deliği: 0.93 AU, ortalama: 16, dedektör kazancı: 800) CS ile 0-4 haftalık tedaviden itibaren her hafta z ekseni yönünde 10 μm aralıklarla gözlendi. Temsili sonuçlar Şekil 3'te gösterilmiştir. Tüm rakamlar, floresan yoğunluğunu karşılaştırmak için aynı işlemle ayarlandı. Tedavi edilmeyen düğümde, -40 μm derinlikte soluk floresan sinyalleri gözlendi (Şekil 3A). Gözlemlenebilir derinlik, CS tedavisinden 1 hafta sonra -60 μm ve 2 hafta sonra -90 μm idi, ancak sitoplazmanın otomatik floresansı her iki haftada da -20 μm derinlikte fark edildi. Otofloresan 3 hafta sonra biraz zayıfladı ve 4 hafta sonra artık görünmüyordu. Floresan sinyaller 3 haftada -80 μm derinlikte ve 4 haftada -100 μm derinlikte gözlendi. İnternodda, floresan sinyalleri CS tedavisi olmadan -60 μm derinlikte gözlenmiştir (Şekil 3B). Gözlemlenebilir derinlik, CS tedavisinin 1 haftasından 3 haftaya kadar -90 μm ve 4 hafta sonra -100 μm idi, ancak sitoplazmadaki otomatik floresan, 1 hafta sonra -20 μm derinlikte fark edildi. Oto-floresan 2 hafta sonra artık görünmüyordu. Yapraklarda, CS tedavisi olmadan -90 μm derinlikte floresan sinyaller gözlenmiştir (Şekil 3C). Floresan sinyaller yapraklarda düğümlerden ve internodlardan daha derin gözlendi. Bununla birlikte, floresan sinyalleri daha zayıftı. Floresan sinyalleri 1 haftalık CS tedavisinden sonra netleşti ve -120 μm derinlikte gözlendi. 2 hafta ve 3 hafta sonra, floresan sinyalleri -110 μm derinlikte ve 4 hafta sonra -120 μm derinlikte gözlendi.
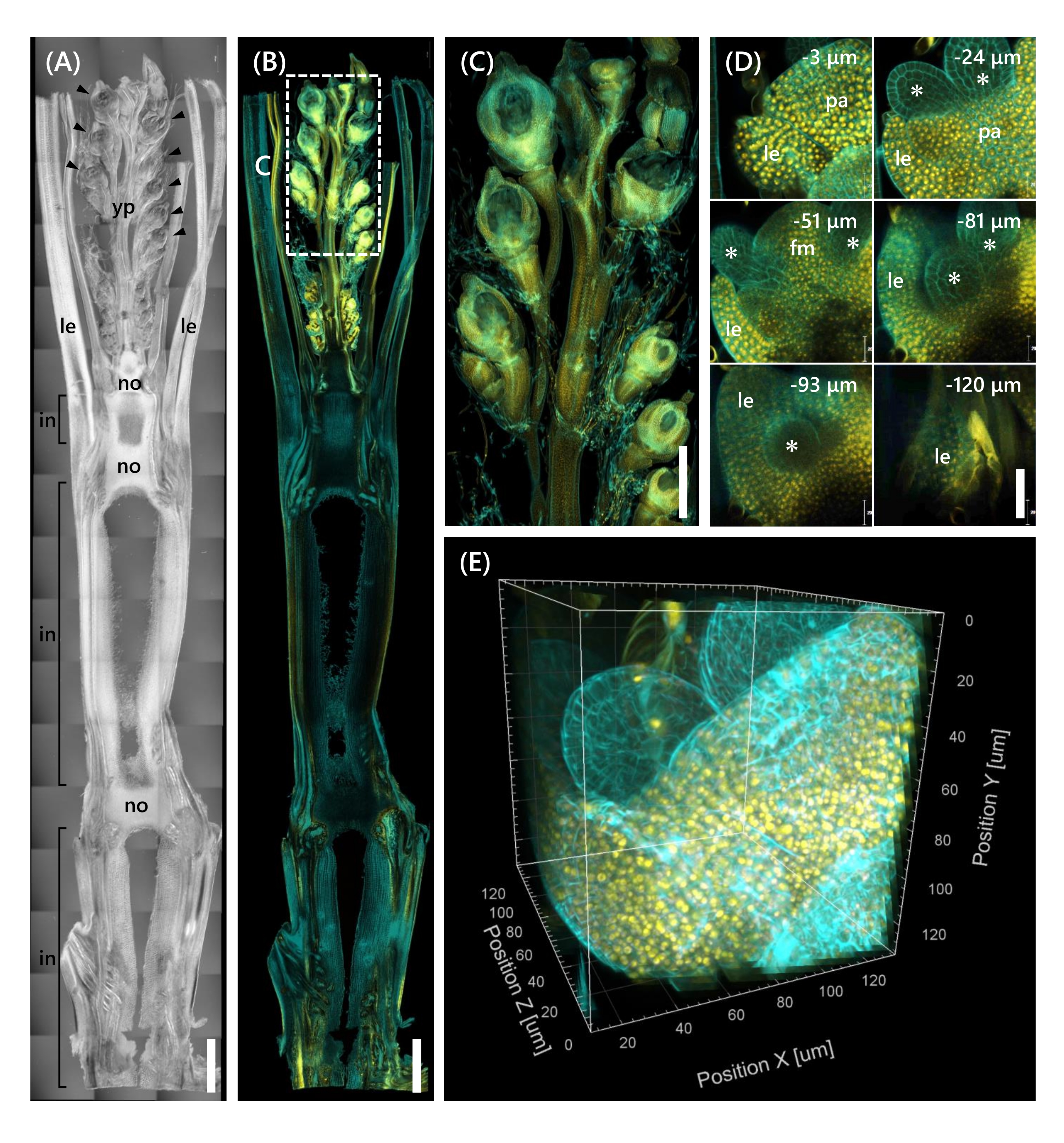
Son olarak, mOrange 12ile kaynaşmış transkripsiyon faktörü OsMADS1511'in ekspresyonu, SAM'ın vejetatiften üreme fazı13'e geçişini düzenleyen gözlenmiştir. OsMADS15-mOrange ile 10 LS'de Nip'ten alınan doku örnekleri sabitlendi, 130 μm kalınlığa kesildi ve 2 hafta boyunca CS ile muamele edildi. Şekil 4 , temizleme çözeltisinin, doğal gen ekspresyonunun floresan proteinlerini (OsMADS15-mOrange) ve floresan boya boyamasını (kalkoflor beyazı) gözlemlemek için aynı anda kullanılabileceğini göstermektedir. 0 ila -130 μm arasındaki derinliklerdeki tüm çiçek gözlemlendi ve 3D görüntü, 3 μm aralıklarla 43 z-yığın görüntüden yeniden oluşturuldu (Şekil 4E).

Şekil 1: Örnekleme konumu ve işleme. (A) Kısa gün koşulu altında 8 LS'de nip fidesi. (B) Tabanın genişletilmiş görünümü. (C) SAM/genç salkımdan tabana doğru bir numune kesilmesi. (D) C'den oval bir kesit (giriş). (E) Sürgünün bir tarafını bıçakla ince bir şekilde tıraş etmek. (F) İnce tıraşlı yüzey. (G) Düğümler arası uzamayı gösteren örnek. Ok uçlarının gösterdiği beyazımsı bölüm düğümlerdir. Üst düğümün üst tarafı genç salkıma sahiptir. Ölçek çubukları: (A) 10 cm, (B-C,E-G) 1 cm, (D) 0,5 cm. Bu şeklin daha büyük bir versiyonunu görmek için lütfen buraya tıklayın.
{kind=link}

Şekil 2: Doku temizliği numune kalınlığına ve CS tedavi süresine bağlıdır . (A) Genç salkım da dahil olmak üzere kısmi sürgünleri olan kesilmemiş örnekler. Maksimum kalınlık: 4 mm. (B) Genç salkımı açığa çıkarmak için tüm yapraklar numunelerden soyulur. Maksimum kalınlık: 2,5 mm. yp: genç salkım, içinde: internod; ok uçları düğümü gösterir. (C) 1 mm kalınlığa kadar elle kesilmiş numuneler. (D) Titreşimli mikro dilimleyici ile 130 μm kalınlığa kadar kesilmiş numuneler. Ölçek çubuğu: 5 mm. Bu şeklin daha büyük bir versiyonunu görüntülemek için lütfen buraya tıklayın.
{kind=link}

Şekil 3: Floresan gözleminin derinliği doku tipine ve CS tedavi süresine bağlıdır. UBQpro::NGCN'nin (A) düğümünde, (B) internodunda ve (C) yaprağında derin görüntülenmesi. "Birleştir", görüntülerin diferansiyel parazit kontrastı (DIC) görüntüsü ve tüm z-stack görüntüleriyle birleştirildiği anlamına gelir. Floresan gözlenmeyen görüntüler "n.d." olarak işaretlendi. (algılanmadı). Konfokal lazer mikroskopisinin ayarları aşağıdaki gibidir: lazer: 488 nm / 13%, hedef: 20x, iğne deliği: 0.93 AU, ortalama: 16, dedektör kazancı: 800, aralık: 10 μm. Ölçek çubukları: 50 μm. Bu şeklin daha büyük bir versiyonunu görüntülemek için lütfen buraya tıklayın.
{kind=link}

Şekil 4: OsMADS15-mOrange'ın derin floresan gözlemi. (A) 10 LS'de Nip'in DIC görüntüsü. yp: genç salkım, le: yaprak, hayır: düğüm, içinde: internod; ok uçları çiçekleri gösterir. (B) OsMADS15-mOrange'ın floresan görüntüsü. Sarı noktalar, çekirdekteki OsMADS15-mOrange'ın floresan proteinlerini gösterir. Hücre duvarı calcofluor beyaz solüsyonu ile camgöbeği boyandı. (C) Genç salkımının genişletilmiş görünümü. (D) Çiçeğin derin görüntülenmesi. pa: palea, le: lemma, fm: çiçek meristemi; yıldız işaretleri stamen primordia'yı gösterir. Konfokal lazer mikroskobu aşağıdaki gibi ayarlandı: OsMADS15-mOrange; lazer: 555 nm/%13, amaç: 20x, iğne deliği: 0,96 AU, ortalama: 16, dedektör kazancı: 800, aralık: 3 μm. Calcofluor beyazı; lazer: 405 nm/%3, hedef: 20x, iğne deliği: 0,93 AU, ortalama: 16, dedektör kazancı: 470, aralık: 3 μm. (E) Z-stack görüntülerinden oluşturulan şeklin (D) 3D görüntüsü. Ölçek çubukları: (A-B) 1 mm, (C) 500 μm, (D) 40 μm. Bu şeklin daha büyük bir versiyonunu görüntülemek için lütfen buraya tıklayın.
{kind=link}
Tartışmalar
Protokolün kritik adımları
Bu protokoldeki kritik adımlar sabitleme ve kırpmadır. Pirinç sürgünleri, fiksatif çözeltinin penetrasyonunu sınırlayan sert, kalın veya katmanlı dokulara sahiptir. Fiksatif çözeltinin geçirgenliğini arttırmak için, Şekil 1E-F'de gösterildiği gibi, dokuların bir tarafı örneklemede ince bir şekilde tıraş edildi. Ek olarak, vakum işlemleri daha yüksek basınç kullanılarak iki kez tekrarlandı. Ayrıca, numuneler 4 ° C'de normal 2 saatlik fiksasyon yerine gece boyunca 4 ° C'de sabitlendi.
Kırpma adımındaki kilit nokta, kısa bir CS tedavisinin ardından iç yapılarını korurken floresan proteinleri gözlemlemek için hazırlanacak dokuların kalınlığını belirlemektir. Şekil 2C'de gösterildiği gibi, mümkün olduğunca ince elle kesilmiş 1 mm kalınlığındaki numuneler, 3 aylık CS tedavisinden sonra bile sadece sınırlı sayıda dokuda saydam hale geldi. Bu nedenle, kırpma adımı, yetişkin pirinç sürgünlerinin derin floresan gözlemi için gereklidir. Bu çalışmada, örnekler Şekil 2D'de gösterildiği gibi 130 μm kalınlığa kesilmiştir. 130 μm kalınlığı, 1 haftalık CS tedavisinden sonra yaprakların ve 2 hafta sonra tüm numunenin temizlenmesine izin verdi. Bu çalışmada 9-10 LS'de yetişkin pirinç sürgünleri kullanılmıştır. Genç pirinç sürgünlerinden gelen kalın fakat daha yumuşak dokular, CS tedavisi ile daha hızlı temizlenebilir. Numunelerin kalınlığı ve CS tedavisinin süresi, gözlemlenecek 3D yapının doku tipine, durumuna ve kalınlığına göre ayarlanmalıdır.
Değişiklikler ve sorun giderme yöntemleri
CS düşük sıcaklıklarda kolayca çökeldi. Çökelmiş CS, floresan proteinleri koruyamaz; Bu nedenle, numuneler uygun sıcaklıkta saklanırken dikkatli olunmalıdır. Ek olarak, hem CS hem de fiksatif çözeltinin antiseptik etkisi yoktur; bu nedenle, floresan proteinler kontamine olursa parçalanacaktır. Toprakta yetişen pirinç mantar büyümesine eğilimlidir; Bu nedenle, numunelerin örneklenmesi ve işlenmesi, kontaminasyonu önlemek için dikkatli bir şekilde yapılmalıdır.
Tampondaki aşırı floresan boyalar arka plan floresansı verebilir ve mikroskobik gözlemlere müdahale edebilir. Örneğin, daha önce Evans mavi boyası içeren bir calcofluor beyaz çözeltisi kullanılmıştır. 1 saat boyama ve 1 saat yıkama işleminden sonra, OsMADS15-mOrange'ın floresan proteinleri 555 nm lazer kullanılarak gözlemlendi. Bununla birlikte, Evans mavi boyasından türetilen arka plan floresansı nedeniyle floresan proteinleri gözlenememiştir. Bu arka plan floresansı, numunelerin 2 saat boyunca yıkanmasıyla neredeyse ortadan kaldırıldı. Dahası, örnekler gece boyunca bırakılırsa floresan proteinler daha netti. Bu nedenle, bu çalışmada saf bir calcofluor beyaz çözeltisi kullanılmıştır. Floresan boyadan elde edilen arka plan floresansı, gözlemlerden önce farklı lazer dalga boyları kullanılarak kontrol edilmelidir.
Yöntemin sınırlamaları
Şekil 3'te gösterildiği gibi, 2 haftalık CS tedavisinden sonra 130 μm kalınlığındaki örneklerde derin floresan proteinler gözlenmiştir. Bu, 130 μm kalınlığındaki numunenin 2 haftalık CS tedavisinden sonra saydam hale geldiği Şekil 2D'de gösterilen sonuçlarla tutarlıdır. Bununla birlikte, Şekil 3A'da gösterildiği gibi, sitoplazmanın oto-floresansı 2 hafta sonra düğümlerde hala fark edildi ve sadece 4 haftalık CS tedavisinden sonra tamamen çıkarıldı. Düğümler yüksek hücre yoğunluğuna sahiptir ve bu nedenle otomatik floresan malzemelerin çıkarılması için daha uzun bir zaman gerektirir.
Şekil 3C'de gösterildiği gibi, CS tedavisi olmayan yapraklarda derin floresan proteinler gözlenmiştir, ancak parlaklık, 20 μm'lik aynı derinlikteki düğümlerde ve internodlardakinden daha zayıftı. 1 haftalık CS tedavisinden sonra, floresan proteinleri daha parlaktı. Klorofil yapraklarda bol miktarda bulunur ve 488 nm uyarma ışığını emer. Ayrıca, 555 nm lazer kullanılarak floresan proteinlerin gözlemlenmesine müdahale edebilen turuncu / kırmızı otomatik floresana sahiptirler. 1 haftalık CS tedavisinden sonra, klorofil ve diğer otomatik floresan malzemeler çıkarıldı ve yüksek sinyal-gürültü oranlı görüntüler elde edildi.
2 hafta ve 4 haftalık CS tedavisinden sonra dokularda gözlenebilen derinlikler, floresan proteinleri 4 hafta sonra daha zayıf görünmesine rağmen, anlamlı olarak farklı değildi (Şekil 3). Normalde, floresan proteinlerinin ve otomatik floresanın parlaklığı zamanla zayıflar ve bu da daha yüksek bir sinyal-gürültü oranı ile sonuçlanır. Bu nedenle, floresan proteinler mikroskobik koşullar ve görüntü işleme ayarlanarak daha net gözlemlenebilir. Bu sonuçlara dayanarak, 2 haftalık CS tedavisinin, numune koşullarımız göz önüne alındığında, derin floresan proteinlerinin gözlemlenmesini kolaylaştırabileceği sonucuna varılmıştır. Bununla birlikte, otomatik floresan malzemeleri tamamen dışlayan daha net görüntüler gözlemlemek için 4 haftaya ihtiyaç vardır.
Vasküler demetler ve çok kollu hücreler gibi güçlü otofloresansı olan yapılar CS'de temizlenemez. Bu yapıları otofloresan olmadan gözlemlemek için, bir zaman geçidi yöntemi12 kullanmak veya floresan spektrumunun spektroskopisi ile görüntü elde etmek gerekir. İki fotonlu mikroskop, daha kalın dokular gözlenirse daha derin dokuları gözlemlemek için daha uygun olabilir.
Yöntemin mevcut ve alternatif yöntemler açısından önemi
Genel olarak, pirinç bitkilerinin iç yapıları kriyostat veya vibratom kesitleme kullanılarak gözlemlenmiştir. Bir kriyostat, daha kolay gözlem sağlayan ince kesitlerin hazırlanması için uygundur, ancak numunelerin hazırlanması ve ekipmanın çalışması zaman alıcıdır. Orijinal 3B yapıyı ince kesitlerden yeniden yapılandırmak da zordur. Vibratomun kullanımı nispeten kolaydır ve kalın bölümler üretmek için uygundur. Bununla birlikte, hedef dokuların kalın bölümleri, ışığın ulaşamayacağı derin dokuların değil, yalnızca kesilmiş yüzeyin gözlemlerine izin verir. Bu nedenlerden dolayı, her iki yöntem de derin floresan gözlemleri için uygun değildir.
Bu çalışma, mevcut yöntemleri birleştirerek CS'nin sınırlı doku penetrasyonu ve konfokal mikroskop altında zayıf nesne çözünürlüğü gibi pirinç sürgünlerinde derin floresan gözlemindeki zorlukları ele almıştır. Şekil 4'te gösterildiği gibi, genç salkımdan tabana kadar yetişkin pirinç sürgünlerinin derin dokularında eksprese edilen floresan proteinleri (OsMADS15-mOrange) gözlemledik. Şekil 4D, çiçeğe odaklanır ve 3 μm aralıklarla derin floresan proteinleri gösterir. 2 haftalık CS tedavisinden sonra -130 μm derinliğin üzerindeki dokular gözlendi, ancak CS tedavisi olmadan aynı büyüklükte ve büyüme aşamasında çiçekte sadece -27 μm derinlik içindeki dokular gözlendi (veriler gösterilmedi). Mevcut geliştirilmiş protokol, sadece genlerin aşırı ekspresyonunun değil, aynı zamanda yetişkin pirinç sürgünlerinin derin dokularındaki doğal gen ekspresyonunun da gözlemlenmesine izin verdi.
Yöntemin belirli araştırma alanlarındaki önemi ve potansiyel uygulamaları
Yetişkin pirinç sürgünlerinin derin floresan gözlemini optimize eden bu protokol, gereksiz dokuları keserek ve CS'nin geçirgenliğini artırarak sert, kalın veya katmanlı dokuların verimli bir şekilde temizlenmesini sağlar. Ek olarak, analiz için numunelerin kalınlığı, normalde kalın veya opak dokuları çözemeyen bir konfokal lazer mikroskobu kullanılarak sürekli ve yapısal derin floresan gözlemine izin verecek şekilde optimize edilmiştir.
Pirinç örneklerini farklı büyüme aşamalarında karşılaştırmak zordur, çünkü floresan proteinler fiksatif ve PBS çözeltilerinde zamanla bozulur. Bununla birlikte, CS'deki floresan proteinler 5 aydan fazla saklanabilir1. CS'nin uzun raf ömrü, pirinçte derin floresan gözlemi için büyük bir avantajdır.
Son zamanlarda, iç yapılarını korurken derin dokuları 3D olarak gözlemlemeyi mümkün kılan birçok temizleme teknolojisi geliştirilmiştir. Bu teknolojiler gelişmeye devam etti ve yeni takas çözümleri geliştirildi. İyi bir örnek, etkili klorofil giderimi ve daha parlak floresan tespiti sağlayan iTOMEI14'tür. Başka bir örnek, temizleme tedavisi sırasında dokuların kahverengileşmesini önleyen ve şeffaf görünmelerini sağlayan ClearSeeAlpha15'tir. Bu takas çözümlerinin mevcut yöntemle birleştirilmesi, daha verimli ve etkili takasın yapılmasına olanak sağlayabilir.
Mevcut yöntemin sadece pirincin değil, diğer bitkilerin de derinlemesine görüntülenmesiyle yeni bilgiler edinilmesine yardımcı olması bekleniyor.
Açıklamalar
Yazarların beyan edecekleri herhangi bir çıkar çatışması yoktur.
Teşekkürler
Dr. R. Terada, Dr. Z. Shimatani ve Dr. H. Tsuji'ye bize OsMADS15-mOrange tohumlarını sağladıkları için teşekkür ederiz; Dr. D. Kurihara, bize NGCN yapısını sağladığı için; ve Dr. R. Shim, makalemizi düzenlediği için. Bu çalışma JSPS KAKENHI (hibe numaraları JP20H05912, 20H05778, 20H05779) ve SATREPS programı (hayır. JPMJSA1706) JST ve JICA'nın.
Malzemeler
| Name | Company | Catalog Number | Comments |
| 1.5 mL microcentrifuge tube | BIO-BIK | ST-0150F | |
| 12-multiwell plate | Corning | 353043 | |
| 50 mL conical tube | Corning | 352070 | |
| Calcofluor white solution | Sigma-Aldrich | 910090 | |
| ClearSee | FUJIFILM Wako Pure Chemical | 031-25151 | This can be made or purchased. |
| Confocal laser microscope | Carl Zeiss | LSM700 | |
| Desiccator | SANPLATEC | Custom made of acrylic. 30 cm (L), 30 cm (W), 14.5 cm (H) | |
| Glass coverslip (18 × 18 No.1) | MATSUNAMI | C018181 | |
| HEPES | FUJIFILM Wako Pure Chemical | 342-01375 | |
| Microscope slide (76 × 26) | MATSUNAMI | S2441 | |
| Paraffin film | Bemis | PM-996 | |
| Paraformaldehyde | FUJIFILM Wako Pure Chemical | 162-16065 | |
| Sodium deoxycholate | Tokyo Chemical Industry | C0316 | |
| Sucrose | FUJIFILM Wako Pure Chemical | 190-00013 | |
| UBQpro::NLS-sGFP-nClover3-mNeonGreen (UBQpro::NGCN) | provided by Dr. Kurihara | ||
| Urea | FUJIFILM Wako Pure Chemical | 211-01213 | |
| Vacuum pump | AS One | AS-01 | |
| Vibrating micro-slicer | DOSAKA | DTK-3000 | |
| Xylitol | FUJIFILM Wako Pure Chemical | 248-00545 |
Referanslar
- Kurihara, D., Mizuta, Y., Sato, Y., Higashiyama, T. ClearSee: A rapid optical clearing reagent for whole-plant fluorescence imaging. Development. 142 (23), 4168-4179 (2015).
- Hasegawa, J., et al. Three-dimensional imaging of plant organs using a simple and rapid transparency technique. Plant and Cell Physiology. 57 (3), 462-472 (2016).
- Musielak, T. J., Slane, D., Liebig, C., Bayer, M. A versatile optical clearing protocol for deep tissue imaging of fluorescent proteins in Arabidopsis thaliana. PLoS One. 11 (8), 0161107 (2016).
- Pandey, K. B., et al. Plant roots sense soil compaction through restricted ethylene diffusion. Science. 371 (6526), 276-280 (2021).
- Ejaza, M., Bencivenga, S., Tavaresa, R., Busha, M., Sablowski, R. ARABIDOPSIS THALIANA HOMEOBOX GENE 1 controls plant architecture by locally restricting environmental responses. Proceedings of the National Academy of Sciences of the United States of America. 118 (17), (2021).
- Smetana, O., et al. High levels of auxin signalling define the stem-cell organizer of the vascular cambium. Nature. 565 (7740), 485-489 (2019).
- Chu, T. T. H., et al. Sub-cellular markers highlight intracellular dynamics of membrane proteins in response to abiotic treatments in rice. Rice. 11 (1), 23 (2018).
- Nagaki, K., Yamaji, N., Murata, M. ePro-ClearSee: A simple immunohistochemical method that does not require sectioning of plant samples. Scientific Reports. 7, 42203 (2017).
- Sato, M., Akashi, H., Sakamoto, Y., Matsunaga, S., Tsuji, H. Whole-tissue three-dimensional imaging of rice at single-cell resolution. International Journal of Molecular Sciences. 23 (1), 40 (2022).
- Matsuo, T., Hoshikawa, K. Science of the Rice Plant: Morphology. Food and Agriculture Policy Research Center. , (1993).
- Kobayashi, K., et al. Inflorescence meristem identity in rice is specified by overlapping functions of three AP1/FUL-Like MADS box genes and PAP2, a SEPALLATA MADS box gene. Plant Cell. 24 (5), 1848-1859 (2012).
- Tamaki, S., et al. FT-like proteins induce transposon silencing in the shoot apex during floral induction in rice. Proceedings of the National Academy of Sciences of the United States of America. 112 (8), 901-910 (2015).
- Kodama, Y. Time gating of chloroplast autofluorescence allows clearer fluorescence imaging In Planta. PLoS One. 11 (3), 0152484 (2016).
- Sakamoto, Y., et al. Improved clearing method contributes to deep imaging of plant organs. Communications Biology. 5 (1), 12 (2022).
- Kurihara, D., Mizuta, Y., Nagahara, S., Higashiyama, T. ClearSeeAlpha: Advanced optical clearing for whole-plant imaging. Plant and Cell Physiology. 62 (8), 1302-1310 (2021).
Yeniden Basımlar ve İzinler
Bu JoVE makalesinin metnini veya resimlerini yeniden kullanma izni talebi
Izin talebiThis article has been published
Video Coming Soon
JoVE Hakkında
Telif Hakkı © 2020 MyJove Corporation. Tüm hakları saklıdır