
Method Article
Osservazione profonda della fluorescenza nei germogli di riso tramite tecnologia di compensazione
In questo articolo
Riepilogo
Il presente protocollo descrive una tecnica di compensazione per i germogli di riso, che sono difficili da preparare per le osservazioni strutturali interne a causa della natura dura, spessa o stratificata dei tessuti. Questo metodo facilita osservazioni continue e profonde di fluorescenza, anche in piante di riso adulte.
Abstract
La tecnologia di clearing recentemente sviluppata che elimina i disallineamenti dell'indice di rifrazione e diminuisce il materiale auto-fluorescente ha permesso di osservare i tessuti vegetali in tre dimensioni (3D) preservando le loro strutture interne. Nel riso (Oryza sativa L.), una pianta modello monocotiledone e una coltura importante a livello globale, la tecnologia di pulizia è stata riportata in organi relativamente facili da osservare, come le radici e le foglie. Sono state riportate anche applicazioni della tecnologia di clearing nel meristema apicale (SAM) e negli steli, ma solo in misura limitata a causa della scarsa penetrazione della soluzione di compensazione (CS) in questi tessuti. La limitata efficienza delle soluzioni di compensazione in questi tessuti è stata attribuita all'autofluorescenza, all'ispessimento e all'indurimento dei tessuti nello stelo mentre i fasci vascolari e l'epidermide si sviluppano e alla stratificazione del SAM con foglie idrorepellenti. Il presente protocollo riporta l'ottimizzazione di un approccio di clearing per l'osservazione continua e 3D dell'espressione genica dalla pannocchia SAM/giovane alla base dei germogli durante lo sviluppo. I campioni di tessuto fissi che esprimono un reporter proteico fluorescente sono stati tagliati in sezioni utilizzando una micro-affettatrice vibrante. Quando è stato raggiunto uno spessore appropriato, è stato applicato il CS. Prendendo di mira specificamente il tessuto centrale, il tasso di penetrazione e l'uniformità del CS sono aumentati e il tempo necessario per rendere il tessuto trasparente è diminuito. Inoltre, la pulizia delle sezioni tagliate ha permesso l'osservazione della struttura interna dell'intero servizio fotografico da una prospettiva macro. Questo metodo ha potenziali applicazioni nell'imaging profondo di tessuti di altre specie vegetali difficili da eliminare.
Introduzione
La tecnologia di pulizia recentemente sviluppata ha permesso di osservare i tessuti profondi delle piante preservando la loro struttura interna 1,2,3. Nell'impianto modello di dicot Arabidopsis, sono stati condotti molti studi sull'imaging delle proteine fluorescenti utilizzando la tecnologia di compensazione per eliminare i disallineamenti dell'indice di rifrazione e rimuovere i materiali auto-fluorescenti 4,5,6. Sebbene l'uso della tecnologia di clearing7,8 e l'imaging 3D a risoluzione cellulare9 siano stati riportati nel riso (Oryza sativa L.), una pianta modello monocotiledone e una coltura importante a livello globale, questi sono limitati a organi relativamente sottili e morbidi, come radici, foglie e meristema apicale (SAM), che sono facili da osservare.
Il germoglio è l'organo principale che costituisce le parti fuori terra delle piante vascolari. Nel riso, i germogli sono composti da una serie di "fitomeri" impilati verticalmente, comprendenti gemme ascellari, foglie e il gambo10. Sulla punta delle riprese, il SAM è composto da cellule staminali indifferenziate al centro. I fitomeri sono formati dalla differenziazione delle cellule derivate dal SAM. Dopo che le piante passano dalla fase vegetativa a quella riproduttiva, gli steli di riso si allungano e il SAM si differenzia in giovani pannocchie10. Questo cambiamento dello sviluppo è accompagnato da fluttuazioni nell'espressione di vari geni negli steli e nelle pannocchie SAM / giovani. Per comprendere i meccanismi alla base della differenziazione cellulare in vari tessuti, è importante osservare strutturalmente la morfologia cellulare e l'espressione genica nei tessuti germogli interni. Tuttavia, l'imaging profondo degli steli (nodi e internodi) nelle riprese rappresenta una sfida a causa dell'inefficienza delle soluzioni di pulizia per penetrare nel tessuto. Gli steli subiscono immediatamente un rapido aumento di volume dalla crescita laterale dopo la differenziazione dal SAM. L'indurimento dei tessuti dei nodi del riso dovuto all'ispessimento dei fasci vascolari e il legame complesso orizzontale dell'anastomosi vascolare nodale, oltre all'elevata repellenza dei germogli di riso, contribuiscono tutti a limitare la penetrazione del CS negli steli10.
Questo studio mirava a osservare i cambiamenti nell'espressione genica nei tessuti dei germogli di riso utilizzando una tecnica strutturale di fluorescenza profonda. Questo lavoro ottimizza un protocollo di compensazione per il riso per osservare continuamente l'espressione genica dalla pannocchia SAM / giovane alla base in una struttura 3D, piuttosto che su una superficie piana, utilizzando un microscopio laser confocale.
Protocollo
1. Preparazione della soluzione fissativa
- Trasferire 70 ml di acqua sterile in una bottiglia di vetro e aggiungere 10 g di paraformaldeide (PFA).
ATTENZIONE: il PFA è tossico; Quindi, si consiglia di indossare guanti. - Aggiungere 2 mL di 1 N NaOH per sciogliere la soluzione di PFA.
- Sciogliere la soluzione di PFA con agitazione continua (300-500 rpm) a 60 °C per circa 1 ora fino a quando la soluzione di PFA è trasparente.
- Aggiungere acqua sterilizzata per aumentare il volume della soluzione di PFA a 100 ml.
NOTA: La soluzione di PFA è stata preparata di fresco prima dell'uso. Inoltre, una soluzione di PFA conservata a 4 °C può essere utilizzata per circa 2 mesi. - Immediatamente prima del campionamento, aggiungere 25 mL di 60 mM HEPES (pH 7,4), 3 mL di 1 M di saccarosio e 2 mL di acqua sterile in 20 mL di 10 % di PFA per preparare una soluzione fissativa da 50 mL.
2. Preparazione della soluzione di compensazione (CS)
- Pesare e mescolare 7,5 g di desossicolato di sodio, 5 g di xilitolo e 12,5 g di urea (vedere Tabella dei materiali) in 50 ml di acqua sterile.
NOTA: La polvere di desossicolato di sodio è stata pesata in una camera di tiraggio di laboratorio generale per prevenire la dispersione nell'aria. - Sciogliere gli ingredienti (punto 2.1.) usando un agitatore magnetico per preparare il CS.
NOTA: Il presente studio ha utilizzato una soluzione di compensazione (CS) preparata internamente, ma può essere utilizzata anche una soluzione di compensazione disponibile in commercio, ClearSee (vedi Tabella dei materiali). - Trasferire il CS in un tubo conico da 50 mL e conservarlo a temperatura ambiente (15-30 °C) al buio.
NOTA: Il CS può essere conservato per più di 1 anno.
3. Campionamento
- Tagliare le radici con cura senza danneggiare le piante. Lavare le piante con acqua per rimuovere lo sporco.
NOTA: Questo metodo ha studiato l'osservazione dei tessuti dallo stadio a tre foglie (LS) alla bandiera LS. Il presente studio ha utilizzato le cultivar di riso Nipponbare (Nip) e Taichung 65 (T65) all'8-10 LS. - Staccare le vecchie foglie esterne con le mani e le pinzette. C'erano circa 2-3 foglie rimaste.
- Tagliare il tessuto dalla pannocchia SAM/giovane alla base con un rasoio a taglio singolo usando un movimento scorrevole per evitare di schiacciare le cellule (Figura 1A-C). Se la posizione della SAM/giovane pannocchia non è visibile, tagliarla più a lungo.
- Poiché la sezione trasversale del germoglio di riso ha una forma ovale (Figura 1D), radere un lato dell'ovale in direzione del suo asse lungo con un rasoio a doppio taglio (Figura 1E-F).
NOTA: questo passaggio ha reso più facile per la soluzione fissativa penetrare nel campione. Una superficie di taglio orizzontale rende il campione più facile da attaccare a un vassoio vibrante micro-affettatrice (vedere Tabella dei materiali). - Confermare la posizione della SAM/giovane pannocchia mediante la comparsa di un colore biancastro nel campione (Figura 1G). Tagliare il tessuto in eccesso.
NOTA: la lunghezza del campione deve essere regolata alla lunghezza massima che può essere rifilata utilizzando una micro-affettatrice vibrante. Se il campione è troppo lungo, dividerlo in più pezzi.
4. Fissazione dei campioni
- Pipettare 1 mL della soluzione fissativa (preparata al punto 1.) in una provetta da microcentrifuga da 1,5 mL e inserire immediatamente i campioni.
NOTA: Assicurarsi che la quantità di campione non superi il 20% del volume della soluzione fissativa. - Tagliare un pezzo di pellicola di paraffina in 2,5 cm2. Modellare il film di paraffina in palline e posizionarle sopra i campioni per evitare che fluttuino fuori dalla soluzione fissativa.
- Porre la provetta contenente il campione in un essiccatore. Quindi, chiudere il coperchio dell'essiccatore e avviare la pompa per vuoto per depressurizzare lentamente l'interno dell'essiccatore fino a -0,095 MPa.
- Dopo aver chiuso l'essiccatore a -0,095 MPa, spegnere la pompa per vuoto e lasciarla riposare per 30 minuti a temperatura ambiente.
NOTA: Il livello di vuoto era tale che le bolle apparivano gradualmente nei campioni. - Aprire leggermente l'essiccatore e riportarlo lentamente alla pressione atmosferica. Ridurre la pressione dell'essiccatore a -0,095 MPa, spegnere la pompa per vuoto e lasciare riposare la miscela per 30 minuti a temperatura ambiente.
- Aprire leggermente l'essiccatore e riportarlo lentamente alla pressione atmosferica. Se il campione è fissato correttamente, affonda nella soluzione fissativa. Se non è affondato, ridurre nuovamente la pressione.
- Rimuovere la pellicola di paraffina, chiudere il coperchio e tenere i tubi per una notte a 4 °C.
NOTA: I campioni sono stati fissati in soluzione fissativa per almeno 2 ore. Può essere conservato per ~ 1 settimana.
5. Taglio del campione fisso
- Rimuovere la soluzione fissativa in eccesso dal campione utilizzando salviette prive di lanugine.
NOTA: la soluzione fissativa in eccesso impedisce al campione di aderire correttamente al vassoio. - Fissare il campione su un vassoio vibrante per micro-affettatrici con colla istantanea. Posizionalo su un vassoio con il lato piatto verso il basso, che è stato rasato durante il campionamento.
NOTA: A causa della struttura del germoglio di riso, deve essere tagliato dalla base verso la SAM / giovane pannocchia. - Impostare le condizioni di rifilatura in base al campione fisso.
NOTA: In questo studio, il taglio è stato impostato su THICK, 130 μm; LARGHEZZA, 15 mm; FREQUENZA, 15%; e VELOCITÀ DI TAGLIO, 32 mm. - Regolare la posizione del campione con il pulsante RVS o FWD e il pulsante SU o GIÙ . Riempire il vassoio con acqua deionizzata fino a quando tutti i campioni sono immersi. Dopo aver riempito il vassoio, premere il pulsante START . Posizionare i campioni tagliati su un vetrino con una goccia di 1x PBS.
NOTA: spostare la posizione del campione sull'altro lato dopo ogni taglio poiché il campione duro consuma la lama del rasoio. - Dopo aver controllato il campione contenente il sito target sotto uno stereomicroscopio, trasferire il campione tagliato su una piastra multipozzetto con 1 mL di 1x PBS.
NOTA: Scegliere il numero di pozzetti in base alla dimensione del campione. In questo studio è stata utilizzata una piastra da 12 pozzetti.
6. Trattamento con la soluzione di compensazione
- Rimuovere il PBS 1x dalla piastra multipozzetto e aggiungere 1x PBS fresco. Lasciare riposare per 1 minuto.
- Rimuovere 1x PBS e aggiungere il CS.
NOTA: la quantità di CS deve essere cinque volte il volume del campione. - Collocare la piastra multipozzetto contenente il campione in un essiccatore. Quindi, chiudere il coperchio dell'essiccatore e avviare la pompa per vuoto per depressurizzare lentamente l'interno dell'essiccatore fino a -0,09 MPa.
- Dopo aver chiuso l'essiccatore a -0,09 MPa, spegnere la pompa per vuoto e lasciarla riposare per 1 ora a temperatura ambiente.
- Aprire leggermente l'essiccatore e riportarlo lentamente alla pressione atmosferica.
- Versare acqua deionizzata nello spazio tra i pozzetti per evitare l'evaporazione del CS. Conservare le piastre multipozzetto a temperatura ambiente al buio per la pulizia.
NOTA: Agitare delicatamente la piastra multipozzetto ogni 1-2 giorni per diffondere la clorofilla e altri componenti. - Quando il CS diventa verde, sostituirlo con una soluzione fresca.
7. Colorazione della parete cellulare con colorante chimico
- Pipet la soluzione bianca di calcofluor (1 mL, concentrazione finale: 1 mg/mL nella soluzione di clearing, vedere Tabella dei materiali) in una piastra multipozzetto. Posizionare i campioni nella soluzione colorante e incubare per 1 ora.
NOTA: Il volume del colorante non deve superare il 10% del volume del CS. - Lavare i campioni macchiati in un'altra piastra multipozzetto con 1 mL di CS per almeno 1 ora.
8. Osservazione con un microscopio laser confocale
- Posizionare una goccia di CS su un vetrino da microscopio e trasferire i campioni trattati (con CS) sul vetrino.
NOTA: Poiché i campioni trattati sono morbidi, devono essere maneggiati con cura con una pinzetta. - Coprire il campione con un vetrino di vetro per evitare l'evaporazione del CS.
NOTA: il CS precipita facilmente. - Osservare i campioni trattati al microscopio laser confocale.
NOTA: Dopo l'osservazione, i campioni trattati possono essere restituiti al CS per ulteriori osservazioni successive.
Risultati
In primo luogo, è stata convalidata la misura in cui i tessuti duri e spessi delle piante di riso adulte potrebbero essere resi trasparenti dal trattamento di pulizia. Nip al 9-10 LS, che è lo stadio di formazione della pannocchia, e sono stati utilizzati tessuti lunghi 12-16 mm tagliati dalla giovane pannocchia alla base delle riprese. Questi campioni, compresa la giovane pannocchia, sono stati fissati nella soluzione fissativa e osservati prima e dopo 1-4 settimane del trattamento di compensazione (Figura 2). I campioni non tagliati sono rimasti verdi e non sono diventati trasparenti dopo 4 settimane o anche dopo 3 mesi di trattamento con la soluzione di compensazione (Figura 2A). Nei campioni che sono stati staccati da tutte le foglie, gli internodi sono diventati bianchi dopo 1 settimana di trattamento con il CS, sebbene i nodi siano rimasti verdi. Questo campione non è diventato trasparente dopo 4 settimane (Figura 2B). Dopo 3 mesi di trattamento con il CS, solo la giovane pannocchia è diventata trasparente. I campioni tagliati a mano sono diventati bianchi, ad eccezione delle foglie esterne e dei fasci vascolari, dopo 1 settimana di trattamento con il CS. Alcune parti delle foglie, la giovane pannocchia e gli internodi sono diventati traslucidi ma non sono diventati trasparenti dopo 4 settimane (Figura 2C). Dopo 3 mesi, solo la giovane pannocchia e le foglie interne sono diventate trasparenti. I campioni che sono stati tagliati a 130 μm di spessore con la micro-affettatrice vibrante sono diventati traslucidi dopo 1 settimana di trattamento con il CS, con le foglie che diventano trasparenti. Dopo 2 settimane, il campione era quasi trasparente ed è rimasto invariato dopo 4 settimane (Figura 2D). Dopo 3 mesi, i campioni sono diventati completamente trasparenti.
Successivamente, è stata convalidata la profondità con cui il trattamento di compensazione ha permesso osservazioni di espressione genica nel germoglio di T65 all'8 LS. Sono stati utilizzati i germogli espressi UBQpro:: NLS-sGFP-nClover3-mNeonGreen (UBQpro ::NGCN, vedi Tabella dei materiali). I campioni di tessuto sono stati fissati nella soluzione fissativa prima di tagliarli a 130 μm di spessore utilizzando una micro-affettatrice vibrante. I campioni sono stati poi osservati al microscopio laser confocale (laser: 488 nm/13%, obiettivo: 20x, foro stenopeico: 0,93 UA, media: 16, guadagno del rivelatore: 800) a intervalli di 10 μm nella direzione dell'asse z ogni settimana da 0-4 settimane di trattamento con il CS. I risultati rappresentativi sono mostrati nella Figura 3. Tutte le cifre sono state regolate con la stessa elaborazione per confrontare l'intensità della fluorescenza. Nel nodo senza trattamento, sono stati osservati deboli segnali fluorescenti a una profondità di -40 μm (Figura 3A). La profondità osservabile era di -60 μm dopo 1 settimana di trattamento con CS e -90 μm dopo 2 settimane, ma l'autofluorescenza del citoplasma era evidente in entrambe le settimane ad una profondità di -20 μm. L'autofluorescenza è diventata leggermente più debole dopo 3 settimane e non era più visibile dopo 4 settimane. Segnali fluorescenti sono stati osservati ad una profondità di -80 μm a 3 settimane e ad una profondità di -100 μm a 4 settimane. Nell'internodo, sono stati osservati segnali fluorescenti a una profondità di -60 μm senza il trattamento CS (Figura 3B). La profondità osservabile era di -90 μm dopo 1-3 settimane di trattamento con CS e -100 μm dopo 4 settimane, ma l'autofluorescenza nel citoplasma era evidente a una profondità di -20 μm dopo 1 settimana. L'autofluorescenza non era più visibile dopo 2 settimane. Nelle foglie, sono stati osservati segnali fluorescenti a una profondità di -90 μm senza il trattamento CS (Figura 3C). I segnali fluorescenti sono stati osservati più in profondità nelle foglie che nei nodi e negli internodi. Tuttavia, i segnali fluorescenti erano più deboli. I segnali fluorescenti sono diventati chiari dopo 1 settimana di trattamento CS e sono stati osservati a una profondità di -120 μm. Dopo 2 settimane e 3 settimane, sono stati osservati segnali fluorescenti a una profondità di -110 μm e, dopo 4 settimane, a una profondità di -120 μm.
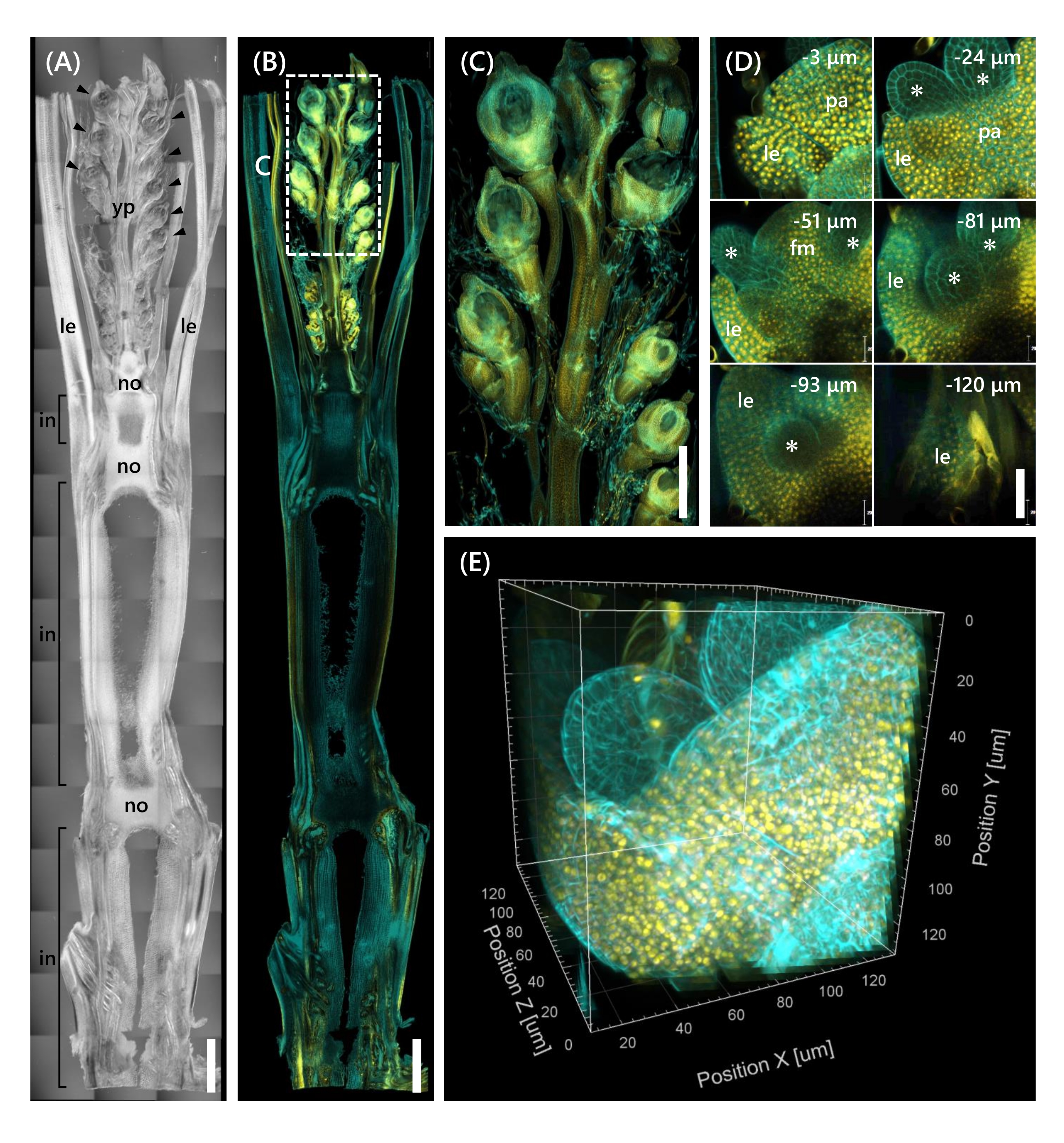
Infine, è stata osservata l'espressione del fattore di trascrizione OsMADS1511 fuso con mOrange 12, che regola il passaggio di SAM dalla fase vegetativa a quella riproduttiva13. I campioni di tessuto di Nip al 10 LS con OsMADS15-mOrange sono stati fissati, tagliati a 130 μm di spessore e trattati con il CS per 2 settimane. La Figura 4 mostra che la soluzione di clearing potrebbe essere utilizzata simultaneamente per osservare le proteine fluorescenti (OsMADS15-mOrange) dell'espressione genica naturale e della colorazione fluorescente (calcofluor white). È stato osservato l'intero fiore a profondità da 0 a -130 μm e l'immagine 3D è stata ricostruita da 43 immagini z-stack a intervalli di 3 μm (Figura 4E).

Figura 1: Posizione di campionamento e lavorazione. (A) Nip seedling a 8 LS nella condizione di giornata corta. (B) Vista ingrandita della base. (C) Un campione ritagliato dalla SAM/giovane pannocchia alla base. (D) Una sezione trasversale ovale da C (riquadro). (E) Rasatura sottile di un lato del germoglio con una lama. (F) Superficie sottilmente rasata. (G) Campione che mostra l'allungamento internodico. La sezione biancastra indicata dalle punte di freccia è nodi. Il lato superiore del nodo superiore ha la giovane pannocchia. Barre della scala: (A) 10 cm, (B-C, E-G) 1 cm, (D) 0,5 cm. Fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}

Figura 2: La pulizia dei tessuti dipende dallo spessore del campione e dal periodo di trattamento CS . (A) Campioni non rifilati con germogli parziali, compresa la giovane pannocchia. Spessore massimo: 4 mm. (B) Tutte le foglie staccate dai campioni per esporre la giovane pannocchia. Spessore massimo: 2,5 mm. YP: Pannocchia giovane, in: internodo; Le punte di freccia indicano il nodo. (C) Campioni che sono stati tagliati a mano a 1 mm di spessore. (D) Campioni rifilati con la microaffettatrice vibrante a 130 μm di spessore. Barra scala: 5 mm. Fare clic qui per visualizzare una versione ingrandita di questa figura.
{kind=link}

Figura 3: La profondità dell'osservazione della fluorescenza dipende dal tipo di tessuto e dal periodo di trattamento con CS. Imaging profondo di UBQpro::NGCN nel nodo (A), nell'internodo (B) e nella foglia (C). "Unisci" significa che le immagini sono state fuse con l'immagine DIC (Differential Interference Contrast) e tutte le immagini z-stack. Le immagini in cui non è stata osservata fluorescenza sono state contrassegnate come "n.d." (non rilevato). Le impostazioni della microscopia laser confocale sono le seguenti: laser: 488 nm/13%, obiettivo: 20x, foro stenopeico: 0,93 UA, media: 16, guadagno del rivelatore: 800, intervallo: 10 μm. Barre della scala: 50 μm. Fare clic qui per visualizzare una versione ingrandita di questa figura.
{kind=link}

Figura 4: Osservazione a fluorescenza profonda di OsMADS15-mOrange. (A) L'immagine DIC di Nip al 10 LS. YP: Pannocchia giovane, LE: foglia, No: Nodo, In: Internodo; Le punte di freccia indicano i fiori. (B) Immagine a fluorescenza di OsMADS15-mOrange. I punti gialli indicano le proteine fluorescenti di OsMADS15-mOrange nel nucleo. La parete cellulare era macchiata di ciano con la soluzione bianca calcofluor. (C) Vista ingrandita della giovane pannocchia. (D) Imaging profondo del fiore. PA: Palea, LE: Lemma, FM: meristema floreale; Gli asterischi indicano lo stame primordia. Il microscopio laser confocale è stato impostato come segue: OsMADS15-mOrange; laser: 555 nm/13%, obiettivo: 20x, foro stenopeico: 0,96 UA, media: 16, guadagno rivelatore: 800, intervallo: 3 μm. Calcofluor bianco; laser: 405 nm/3%, obiettivo: 20x, foro stenopeico: 0,93 UA, media: 16, guadagno del rivelatore: 470, intervallo: 3 μm. (E) L'immagine 3D della figura (D) costruita dalle immagini z-stack. Barre della scala: (A-B) 1 mm, (C) 500 μm, (D) 40 μm. Fare clic qui per visualizzare una versione ingrandita di questa figura.
{kind=link}
Discussione
Passaggi critici del protocollo
I passaggi critici in questo protocollo sono la fissazione e il taglio. I germogli di riso hanno tessuti duri, spessi o stratificati che limitano la penetrazione della soluzione fissativa. Per migliorare la permeabilità della soluzione fissativa, un lato del tessuto è stato rasato sottilmente al campionamento, come mostrato nella Figura 1E-F. Inoltre, i trattamenti sottovuoto sono stati ripetuti due volte utilizzando una pressione più elevata. Inoltre, i campioni sono stati fissati per una notte a 4 °C invece della consueta fissazione di 2 ore a 4 °C.
Il punto chiave nella fase di taglio è determinare lo spessore dei tessuti da preparare per osservare le proteine fluorescenti preservando la loro struttura interna dopo un breve periodo di trattamento CS. Come mostrato nella Figura 2C, i campioni spessi 1 mm, tagliati a mano il più sottili possibile, sono diventati trasparenti solo in un numero limitato di tessuti anche dopo 3 mesi di trattamento CS. Pertanto, la fase di taglio è essenziale per l'osservazione della fluorescenza profonda dei germogli di riso adulti. In questo studio, i campioni sono stati tagliati a uno spessore di 130 μm, come mostrato nella Figura 2D. Lo spessore di 130 μm ha permesso di liberare le foglie dopo 1 settimana di trattamento CS e l'intero campione dopo 2 settimane. In questo studio sono stati utilizzati germogli di riso adulti al 9-10 LS. I tessuti spessi ma più morbidi dei germogli di riso più giovani possono essere eliminati più velocemente con il trattamento CS. Lo spessore dei campioni e la durata del trattamento CS devono essere regolati in base al tipo di tessuto, alle condizioni e allo spessore della struttura 3D da osservare.
Modifiche e metodi di risoluzione dei problemi
Il CS precipitava facilmente a basse temperature. Il CS precipitato non può preservare le proteine fluorescenti; Pertanto, è necessario prestare attenzione quando si conservano i campioni alla temperatura corretta. Inoltre, sia il CS che la soluzione fissativa non hanno alcun effetto antisettico; Pertanto, le proteine fluorescenti saranno degradate se contaminate. Il riso coltivato nel suolo è soggetto a crescita fungina; Pertanto, il campionamento e la manipolazione dei campioni devono essere eseguiti con cura per evitare la contaminazione.
I coloranti fluorescenti in eccesso nel tampone possono emettere fluorescenza di fondo e interferire con le osservazioni microscopiche. Ad esempio, in precedenza veniva utilizzata una soluzione bianca di calcofluor contenente colorante blu Evans. Dopo la colorazione per 1 ora e il lavaggio per 1 ora, le proteine fluorescenti di OsMADS15-mOrange sono state osservate utilizzando un laser a 555 nm. Tuttavia, le proteine fluorescenti non potevano essere osservate a causa della fluorescenza di fondo derivata dal colorante blu di Evans. Questa fluorescenza di fondo è stata quasi eliminata lavando i campioni per 2 ore. Inoltre, le proteine fluorescenti erano più chiare se i campioni venivano lasciati durante la notte. Pertanto, in questo studio è stata utilizzata una soluzione bianca di calcofluor puro. La fluorescenza di fondo derivata dal colorante fluorescente deve essere controllata utilizzando diverse lunghezze d'onda laser prima delle osservazioni.
Limitazioni del metodo
Come mostrato nella Figura 3, sono state osservate proteine fluorescenti profonde nei campioni spessi 130 μm dopo 2 settimane di trattamento con CS. Ciò è coerente con i risultati mostrati nella Figura 2D, dove il campione spesso 130 μm è diventato trasparente dopo 2 settimane di trattamento con CS. Tuttavia, come mostrato nella Figura 3A, l'autofluorescenza del citoplasma era ancora evidente nei linfonodi dopo 2 settimane ed è stata completamente rimossa solo dopo 4 settimane di trattamento con CS. I nodi hanno un'alta densità di celle e, pertanto, richiedono un tempo più lungo per rimuovere i materiali auto-fluorescenti.
Come mostrato nella Figura 3C, sono state osservate proteine fluorescenti profonde nelle foglie senza trattamento CS, ma la luminosità era più debole di quella nei nodi e negli internodi alla stessa profondità di 20 μm. Dopo 1 settimana di trattamento con CS, le proteine fluorescenti erano più luminose. La clorofilla è abbondante nelle foglie e assorbe 488 nm di luce di eccitazione. Hanno anche autofluorescenza arancione / rossa, che può interferire con l'osservazione di proteine fluorescenti utilizzando un laser a 555 nm. Dopo 1 settimana di trattamento CS, la clorofilla e altri materiali auto-fluorescenti sono stati rimossi, ottenendo immagini ad alto rapporto segnale-rumore.
Le profondità che potevano essere osservate nei tessuti dopo 2 settimane e 4 settimane di trattamento con CS non erano significativamente diverse, sebbene le proteine fluorescenti apparissero più deboli dopo 4 settimane (Figura 3). Normalmente, la luminosità delle proteine fluorescenti e l'autofluorescenza si indeboliscono con il tempo, con un conseguente rapporto segnale-rumore più elevato. Pertanto, le proteine fluorescenti possono essere osservate più chiaramente regolando le condizioni microscopiche e l'elaborazione delle immagini. Sulla base di questi risultati, si è concluso che 2 settimane di trattamento con CS potrebbero facilitare l'osservazione delle proteine fluorescenti profonde, date le condizioni del nostro campione. Tuttavia, sono necessarie 4 settimane per osservare immagini più chiare che escludono completamente i materiali auto-fluorescenti.
Le strutture con forte autofluorescenza, come i fasci vascolari e le cellule multi-braccio, non possono essere eliminate nel CS. Per osservare queste strutture senza autofluorescenza, è necessario utilizzare un metodo di time-gating12 o ottenere immagini mediante spettroscopia dello spettro di fluorescenza. Un microscopio a due fotoni può essere più adatto per osservare tessuti più profondi se si osservano tessuti più spessi.
Significato del metodo rispetto ai metodi esistenti e alternativi
Generalmente, le strutture interne delle piante di riso sono state osservate utilizzando il sezionamento criostato o vibratomo. Un criostato è adatto per preparare sezioni sottili, che consentono un'osservazione più facile, ma la preparazione dei campioni e il funzionamento dell'apparecchiatura richiedono molto tempo. Anche ricostruire la struttura 3D originale da sezioni sottili è difficile. La vibratoma è relativamente facile da usare e adatta alla produzione di sezioni spesse. Tuttavia, sezioni spesse di tessuti bersaglio consentono solo osservazioni della superficie tagliata e non tessuti profondi che la luce non può raggiungere. Per questi motivi, nessuno dei due metodi è adatto per osservazioni di fluorescenza profonda.
Questo studio ha affrontato le sfide nell'osservazione della fluorescenza profonda nei germogli di riso, come la limitata penetrazione tissutale del CS e la scarsa risoluzione dell'oggetto al microscopio confocale, combinando i metodi esistenti. Come mostrato nella Figura 4, abbiamo osservato proteine fluorescenti (OsMADS15-mOrange) espresse nei tessuti profondi dei germogli di riso adulti dalla giovane pannocchia alla base. La Figura 4D si concentra sul fiore e mostra proteine fluorescenti profonde a intervalli di 3 μm. Tessuti di profondità superiore a -130 μm sono stati osservati dopo 2 settimane di trattamento con CS, ma solo i tessuti entro -27 μm di profondità sono stati osservati (dati non mostrati) nel fiore alla stessa dimensione e stadio di crescita senza trattamento CS. L'attuale protocollo migliorato ha permesso l'osservazione non solo della sovraespressione dei geni, ma anche dell'espressione genica naturale nei tessuti profondi dei germogli di riso adulti.
Importanza e potenziali applicazioni del metodo in specifiche aree di ricerca
Questo protocollo, che ottimizza l'osservazione della fluorescenza profonda dei germogli di riso adulti, consente l'eliminazione efficiente di tessuti duri, spessi o stratificati tagliando i tessuti non necessari e aumentando la permeabilità del CS. Inoltre, lo spessore dei campioni per l'analisi è stato ottimizzato per consentire l'osservazione continua e strutturale della fluorescenza profonda utilizzando un microscopio laser confocale, che normalmente non può risolvere tessuti spessi o opachi.
È difficile confrontare campioni di riso in diversi stadi di crescita perché le proteine fluorescenti si degradano nel tempo nelle soluzioni fissative e PBS. Tuttavia, le proteine fluorescenti nel CS possono essere conservate per più di 5 mesi1. La lunga durata di conservazione del CS è un grande vantaggio per l'osservazione della fluorescenza profonda nel riso.
Recentemente, sono state sviluppate molte tecnologie di pulizia, che consentono di osservare i tessuti profondi in 3D preservando le loro strutture interne. Queste tecnologie hanno continuato ad evolversi e sono state sviluppate nuove soluzioni di compensazione. Un buon esempio è iTOMEI14, che consente un'efficiente rimozione della clorofilla e un rilevamento della fluorescenza più brillante. Un altro esempio è ClearSeeAlpha15, che impedisce l'imbrunimento dei tessuti durante il trattamento di pulizia e li fa apparire trasparenti. La combinazione di queste soluzioni di compensazione con il metodo attuale può consentire una compensazione più efficiente ed efficace.
Si prevede che il metodo attuale aiuterà a ottenere nuove conoscenze attraverso l'imaging approfondito non solo del riso ma anche di altre piante.
Divulgazioni
Gli autori non hanno conflitti di interesse da dichiarare.
Riconoscimenti
Ringraziamo il Dr. R. Terada, il Dr. Z. Shimatani e il Dr. H. Tsuji per averci fornito i semi OsMADS15-mOrange; Dr. D. Kurihara per averci fornito la costruzione NGCN; e il Dr. R. Shim per aver modificato il nostro manoscritto. Questo lavoro è stato finanziato da JSPS KAKENHI (numero di sovvenzione JP20H05912, 20H05778, 20H05779) e dal programma SATREPS (no. JPMJSA1706) del GVC e JICA.
Materiali
| Name | Company | Catalog Number | Comments |
| 1.5 mL microcentrifuge tube | BIO-BIK | ST-0150F | |
| 12-multiwell plate | Corning | 353043 | |
| 50 mL conical tube | Corning | 352070 | |
| Calcofluor white solution | Sigma-Aldrich | 910090 | |
| ClearSee | FUJIFILM Wako Pure Chemical | 031-25151 | This can be made or purchased. |
| Confocal laser microscope | Carl Zeiss | LSM700 | |
| Desiccator | SANPLATEC | Custom made of acrylic. 30 cm (L), 30 cm (W), 14.5 cm (H) | |
| Glass coverslip (18 × 18 No.1) | MATSUNAMI | C018181 | |
| HEPES | FUJIFILM Wako Pure Chemical | 342-01375 | |
| Microscope slide (76 × 26) | MATSUNAMI | S2441 | |
| Paraffin film | Bemis | PM-996 | |
| Paraformaldehyde | FUJIFILM Wako Pure Chemical | 162-16065 | |
| Sodium deoxycholate | Tokyo Chemical Industry | C0316 | |
| Sucrose | FUJIFILM Wako Pure Chemical | 190-00013 | |
| UBQpro::NLS-sGFP-nClover3-mNeonGreen (UBQpro::NGCN) | provided by Dr. Kurihara | ||
| Urea | FUJIFILM Wako Pure Chemical | 211-01213 | |
| Vacuum pump | AS One | AS-01 | |
| Vibrating micro-slicer | DOSAKA | DTK-3000 | |
| Xylitol | FUJIFILM Wako Pure Chemical | 248-00545 |
Riferimenti
- Kurihara, D., Mizuta, Y., Sato, Y., Higashiyama, T. ClearSee: A rapid optical clearing reagent for whole-plant fluorescence imaging. Development. 142 (23), 4168-4179 (2015).
- Hasegawa, J., et al. Three-dimensional imaging of plant organs using a simple and rapid transparency technique. Plant and Cell Physiology. 57 (3), 462-472 (2016).
- Musielak, T. J., Slane, D., Liebig, C., Bayer, M. A versatile optical clearing protocol for deep tissue imaging of fluorescent proteins in Arabidopsis thaliana. PLoS One. 11 (8), 0161107 (2016).
- Pandey, K. B., et al. Plant roots sense soil compaction through restricted ethylene diffusion. Science. 371 (6526), 276-280 (2021).
- Ejaza, M., Bencivenga, S., Tavaresa, R., Busha, M., Sablowski, R. ARABIDOPSIS THALIANA HOMEOBOX GENE 1 controls plant architecture by locally restricting environmental responses. Proceedings of the National Academy of Sciences of the United States of America. 118 (17), (2021).
- Smetana, O., et al. High levels of auxin signalling define the stem-cell organizer of the vascular cambium. Nature. 565 (7740), 485-489 (2019).
- Chu, T. T. H., et al. Sub-cellular markers highlight intracellular dynamics of membrane proteins in response to abiotic treatments in rice. Rice. 11 (1), 23 (2018).
- Nagaki, K., Yamaji, N., Murata, M. ePro-ClearSee: A simple immunohistochemical method that does not require sectioning of plant samples. Scientific Reports. 7, 42203 (2017).
- Sato, M., Akashi, H., Sakamoto, Y., Matsunaga, S., Tsuji, H. Whole-tissue three-dimensional imaging of rice at single-cell resolution. International Journal of Molecular Sciences. 23 (1), 40 (2022).
- Matsuo, T., Hoshikawa, K. Science of the Rice Plant: Morphology. Food and Agriculture Policy Research Center. , (1993).
- Kobayashi, K., et al. Inflorescence meristem identity in rice is specified by overlapping functions of three AP1/FUL-Like MADS box genes and PAP2, a SEPALLATA MADS box gene. Plant Cell. 24 (5), 1848-1859 (2012).
- Tamaki, S., et al. FT-like proteins induce transposon silencing in the shoot apex during floral induction in rice. Proceedings of the National Academy of Sciences of the United States of America. 112 (8), 901-910 (2015).
- Kodama, Y. Time gating of chloroplast autofluorescence allows clearer fluorescence imaging In Planta. PLoS One. 11 (3), 0152484 (2016).
- Sakamoto, Y., et al. Improved clearing method contributes to deep imaging of plant organs. Communications Biology. 5 (1), 12 (2022).
- Kurihara, D., Mizuta, Y., Nagahara, S., Higashiyama, T. ClearSeeAlpha: Advanced optical clearing for whole-plant imaging. Plant and Cell Physiology. 62 (8), 1302-1310 (2021).
Ristampe e Autorizzazioni
Richiedi autorizzazione per utilizzare il testo o le figure di questo articolo JoVE
Richiedi AutorizzazioneEsplora altri articoli
This article has been published
Video Coming Soon
Personale delle biblioteche
Copyright © 2025 MyJoVE Corporation. Tutti i diritti riservati