
Method Article
جيل من الكروماتين على نطاق الجينوم تكيف التقاط المكتبات من الأجنة المورفولوجية محكم نظموا في وقت مبكر
In This Article
Summary
يصف هذا العمل بروتوكول لتوليد عالية الدقة في الموقع مرحبا-C مكتبات من أحكام نظموا قبل جاستروليشن melanogaster المورفولوجية الأجنة.
Abstract
ويقدم أفكاراً قيمة حول آليات تنظيم الجينات التحقيق بنية ثلاثية الأبعاد من الكروماتين. هنا، يمكننا وصف بروتوكول لأداء في الكروماتين تكيف التقاط تقنية في الموقع مرحبا-ج على نظم السكان الجنين melanogaster المورفولوجية . والنتيجة هي مكتبة تسلسل الذي يسمح لرسم خرائط لجميع الكروماتين التفاعلات التي تحدث في النواة في تجربة واحدة. ويتم فرز الأجنة يدوياً باستخدام مجهر فلوري استريو وخط الطيران المعدلة وراثيا يحتوي على علامة نووية. يمكن الحصول عليها باستخدام هذا الأسلوب، والسكان الجنين من كل دورة من دورات شعبة النووية، ومع وضع دورة الخلية المحددة، مع درجة نقاء عالية جداً. البروتوكول يمكن تكييفها لفرز الأجنة كبار السن تتجاوز gastrulation أيضا. فرز الأجنة تستخدم كمدخلات في الموقع مرحبا-جيم. جميع التجارب، بما في ذلك تسلسل إعداد مكتبة، يمكن أن تكتمل في غضون خمسة أيام. البروتوكول متطلبات المدخلات المنخفضة ويعمل موثوق بها باستخدام أجنة المرحلة بلاستوديرم 20 كمواد الإدخال. والنتيجة النهائية مكتبة تسلسل لتسلسل الجيل القادم. بعد التسلسل، يمكن معالجة البيانات إلى خرائط التفاعل الكروماتين على نطاق الجينوم التي يمكن تحليلها باستخدام مجموعة واسعة من الأدوات المتاحة للحصول على معلومات حول اقتران توبولوجيكالي بنية المجال (صبي) والحلقات الكروماتين الكروماتين المقصورات خلال التنمية المورفولوجية .
Introduction
التقاط تكيف الكروماتين (3 ج) برز كوسيلة استثنائية مفيدة لدراسة طوبولوجيا الكروماتين في نواة1. يسمح الخيار ج 3 ج مرحبا قياس ترددات الاتصال كل الكروماتين التفاعلات التي تحدث في النواة في تجربة واحدة2. تطبيق مرحبا-ج وقد لعبت دوراً هاما في اكتشاف وتوصيف العديد من المبادئ الأساسية للمنظمة الكروماتين، مثل الأمراض والمقصورات والحلقات3،،من45.
الدراسات المعمارية الكروماتين في سياق التحولات الإنمائية وتمايز خلية تستخدم بشكل متزايد لكشف آليات تنظيم الجينات أثناء هذه العمليات6،،من78، 9. واحد من الكائنات نموذج من اهتمام كبير هو melanogaster المورفولوجية، وتتسم أيضا الذين التنمية والجينوم. ومع ذلك، إجراء دراسات قليلة أن التحقيق في العمارة الكروماتين في المورفولوجية خارج زراعة الأنسجة في المختبر كانت إعدادات10،11. في الأجنة كانت ح 16 – 18 وظيفة الإخصاب، تادس والمقصورات يذكرنا بهياكل مماثلة في الثدييات حددت10، الذي يثير مسألة الدور الذي كانوا يلعبون في تنظيم الجينات خلال الجنين المورفولوجية التنمية. لا سيما في المراحل الأولى من التنمية، قبل جاستروليشن، مثل هذه الدراسات صعبة من الناحية التقنية. قبل جاستروليشن، تخضع الأجنة المورفولوجية 13 الانقسامات النووية متزامن أن المضي قدما بوتيرة سريعة جداً من 8 – 60 دقيقة لكل دورة12،13. وبالإضافة إلى ذلك، تجعل انعدام الميزات المرئية التمييز بين المراحل المختلفة من الصعب الحصول على الجنين نظموا أحكام المواد بكميات كافية.
من أجل وضع بروتوكول يسمح لدراسة الهندسة المعمارية الكروماتين في الإسراع بتطوير المورفولوجية في قرار الدورة النووية، نحن الجمع بين اثنين من التقنيات القائمة: في الموقع مرحبا جيم، الذي يتيح توليد عالية الدقة كاملة الاتصال خرائط الجينوم5، والتدريج الجنين باستخدام خط المورفولوجية المعدلة وراثيا معربا عن13،اجفب-منها التحوير14. هذا التحوير يموضع إلى النواة أثناء الطور البيني وتسارع شرط في جميع أنحاء بلاستوديرم المخلوي أثناء الانقسام. باستخدام هذه الخاصية، من الممكن بسهولة التمييز بين المراحل المختلفة بكثافتها النووية والأجنة الانقسامية التي تشتت إشارة التجارة والنقل.
معا، هذه التقنيات تمكن دراسة هيكل ثلاثي الأبعاد من الكروماتين بدقة عالية من عدد قليل من 20 المورفولوجية الأجنة. ويشمل هذا البروتوكول الإرشادات الخاصة بتجميع وفرز الأجنة المورفولوجية للحصول على سكان أجنة من دورة شعبة نووية واحدة. فهو يصف أيضا كيف تستخدم الأجنة التي تم الحصول عليها لتنفيذ في الموقع مرحبا--جيم والنتيجة النهائية مكتبة النوكليوتيدات مناسبة للتسلسل في آلات تسلسل الجيل القادم. ثم يمكن معالجة ما يلي التسلسل الناتج إلى خرائط التفاعل الكروماتين مفصلة تغطي كامل الجينوم المورفولوجية .
Protocol
1-جمع الجنين المورفولوجية
ملاحظة: يمكن أن يؤديها لجمع ما يعادل جنين كما هو موضح في منشور سابق15.
- نقل الذباب منها اجفب الشباب (< 1 أسبوع من العمر) في أقفاص جمع البيض مع ييستيد جمع اللوحات16 (الإيثانول 1% وحمض الخليك 1% واجار 4%).
- نقل مجموعة أقفاص إلى حاضنة تعيين عند 25 درجة مئوية. حضانة لمدة 1 – 2 أيام قبل جمع البيض يحسن إنتاج البيض إلى حد كبير. تغيير لوحات جمع مرتين في يوم.
- إزالة لوحات تحتوي على الأجنة من الدائرة جمع في فترات 30 – 60 دقيقة. أصغر فترات يؤدي الأجنة أقل، لكن توزيع أشد صرامة من مراحل النمو. جمع من أقفاص متعددة في نفس الوقت حيث أن المثالية > 200 بيضة وضعت كل 30-60 دقيقة.
- تخزين لوحات عند 25 درجة مئوية يبلغ العمر المطلوب الأجنة. لمرحلة بلاستوديرم احتضان الأجنة (دورة النووي 14)، لما يقرب من 2 ح.
- بعد 2 ساعة حضانة، إضافة ماء الصنبور من زجاجة بخ لوحة جمع حتى تغطي كامل السطح بالماء. تعليق الأجنة والخميرة باستخدام فرشاة ناعمة.
- صب حراكه الأجنة من لوحة جمع سلة جمع جنين (مصافي خلية تجارية مع 100 من حجم المسام ميكرومتر أو سلال محلية الصنع17 تعمل بشكل جيد)، مضيفاً إضافية حنفية الماء من زجاجة بخ، إذا لزم الأمر. وفي هذه المرحلة، الجمع بين أجنة من كل الألواح التي تم جمعها في نفس الوقت. وتمثل العينة المجمعة دفعة واحدة.
- تغسل الأجنة جيدا قبل الشطف سلة بماء الصنبور من زجاجة بخ ل 30 ق حتى جرفت جميع الخميرة بقايا.
- ديتشوريوناتي الأجنة بوضع سلة جمع إلى حل 2.5% هيبوكلوريت الصوديوم في الماء. الانفعالات الخفيفة التي يحوم يسهل إزالة المشيمة. تستمر حتى الأجنة مسعور بما فيه الكفاية بحيث أنها تطفو على سطح الحل عندما رفعت إلى السلة وأغرقت مرة أخرى، والتي ينبغي أن ~1.75–2 دقيقة.
تنبيه: تحت كلوريت الصوديوم التآكل. ارتداء معدات الوقاية الشخصية المناسبة. الحلول التي تحتوي على < تحت كلوريت الصوديوم 10% يمكن التخلص منها عادة في الحوض، وتأكد من أن التحقق من النظام الأساسي للمعهد المضيف. - إزالة السلة من الحل واشطف بعناية بماء الصنبور من زجاجة بخ حتى رائحة التبييض لم يعد ملحوظ.
2-الجنين التثبيت
ملاحظة: شروط التثبيت الأمثل، أساسا على تركيز المنظفات، والفورمالديهايد ومدة التثبيت، تحتاج إلى أن تكون تجريبية مصممة لتناسب مرحلة الأجنة. لمراحل حول بلاستوديرم المخلوي، تركيز 0.5% Triton X-100 و 1.8 في المائة فورمالدهايد نهائية في المرحلة مائي يعمل بشكل جيد. لمراحل لاحقة إلى ما بعد مرحلة الجنين 9، كذلك الاستفادة المثلى من هذه المعايير قد تكون ضرورية. ينبغي أن تتضمن جميع الحلول المستخدمة أثناء التثبيت والفرز مثبطات البروتياز.
- عكس سلة جمع ووضعه على أنبوب الطرد المركزي مخروطية 15 مل. مسح الأجنة من السلة في أنبوب باستخدام ماصة باستور الاستغناء عن برنامج تلفزيوني-T (برنامج تلفزيوني، 0.5% Triton X-100).
- واسمحوا الأجنة تستقر عند القاع وضبط الحجم الإجمالي إلى 2 مل مع برنامج تلفزيوني-ت.
- إضافة 6 مل هيبتان و 100 ميليلتر من 37% فورمالدهيد في المياه.
تنبيه: هيبتان وفورمالدهايد سامة عند استنشاقها أو بعد الجلد الاتصال. ارتداء معدات الوقاية الشخصية المناسبة، والعمل في غطاء دخان. وقد النفايات المحتوية على هيبتان أو الفورمالديهايد التخلص منها بشكل منفصل وفقا للنظام الأساسي للمعهد المضيف. - بعد إضافة الفورمالدهايد، بدء تشغيل جهاز ضبط وقت 15 دقيقة وقوة يهز الأنبوب صعودا وهبوطاً لمدة 1 دقيقة باليد. سوف تجمع بين المرحلة المائية والعضوية بشكل شامبو-مثل الاتساق.
- تحرض على خلاط الدنمارك حتى 10 دقيقة بعد إضافة فورمالدهايد.
- الطرد المركزي في 500 x ز لمدة 1 دقيقة في درجة حرارة الغرفة لجمع الأجنة في الجزء السفلي من الأنبوب.
- نضح السائل مثل الشامبو كامل وتخلص منه، مع الحرص على عدم نضح أي الأجنة. الكميات الصغيرة المتبقية من المادة طافية مثل الشامبو لا تسبب مشاكل.
- 15 دقيقة بعد إضافة فورمالدهايد، ريسوسبيند الأجنة في 5 مل من برنامج تلفزيوني-T مع 125 مم جليكاين آرو والفورمالديهايد. مزيج من قوة اهتزاز صعودا وهبوطاً لمدة 1 دقيقة.
- الطرد المركزي في 500 x ز في درجة حرارة الغرفة لمدة 1 دقيقة ونضح المادة طافية.
- تغسل الأجنة التي ريسوسبيندينج عليها في 5 مل من برنامج تلفزيوني المثلج-ت. اسمحوا تسوية الأجنة وأسبيراتي كلها طافية.
- تكرار الغسيل في خطوة 2.10 مرتين أخريين.
- الحفاظ على الأجنة على الجليد حتى الفرز. عادة, أنها فكرة جيدة لجمع 3 – 4 دفعات من الأجنة يطير قبل الشروع في الفرز. ومع ذلك، يجب فرز الأجنة وفي نفس اليوم. التخزين الموسعة على الجليد أو في الثلاجة يؤدي إلى مورفولوجيا الجنين المتغيرة.
3-الجنين الفرز
ملاحظة: يمكن أن يتم الفرز على أي مجهر ستيريو الفلورسنت مزودة بعامل تصفية بروتينات فلورية خضراء في 60 – 80 X التكبير.
- استخدام ماصة ميليلتر 1,000، نقل دفعة أجنة حوالي 100 سفينة زجاجية صغيرة مناسبة للفرز، يفضل أن يكون من لون داكن، ووضعه على الجليد.
-
فرز الأجنة قبل النووي حالة الكثافة ودورة الخلية (الشكل 1) عن طريق دفع الأجنة المرغوب في كومة منفصلة باستخدام تلميح إبرة أو حقنه.
- قم بإزالة جميع الأجنة مع توزيع اجفب-منها (الشكل 1E) مشتتة، غير النووية. أيضا، يجب إزالة الأجنة التي تظهر جزئيا إشارة بروتينات فلورية خضراء غير نووية.
- للمساعدة في الفرز، قم بتجميع تشكيلة أجنة مرجع في الدورة النووية 12 و 13، و 14 في كل دفعة باستخدام الصور في الشكل 1 كدليل. استخدام هذه التشكيلة لمطابقة الأجنة لمرحلة مجهولة مع واحد من الأجنة مرجع بغية تحديد مرحلة نموها.
- للتحقق من مرحلة النمو للأجنة مرجع، قياس كثافة النووية بتصوير الجنين وإحصاء عدد الأنوية في السطح للجنين في منطقة 2,500 مكم2 باستخدام برامج التصوير التي توفر المعلومات عن بعد.
ملاحظة: العدد المتوقع من نواة لمنطقة 2,500 مكم2 هو نوى 12 إلى 16 في الدورة النووية 12، ونوى 20 إلى 30 في الدورة النووية 1313.
- متى يتم فصل جميع الأجنة في المرحلة المناسبة، التقاط صور للأجنة للتوثيق ومراقبة الجودة. إذا كان ستيريو المجهر ليست في حد ذاتها مزودة بوحدة نمطية كاميرا، يمكن استخدام أي مجهر ابيفلوريسسينسي مع عوامل تصفية التجارة والنقل.
- "الماصة؛" حتى الأجنة المرغوب باستخدام ماصة ميليلتر 1,000، نقل إلى أنابيب جديدة، ومكان على الجليد.
- تستمر حتى يتم فرز الأجنة ما يكفي للتجربة المزمعة. للأجنة الأكبر سنا من المرحلة 9، عموما الأجنة 20 تكفي لتجربة واحدة في الموقع مرحبا-ج. في 12 دورة النووي، الأجنة 80 نقطة انطلاق جيدة. في الدورات السابقة، ينبغي أن تضاعف تقريبا عدد الأجنة لكل دورة.
- تجربة تجمع وانقسام الأجنة في أنابيب 1.5 مل في مثل هذه طريقة أن أنبوب واحد يحتوي على أجنة كافية لواحد في الموقع مرحبا-ج. فمن المستحسن استخدام أنابيب مع خصائص ربط الحمض النووي منخفضة، حيث ستستخدم الأنبوب نفسه للبروتوكول بأكمله والامتزاز من الحمض النووي يمكن أن تؤدي إلى خسائر كبيرة في تركيزات منخفضة من الحمض النووي.
- تدور أنابيب بإيجاز في 100 x ز في درجة حرارة الغرفة وإزالة المادة طافية. ينبغي أن تكون الأجنة جافا قدر الإمكان للتجميد.
- فلاش تجميد الأجنة بغمر الأنابيب في النتروجين السائل ومخزن في-80 درجة مئوية.
4. في الموقع مرحبا-ج
- تحلل
- وضع أنابيب بأجنة مجمدة في الجليد.
- ريسوسبيند الأجنة في 500 ميليلتر من المخزن المؤقت تحلل المثلج (10 ملم تريس Cl pH 8.0، 10 مم كلوريد الصوديوم، 0.2% CA إيجيبال-630، ومثبطات البروتياز؛ المذابة في الماء). ثم انتظر 1 دقيقة ترك الأجنة تسوية في الجزء السفلي من الأنبوب.
- طحن مدقة الصغرى معدنية، قبل تبريده في الجليد، والتي تم تصميمها لتناسب محكم أنبوب ميكروسينتريفوجي 1.5 مل باستخدام أجنة.
- إلى تجنب التحريض الأجنة، وإدراج المدقة ببطء حتى أنها تمس الجزء السفلي من الأنبوب، دفع إلى الأسفل وطحن ثم عن طريق تناوب مدقة مرتين في كلا الاتجاهين.
- رفع المدقة قليلاً جداً، ودفع إلى الجزء السفلي من الأنبوب مرة أخرى، وكرر طحن.
- كرر 4.1.3.2 10 مرات، أو حتى يتم تفكيك الأجنة تماما. أن الحل ينبغي أن تكون متجانسة، وينبغي أن يظل لا القطع الكبيرة المتبقية من الأجنة.
- احتضان تعليق المتجانس على الجليد ل 15 دقيقة تدور في س 1,000 ز، 4 درجة مئوية لمدة 5 دقائق، وتجاهل المادة طافية.
- أغسل بيليه ريسوسبيندينج في 500 المخزن المؤقت لتحلل المثلج ميليلتر، بيبيتينج صعودا وهبوطاً.
- تدور مرة أخرى كما هو الحال في 4.1.4، وتجاهل المادة طافية.
- ريسوسبيند بيليه غسلها في 100 ميليلتر من 0.5% الصوديوم دوديسيل كبريتات (SDS)، بيبيتينج صعودا وهبوطاً. بيرميبيليزي الأنوية بحضانة لمدة 10 دقائق عند 65 درجة مئوية في كتلة تدفئة. إخماد الحزب الديمقراطي الصربي بإضافة 50 ميليلتر من الماء ميليلتر X-100 و 120 تريتون 10%. مزيج من خلال يسددها الأنبوب.
- احتضان في 37 درجة مئوية لمدة 15 دقيقة في كتلة الحرارة.
- هضم إنزيم التقييد
- إضافة 25 ميليلتر من 10 × إنزيم التقييد المخزن المؤقت و 20 يو من 5 U/ميليلتر مبوي. مزيج من خلال يسددها الأنبوب.
- هضم الحمض النووي بحضانة لمدة 90 دقيقة في 37 درجة مئوية في كتلة الحرارة تحت الانفعالات طفيف (750 دورة في الدقيقة).
- إضافة 20 آخر يو مبوي وتستمر الحضانة لمدة 90 دقيقة.
- إلغاء تنشيط الحرارة مبوي التي تفرخ في 62 درجة مئوية لمدة 20 دقيقة.
- التعبئة المتراكمة
ملاحظة: ملء المتراكمة مع داتب بيوتينيلاتيد يسمح التحديد الشظايا الواصلة محددة. البيوتين-داتب في تقاطعات ربط محمي من [ااكسونوكلس] نشاط من T4 دنا بوليميريز (قسم 4.6)، بينما هو كفاءة إزالة داتب البيوتين في نهايات كليلة أونليجاتيد. يثري المنسدلة استخدام الخرز المغلفة ستريبتافيدين في قسم 4.7 ولذلك تحديداً لشظايا من الحمض النووي الواصلة، تشيميريك.- إضافة 18 ميليلتر من 0.4 مم البيوتين-14-داتب وميليلتر 2.25 مزيج دكتب/دجتب/دتتب غير معدلة (3.3 ملم في كل) ميليلتر 8 5 U/ميليلتر دنا بوليميريز I كلينوو جزء.
- مزج بالتحريك الأنبوب واحتضان في 37 درجة مئوية لمدة 90 دقيقة في كتلة الحرارة.
- عملية ربط
- إضافة 657 ميليلتر من الماء، ميليلتر 120 10 x T4 الحمض النووي ليجاسى المخزن المؤقت، 100 ميليلتر من 10% Triton X-100، ميليلتر 6 من 20 ملغ/مل ألبومين المصل البقري (BSA)، ومزيج من قبل التحريك الأنبوب. أخيرا إضافة 5 ميليلتر U/ميليلتر 5 T4 الحمض النووي ليجاسى ومزيج بالتحريك الأنبوب.
- قم بتدوير الأنبوب بلطف (20 لفة في الدقيقة) في درجة حرارة الغرفة ح 2.
- إضافة الدفعة ثانية من 5 ميليلتر من 5 U/ميليلتر T4 الحمض النووي ليجاسى ومواصلة التناوب 2 ح المزيد.
- تدور أسفل النوى في س 2,500 ز لمدة 5 دقائق وتجاهل المادة طافية.
- استخراج الحمض النووي
- ريسوسبيند بيليه في 500 ميليلتر من المخزن المؤقت الاستخراج (50 مم تريس Cl pH 8.0، 50 مم كلوريد الصوديوم، 1 ملم حمض الإيثيلين (يدتا)، الحزب الديمقراطي الصربي 1%؛ والمذابة في الماء) وإضافة 20 ميليلتر من 20 ملغ/مل بروتيناز ميكس ك بالتحريك الأنبوب.
- هضم البروتين التي تفرخ في 55 درجة مئوية مدة 30 دقيقة، تهتز 1,000 لفة في الدقيقة.
- دي-التشعب، إضافة 130 ميليلتر من 5 م كلوريد الصوديوم واحتضان بين عشية وضحاها في 68 درجة مئوية، تهتز 1,000 لفة في الدقيقة.
- ماصة العينة في أنبوب 2 مل جديدة، تفضيلي مع خصائص ربط الحمض النووي منخفضة.
- إضافة مجلدات x 0.1 (63 ميليلتر) من الصوديوم م 3 خلات pH 5-2 و 2 ميليلتر من 15 ملغ/مل جليكوبلوي. مزيج جيد من قبل عكس. إضافة 1.6 x مجلدات (1,008 ميليلتر) من الإيثانول النقي المطلق ومزيج بعكس.
- احتضان في-80 درجة مئوية عن 15 دقيقة للطرد المركزي في 20,000 x ز عند 4 درجة مئوية لمدة 30 دقيقة على الأقل. بيليه الحمض النووي في كثير من الأحيان صغيرة جداً، ويكاد يكون غير مرئي، ويمكن رصدها فقط بسبب اللون الأزرق جليكوبلوي.
- إزالة المادة طافية بعناية فائقة، تتحرك غيض ماصة في الأنبوب على طول الجدار المقابل من حيث يقع بيليه الحمض النووي. يتم غالباً بسهولة إزالة القطرات الصغيرة المتبقية أثناء هذه الخطوة ويغسل التالية بدفعهم خارج الأنابيب استخدام تلميح P10 بدلاً من بيبيتينج بها.
- أغسل بيليه بإضافة 800 ميليلتر من الإيثانول 70%. مزيج من عكس والطرد المركزي في 20,000 x ز في درجة حرارة الغرفة للحد الأدنى 5 كرر هذا يغسل مرة واحدة على الأقل.
- إزالة آثار كل من الإيثانول وترك مكانة أنبوب مع فتح الغطاء لمدة تصل إلى 5 دقائق أيردري. مرة واحدة متبقية لا سائل، إضافة 50 ميليلتر من 10 ملم تريس Cl pH 8.0. مرارا وتكرارا "الماصة؛" أن الحل في المنطقة على جدار الأنبوبة حيث يقع بيليه جعل الحمض النووي.
- أضف 1 ميليلتر من 20 ملغ/مل رناسي A، مزيج من قبل التحريك الأنبوب، واحتضان في 37 درجة مئوية لمدة 15 دقيقة لهضم الحمض النووي الريبي. يمكن الآن المخزنة في الثلاجة بين عشية وضحاها العينة أو المجمدة في-20 درجة مئوية إلى أجل غير مسمى.
- أساس الاختيار تركيز الحمض النووي باستخدام صبغة فلورسنت المقايسة وفقا لإرشادات الشركة المصنعة. ينبغي أن يكون المبلغ الإجمالي للحمض النووي للعينة على الأقل 10 نغ، إلا القليل جداً من المواد متوفر للتضخيم والمكتبة تعقد سيكون على الأرجح منخفضة. وعندما يحدث ذلك، مبلغ ابتداء من المواد ربما ليس كافياً، أو انقطع المواد على طول الطريق، وربما خلال تحلل وهطول الأمطار.
- إزالة البيوتين وقص الحمض النووي
- إضافة معا ميليلتر 12 x 10 T4 دنا بوليميريز المخزن المؤقت 3 ميليلتر من داتب 1 مم، 3 ميليلتر من دجتب 1 مم و 46 ميليلتر من الماء. مزيج من خلال يسددها الأنبوب. إضافة 5 ميليلتر من 3 U/mL T4 دنا بوليميريز ومزج بالتحريك الأنبوب واحتضان في 20 درجة مئوية لمدة 30 دقيقة.
- إضافة 3 ميليلتر يدتا 0.5 M لوقف رد الفعل، واستخدام المياه لإحضار العينة لوحدة تخزين من حوالي 120 ميليلتر.
- قص الحمض النووي لحجم 200 – 400 بي بي باستخدام جهاز سونيكيشن وفقا لإرشادات الشركة المصنعة. استخدام سونيكاتور المذكورة في الجدول للمواد، البرنامج التالي المناسب: 2 دورات كل منها 50 s، واجب 10%، وكثافة 5، 200 دورات/الاندفاع.
- المنسدلة البيوتين
- "الماصة؛" 30 ميليلتر من 10 ملغ/مل streptavidin المغلفة المغناطيسي الخرز في أنبوب جديد وتفصل بينهما على الوقوف مغناطيسية، وتجاهل المادة طافية.
- ريسوسبيند الخرز في 1 × المخزن المؤقت ب ث (5 ملم تريس Cl الرقم الهيدروجيني 7.4، يدتا 0.5 مم، 1 م كلوريد الصوديوم؛ المذابة في الماء) + 0.1% X-100 تريتون ومزيج من فورتيكسينج. ضع الأنبوبة على الوقوف مغناطيسية والانتظار لمدة 1-5 دقائق حتى مفصولة الخرز، اعتماداً على وطرازه.
- نضح وتجاهل المادة طافية حين ينزلق طرف الماصة على طول الجدار المقابل من حيث توجد الخرز. ريسوسبيند الخرز في ميليلتر 120 2 × المخزن المؤقت ب ث (10 ملم تريس Cl الرقم الهيدروجيني 7.4، يدتا 1 مم، وكلوريد الصوديوم م 2). مزيج من فورتيكسينج.
- نقل الحمض النووي المنفصمة إلى أنبوب ربط الحمض النووي جديدة منخفضة، وخلط مع ميليلتر 120 تعليق حبة في المخزن المؤقت ب X و W 2 من فورتيكسينج. قم بتدوير الخرز مع عينات الحمض النووي 20 لفة في الدقيقة لمدة 15 دقيقة.
- فصل الخرز على الوقوف مغناطيسية وتجاهل المادة طافية.
- ريسوسبيند الخرز في 600 ميليلتر من 1 × ب & W + 0.1% X-100 تريتون، واحتضان في 55 درجة مئوية لمدة دقيقة 2، تهتز 1,000 لفة في الدقيقة. بعد الانفصال، وتجاهل المادة طافية. كرر هذا يغسل مرة واحدة.
- أغسل حبات مرة واحدة مع 600 ميليلتر من 10 ملم تريس Cl pH 8.0، وتجاهل المادة طافية بعد الانفصال.
- ريسوسبيند الخرز في 50 ميليلتر من 10 ملم تريس Cl pH 8.0.
5-تسلسل إعداد المكتبة
ملاحظة: تتم جميع الخطوات المكتبة باستخدام مكونات من إعداد مكتبة الحمض النووي تجاري كيت (انظر الجدول للمواد). ومع ذلك، يمكن الاستعاضة مجموعات بديلة أو الكواشف الأخرى. هطول الأمطار يميل إلى النموذج في عوامل إعداد المكتبة أثناء التخزين المجمد. ولذا من المهم التأكد من أن يحل جميع هطول الأمطار قبل استخدام الكواشف.
-
الإصلاح نهاية
- نقل تعليق حبة في 50 ميليلتر من 10 ملم تريس Cl pH 8.0 في أنبوب بكر جديدة.
- إضافة 3 ميليلتر من نهاية إنزيم الإعدادية خليط و 7 ميليلتر من نهاية الإعدادية رد فعل المخزن المؤقت. مزيج من بيبيتينج صعودا وهبوطاً.
- نقل الأنبوب إلى cycler حرارية وتشغيل البرنامج التالي: 20 درجة مئوية لمدة 30 دقيقة، 65 درجة مئوية و 30 دقيقة، وعقد في 4 درجات مئوية.
-
ربط محول
- إضافة 30 ميليلتر من مزيج الرئيسي ربط، 2.5 ميليلتر من 1.5 ميكرومتر "تسلسل محول" (مخفف إلى 1.5 ميكرومتر من الأسهم)، وميليلتر 1 "ربط محسن" إلى تعليق حبة. مزيج من بيبيتينج صعودا وهبوطاً.
- تبني في 20 درجة مئوية لمدة 15 دقيقة في cycler حرارية.
- إضافة 3 ميليلتر من الإنزيم المستخدم. مزيج من بيبيتينج صعودا وهبوطاً.
- احتضان في 37 درجة مئوية لمدة 15 دقيقة في cycler حرارية.
- فصل الخرز على الوقوف مغناطيسية وإزالة المادة طافية.
- ليغسل الخرز، ريسوسبيند الخرز في 100 ميليلتر من 1 × المخزن المؤقت ب ث + 0.1% X-100 تريتون. مزيج من فورتيكسينج، ونقل إلى أنبوب ميكروسينتريفوجي جديدة. فصل الخرز على الوقوف مغناطيسية وإزالة المادة طافية.
- كرر هذا يغسل مرة واحدة باستخدام 600 ميليلتر من نفس المخزن المؤقت.
- ريسوسبيند الخرز في 600 ميليلتر من 10 ملم تريس Cl pH 8.0، مزيج من فورتيكسينج، ونقل الخرز إلى أنبوب جديد.
- فصل الخرز على الوقوف مغناطيسية وتجاهل المادة طافية ريسوسبيند الخرز في 50 ميليلتر من 10 ملم تريس Cl pH 8.0.
-
[بكر] تضخيم
- إعداد أنابيب PCR اثنين وفي كل مجال، مزيج 25 ميليلتر من مزيج ماجستير بوليميريز و 1.5 ميليلتر من 10 ميكرون إلى الأمام (غير المفهرسة) بكر التمهيدي 1.5 ميليلتر من 10 ميكرون عكس (مفهرس) بكر التمهيدي.
ملاحظة: إلى الأمام (غير المفهرسة) بكر التمهيدي:
5-آتجاتاكجكجاككاككجاجاتكتاكاكتكتتكككتاكاكجاكجكتكتككجاتك * تي-3´.
عكس (مفهرس) بكر التمهيدي:
5-كاجكاجاجاكجكاتاكجاجاتنننننجتجاكتجاجتكاجاكجتجتجكتكتككجاتك * تي-3´. * يشير إلى سندات فوسفوروثيواتي و Ns في التمهيدي PCR المفهرسة. - في كل أنبوب، إضافة ميليلتر 22 تعليق حبة ومزيج من بيبيتينج صعودا وهبوطاً.
- تشغيل الاسترداد باستخدام البرنامج التالي: 98 درجة مئوية لمدة 1 دقيقة، (98 درجة مئوية لمدة 15 ثانية، 65 درجة مئوية مقابل 75 ق، تكثف 1.5 ° C/s) مرات 9-12، 65 درجة مئوية لمدة 5 دقائق، وعقد في 4 درجات مئوية.
ملاحظة: عدد دورات التضخيم قد يتحدد تجريبيا. ومع ذلك، وجدنا أن المكتبات التي تتطلب دورات أكثر من 12 بوجه عام من التعقيد منخفضة ولم تسفر عن جودة عالية خرائط مرحبا-ج. من ناحية أخرى، المكتبات التي تتطلب دورات أقل من 12 لم تتأثر سلبا بتضخيم دورات 12 بصورة كاملة. ولذلك، من الممكن لدورات 12 التضخيم الافتراضي. - تجميع ردود فعل بكر اثنين في أنبوب ميكروسينتريفوجي واحدة وفصل الخرز على الوقوف مغناطيسية، ونقل المادة طافية يحتوي على المكتبة لأنبوب جديد.
- إعداد أنابيب PCR اثنين وفي كل مجال، مزيج 25 ميليلتر من مزيج ماجستير بوليميريز و 1.5 ميليلتر من 10 ميكرون إلى الأمام (غير المفهرسة) بكر التمهيدي 1.5 ميليلتر من 10 ميكرون عكس (مفهرس) بكر التمهيدي.
-
حجم التحديد
- جلب أمبوري إكس بي تعليق حبة إلى درجة حرارة الغرفة ومزيج جيد بالهز.
- إعادة التخزين لتفاعل PCR المجمعة بالضبط 200 ميليلتر بالماء. خلال PCR والفصل المغناطيسي، يتم عادة فقدان بعض من وحدة التخزين الأصلية. تحقق من حجم بواسطة إعداد ماصة إلى 200 ميليلتر ونضح وحدة التخزين بأكملها من رد فعل. إذا يستنشق الهواء، الماء أكثر الاحتياجات المراد إضافتها. إذا تجاوز الحجم 200 ميليلتر، ضبط حجم الخرز إضافتها في الخطوات 5.4.3 و 5.4.6 نسبيا.
ملاحظة: وحدات التخزين في الأقواس صالحة إذا كان إجمالي حجم ردود الفعل PCR المجمعة بالضبط 200 ميليلتر. - إضافة 0.55 x أحجام (110 ميليلتر) تعليق حبة أمبوري إكس بي ومزيج من بيبيتينج صعودا وهبوطاً على الأقل 10 مرات.
- احتضان في درجة حرارة الغرفة لمدة 5 دقائق، الخرز منفصل على الوقوف مغناطيسية لمدة 5 دقائق.
- نقل المادة طافية إلى أنبوب جديد. تجاهل الأنبوب الذي يحتوي على الخرز. الخرز وقد ربط الحمض النووي > 700 شركة بريتيش بتروليوم، التي كبيرة جداً لتكون متسلسلة.
- إضافة إلى المادة طافية، 0.2 x وحدات التخزين (40 ميكروليتر، أسفر عن مجموعة المخزن المؤقت x Ampure 0.75 في العينة) تعليق حبة أمبوري إكس بي ومزيج من بيبيتينج صعودا وهبوطاً 10 مرات.
- احتضان في درجة حرارة الغرفة لمدة 5 دقائق، الخرز منفصل على الوقوف مغناطيسية لمدة 5 دقائق.
- تجاهل المادة طافية التي تحتوي على الحمض النووي < 200 شركة بريتيش بتروليوم، التي تشمل الإشعال الحرة، مبلمر، وشظايا صغيرة جداً تكون متسلسلة.
- اترك الأنبوب على الوقوف المغناطيسية. أغسل حبات، إضافة 700 ميليلتر من الإيثانول 80%، مع الحرص على عدم تعكير بيليه حبة، واحتضان لمدة 30 ثانية.
- تجاهل المادة طافية، ثم أخذ الأنبوب قبالة موقف المغناطيسية وريسوسبيند الخرز في 100 ميليلتر من 10 ملم تريس Cl pH 8.0. مزيج من بيبيتينج صعودا وهبوطاً 10 مرات، واحتضان في درجة حرارة الغرفة لمدة 1 دقيقة.
- إضافة مجلدات x 0.8 (80 ميليلتر) من تعليق حبة أمبوري إكس بي. مزيج من بيبيتينج صعودا وهبوطاً 10 مرات واحتضان في درجة حرارة الغرفة لمدة 5 دقائق. هذه الجولة الثانية من اختيار حجم الحد الأدنى يضمن أن المكتبة نهائية خالية تماما من الإشعال ومبلمر.
- فصل الخرز على الوقوف مغناطيسية لمدة 5 دقائق وتجاهل المادة طافية.
- تغسل بيليه حبة مرتين مع 700 ميليلتر من الإيثانول 80% لمدة 30 ثانية كل منهما، بينما تترك الأنبوبة على الوقوف المغناطيسي، كما ذكر أعلاه.
- مع الأنبوب لا يزال على الوقوف المغناطيسي، إزالة آثار كل من الإيثانول. ويساعد على دفع قطرات الإيثانول من الأنبوب باستخدام ماصة P10. واسمحوا الإيثانول المتبقية تتبخر لمدة أقصاها 5 دقائق.
- أخذ أنبوب قبالة موقف المغناطيسية وريسوسبيند الخرز في 50 ميليلتر من 10 ملم تريس Cl pH 8.0. مزيج من بيبيتينج صعودا وهبوطاً 10 مرات.
- احتضان في درجة حرارة الغرفة لمدة 5 دقائق، ثم الخرز منفصل على الوقوف مغناطيسية.
- نقل طافية على أنبوب جديد. هذا هو مكتبة مرحبا-C النهائي، مستعد لأن يكون كمياً ومتسلسلة على آلات تسلسل الجيل القادم، طبقاً لإرشادات الشركة المصنعة.
النتائج
فرز السكان الجنين في الدورة النووية 12 و 13 و 14 (الموافق 01:30، 01:45، و 02:10 ساعات ما بعد الإخصاب، على التوالي12) وتم الحصول على 3 – 4 ح وظيفة التسميد (hpf) وفقا للإجراءات المبينة في البروتوكول. بالتقاط صور لإشارة منها اجفب من كل دفعة الجنين تم فرزها، فمن الممكن لتوثيق الوضع المرحلة ودورة الخلية الدقيق لكل جنين واحد يستخدم في تجارب المتلقين للمعلومات. مثال على صور للأجنة من السكان تم فرزها مبينة في الشكل 1 باء-ه. إخراج البروتوكول مرحبا-C في الموقع مكتبة النوكليوتيدات جاهزة تكون متسلسلة على آلات تسلسل الجيل القادم. ولهذا الغرض، تركيز مكتبة نهائي من شمال البحر الأبيض المتوسط على الأقل 2-4 مطلوب عادة. استخدام المبالغ الموصى بها لإدخال المواد، وهذا التركيز موثوق تحقق (الجدول 1).
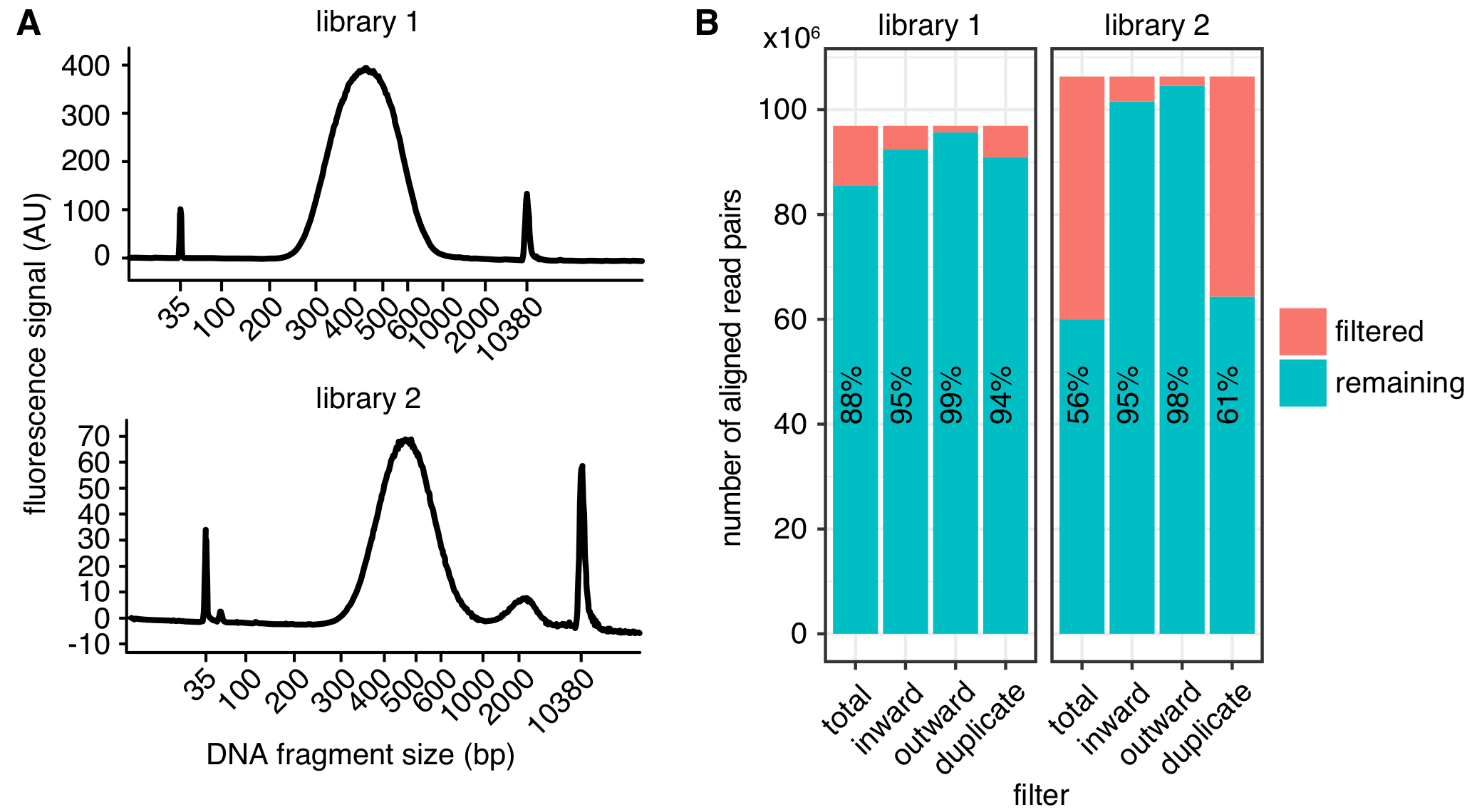
توزيع الحجم المتوقع من شظايا من الحمض النووي بعد تحديد حجم ما بين 300-600 شركة بريتيش بتروليوم، مع حد أقصى في حوالي 500 شركة بريتيش بتروليوم (الشكل 2A)، اعتماداً على القص الدقيق ومعايير اختيار حجم. للتسلسل، نوصي بما يلي نهاية الاقتران من طول bp 75 على الأقل لتقليل عدد الأجزاء تقييد مصادفة في الجينوم. يمكن الحصول على خرائط عالية الدقة بحجم بن 1 – 2 كيلو بايت من يقرأ 400 مليون. ونحن نوصي التسلسل replicates بيولوجية متعددة على عمق أقل من 150 مليون يقرأ كل، بدلاً من التسلسل تكرار واحد في عمق عالية جداً. وهذا يسمح تقييم الاختلاف البيولوجي ويؤدي إلى انخفاض عدد القراءات المهملة بسبب ازدواجية PCR. لتمثيل مرئي، يمكن دمجها في replicates. قبل الالتزام بالتسلسل عينة في عمق عالية، نوصي بتشغيل العينات باستخدام تسلسل الضحلة (بضعة ملايين ما يلي كل عينة) لتحديد معلمات نوعية المكتبة الأساسية كما هو الحال في الشكل 2.
تحليل البيانات مرحبا-ج يتطلب خبرة كبيرة من الموارد والمعلوماتية الحيوية الحسابية. كنظرة الخام، يقرأ إقران يتم تعيينها بشكل مستقل للجينوم المرجعية، وأن التحالفات الناتجة يتم تصفيتها للجودة والتوجه، ثم يمكن إنشاء مصفوفة اتصالات على مستوى القرار أو جزء معين بن من تمت تصفيتها التحالفات. مصفوفة الاتصال هو الأساس لتحليل جميع المتلقين للمعلومات كذلك استكشاف تادس والحلقات والأجزاء الأخرى. تتوفر العديد من خطوط الأنابيب المعلوماتية الحيوية للتحليلات الأولية لقراءة التسلسل، تمكن معالجة الخام ما يلي إلى مصفوفات الاتصال دون كثير المعلوماتية المتخصصة المعرفة18،19، 2021،،،من2223. كيف يتم إجراء مزيد من التحليل يعتمد إلى حد كبير على مسألة البيولوجية الدقيقة قيد الدراسة وقد تتطلب خبرة كبيرة في البرمجة والبرمجة في البحث والتطوير أو بايثون. عدة أدوات وخوارزميات لاستدعاء تادس غير متاح5،24،25،26،،من2728، فضلا عن برامج لتحليل و استكشاف البيانات C مرحبا في مستعرض ويب وتطبيقات سطح المكتب قائمة بذاتها29،30،،من3132.
وفور تجهيز، يمكن تحديد نوعية المكتبة استخدام مقاييس مختلفة (الشكل 2). أولاً، ينبغي أن يكون معدل التكرارات بكر، وهو عدد أزواج القراءة المتتابعة الناشئة عن الجزيء الأصلي نفسه، منخفضة قدر الإمكان الحد من مقدار ما يلي تسلسل مهدرة. ومع ذلك، وحتى المكتبات مع > يمكن معالجة الازدواجية بكر 40% إلى خرائط عالية الجودة الاتصال إذا كانت التكرارات يتم تصفيتها. ثانيا، القراءات التي تمت تصفيتها بسبب ميولهم، كما هو موضح في4، ينبغي دائماً يكون المعدل أقل من 10% أزواج القراءة المنحازة.
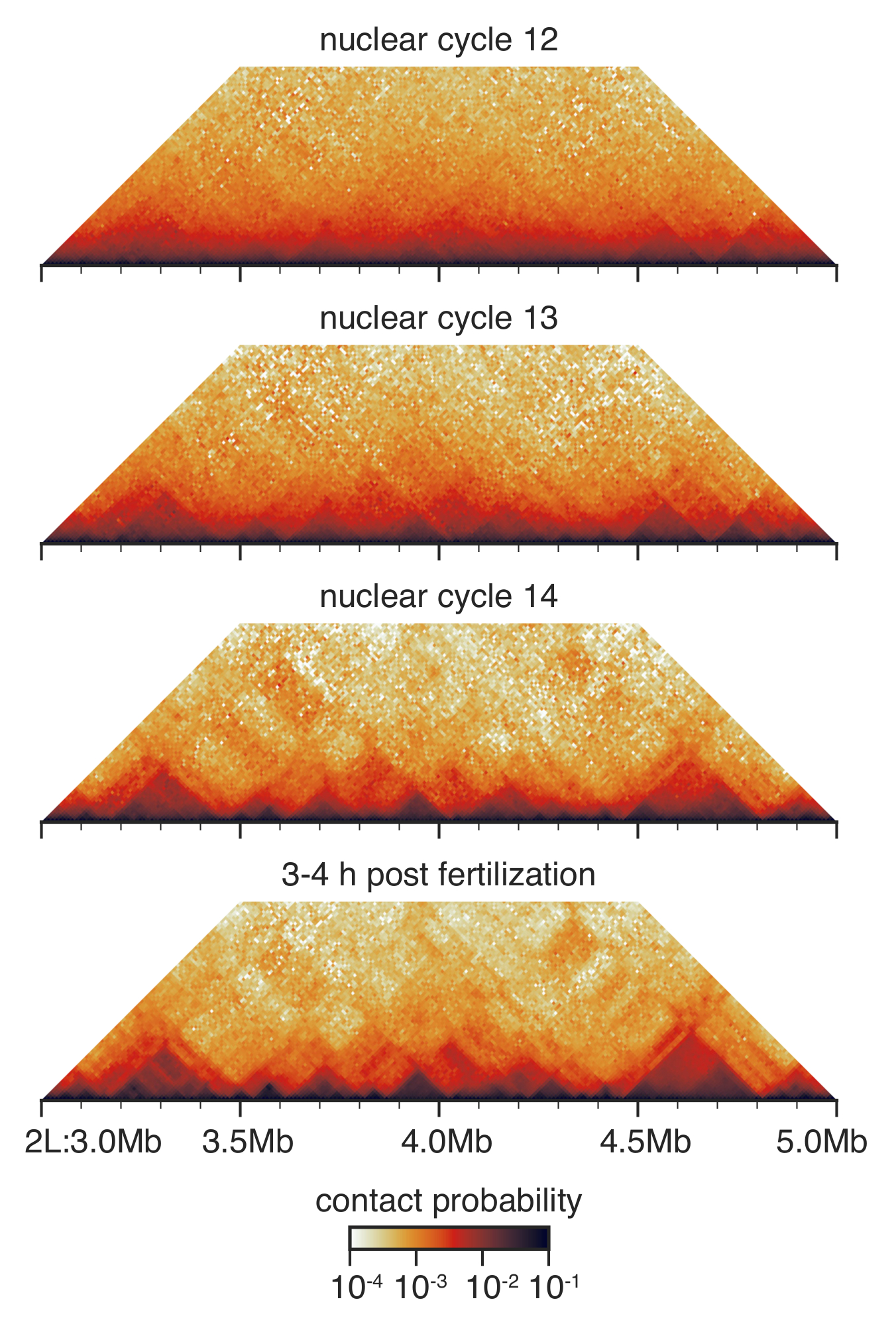
أثناء التطوير قبل جاسترولار من المورفولوجية بين دورة النووي 12 و 14، هو البنية النووية جذريا تشكيلها33 (الشكل 3). في الدورة النووية 12، يتم الكشف عن بعض الأمراض، والتوزيع العام للاتصالات سلس جداً دون العديد من الميزات ملموس. وهذا هو تغيرا في الدورة النووية 13 و 14، عندما تادس بارز على نحو متزايد، وتستنفد الاتصالات البعيدة المدى غير محدد.

رقم 1: صور الممثل للأجنة اجفب منها أثناء الفرز. (أ) اجفب-منها إشارة من سكان لم يتم فرزها بالأجنة بعد جمع 60 دقيقة وحضانة ح 2 في 25 درجة مئوية (ب ه) أمثلة على الأجنة من السكان تم فرزها في الدورة النووية 12 (ب)، النووي دورة 13 (ج)، (14 دورة النووي د)، ومن الأجنة يمر الانقسام متزامن (E). تغيير حجم أشرطة = 200 ميكرومتر. الرجاء انقر هنا لمشاهدة نسخة أكبر من هذا الرقم-
{kind=link}

الشكل 2: أمثلة في الموقع مرحبا-C مكتبة نوعية المقاييس. آثار بيواناليزير (A) يبين التوزيع للحمض النووي أحجام جزء من مكتبة مرحبا-ج ناجحة (مكتبة 1، الأعلى) ومن مكتبة الذي يعرض وه شظايا التي تكون كبيرة جداً لتسلسل (مكتبة 2، أسفل). مكتبة 2 كان التسلسل بنجاح، ولكن كميات أكبر من شظايا من الحمض النووي غير مرغوب فيها قد تؤدي إلى تسلسل تناقص الغلة. (ب) تصفية إحصاءات مكتبتين مرحبا-ج: عرض هو عدد أزواج القراءة المنحازة المستبعدة من مزيد من التحليل بسبب قراءة الاتجاه والمسافة (إلى الداخل وإلى الخارج)4 أو الازدواجية بكر (مكررة). في كل شريط، يتم رسم عدد القراءات تمرير عامل التصفية (المتبقية) والفشل (تصفية). بالإضافة إلى ذلك يتم عرض النسبة المئوية لتمرير عامل التصفية على ما يلي كنص. الرجاء انقر هنا لمشاهدة نسخة أكبر من هذا الرقم-
{kind=link}

الشكل 3: التفاعل ج خرائط من الأجنة نظموا. خرائط التفاعل ج إهمال في القرار 10 كيلو بايت ومتوازن كما هو موضح قبل33. سيظهر منطقة على الصبغي 2 ل. الرجاء انقر هنا لمشاهدة نسخة أكبر من هذا الرقم-
{kind=link}
| مكتبة | المرحلة | عدد الأجنة | كمية الحمض النووي قبل القص (الحرس الوطني) | دورات بكر | مكتبة النهائي تركيز (nM) |
| 1 | دورة النووي 12 | 71 | 46 | 12 | 28.2 |
| 2 | دورة النووي 12 | 46 | 40 | 12 | 22.2 |
| 3 | دورة النووي 12 | 60 | 13 | 13 | 12.3 |
| 4 | دورة النووي 13 | 36 | 39 | 12 | 22.2 |
| 5 | دورة النووي 13 | 35 | 10 | 12 | 5.0 |
| 6 | دورة النووي 13 | 48 | 18 | 12 | 8.7 |
| 7 | دورة النووي 14 | 33 | 30 | 12 | 39.8 |
| 8 | دورة النووي 14 | 24 | 36 | 12 | 20.4 |
| 9 | دورة النووي 14 | 14 | 8 | 12 | 4.2 |
| 10 | 3-4 هبف | 17 | 30 | 12 | 24.0 |
| 11 | 3-4 هبف | 18 | 42 | 11 | 19.1 |
| 12 | 3-4 هبف | 22 | 63 | 11 | 48.4 |
الجدول 1: قائمة بتسلسل الممثل مكتبة الإحصاءات. لكل مكتبة في القائمة، عدد الأجنة التي تم استخدامها لجيلها، مبلغ إجمالي الحمض النووي قبل المنسدلة البيوتين والقص تقاس كوبيت، دورات عدد PCR يستخدم للتضخيم، وتركيز المكتبة التسلسل النهائي بعد أن يتم الإشارة إلى تنقية واختيار حجم.
Discussion
البروتوكول المعروضة هنا فعالة جداً في توليد خرائط عالية الجودة للهندسة المعمارية الكروماتين في الأجنة المورفولوجية في وقت مبكر. مقارنة بروتوكول سابق34، يستخدم النهج الموصوفة هنا حديثة في الموقع مرحبا-ج الداخلي5، مما أسفر عن تجهيز أسرع ودقة أعلى، وأقل استخدام كاشف. الداخلي الإجمالي بما في ذلك البروتوكول مرحبا-C في الموقع المتوقع للعمل على مجموعة واسعة من المراحل والنظم التجريبية إلى جانب المورفولوجية. منذ البروتوكول بشرط إدخال منخفضة، لكن يمكن أيضا استخدامه في السكان خلية معزولة. في المورفولوجية، عند استخدام البروتوكول للأجنة خارج النطاق الموصوفة هنا، بعض المعلمات، لا سيما تثبيت المواد، قد تحتاج إلى تعديل. منذ وضع الأجنة الأكبر سنا بشرة شديدة كتيمة، زيادة تركيز فورمالدهايد وإطالة أمد التثبيت قد يكون مناسباً. لمجموعة من الأجنة في مراحل عدا دورة النووي 14، مرات حضانة الأجنة عند 25 درجة مئوية في الخطوة 1، 4 تحتاج إلى تعديلها على النحو التالي: الدورة النووية 12, 70 دقيقة؛ دورة النووي 13، 90 دقيقة؛ 3 – 4 هبف، 03:30 ح.
خلال الانقسامات الانقسام 13 (المرحلة 1-4)، الزوجي كثافة الأنوية تقريبا مع كل شعبة. يمكن بسهولة تحديد الأنوية الأسفار بروتينات فلورية خضراء مشرقة. أثناء الانقسام، اجفب منها غير موجود في النواة، والإشارة هو موزعة الجنين. هذه الميزة يجعل تحديد الأجنة التي تشهد تقسيما انشقاق متزامن ممكن. لدراسة تشكيل الكروماتين، هذه الأجنة الانقسامية ليست عادة مرغوباً فيه، منذ الانقسامية تنظيم الكروماتين يختلف جذريا عن المنظمة الطور البيني35. فمن الممكن التكيف مع بروتوكول لتحديد الأجنة تمر تقسيم الانقسامية متزامن على وجه التحديد. في هذه الحالة، ينبغي أن تظل الأجنة فقط مع توزيع اجفب-منها مشتتة، غير النووية، ويجب أن يتم تجاهل جميع الأجنة الأخرى. إذ لا يمكن تحديد كثافة النووي، يجب استخدام أساليب بديلة لمرحلة الأجنة بالتشكل على عرضها في الفحص المجهري الخفيفة المنقولة. وجود خلايا القطب ونوى في محيط جنين تشير إلى أن الجنين قد أكمل دورة النووي على الأقل 9، بينما يشير سيلولاريزيشن مرئية على الهامش الدورة النووية 1412.
ج تجارب يمكن أن يؤديها بنجاح باستخدام تشكيلة واسعة من إنزيمات التقييد5. عادة ما تستخدم النهج الحالي الإنزيمات التي تعترف بأي تسلسل 4-قاعدة، مثل مبوي، أو موقع 6-قاعدة اعتراف، مثل هنديي. ميزة القواطع 4-قاعدة على تقطيع 6-قاعدة أنها توفر دقة أعلى المحتملة، نظراً لعمق كاف التسلسل، وتغطية أكثر من تقييد مواقع عبر الجينوم. لا يوجد أي ميزة واضحة في اختيار واحد 4-قاعدة القاطع على آخر5،23،،من3637. الإنزيمات هما الأكثر استخداماً، مبوي ودبنيي، على حد سواء الاعتراف جتك نفس الاعتراف بالموقع. دبنيي أقل حساسية مثلايشن البد، ولا تشكل مصدر قلق في المورفولوجية. البروتوكول المعروضة هنا يمكن أيضا أن نجاح استخدام دبنيي إنزيم التقييد. في الجزء 4-2. المخزن المؤقت لأنزيم التقييد قد يتعين تعديلها للتوافق مع دبنيي، وفقا لتوصيات الشركة المصنعة.
إذا كان حجم جزء من المكتبة التسلسل ينحرف كثيرا عن النطاق المبين في الشكل 2 أ، قد تكون أقل كفاءة تشكيل الكتلة أثناء تسلسل أو تفشل تماما. وفي هذه الحالة، يجب التحقق من توزيع حجم بعد القص والقص معلمات تعديلها تبعاً لذلك. قمم في توزيع الحمض النووي شظايا صغيرة جداً (< 100 bp) أو كبير جداً (> 1,000 bp) أحجام يشير مشاكل مع حجم التحديد، مثل حمل أكثر من الخرز أو المادة طافية التي من المفترض أن يتم تجاهل. غالباً ما هذه المكتبات مع قمم صغيرة في هذه الأحجام غير مرغوب فيها، مثل واحد ويتم المصورة، لا يزال التسلسل بنجاح مع انخفاض طفيف فقط في تجميع الكفاءة.
وينبغي تجنب معدلات عالية من الازدواجية PCR لأن هذا يخفض بشكل كبير عدد ما يلي تسلسل للاستخدام. معدل التكرار PCR ارتباطاً مباشرا بكمية المواد المدخلة. استخدام المزيد من المدخلات ولذلك عادة ما يخفف من المشاكل مع ازدواجية PCR.
تشير أرقام أعلى من القراءات التي تمت تصفيتها بسبب اتجاه القراءة (الشكل 2B) الهضم غير كافية، والتي يمكن أن تكون نتيجة لاستخدام القليل جداً إنزيم أو إدخال الكثير من المواد أو التجانس غير مكتملة للأجنة.
Disclosures
الكتاب ليس لها علاقة بالكشف عن.
Acknowledgements
تم تمويل هذا البحث من "جمعية ماكس بلانك". وأيد C.B.H. على زمالة من المدرسة البحوث الدولية ماكس بلانك – "الطب الحيوي الجزيئي". ونحن نشكر ليث شيلبي واريك ف. فيشاوس فازوا يرجى توفير خط melanogaster المورفولوجية اجفب منها.
Materials
| Name | Company | Catalog Number | Comments |
| Biotin-14-dATP | Life Technologies | 19524016 | |
| MboI | New England Biolabs | R0147L | |
| DNA Polymerase I Klenow Fragment | New England Biolabs | M0210L | |
| T4 DNA Ligase | Thermo Fisher | EL0012 | T4 DNA Ligase Buffer included |
| T4 DNA Polymerase | New England Biolabs | M0203L | |
| Proteinase K | AppliChem | A4392 | |
| GlycoBlue | Life Technologies | AM9516 | |
| Complete Ultra EDTA-free protease inhibitors | Roche | 5892791001 | |
| NEBNext Multiplex Oligos for Illumina (Index Primers Set 1) | New England Biolabs | E7335 | Sequencing Adaptor, Forward (unindexed) PCR primer and Reverse (indexed) PCR primer and USER enzyme used in the Library preparation section are components of this kit |
| NEBNext Ultra II DNA Library Prep Kit | New England Biolabs | E7645 | End Prep Enzyme Mix, End Prep Reaction Buffer, Ligation Enhancer, Ligation Master Mix and Polymerase Master Mix used in the Library preparation section are components of this kit |
| Covaris S2 AFA System | Covaris | ||
| DNA LoBind Tubes, 1.5 mL | Eppendorf | 0030108051 | |
| Falcon cell strainer 100 µm | Corning | 352360 | Embryo collection baskets |
| 37% formaldehyde | VWR | 437536C | |
| Heptane | AppliChem | 122062.1612 | |
| M165 FC fluorescent stereo microscope | Leica | ||
| M165 FC DFC camera | Leica | ||
| Metal micro pestle | Carl Roth | P985.1 | Used to lyse embryos in step 4.1.4 |
| RNase A | AppliChem | A3832,0050 | |
| Dynabeads MyOne Streptavidin C1 | Life Technologies | 65002 | Streptavidin coated magnetic beads |
| Ampure XP beads | Beckman Coulter | A63881 | |
| Qubit 3.0 Fluorometer | Thermo Fisher Scientific | Q33216 | |
| Qubit assay tubes | Thermo Fisher Scientific | Q32856 | |
| Qubit dsDNA HS Assay Kit | Thermo Fisher Scientific | Q32854 | |
| Phosphate buffered saline (PBS) | Sigma-Aldrich | P4417 | |
| eGFP-PCNA flies | Gift from S. Blythe and E. Wieschaus | ||
| Sodium hypochlorite 13% | Thermo Fisher | AC219255000 | |
| Triton X-100 | AppliChem | A4975 | |
| Tris buffer pH 8.0 (1 M) for molecular biology | AppliChem | A4577 | |
| NaCl | AppliChem | A2942 | |
| IGEPAL CA-630 | Sigma-Aldrich | I8896 | |
| 1.5 mL microcentrifuge tubes | Greiner Bio-One | 616201 | |
| SDS for molecular biology | AppliChem | A2263 | |
| 10x CutSmart buffer | New England Biolabs | B7204S | Restriction enzyme buffer |
| PCR Nucleotide Mix | Sigma-Aldrich | 11814362001 | Unmodified dCTP, dGTP, dTTP |
| BSA, Molecular Biology Grade | New England Biolabs | B9000S | |
| EDTA 0.5 M solution for molecular biology | AppliChem | A4892 | |
| Sodium acetate 3 M pH 5.2 | Sigma-Aldrich | S7899 | |
| DynaMag-2 Magnet | Life Technologies | 12321D | Magnetic stand |
| Intelli-Mixer RM-2L | Omnilab | 5729802 | Rotator |
| ThermoMixer F1.5 | Eppendorf | 5384000012 | Mixer |
| Small Embryo Collection Cages | Flystuff.com | 59-100 | Egg collection cage |
| Centrifuge 5424 R | Eppendorf | 5404000413 | |
| C1000 Touch Thermal Cycler | Bio-Rad | 1851148 | |
| PCR tube strips | Greiner Bio-One | 673275 | |
| NEBuffer 2.1 | New England Biolabs | B7202S | T4 DNA Polymerase buffer |
References
- Bonev, B., Cavalli, G. Organization and function of the 3D genome. Nat Rev Genet. 17 (11), 661-678 (2016).
- Lieberman-Aiden, E., et al. Comprehensive mapping of long-range interactions reveals folding principles of the human genome. Science. 326 (5950), 289-293 (2009).
- Dixon, J. R., et al. Topological domains in mammalian genomes identified by analysis of chromatin interactions. Nature. 485 (7398), 376-380 (2012).
- Jin, F., et al. A high-resolution map of the three-dimensional chromatin interactome in human cells. Nature. , (2013).
- Rao, S. S. P., et al. A 3D Map of the Human Genome at Kilobase Resolution Reveals Principles of Chromatin Looping. Cell. 159 (7), 1665-1680 (2014).
- Darbellay, F., Duboule, D. Topological Domains, Metagenes, and the Emergence of Pleiotropic Regulations at Hox Loci. Current topics in developmental biology. 116, 299-314 (2016).
- Beagan, J. A., et al. Local Genome Topology Can Exhibit an Incompletely Rewired 3D-Folding State during Somatic Cell Reprogramming. Cell stem cell. 18 (5), 611-624 (2016).
- Andrey, G., et al. Characterization of hundreds of regulatory landscapes in developing limbs reveals two regimes of chromatin folding. Genome Res. 27 (2), 223-233 (2017).
- Krijger, P. H. L., de Laat, W. Regulation of disease-associated gene expression in the 3D genome. Nature Reviews. Molecular Cell Biology. 17 (12), 771-782 (2016).
- Sexton, T., et al. Three-dimensional folding and functional organization principles of the Drosophila genome. Cell. 148 (3), 458-472 (2012).
- Ghavi-Helm, Y., et al. Enhancer loops appear stable during development and are associated with paused polymerase. Nature. 512 (7512), 96-100 (2014).
- Foe, V. E., Alberts, B. M. Studies of nuclear and cytoplasmic behaviour during the five mitotic cycles that precede gastrulation in Drosophila embryogenesis. J Cell Sci. 61, 31-70 (1983).
- Blythe, S. A., Wieschaus, E. F. Zygotic Genome Activation Triggers the DNA Replication Checkpoint at the Midblastula Transition. Cell. 160 (6), 1169-1181 (2015).
- Blythe, S. A., Wieschaus, E. F. Establishment and maintenance of heritable chromatin structure during early Drosophila embryogenesis. eLife. 5, e20148(2016).
- JoVE Science Education Database. Embryo and Larva Harvesting and Preparation. Biology I: yeast, Drosophila and C. elegans. Drosophila melanogaster. , JoVE, Cambridge, MA. (2017).
- Sicaeros, B., O'Dowd, D. K. Preparation of Neuronal Cultures from Midgastrula Stage Drosophila Embryos. Journal of Visualized Experiments. (5), (2007).
- Shermoen, A. W. Preparation of Baskets for Drosophila Egg Collections, Treatments, and Incubations. Cold Spring Harbor Protocols. (10), (2008).
- Ay, F., Noble, W. S. Analysis methods for studying the 3D architecture of the genome. Genome biology. 16 (1), 183(2015).
- Lazaris, C., Kelly, S., Ntziachristos, P., Aifantis, I., Tsirigos, A. HiC-bench: comprehensive and reproducible Hi-C data analysis designed for parameter exploration and benchmarking. BMC Genomics. 18 (1), (2017).
- Servant, N., et al. HiC-Pro: an optimized and flexible pipeline for Hi-C data processing. Genome Biology. 16 (1), (2015).
- Durand, N. C., et al. Juicer Provides a One-Click System for Analyzing Loop-Resolution Hi-C Experiments. Cell systems. 3 (1), 95-98 (2016).
- Lajoie, B. R., Dekker, J., Kaplan, N. The Hitchhiker's guide to Hi-C analysis: Practical guidelines. Methods. 72, 65-75 (2015).
- Schmitt, A. D., Hu, M., Ren, B. Genome-wide mapping and analysis of chromosome architecture. Nature Reviews. Molecular Cell Biology. 17 (12), 743-755 (2016).
- Shin, H., et al. TopDom: an efficient and deterministic method for identifying topological domains in genomes. Nucleic Acids Res. 44 (7), e70(2016).
- Kruse, K., Hug, C. B., Hernández-Rodríguez, B., Vaquerizas, J. M. TADtool: visual parameter identification for TAD-calling algorithms. Bioinformatics. 32 (20), 3190-3192 (2016).
- Lévy-Leduc, C., Delattre, M., Mary-Huard, T., Robin, S. Two-dimensional segmentation for analyzing Hi-C data. Bioinformatics. 30 (17), Oxford, England. i386-i392 (2014).
- Filippova, D., Patro, R., Duggal, G., Kingsford, C. Identification of alternative topological domains in chromatin. Algorithms for molecular biology: AMB. 9 (1), 14(2014).
- Crane, E., et al. Condensin-driven remodelling of X chromosome topology during dosage compensation. Nature. 523 (7559), 240-244 (2015).
- Durand, N. C., et al. Juicebox Provides a Visualization System for Hi-C Contact Maps with Unlimited Zoom. Cell systems. 3 (1), 99-101 (2016).
- Zhou, X., et al. Exploring long-range genome interactions using the WashU Epigenome Browser. Nature Methods. 10 (5), 375-376 (2013).
- Ramírez, F., et al. High-resolution TADs reveal DNA sequences underlying genome organization in flies. bioRxiv. , 115063(2017).
- Kerpedjiev, P., et al. HiGlass: Web-based Visual Comparison And Exploration Of Genome Interaction Maps. bioRxiv. , 121889(2017).
- Hug, C. B., Grimaldi, A. G., Kruse, K., Vaquerizas, J. M. Chromatin Architecture Emerges during Zygotic Genome Activation Independent of Transcription. Cell. 169 (2), (2017).
- Berkum, N. L., et al. Hi-C: a method to study the three-dimensional architecture of genomes. Journal of Visualized Experiments: JoVE. (39), (2010).
- Naumova, N., et al. Organization of the mitotic chromosome. Science. 342 (6161), 948-953 (2013).
- Denker, A., de Laat, W. The second decade of 3C technologies: detailed insights into nuclear organization. Genes & development. 30 (12), 1357-1382 (2016).
- Belaghzal, H., Dekker, J., Gibcus, J. H. Hi-C 2.0: An optimized Hi-C procedure for high-resolution genome-wide mapping of chromosome conformation. Methods (San Diego, Calif). 123, 56-65 (2017).
Reprints and Permissions
Request permission to reuse the text or figures of this JoVE article
Request PermissionExplore More Articles
This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. All rights reserved