
Method Article
Genom çapında kromatin Conformation yakalama kitaplıklardan sıkıca aşamalı erken Drosophila embriyo nesil
Bu Makalede
Özet
Bu eser bir protokolünü açıklar yüksek çözünürlüklü in situ Hi-C kitaplıklardan nesil sıkıca öncesi gastrulasyon Drosophila melanogaster sahnelenen için embriyo.
Özet
Kromatin üç boyutlu mimari soruşturma gen düzenlemesi mekanizmalarının içine çok değerli fikir sunar. Burada, kromatin conformation yakalama tekniği in situ Hi-C üzerinde gerçekleştirmek için bir protokol tarif Drosophila melanogaster embriyo nüfus düzenledi. Tek bir deney çekirdeğinde ortaya tüm kromatin etkileşimler eşleme sağlar bir sıralama kitaplığı sonucudur. Embriyo sıralama floresan stereo mikroskop ve nükleer bir işaretçi içeren bir transgenik sinek çizgi kullanarak el ile yapılır. Bu teknik, embriyo nüfus her nükleer bölünme döngüsüne ve tanımlı hücre döngüsü durumundaki kullanarak çok yüksek saflık ile elde edilebilir. İletişim kuralı da gastrulasyon ötesinde Büyük Embriyo sıralamak için adapte olabilir. Sıralanmış embriyo situ için Hi-c girişleri kullanılır Tüm deneyler Kütüphane hazırlık, sıralama dahil olmak üzere, beş gün içinde tamamlanabilir. Protokol düşük giriş gereksinimleri ve güvenilir bir şekilde 20 blastoderm sahne embriyo giriş malzeme olarak kullanarak çalışır. Sonraki nesil sıralaması için bir sıralama kitaplığı sonucudur. Sonra sıralama, veri topolojik etki alanı (TAD) yapısı, kromatin döngüler ve kromatin ilişkilendirme hakkında bilgi elde etmek için çok çeşitli araçlar kullanılarak analiz genom çapında kromatin etkileşim haritalar içine işlenebilir Drosophila geliştirme sırasında bölmeleri.
Giriş
Kromatin conformation yakalama (3C) çekirdeği1kromatin topolojisinin çalışmaya son derece yararlı bir yöntem olarak ortaya çıkmıştır. 3C değişken Hi-C tek deneme2çekirdeğinde ortaya tüm kromatin etkileşimler iletişim frekanslarını ölçmek sağlar. Hi-c uygulama bulma ve kromatin kuruluşun TADs, bölmeleri ve döngüler3,4,5gibi birçok temel ilkeleri karakterizasyonu önemli bir rol oynamıştır.
Kromatin mimarisi, gelişimsel geçişler ve hücre farklılaşması bağlamda çalışmaların giderek gen düzenlemesi sırasında bu işlemleri6,7,8mekanizmaları çözülmeye kullanılır, 9. Drosophila melanogasterolan geliştirme ve genom de karakterizedir, model organizmalar büyük ilgi biridir. Ancak,10,11 Drosophila kromatin mimarisinde vitro doku kültürü ayarları olmuştur dışında araştırmak kaç araştırmalar yapılmıştır. Embriyo 16-18 h sonrası döllenme, TADs ve bölmeleri memelilerde benzer yapıları anımsatan tanımlanan10, hangi yükseltir soru hangi rolü gen düzenlemesi sırasında Drosophila embriyo içinde oynuyorlar vardı geliştirme. Gastrulasyon önce geliştirme, erken evrelerinde özellikle bu tür çalışmalar Teknik olarak zor olan. Gastrulasyon önce Drosophila embriyo 8-60 dk döngüsü12,13başına son derece hızlı bir tempoda devam 13 zaman uyumlu nükleer bölünmeler tabi. Buna ek olarak, farklı aşamaları ayırt etmek için görsel özellikler eksikliği zor yapmak sıkıca aşamalı embriyo malzemesi yeterli miktarda elde etmek.
Kromatin nükleer döngüsü çözünürlükte erken Drosophila gelişiminde Hochschule'a sağlayan bir protokol geliştirmek için biz iki varolan teknikleri kombine: yüksek çözünürlüklü nesil tüm sağlayan in situ Hi-C, Genom kişi haritalar5ve eGFP-PCNA transgene13,14ifade bir transgenik Drosophila satırını kullanarak embriyo evreleme. Bu transgene çekirdeğine Interphase sırasında yerelleştirir ve mitoz sırasında sinsisyal blastoderm yayılır. Bu özelliği kullanarak, kolayca kendi nükleer yoğunluğu tarafından farklı aşamaları ve mitotik embriyo GFP sinyal dağılım tarafından ayırt etmek mümkündür.
Birlikte, bu tekniklerin kromatin olabildiğince az 20 Drosophila embriyo olarak gelen yüksek çözünürlüklü üç boyutlu yapısını eğitim etkinleştirin. Bu iletişim kuralı bir tek nükleer bölümü döngüsü embriyo olasılığını elde etmek için hasat ve Drosophila embriyo sıralama için yönergeler içerir. Daha fazla elde edilen embriyolar in situ Hi-c gerçekleştirmek için nasıl kullanılacağını açıklar Sonuçta bir sonraki nesil sıralama makinelerde sıralama için uygun bir nükleotit kütüphanesidir. Elde edilen sıralama okuma sonra tüm Drosophila genom kapsayan detaylı kromatin etkileşim haritalar işlenebilir.
Protokol
1. drosophila embriyo koleksiyonu
Not: Bir önceki yayın15' te gösterildiği gibi bir eşdeğer embriyo koleksiyon gerçekleştirilebilir.
- Genç eGFP-PCNA sinekler transfer (< 1 haftalık)16 (% 1 etanol, % 1 asetik asit ve % 4 agar) içine yumurta toplama kafesleri mayalanarak koleksiyonu ile tabaklar.
- 25 ° C'de ayarla bir kuluçka koleksiyonu kafesleri taşımak 1-2 gün önce yumurta toplama için kuluçka yumurta verimi önemli ölçüde artırır. Koleksiyon tabakları günde iki kez değiştirin.
- Embriyo toplama odası 30-60 dk aralıklarla gelen içeren plakaları kaldırın. Ama gelişim aşamalarında daha sıkı dağıtım daha az embriyolar daha küçük aralıklar neden. Paralel olarak birden fazla kafes kadar bu ideal toplamak > 200 yumurta her 30-60 dk koyulur.
- Embriyoların istenen yaşına kadar tabak 25 ° C'de depolayın. Blastoderm aşaması için embriyo (nükleer döngüsü 14), kuluçkaya yaklaşık 2 h için.
- Tüm yüzeyi su ile kaplıdır, kuluçka 2 h sonra musluk suyu bir fışkırmak şişe toplama plaka ekleyin. Embriyo askıya alma ve yumuşak bir fırça kullanarak Maya.
- Koleksiyon plaka resuspended embriyo bir embriyo toplama sepeti (ticari hücre süzgeçler) 100 µm gözenek boyutu veya ev yapımı sepet17 iş iyi fışkırmak şişe ek musluk suyu ekleyerek gerekirse dökün. Bu aşamada, embriyo paralel olarak toplanan tüm plakaları dan birleştirin. Havuza alınan örnek tek bir toplu iş temsil eder.
- Embriyo de musluk suyu sepetle fışkırmak şişe 30 dan durulama tarafından yıkayın kadar tüm Maya kalıntı akıp s.
- Dechorionate embriyo içine su % 2.5 sodyum hipoklorit çözeltilerine toplama sepeti yerleştirerek. Dönen tarafından ışık ajitasyon koryon kaldırılmasını kolaylaştırır. Böylece ne zaman sepet dışarı kaldırdı ve tekrar, sular altında onlar çözüm yüzeyde float embriyo yeterince hidrofobik kadar devam ~1.75–2 min almalı.
Dikkat: Sodyum hipoklorit aşındırıcı. Uygun kişisel koruyucu ekipman giymek. Çözümler içeren < % 10 sodyum hipoklorit genelde lavaboda tasfiye edilmesi, ana bilgisayar Enstitüsü yönetmeliklerine kontrol etmek emin olun. - Sepet çözümden kaldırmak ve çamaşır suyu kokusu artık fark edilir kadar fışkırmak şişe dokunun su ile iyice durulayın.
2. embriyo fiksasyon
Not: Optimum fiksaj koşulları, deterjan, formaldehit ve fiksasyon, süresi öncelikle konsantrasyon embriyo aşamasında uygun ampirik olarak belirlenmiş olması gerekir. Sinsisyal blastoderm çevresinde aşamaları için % 0,5 Triton X-100 ve % 1,8 formaldehit son bir konsantrasyon sulu faz iş de. Embriyo sahne 9 ötesinde daha sonraki aşamaları için daha fazla en iyi duruma getirme bu parametreler gerekli olabilir. Tüm çözümler fiksasyon ve sıralama sırasında kullanılan proteaz inhibitörleri içermelidir.
- Toplama sepeti ters çevir ve 15 mL konik Santrifüjü tüp yerleştirin. Embriyo sepeti üzerinden bir Pasteur pipet PBS-T dağıtımı kullanarak tüpüne sifonu (PBS, % 0.5 Triton X-100).
- Alt kısmında yerleşmek ve PBS-T. ile 2 ml toplam ses seviyesini embriyo izin
- Heptane 6 mL ve 100 µL % 37 formaldehit suya ekleyin.
Dikkat: Heptane ve formaldehit teneffüs ne zaman zehirli veya cilt sonra başvurun. Uygun kişisel koruyucu donanımları ve duman mahallede çalışma. Heptane veya formaldehit içeren atık ayrı ayrı ana bilgisayar Enstitü nün düzenlemelere göre tasfiye edilmesi gerekiyor. - Formaldehit eklenmesi sonra 15 dk sayacýný baþlatmak ve şiddetle tüp yukarı ve aşağı 1 dk. için el ile çalkalanır. Sulu ve Organik faz formu bir şampuan gibi tutarlılık için birleştirir.
- Bir çevirme karıştırıcı formaldehit eklenmesinden sonra 10 dk kadar tahrik.
- Vasıl 500 x g embriyo tüp alt toplamak için oda sıcaklığında 1 dk santrifüj kapasitesi.
- Tüm şampuan gibi sıvı Aspire edin ve, herhangi bir embriyo Aspire edin değil için dikkat çekici atın. Şampuan gibi süpernatant az kalan miktarda sorunları neden olmaz.
- 15 dakika sonra formaldehit, ek PBS-T 5 ml formaldehit gidermek için 125 mM glisin ile embriyo resuspend. 1 dk. için yukarı ve aşağı sallayarak kuvvetle karıştırın.
- 500 x g 1 dk. için oda sıcaklığında, santrifüj kapasitesi ve süpernatant Aspire edin.
- Embriyo 5 mL buz gibi PBS-T. resuspending tarafından yıkama Embriyo yerleşmeye ve Aspire edin tüm süpernatant izin.
- Adım 2.10 makinaya iki kez daha tekrarlayın.
- Embriyo buz üzerinde sıralama kadar tutun. Genellikle, sıralama için 3-4 toplu işlemleri devam etmeden önce sinek embriyo toplamak için iyi bir fikirdir. Ancak, embriyo aynı gün sıralanması. Genişletilmiş depolama buz üzerinde veya buzdolabında değişmiş embriyo morfoloji için yol açar.
3. embriyo sıralama
Not: Sıralama 60-80 X büyütme, GFP filtre ile donatılmış flüoresan herhangi bir stereo mikroskop üzerinde yapılabilir.
- 1000 µL pipet kullanarak, küçük cam kap, tercihen bir koyu renk, sıralamak için uygun bir toplu iş yaklaşık 100 embriyo transfer ve buz üzerinde yerleştirin.
-
Sıralama embriyo tarafından arzu edilen embriyo bir iğne veya şırınga ucu kullanarak ayrı bir yığın iterek nükleer yoğunluğu ve hücre döngüsü duruma göre (Şekil 1).
- EGFP-PCNA (Şekil 1E) dağınık, nükleer olmayan dağılımı ile tüm embriyoların kaldırın. Ayrıca, nükleer olmayan GFP sinyal kısmen gösteren embriyolar kaldırılması gerekir.
- Sıralama yardım resimleri resim 1 ' de bir kılavuz olarak kullanarak her yığında nükleer döngüsü 12, 13 ve 14 başvuru embriyo bir line-up araya. Bu line-up onların sahne belirlemek için bilinmeyen bir sahne embriyo bir başvuru embriyoların ile eşleştirmek için kullanın.
- Gelişim aşaması için başvuru embriyo doğrulamak için embriyo görüntüleme ve çekirdekleri bir alanda mesafe bilgi sağlar görüntüleme yazılımı kullanarak 2500 µm2 embriyo yüzeyde sayılması nükleer yoğunluğu ölçmek.
Not: Beklenen çekirdek için 2.500 µm2 yüzölçümü 12-16 çekirdek nükleer döngüsü 12 ve 20-30 çekirdekleri nükleer döngüsü 1313sayısıdır.
- Bir kez tüm embriyoların uygun aşamada ayrılır, belgelendirme ve kalite kontrol için embriyo fotoğraf çekmek. Stereo mikroskop kendisi donatmak ile bir fotoğraf makinesi ölçü birimi değilse, herhangi bir epifluorescence mikroskop GFP filtreleri ile kullanılabilir.
- Pipet 1000 µL pipet, taze tüp ve yer buz üzerinde kullanarak istenen embriyolar kadar.
- Yeterli embriyo için planlanan deney sıralanır kadar devam edin. İçin embriyo büyük 9, sahne daha genellikle 20 embriyo bir in situ Hi-C deney için yeterlidir. 12 nükleer döngüsü 80 embriyo iyi bir başlangıç noktası vardır. Önceki döngüleri, embriyo sayısı yaklaşık her döngüsü için iki katına.
- Havuzu ve split embriyo bir tüp bir tek in situ Hi-C için yeterli embriyo içerir şekilde 1,5 mL tüpler içine deneyin. Bu tüpler düşük DNA bağlama özellikleri ile aynı tüp için tüm iletişim kuralının kullanılacağını ve DNA adsorpsiyon düşük DNA konsantrasyonlarda önemli zararlar açabilir kullanmadan önce tavsiye edilir.
- Kısaca, 100 x g oda sıcaklığında tüpler spin ve süpernatant kaldırın. Embriyo gibi kuru olmalıdır dondurma için mümkün olduğunca.
- Flaş dondurma embriyo sıvı azot ve mağaza-80 ° C'de tüplerde batış tarafından
4. in Situ Hi-C
- Lizis
- Dondurulmuş embriyo ile tüplerin buza koyun.
- Embriyo buz gibi lizis arabellek (10 mM Tris-Cl pH 8.0, 10 mM NaCl, %0.2 IGEPAL CA-630, proteaz inhibitörleri; içinde çözünmüş su) 500 µL içinde resuspend. Daha sonra embriyo tüp alt kısmında yerleşmek izin için 1 dakika bekleyin.
- 1.5 mL microcentrifuge tüp sıkıca uyacak şekilde tasarlanmıştır buz üzerinde önceden soğutmalı metal bir mikro havaneli kullanarak embriyo eziyet.
- Embriyo Tantanacı önlemek için tüp alt dokunana kadar havaneli yavaş yavaş Ekle, aşağı doğru itin ve havaneli ile iki kez her iki yönde döndürerek eziyet.
- Havaneli ile çok hafif kaldırın, tekrar tüp altına itmek ve taşlama tekrarlayın.
- 4.1.3.2 10 kez yineleyin veya embriyo tamamen lysed kadar. Çözüm homojen olmalıdır ve embriyo hiçbir kalıntı büyük parçaları kalmalıdır.
- 15 dk. Spin 1000 x g, 5 min için 4 ° C de buzda homojenize süspansiyon kuluçkaya ve süpernatant atın.
- Pelet yukarı ve aşağı pipetting 500 µL buz gibi lizis arabellekte resuspending tarafından yıkayın.
- 4.1.4 olduğu gibi yine spin ve süpernatant atın.
- Yukarı ve aşağı pipetting yıkanmış Pelet 100 µL % 0.5 Sodyum Lauryl Sülfat (SDS), içinde resuspend. Çekirdeği bir Isıtma blok içinde 65 ° C'de 10 dakika için kuluçka tarafından permeabilize. SDS 50 µL % 10 Triton X-100 ve 120 µL su ekleyerek gidermek. Tüp flicking tarafından karıştırın.
- Isı blok 15 dakika 37 ° C'de kuluçkaya.
- Restriksiyon enzimi sindirim
- Restriksiyon enzimi arabellek x 10 ve 20 5 U/µL MboI U 25 µL ekleyin. Tüp flicking tarafından karıştırın.
- DNA 37 ° c ısı blok hafif ajitasyon (750 devir/dakika) altında 90 dk için kuluçka sindirmek.
- Başka bir 20 U MboI ekleyin ve kuluçka 90 dk için devam edin.
- Isı-MboI 20 dk 62 ° C'de kuluçka tarafından devre dışı bırakabilirsiniz.
- Çıkıntı doldurma
Not: biotinylated dATP ile çıkıntı doldurma belirli ve birleştirilmiş parçaları seçim sağlar. Biotin-dATP unligated künt ucunda verimli bir şekilde kaldırılır, ancak biotin-dATP tüp ligasyonu kavşaklar, eksonükleaz aktivitesi T4 DNA polimeraz (Bölüm 4.6), koruma altındadır. Streptavidin kaplı boncuk 4.7 bölümünde bu nedenle özellikle kullanarak açılan ve birleştirilmiş, chimeric DNA parçaları için zenginleştirir.- 18 µL 0.4 mM biotin-14-dATP, 2.25 µL değiştirilmemiş bir dCTP/dGTP/dTTP karışımı (3,3 mM her) ve 8 µL parçasının 5 U/µL DNA polimeraz ı Klenow ekleyin.
- Tüp flicking tarafından mix ve 37 ° c ısı blok içinde 90 dk için kuluçkaya.
- Tüp ligasyonu
- 657 µL su, 10 x T4 DNA ligaz arabelleği, %10 100 µL 120 µL ekleyin Triton X-100, 20 mg/mL Sığır serum albumin (BSA) ve karışımı tüp flicking tarafından 6 µL. Son olarak tüp flicking tarafından 5 µL 5 U/µL T4 DNA ligaz ve karışımı ekleyin.
- Tüpü yavaşça 2s için oda sıcaklığında (20 devir/dakika) döndürün.
- 5 U/µL T4 DNA ligaz 5 µL ikinci taksit ekleyin ve 2 için daha fazla h dönen devam edin.
- Çekirdeklerin 2500 x g 5 min için de aşağı spin ve süpernatant atın.
- DNA ekstraksiyon
- Pelet ayıklama arabellek (50 mM Tris-Cl pH 8.0, 50 mM NaCl, 1 mM Ethylenediaminetetraacetic asit (EDTA), % 1 SDS; içinde çözünmüş su) 500 µL içinde resuspend ve tüp flicking tarafından 20 mg/mL İndinavir K. Mix 20 µL ekleyin.
- Protein 55 ° c 30 dk 1000 devir / dakikada sallayarak için kuluçka sindirmek.
- İçin de-crosslink, 5 M NaCl 130 µL ekleyin ve bir gecede 1000 devir / dakikada sallayarak 68 ° C'de kuluçkaya.
- Pipet örnek tercihen düşük DNA bağlama özelliklere sahip yeni bir 2 mL tüp içine.
- 0.1 x birimleri (63 µL) 3 M sodyum asetat pH 5.2 ve 2 µL 15 mg/ml GlycoBlue ekleyin. Mix de ters çevirme. Ters çevirme tarafından 1.6 x birimleri (1,008 µL) saf mutlak etanol ve karışımı ekleyin.
- 20.000 x g için en az 30 dk 4 ° c de 15 dakika santrifüj süreyle-80 ° C'de kuluçkaya. DNA Pelet genelde çok küçük, neredeyse görünmez ve yalnızca GlycoBlue mavi renk nedeniyle lekeli olabilir.
- Süpernatant pipet ucu ters duvar boyunca tüp içine DNA Pelet olduğu üzerinden hareket kaldırmak çok dikkatli. Küçük kalan damlacıkları genellikle kolayca kaldırılır Bu adım ve aşağıdaki yıkama sırasında P10 ucu kullanarak tüpler dışına iterek yerine onları pipetting.
- Pelet 800 µL % 70 etanol ekleyerek yıkayın. Mix ters çevirme tarafından ve santrifüj kapasitesi 20.000 x g , oda sıcaklığında 5 dakika süreyle, bu yıkama en az bir kez yinelenir.
- Etanol bütün izlerini kaldırmak ve tüp ayakta kurumasını 5 min için kapağı açık bırakın. Hiçbir sıvı kalan kez, 10 mM Tris-Cl pH 8.0 50 µL ekleyin. Art arda çözüm tüp duvarına nerede Pelet DNA solubilize için yer oldu alana pipette.
- 20 mg/ml RNase A, Tüp, flicking tarafından mix 1 µL ekleyin ve 15 dakika sindirimi RNA 37 ° C'de kuluçkaya. Örnek şimdi gece buzdolabında saklanan ya da süresiz olarak-20 ° C'de donmuş.
- Onay floresan bir boya kullanarak DNA konsantrasyonu tahlil üretici yönergelerine göre temel. DNA'ın toplam tutarı örnek en az 10 olmalıdır ng, aksi halde çok az malzeme amplifikasyon için mevcuttur ve Kütüphane karmaşıklığı büyük olasılıkla düşük. Bu durumda, malzeme başlangıç miktarını muhtemelen yeterli değildi veya malzeme yol boyunca, belki de lizis ve yağış sırasında kayboldu.
- Biotin kaldırma ve DNA kesme
- Birlikte 10 x T4 DNA polimeraz arabelleği 12 µL, 1 mM dATP, 3 µL, 1 mM dGTP, 3 µL ve 46 µL su ekleyin. Tüp flicking tarafından karıştırın. 3 U/mL T4 DNA polimeraz 5 µL eklemek, tüp flicking tarafından mix ve 30 dakika için 20 ° C'de kuluçkaya.
- Reaksiyon durdurmak için 0, 5 M EDTA 3 µL ekleyip su yaklaşık 120 µL bir birim için örnek getirmek için kullanabilirsiniz.
- DNA sonication aygıtıyla üreticinin talimatlarına göre 200-400 bp boyutuna kesme. Malzemeler tablobelirtilen sonicator kullanarak, aşağıdaki programı uygundur: 2 döngüleri her 50 s, % 10 görev, yoğunluğu 5, 200 devir/seri çekim.
- Biotin açılan
- 10 mg/mL streptavidin kaplı manyetik boncuklar 30 µL yeni bir tüp pipet, bunları manyetik bir stand üzerine ayırın ve süpernatant atın.
- Boncuk 1 x B & W arabellek (5 mM Tris-Cl pH 7.4, 0,5 mM EDTA, 1 M NaCl; içinde çözünmüş su) + %0.1 Triton X-100 ve mix vortexing tarafından resuspend. Tüp manyetik bir stand üzerine yerleştirin ve boncuklar, marka ve model bağlı olarak ayrılır 1 – 5 min için bekleyin.
- Aspire edin ve süpernatant pipet ucu boncuk konumlandırıldığı duvar boyunca kayma sırasında atın. Boncuk 2 B & W arabellek (10 mM Tris-Cl pH 7.4, 1 mM EDTA ve 2 M NaCl) x 120 µL resuspend. Vortexing tarafından mix.
- Yamultulmuş DNA bir yeni düşük DNA bağlama tüp için transfer ve vortexing tarafından 120 µL boncuk süspansiyon içinde 2 X B & W tampon ile karıştırın. Boncuk DNA örneği 20 devirde 15 dakika ile döndürün.
- Boncuk manyetik bir stand ayırmak ve süpernatant atın.
- Resuspend 1 x B & W + %0,1 600 µL Triton X-100 boncuk ve 55 ° c için 2 dk 1000 devir / dakikada sallayarak, kuluçkaya. Ayrılıktan sonra süpernatant atmak. Bu yıkama bir kez yinelenir.
- Boncuk 10 mM Tris-Cl pH 8.0 600 µL hemen ile yıkayın ve süpernatant ayrılıktan sonra atın.
- Boncuk 10 mM Tris-Cl pH 8.0 50 µL içinde resuspend.
5. sıralama kitaplığı hazırlık
Not: Tüm kütüphane adımları yapılır bir ticari DNA Kütüphane hazırlık bileşenleri kullanarak ( Tablo malzemelerigörmek) kit. Ancak, alternatif kitleri veya diğer kimyasalları yerine olabilir. Yağış dondurucu depolama sırasında form kitaplığı hazırlık aracıları eğilimindedir. Bu nedenle tüm yağış reaktifleri kullanmadan önce tasfiye edilir emin olmak önemlidir.
-
Son onarım
- 10 mM Tris-Cl pH 8.0 50 µL boncuk süspansiyon yeni bir PCR tüp içine aktarın.
- 3 µL son hazırlık enzim karışımı ve son hazırlık tepki arabelleği 7 µL ekleyin. Yukarı ve aşağı pipetting tarafından karıştırın.
- Tüp bir termal cycler aktarmak ve aşağıdaki programı çalıştır: 20 ° C 30 dk, 30 dk ve 4 ° C'de tutmak için 65 ° C
-
Bağdaştırıcı ligasyonu
- Tüp ligasyonu Master Mix, 1,5 µM 2.5 µL 30 µL eklemek adaptör (stoktan için 1,5 µM seyreltik) sıralama ve ligasyonu artırıcı 1 µL boncuk süspansiyon için. Yukarı ve aşağı pipetting tarafından karıştırın.
- 20 ° c termal cycler 15dk için kuluçkaya.
- Kullanıcı enzim 3 µL ekleyin. Yukarı ve aşağı pipetting tarafından karıştırın.
- 15 dakika içinde bir termal cycler 37 ° C'de kuluçkaya.
- Boncuk manyetik bir stand ayırmak ve süpernatant kaldırın.
- Boncuk yıkamak, resuspend için Triton X-100 1 x B & W arabellek + %0,1 100 µL içinde boncuk. Karıştırmak yanında vortexing ve yeni microcentrifuge tüp transferi. Boncuk manyetik bir stand ayırmak ve süpernatant kaldırın.
- Bu yıkama bir kez aynı arabelleği 600 µL kullanarak yineleyin.
- 10 mM Tris-Cl pH 8.0, karıştırmak vortexing, yanında 600 µL boncuk resuspend ve yeni bir tüp boncuk aktarın.
- Boncuk manyetik bir stand ayrı süpernatant atmak ve boncuk 10 mM Tris-Cl pH 8.0 50 µL içinde resuspend.
-
PCR güçlendirme
- İki PCR tüpleri hazırlayın ve her, 25 µL polimeraz Master Mix, 1,5 µL 10 µM ileri (Sıralanmayan) PCR astar ve 10 µM ters (Dizinli) PCR astar 1,5 µL karıştırın.
Not: İleri (Sıralanmayan) PCR astar:
5'-AATGATACGGCGACCACCGAGATCTACACTCTTTCCCTACACGACGCTCTTCCGATC * T-3´.
Geriye doğru (Dizinli) PCR astar:
5'-CAAGCAGAAGACGGCATACGAGATNNNNNNGTGACTGGAGTTCAGACGTGTGCTCTTCCGATC * T-3´. * dizinli PCR astar phosphorothioate tahvil ve Ns gösterir. - Her tüpün içinde yukarı ve aşağı pipetting tarafından 22 µL boncuk süspansiyon ve karışımı ekleyin.
- Aşağıdaki programı kullanarak PCR çalıştırın: 98 ° C için 1 dakika, (98 ° C 15 s, 75 65 ° C s, 1.5 ramping ° C/s) 65 ° C 5 min ve 4 ° C'de tutmak için 9 - 12 kez, tekrarlanan
Not: Ampirik olarak belirlenmesini amplifikasyon döngü sayısı vardır. Ancak, 12'den fazla döngüleri gerekli kütüphaneleri genellikle düşük karmaşıklığı vardı ve yüksek kalitede Hi-C haritalar sonuçlanmamıştır bulduk. Öte yandan, az 12 döngüleri gerekli kütüphaneleri olumsuz için tam 12 döngüleri yükseltecek tarafından etkilenen değil. Bu nedenle, varsayılan amplifikasyon 12 döngüleri için mümkündür. - Havuzu iki PCR reaksiyonları bir tek microcentrifuge tüp boncuk manyetik bir stand ayrı ve yeni bir tüp kitaplığına içeren süpernatant aktarın.
- İki PCR tüpleri hazırlayın ve her, 25 µL polimeraz Master Mix, 1,5 µL 10 µM ileri (Sıralanmayan) PCR astar ve 10 µM ters (Dizinli) PCR astar 1,5 µL karıştırın.
-
Boyut seçimi
- Ampure XP de sallayarak oda sıcaklığında ve mix boncuk süspansiyon getirmek.
- Havuza alınan su ile tam olarak 200 µL PCR tepki hacmi getir. PCR ve manyetik ayırma sırasında bazı orijinal hacminin genellikle kayıp. Birim için 200 µL pipet ayarlayarak doğrulayın ve birimin tümü reaksiyon Aspire edin. Hava emişli, daha eklenmesi gereken su. Birimin 200 µL aşarsa, adım 5.4.3 ve 5.4.6 orantılı olarak eklenen boncuk seviyesini ayarlamak.
Not: güç parantez içinde havuzlu PCR reaksiyonları hacmi tam olarak 200 µL ise geçerlidir. - 0.55 x (110 µL) hacimleri Ampure XP boncuk süspansiyon ve yukarı ve aşağı en az 10 kez pipetting tarafından karıştırın.
- 5 dk, 5 dk için manyetik bir stand ayrı boncuk için oda sıcaklığında kuluçkaya.
- Süpernatant için yeni bir tüp taşıyın. Boncuk içeren tüp atmak. Boncuk DNA bağladıktan > 700 bp, sıralı için çok büyük.
- Süpernatant için 0.2 x birimleri (40 µL, 0,75 x Ampure arabellek örnekteki toplam sonuçlanan) eklemek Ampure XP boncuk süspansiyon ve aşağı yukarı 10 kez pipetting tarafından mix.
- 5 dk, 5 dk için manyetik bir stand ayrı boncuk için oda sıcaklığında kuluçkaya.
- DNA içeren süpernatant atmak < 200 ücretsiz astar, astar dimer ve çok küçük parçaları içeren sıralı için bp,.
- Tüp manyetik kürsüye bırakın. Boncuk yıkayın, 700 µL % 80 etanol eklemek için dikkat çekici boncuk Pelet rahatsız eder ve 30 için kuluçkaya için s.
- Süpernatant atmak, sonra tüp manyetik stand off al ve boncuk 10 mM Tris-Cl pH 8.0 100 µL içinde resuspend. Aşağı yukarı 10 kez pipetting tarafından mix ve 1 dk. için oda sıcaklığında kuluçkaya.
- 0.8 x (80 µL) hacimleri Ampure XP boncuk süspansiyon ekleyin. Aşağı yukarı 10 kez pipetting tarafından mix ve 5 min için oda sıcaklığında kuluçkaya. Alt sınır boyutu seçimi bu ikinci turunda son Kütüphane astar ve astar dimer tamamen ücretsiz sağlanır.
- Boncuk 5 min için manyetik bir stand ayırmak ve süpernatant atın.
- Boncuk Pelet 700 µL % 80 etanol için 30 ile iki kez yıkayın s her yukarıdaki gibi manyetik standında tüp bırakarak.
- Tüp ile hala manyetik stand, etanol bütün izlerini kaldırmak. Bu damlacıkları etanol P10 pipet kullanarak tüp dışarı itmek için yardımcı olur. Kalan etanol için en fazla 5 dk buharlaşmasına izin.
- Tüp manyetik stand off al ve boncuk 10 mM Tris-Cl pH 8.0 50 µL içinde resuspend. Aşağı yukarı 10 kez pipetting karıştırın.
- 5 dk sonra ayrı boncuk manyetik bir stand için oda sıcaklığında kuluçkaya.
- Transfer süpernatant temiz bir tüp. Bu son Hi-C Kütüphane, sayısal ve sonraki nesil sıralama makinelerde, üreticinin yönergelerine göre sıralı hazır olduğunu.
Sonuçlar
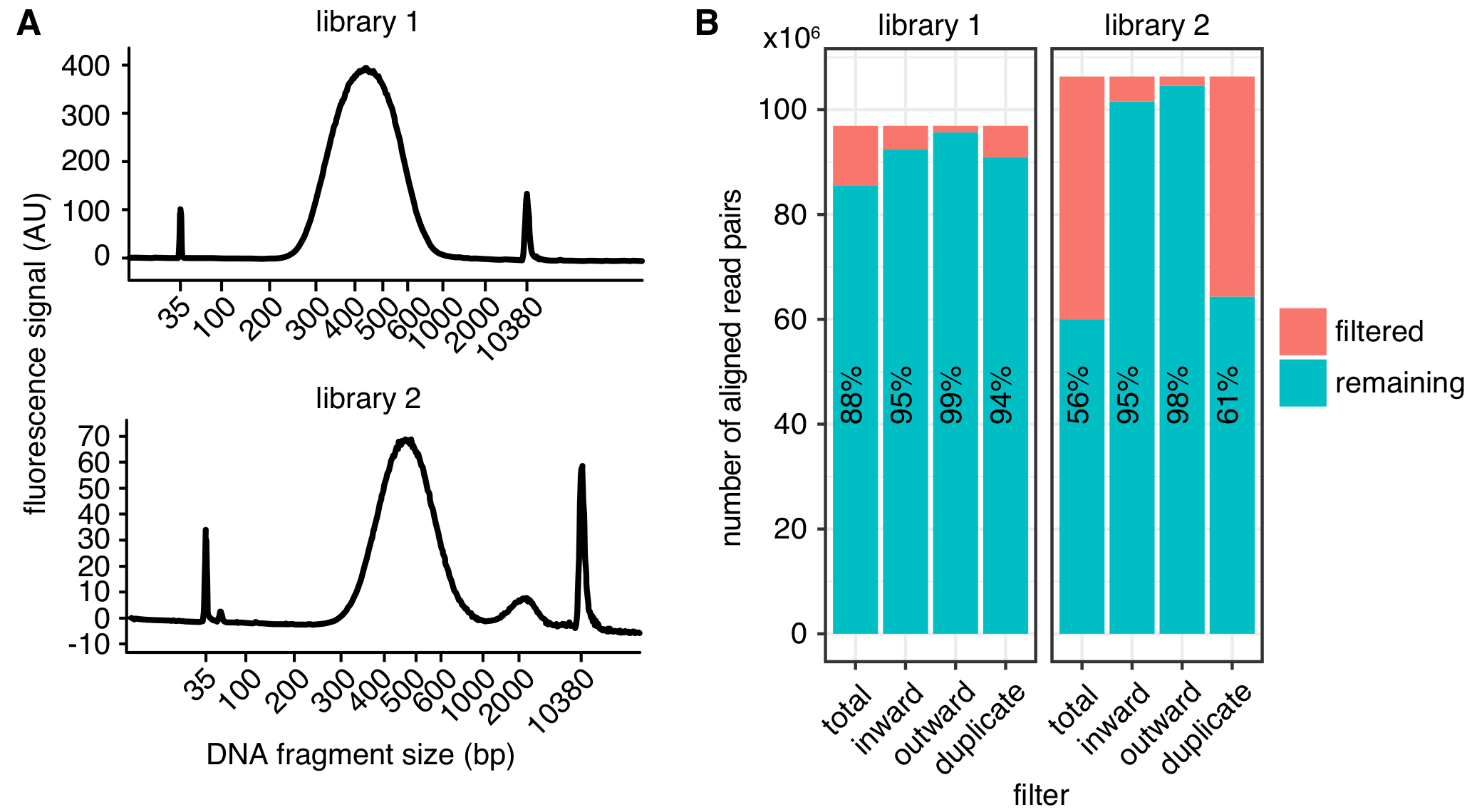
Embriyo nüfus nükleer döngüsü 12, 13 ve 14 sıralı (1: 30'a karşılık gelen 1:45 ve 2:10 saatleri sonrası döllenme, sırasıyla12) ve iletişim kuralında tanımlanan yordamlara göre belirleme yapılacağı elde edilen 3-4 h sonrası döllenme (hpf). Her sıralanmış embriyo toplu eGFP-PCNA sinyal fotoğraflarını alarak, aşağı akım deneylerde kullanılan her tek embriyo kesin sahne ve hücre döngüsü durumunu belgelemek mümkündür. Örnek resimler sıralanmış nüfus üzerinden embriyo Şekil 1B-Egösterilir. Situ Hi-C iletişim kuralı çıkışını sonraki nesil sıralama makinelerde sıralı hazır bir nükleotit kütüphanesidir. Bu amaçla, en az 2-4 nM son Kütüphane konsantrasyonu genellikle gereklidir. Giriş malzeme önerilen miktarda kullanarak, bu güvenilir bir şekilde bölgedir (Tablo 1)elde.
Boyut seçimi arasında yaklaşık 500 bp bağlı olarak tam yamultma (2A rakam), ve boyut seçimi parametreleri maksimum 300-600 bp sonra DNA parçalarının beklenen boyut dağılımı. Sıralama için genom içinde eşlenemez kısıtlama parçaları sayısını en aza indirmek için en az 75 bp uzunluğu eşleştirilmiş uç okuma öneririz. Yüksek çözünürlüklü haritalar 1 – 2 kb kutusu boyutu ile 400 milyon Okunma elde edilebilir. ~ 150 milyon daha düşük derinlikte birden çok biyolojik çoğaltır sıralama sıralama çok yüksek derinlikte bir tek REPLICATE yerine her okur tavsiye ediyoruz. Bu değerlendirme biyolojik çeşitlilik sağlar ve daha küçük bir numarayla PCR çoğaltma nedeniyle atılan okur yol açar. Görsel temsili için çoğaltır birleştirilebilir. Bir örnek yüksek derinlikte sıralama için taahhütte bulunmadan önce Şekil 2Bolduğu gibi temel Kütüphane kalite parametreleri belirlemek için sığ sıralama (örnek başına bir kaç milyon defa okundu) kullanarak örnekleri çalışan öneririz.
Hi-C veri analizi önemli Hesaplamalı kaynakları ve Biyoinformatik uzmanlık gerektirir. Kaba bir genel olarak eşleştirilmiş okuma bağımsız olarak başvuru genom eşleştirilir, elde edilen hizalamaları kalite ve yönlendirme için filtre uygulanır sonra bir matris kişilerin belirli depo gözü çözünürlük veya parça düzeyinde oluşturulan süzülmüş gelen hizalamaları. İletişim matris daha da aşağı akım Analizi TADs, döngüler ve bölmeleri keşfetmek için temelidir. Sıralama okuma ilk analiz için birkaç Biyoinformatik boru hatları kullanılabilir çok özel Biyoinformatik bilgi18,19, olmadan iletişim matrisler içine ham okuma işleme Etkinleştir 20,21,22,23. Daha fazla çözümleme yapılır nasıl out tam biyolojik soru altında eğitim büyük ölçüde bağlıdır ve programlama ve R veya Python komut dosyası önemli deneyim gerektirebilir. Ancak, mevcut5,24,25,26,27,28gibi yazılım analiz etmek için çeşitli araçlar ve algoritmaları TADs aramak için vardır ve Hi-C veri içinde örümcek ağı kaş ve tek başına masaüstü uygulamaları29,30,31,32olarak keşfedin.
Bir kez işlenir, Kütüphane kalitesi farklı ölçümleri (Şekil 2B) kullanarak belirlenebilir. İlk olarak, aynı özgün molekül doğan sıralı okuma çiftlerinin sayısı olan PCR çoğaltmaları, oranı boşa sıra okuma miktarını sınırlamak mümkün olduğunca düşük olmalıdır. Ancak, kütüphaneleri ile bile > % 40 PCR çoğaltma işleme yüksek kaliteli iletişim haritalar yinelenenleri filtre durumunda. İkinci olarak, yönelimleri, nedeniyle filtre uygulanmış okuma oranı4' te açıklandığı gibi sürekli olarak hizalanmış okuma çiftlerinin % 10 düşük olmalıdır.
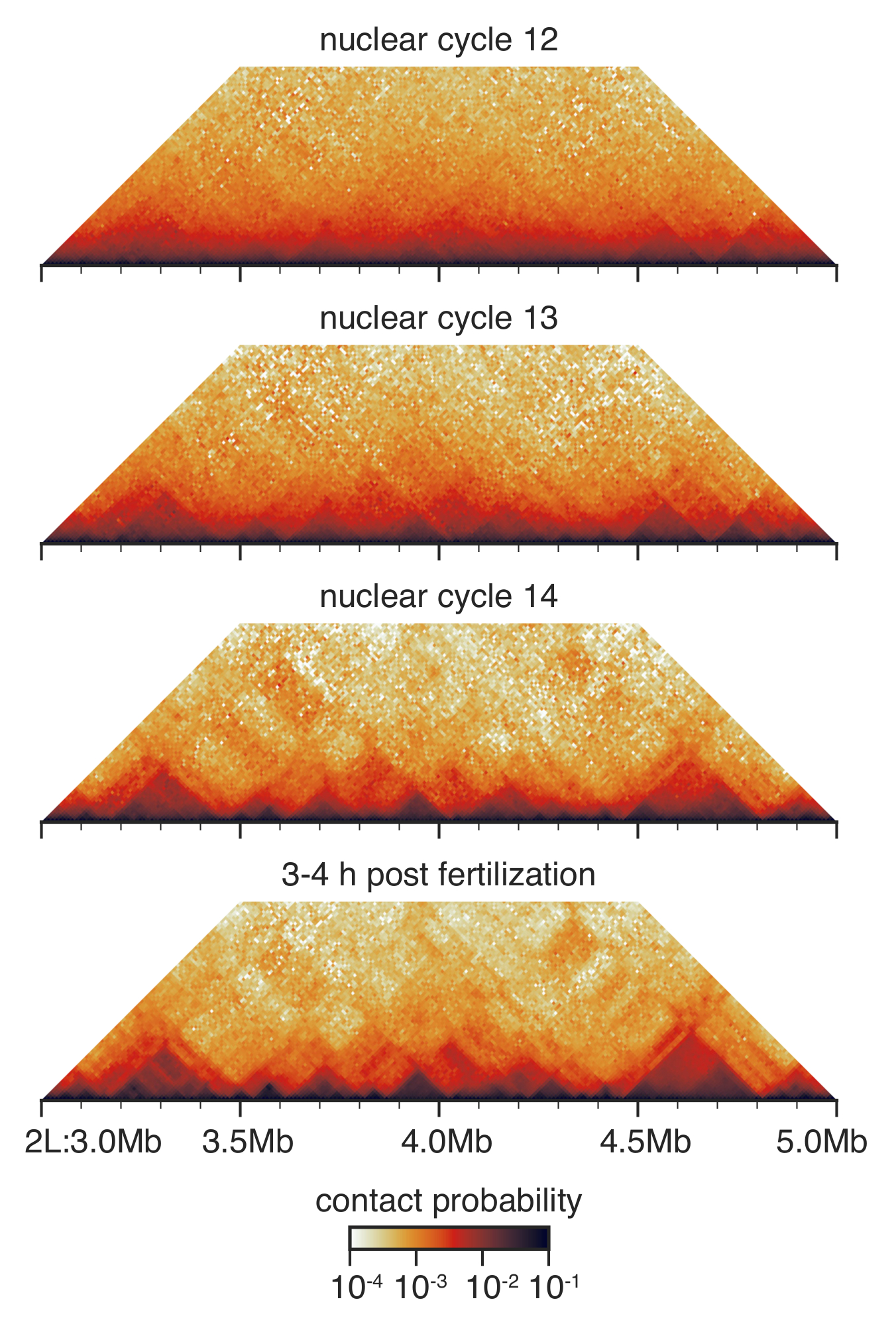
Drosophila nükleer döngüsü 12 ve 14 arasında önceden gastrular geliştirme sırasında büyük ölçüde yenilenmiş33 (Şekil 3) nükleer bir mimaridir. Nükleer döngüsü 12, kaç TADs algılanır ve kişilerin genel dağıtım birçok discernable özellikleri olmadan çok düzgün. Bu önemli ölçüde nükleer döngüsü 13 ve 14, TADs giderek daha belirgindir ve belirsiz uzun menzilli kişiler tükenmiş değiştirilir.

Şekil 1: sıralama sırasında eGFP-PCNA embriyo temsilcisi resimleri. (A)eGFP-PCNA sinyal 60 dk toplama ve nükleer döngüsü 12 (B), nükleer embriyo 25 ° C (B-E) örneklerden sıralanmış nüfus itibariyle 2 h kuluçka 13 (C), nükleer döngüsü 14 (döngüsü sonra embriyo sıralanmamış bir popülasyondan D) ve zaman uyumlu mitoz (E) geçiren embriyo. Ölçek çubukları 200 µm. = Bu rakam daha büyük bir versiyonunu görüntülemek için buraya tıklayınız.
{kind=link}

Resim 2: in situ Hi-C Kitaplığı kalite ölçümleri örnekleri. DNA parçası boyutları başarılı bir Hi-C Library (kütüphane 1, en iyi) ve parçaları (Library 2, alt) sıralama için çok büyük bir zirve görüntüler bir kitaplıktan dağılımını gösteren(a)Bioanalyzer izlemeler. Kütüphane 2 başarıyla sıralı, ancak daha büyük miktarlarda istenmeyen DNA parçalarının azalan sıralama verimleri için neden olabilir. (B) iki Hi-C Kitaplığı istatistiklerinin filtreleme: görüntülenen okuma yön ve mesafe (içe, dışa doğru)4 veya PCR çoğaltma (yinelenen) nedeniyle daha fazla analiz bırakılır hizalanmış okuma çiftleri sayısıdır. Her barda (kalan) filtre geçen defa okundu ve başarısız (filtre) çizilir. Filtre geçen okuma yüzdesi Ayrıca metin olarak gösterilir. Bu rakam daha büyük bir versiyonunu görüntülemek için buraya tıklayınız.
{kind=link}

Şekil 3: Merhaba-C etkileşim haritaları hazırlanmış embriyolar. Merhaba C etkileşim haritalar 10 kb çözünürlükte binned ve33daha önce açıklandığı gibi dengeli. Kromozom 2 bir bölge gösterilir L. Bu rakam daha büyük bir versiyonunu görüntülemek için buraya tıklayınız.
{kind=link}
| Kütüphane | Sahne | Embriyo sayısı | (Ng) kesme önce DNA miktarı | PCR döngüleri | Son Kütüphane konsantrasyonu (nM) |
| 1 | Nükleer döngüsü 12 | 71 | 46 | 12 | 28.2 |
| 2 | Nükleer döngüsü 12 | 46 | 40 | 12 | 22,2 |
| 3 | Nükleer döngüsü 12 | 60 | 13 | 13 | 12,3 |
| 4 | Nükleer döngüsü 13 | 36 | 39 | 12 | 22,2 |
| 5 | Nükleer döngüsü 13 | 35 | 10 | 12 | 5.0 |
| 6 | Nükleer döngüsü 13 | 48 | 18 | 12 | 8,7 |
| 7 | Nükleer döngüsü 14 | 33 | 30 | 12 | 39.8 |
| 8 | Nükleer döngüsü 14 | 24 | 36 | 12 | 20,4 |
| 9 | Nükleer döngüsü 14 | 14 | 8 | 12 | 4.2 |
| 10 | 3-4 hpf | 17 | 30 | 12 | 24.0 |
| 11 | 3-4 hpf | 18 | 42 | 11 | 19,1 |
| 12 | 3-4 hpf | 22 | 63 | 11 | 48,4 |
Tablo 1: temsilcisi sıralama kitaplığı istatistik listesi. Her kitaplık listesinde, onun üretme, biotin açılan ve qubit'e tarafından ölçülen kesme önce toplam DNA miktarı için kullanılan embriyo sayısı için PCR güçlendirme ve sıralama kitaplığı son konsantrasyonu için kullanılan döngüleri sayısı Arıtma ve boyutu seçimden sonra gösterilir.
Tartışmalar
Burada sunulan yüksek kaliteli haritalar kromatin mimarisinin erken Drosophila embriyo çok daha etkili kuralıdır. Bir önceki iletişim kuralı34için karşılaştırıldığında, açıklanan yaklaşım burada daha yüksek çözünürlük, daha hızlı işleme ve daha az reaktif kullanımı kaynaklanan bir güncel in situ Hi-C yordamı5, kullanır. Situ Hi-C protokolü de dahil olmak üzere genel yordam çok çeşitli aşamaları ve Drosophilayanı sıra deneysel sistemleri üzerinde çalışmak için bekleniyor. Protokol düşük giriş zorunlu olduğundan, izole hücre popülasyonlarının üzerinde de kullanılabilir. Drosophilaiçinde Aralık dışında embriyo için iletişim kuralını kullanarak, bazı parametreler tarif ederken özellikle malzeme, fiksasyonu ayarlanması etmeniz gerekebilir. Yana son derece geçirimsiz bir manikür büyük embriyo geliştirmek, formaldehit konsantrasyonu yükseltmek ve fiksasyon uzatan uygun olabilir. Embriyo nükleer döngüsü 14 farklı aşamalarında koleksiyonu için 25 ° c adım 1.4 embriyo kuluçka zamanlarında aşağıdaki gibi ayarlanması gerekir: nükleer döngüsü 12, 70 dk; Nükleer döngüsü 13, 90 dk; 3-4 hpf, 3:30 s.
13 göğüs arası bölümler (sahne 1-4), sırasında çekirdek yoğunluğu kabaca her bölümü ile iki katına çıkar. Çekirdeklerin kolayca onların parlak GFP floresans tarafından tespit edilebilir. Mitoz sırasında eGFP-PCNA çekirdeğinde yer ve onun sinyal embriyo dağınık olduğunu. Bu özellik bir zaman uyumlu bölünme bölünme mümkün geçiren tanımlayıcı embriyo yapar. Kromatin mitotik organizasyonu Interphase organizasyon35büyük ölçüde farklı olduğu için Kromatin uyum eğitimi için mitotik Bu embriyolar genellikle, istenmez. Özellikle zaman uyumlu bir mitotik bölünme geçirecek embriyo seçmek için iletişim kuralı uyum sağlamak mümkündür. Bu durumda, eGFP-PCNA dağınık, nükleer olmayan dağılımı ile tek embriyo tutulmalıdır ve diğer embriyo atılmalıdır. Nükleer yoğunluğu tespit edilemez beri sahne embriyo tarafından iletilen ışık mikroskobu izlendi onların morfolojisi için alternatif yöntemler istihdam gerekir. Kutup hücreleri ve hücre çekirdeği embriyo çevre varlığı çevre görünür cellularization nükleer döngüsü 1412gösterir, ancak gösterir embriyo en az nükleer döngüsü 9, tamamladı.
Merhaba C deneyler başarıyla enzimleri5geniş bir seçki kullanılarak gerçekleştirilebilir. Geçerli yaklaşımlar genellikle ya bir 4-base sırası, MboI veya HindIII gibi bir 6-base tanıma sitesi gibi tanımak enzimler kullanılır. 4-base kesiciler avantajı 6-base kesiciler yeterli sıralama derinlik ve kısıtlama sitelerin daha bile kapsama genom verilen yüksek potansiyel çözünürlük sundukları var. Başka bir5,23,36,37bir 4-base kesici seçiminde hiçbir avantaj olduğunu. İki en yaygın olarak kullanılan enzimler, MboI ve DpnII, her ikisi de aynı GATC tanıma sitesi tanıdım. DpnII ilgilendirmiyor DrosophilaCpG metilasyonu daha az duyarlıdır. Burada sunulan protokolü de başarılı bir şekilde DpnII Restriksiyon enzimi kullanılarak tamamlanabilir. 4.2 bölümünde. Restriksiyon enzimi ve tampon DpnII uyumluluk için ayarlanacak üreticinin önerilerini göre var.
Şekil 2Agösterildiği aralığından sıralama kitaplığı parçası boyutunu önemli ölçüde sapma sergiliyor, küme oluşumu sırasında sıralama daha az verimli veya tamamen başarısız. Bu durumda, yamultma sonra boyutu dağıtım kontrol edilmelidir ve parametreleri yamultma buna göre ayarlanır. DNA parçalarının çok küçük dağılımında tepeler (< 100 bp) veya çok büyük (> 1000 bp) boyutları gösterir taşımak örneğin boyut seçimi ile ilgili sorunlar üzerine boncuk veya atılacak beklenen süpernatant. Sık sık bu kütüphanelerin bu istenmeyen büyüklüktedir, hala bir resimde, olan gibi küçük tepeler ile başarıyla verimliliği kümeleme içinde sadece küçük bir düşüş ile sıralı.
Kullanılabilir sıra okuma sayısı büyük ölçüde azalttığı yüksek oranda PCR tekrarından kaçınılmalıdır. PCR çoğaltmaları oranı doğrudan giriş malzeme miktarı ile ilgilidir. Daha fazla giriş'i kullanarak bu nedenle genellikle PCR çoğaltma sorunları azaltır.
Okuma okuma yönelimi nedeniyle (2B rakam) filtre daha yüksek sayılar çok az enzim, çok fazla giriş malzeme veya eksik homojenizasyon embriyoların kullanılması sonucunda olabilir yetersiz sindirim gösterir.
Açıklamalar
Yazarlar ifşa gerek yok.
Teşekkürler
Bu araştırma Max Planck toplum tarafından finanse edildi. C.B.H. moleküler Biyomedikal Uluslararası Max Planck Araştırma Okulu – bir bursu tarafından desteklenmiştir. Biz Shelby Blythe ve Eric Wieschaus nazik eGFP-PCNA Drosophila melanogaster satır verdiğiniz için teşekkür ederiz.
Malzemeler
| Name | Company | Catalog Number | Comments |
| Biotin-14-dATP | Life Technologies | 19524016 | |
| MboI | New England Biolabs | R0147L | |
| DNA Polymerase I Klenow Fragment | New England Biolabs | M0210L | |
| T4 DNA Ligase | Thermo Fisher | EL0012 | T4 DNA Ligase Buffer included |
| T4 DNA Polymerase | New England Biolabs | M0203L | |
| Proteinase K | AppliChem | A4392 | |
| GlycoBlue | Life Technologies | AM9516 | |
| Complete Ultra EDTA-free protease inhibitors | Roche | 5892791001 | |
| NEBNext Multiplex Oligos for Illumina (Index Primers Set 1) | New England Biolabs | E7335 | Sequencing Adaptor, Forward (unindexed) PCR primer and Reverse (indexed) PCR primer and USER enzyme used in the Library preparation section are components of this kit |
| NEBNext Ultra II DNA Library Prep Kit | New England Biolabs | E7645 | End Prep Enzyme Mix, End Prep Reaction Buffer, Ligation Enhancer, Ligation Master Mix and Polymerase Master Mix used in the Library preparation section are components of this kit |
| Covaris S2 AFA System | Covaris | ||
| DNA LoBind Tubes, 1.5 mL | Eppendorf | 0030108051 | |
| Falcon cell strainer 100 µm | Corning | 352360 | Embryo collection baskets |
| 37% formaldehyde | VWR | 437536C | |
| Heptane | AppliChem | 122062.1612 | |
| M165 FC fluorescent stereo microscope | Leica | ||
| M165 FC DFC camera | Leica | ||
| Metal micro pestle | Carl Roth | P985.1 | Used to lyse embryos in step 4.1.4 |
| RNase A | AppliChem | A3832,0050 | |
| Dynabeads MyOne Streptavidin C1 | Life Technologies | 65002 | Streptavidin coated magnetic beads |
| Ampure XP beads | Beckman Coulter | A63881 | |
| Qubit 3.0 Fluorometer | Thermo Fisher Scientific | Q33216 | |
| Qubit assay tubes | Thermo Fisher Scientific | Q32856 | |
| Qubit dsDNA HS Assay Kit | Thermo Fisher Scientific | Q32854 | |
| Phosphate buffered saline (PBS) | Sigma-Aldrich | P4417 | |
| eGFP-PCNA flies | Gift from S. Blythe and E. Wieschaus | ||
| Sodium hypochlorite 13% | Thermo Fisher | AC219255000 | |
| Triton X-100 | AppliChem | A4975 | |
| Tris buffer pH 8.0 (1 M) for molecular biology | AppliChem | A4577 | |
| NaCl | AppliChem | A2942 | |
| IGEPAL CA-630 | Sigma-Aldrich | I8896 | |
| 1.5 mL microcentrifuge tubes | Greiner Bio-One | 616201 | |
| SDS for molecular biology | AppliChem | A2263 | |
| 10x CutSmart buffer | New England Biolabs | B7204S | Restriction enzyme buffer |
| PCR Nucleotide Mix | Sigma-Aldrich | 11814362001 | Unmodified dCTP, dGTP, dTTP |
| BSA, Molecular Biology Grade | New England Biolabs | B9000S | |
| EDTA 0.5 M solution for molecular biology | AppliChem | A4892 | |
| Sodium acetate 3 M pH 5.2 | Sigma-Aldrich | S7899 | |
| DynaMag-2 Magnet | Life Technologies | 12321D | Magnetic stand |
| Intelli-Mixer RM-2L | Omnilab | 5729802 | Rotator |
| ThermoMixer F1.5 | Eppendorf | 5384000012 | Mixer |
| Small Embryo Collection Cages | Flystuff.com | 59-100 | Egg collection cage |
| Centrifuge 5424 R | Eppendorf | 5404000413 | |
| C1000 Touch Thermal Cycler | Bio-Rad | 1851148 | |
| PCR tube strips | Greiner Bio-One | 673275 | |
| NEBuffer 2.1 | New England Biolabs | B7202S | T4 DNA Polymerase buffer |
Referanslar
- Bonev, B., Cavalli, G. Organization and function of the 3D genome. Nat Rev Genet. 17 (11), 661-678 (2016).
- Lieberman-Aiden, E., et al. Comprehensive mapping of long-range interactions reveals folding principles of the human genome. Science. 326 (5950), 289-293 (2009).
- Dixon, J. R., et al. Topological domains in mammalian genomes identified by analysis of chromatin interactions. Nature. 485 (7398), 376-380 (2012).
- Jin, F., et al. A high-resolution map of the three-dimensional chromatin interactome in human cells. Nature. , (2013).
- Rao, S. S. P., et al. A 3D Map of the Human Genome at Kilobase Resolution Reveals Principles of Chromatin Looping. Cell. 159 (7), 1665-1680 (2014).
- Darbellay, F., Duboule, D. Topological Domains, Metagenes, and the Emergence of Pleiotropic Regulations at Hox Loci. Current topics in developmental biology. 116, 299-314 (2016).
- Beagan, J. A., et al. Local Genome Topology Can Exhibit an Incompletely Rewired 3D-Folding State during Somatic Cell Reprogramming. Cell stem cell. 18 (5), 611-624 (2016).
- Andrey, G., et al. Characterization of hundreds of regulatory landscapes in developing limbs reveals two regimes of chromatin folding. Genome Res. 27 (2), 223-233 (2017).
- Krijger, P. H. L., de Laat, W. Regulation of disease-associated gene expression in the 3D genome. Nature Reviews. Molecular Cell Biology. 17 (12), 771-782 (2016).
- Sexton, T., et al. Three-dimensional folding and functional organization principles of the Drosophila genome. Cell. 148 (3), 458-472 (2012).
- Ghavi-Helm, Y., et al. Enhancer loops appear stable during development and are associated with paused polymerase. Nature. 512 (7512), 96-100 (2014).
- Foe, V. E., Alberts, B. M. Studies of nuclear and cytoplasmic behaviour during the five mitotic cycles that precede gastrulation in Drosophila embryogenesis. J Cell Sci. 61, 31-70 (1983).
- Blythe, S. A., Wieschaus, E. F. Zygotic Genome Activation Triggers the DNA Replication Checkpoint at the Midblastula Transition. Cell. 160 (6), 1169-1181 (2015).
- Blythe, S. A., Wieschaus, E. F. Establishment and maintenance of heritable chromatin structure during early Drosophila embryogenesis. eLife. 5, e20148(2016).
- JoVE Science Education Database. Embryo and Larva Harvesting and Preparation. Biology I: yeast, Drosophila and C. elegans. Drosophila melanogaster. , JoVE, Cambridge, MA. (2017).
- Sicaeros, B., O'Dowd, D. K. Preparation of Neuronal Cultures from Midgastrula Stage Drosophila Embryos. Journal of Visualized Experiments. (5), (2007).
- Shermoen, A. W. Preparation of Baskets for Drosophila Egg Collections, Treatments, and Incubations. Cold Spring Harbor Protocols. (10), (2008).
- Ay, F., Noble, W. S. Analysis methods for studying the 3D architecture of the genome. Genome biology. 16 (1), 183(2015).
- Lazaris, C., Kelly, S., Ntziachristos, P., Aifantis, I., Tsirigos, A. HiC-bench: comprehensive and reproducible Hi-C data analysis designed for parameter exploration and benchmarking. BMC Genomics. 18 (1), (2017).
- Servant, N., et al. HiC-Pro: an optimized and flexible pipeline for Hi-C data processing. Genome Biology. 16 (1), (2015).
- Durand, N. C., et al. Juicer Provides a One-Click System for Analyzing Loop-Resolution Hi-C Experiments. Cell systems. 3 (1), 95-98 (2016).
- Lajoie, B. R., Dekker, J., Kaplan, N. The Hitchhiker's guide to Hi-C analysis: Practical guidelines. Methods. 72, 65-75 (2015).
- Schmitt, A. D., Hu, M., Ren, B. Genome-wide mapping and analysis of chromosome architecture. Nature Reviews. Molecular Cell Biology. 17 (12), 743-755 (2016).
- Shin, H., et al. TopDom: an efficient and deterministic method for identifying topological domains in genomes. Nucleic Acids Res. 44 (7), e70(2016).
- Kruse, K., Hug, C. B., Hernández-Rodríguez, B., Vaquerizas, J. M. TADtool: visual parameter identification for TAD-calling algorithms. Bioinformatics. 32 (20), 3190-3192 (2016).
- Lévy-Leduc, C., Delattre, M., Mary-Huard, T., Robin, S. Two-dimensional segmentation for analyzing Hi-C data. Bioinformatics. 30 (17), Oxford, England. i386-i392 (2014).
- Filippova, D., Patro, R., Duggal, G., Kingsford, C. Identification of alternative topological domains in chromatin. Algorithms for molecular biology: AMB. 9 (1), 14(2014).
- Crane, E., et al. Condensin-driven remodelling of X chromosome topology during dosage compensation. Nature. 523 (7559), 240-244 (2015).
- Durand, N. C., et al. Juicebox Provides a Visualization System for Hi-C Contact Maps with Unlimited Zoom. Cell systems. 3 (1), 99-101 (2016).
- Zhou, X., et al. Exploring long-range genome interactions using the WashU Epigenome Browser. Nature Methods. 10 (5), 375-376 (2013).
- Ramírez, F., et al. High-resolution TADs reveal DNA sequences underlying genome organization in flies. bioRxiv. , 115063(2017).
- Kerpedjiev, P., et al. HiGlass: Web-based Visual Comparison And Exploration Of Genome Interaction Maps. bioRxiv. , 121889(2017).
- Hug, C. B., Grimaldi, A. G., Kruse, K., Vaquerizas, J. M. Chromatin Architecture Emerges during Zygotic Genome Activation Independent of Transcription. Cell. 169 (2), (2017).
- Berkum, N. L., et al. Hi-C: a method to study the three-dimensional architecture of genomes. Journal of Visualized Experiments: JoVE. (39), (2010).
- Naumova, N., et al. Organization of the mitotic chromosome. Science. 342 (6161), 948-953 (2013).
- Denker, A., de Laat, W. The second decade of 3C technologies: detailed insights into nuclear organization. Genes & development. 30 (12), 1357-1382 (2016).
- Belaghzal, H., Dekker, J., Gibcus, J. H. Hi-C 2.0: An optimized Hi-C procedure for high-resolution genome-wide mapping of chromosome conformation. Methods (San Diego, Calif). 123, 56-65 (2017).
Yeniden Basımlar ve İzinler
Bu JoVE makalesinin metnini veya resimlerini yeniden kullanma izni talebi
Izin talebiThis article has been published
Video Coming Soon
JoVE Hakkında
Telif Hakkı © 2020 MyJove Corporation. Tüm hakları saklıdır