
Method Article
Génération de chromatine Genome-wide Conformation Capture bibliothèques d’embryons de drosophile début étroitement mise en scène
Dans cet article
Résumé
Cet ouvrage décrit un protocole pour la génération de haute résolution en situ CIH bibliothèques d’étroitement mis en scène la gastrulation Drosophila melanogaster embryons.
Résumé
Étudier l’architecture tridimensionnelle de la chromatine offre un aperçu précieux sur les mécanismes de régulation génique. Nous décrivons ici un protocole permettant d’effectuer la chromatine conformation capture technique in situ CIH sur mise en scène de populations d’embryon de drosophile . Le résultat est une bibliothèque de séquençage qui permet la cartographie de toutes les interactions de la chromatine qui se produisent dans le noyau en une seule expérience. Tri des embryons se fait manuellement à l’aide d’un microscope stéréo fluorescent et une ligne mouche transgénique contenant un marqueur nucléaire. En utilisant cette technique, les populations d’embryon de chaque cycle de division nucléaire et possédant un statut défini cycle cellulaire, peut être obtenue avec une très grande pureté. Le protocole peut également être adapté pour trier les embryons âgés au-delà de gastrulation. Trié les embryons sont utilisés comme entrées pour in situ Hi-C. Toutes les expériences, y compris le séquençage de préparation de la bibliothèque, peuvent être complétées en cinq jours. Le protocole a de faibles exigences d’entrée et fonctionne de manière fiable à l’aide de 20 embryons au stade blastoderme comme matières. Le résultat final est une bibliothèque de séquençage pour le séquençage de la génération suivante. Après le séquençage, les données peuvent être transformées en cartes d’interaction de chromatine Génome-large qui peuvent être analysés à l’aide d’un large éventail d’outils disponibles pour obtenir des informations sur l’Association topologiquement structure du domaine (TAD), des boucles de la chromatine et la chromatine compartiments au cours du développement de la drosophile .
Introduction
Capture de conformation de la chromatine (3C) est devenue une méthode particulièrement utile pour étudier la topologie de la chromatine dans le noyau1. La variante 3C Hi-C permet de mesurer les fréquences de contact de toutes les interactions de la chromatine qui se produisent dans le noyau en une seule expérience2. Application du CIH a joué un rôle important dans la découverte et la caractérisation de nombreux principes fondamentaux d’organisation de la chromatine, comme dat, compartiments et boucles3,4,5.
Études d’architecture de la chromatine dans le contexte des transitions du développement et de différenciation cellulaire sont de plus en plus utilisés à élucider les mécanismes de régulation des gènes au cours de ces processus6,7,8, 9. Parmi les organismes modèles d’un grand intérêt est Drosophila melanogaster, dont développement et génome sont bien caractérisés. Cependant, peu d’études qui examinent l’architecture de la chromatine dans la drosophile en dehors de l’in vitro culture tissulaire, les paramètres ont été réalisées10,11. Chez les embryons, 16 – 18 h après la fécondation, DAT et compartiments qui rappelle des structures similaires chez les mammifères ont été identifiés10, qui soulève la question de quel rôle ils jouent dans la régulation des gènes au cours de l’embryon de drosophile développement. Surtout dans les premiers stades de développement, avant la gastrulation, ces études sont techniquement difficiles. Avant la gastrulation, des embryons de drosophile subissent 13 divisions nucléaires synchrones qui procèdent à un rythme extrêmement rapid de 8 – 60 min par cycle12,13. En outre, l’absence de caractéristiques visuelles de distinguer les différentes étapes le rendent difficile obtenir un embryon solidement mises en scène matériel en quantité suffisante.
Afin d’élaborer un protocole qui permet d’étudier l’architecture de la chromatine dans le développement précoce de drosophile à résolution du cycle nucléaire, nous avons combiné les deux techniques existantes : in situ Hi-C, qui permet à toute la génération des images haute résolution génome contact cartes5et mise en scène d’embryon à l’aide d’une ligne de Drosophila transgénique exprimant eGFP-PCNA transgène13,14. Ce transgène se localise dans le noyau pendant l’interphase et se disperse dans le blastoderme syncytial au cours de la mitose. À l’aide de cette propriété, il est possible de distinguer facilement les différentes étapes de leur densité nucléaire et embryons mitotiques de la dispersion du signal GFP.
Ensemble, ces techniques permettent l’étude de la structure tridimensionnelle de la chromatine en haute résolution à partir d’aussi peu que 20 embryons de drosophile . Ce protocole comprend les instructions pour la récolte et le tri des embryons de drosophile pour obtenir les populations d’embryons d’un cycle de division nucléaire unique. Il décrit également comment les embryons obtenus sont utilisés pour effectuer sur place Hi-C. Le résultat final est une bibliothèque de nucléotides appropriée pour l’ordonnancement sur machines de séquençage prochaines génération. Les lectures de séquençage qui en résultent peuvent alors être traités dans des cartes d’interaction chromatine détaillées couvrant tout le génome de la drosophile .
Protocole
1. collecte d’embryons de drosophile
Remarque : Une collecte d’embryons équivalent peut être effectuée comme indiqué dans une précédente publication15.
- Transférer les jeunes eGFP-PCNA mouches (< 1 semaine vieux) dans des cages collection oeuf avec une collection plaques16 (1 % éthanol, acide acétique à 1 % et 4 % d’agar).
- Déplacer les cages de la collection dans un incubateur à 25 ° C. Une incubation de 1 à 2 jours avant la collecte des œufs améliore de manière significative le rendement de l’oeuf. Changer les plaques collectrices deux fois par jour.
- Enlever les plaques contenant des embryons de la chambre de collecte en intervalles de 30 à 60 min. Des intervalles entraîner dans des embryons de moins, mais plus stricte répartition des stades de développement. Recueillir auprès des cages multiples en parallèle alors qu’idéalement > 200 œufs sont pondus à chaque 30 à 60 min.
- Stocker les plaques à 25 ° C jusqu'à ce que les embryons atteignent l’âge désiré. Pour les embryons au stade blastoderme (cycle nucléaire 14), incuber pendant environ 2 h.
- Après 2 h d’incubation, ajouter l’eau du robinet d’un vaporisateur à la plaque de collection afin que la totalité de la surface est recouverte d’eau. Suspendre les embryons et levure à l’aide d’une brosse douce.
- Versez resuspendues embryons de la plaque de la collection dans un bac de ramassage de l’embryon (crépines cellule commerciale avec la taille des pores 100 µm ou paniers maison17 fonctionnent bien), ajoutant de l’eau du robinet supplémentaire d’un vaporisateur, si nécessaire. À ce stade, combiner les embryons des boîtes qui ont été recueillis en parallèle. L’échantillon représente un lot unique.
- Lavez les embryons bien en rinçant le panier avec l’eau du robinet d’un vaporisateur pour 30 s jusqu'à ce que tous les résidus de levure sont emporté.
- Dechorionate embryons en plaçant le bac de ramassage dans une solution d’hypochlorite de sodium à 2,5 % dans l’eau. Agitation légère en agitant facilite l’élimination du chorion. Continuer jusqu'à ce que les embryons soient suffisamment hydrophobes afin qu’elles flottent à la surface de la solution lorsque le panier est soulevé et immergé à nouveau, qui devrait prendre la ~1.75–2 min.
ATTENTION : l’hypochlorite de Sodium est corrosif. Porter un équipement de protection individuelle approprié. Solutions contenant < 10 % d’hypochlorite de sodium peuvent généralement être éliminés dans l’évier, vérifiez le règlement de l’Institut de l’hôte. - Retirer le panier de la solution et rincer abondamment à l’eau du robinet d’un vaporisateur, jusqu'à ce que l’odeur de l’eau de Javel n’est plus perceptible.
2. Fixation de l’embryon
Remarque : Des conditions de fixation optimale, principalement la concentration de détergent, de formaldéhyde et de la durée de fixation, doivent être déterminées empiriquement pour s’adapter à la scène des embryons. Pour les étapes autour du blastoderme syncytial, une concentration finale de 0,5 % Triton X-100 et 1,8 % de formaldéhyde dans la phase aqueuse fonctionnent bien. Pour les étapes ultérieures au stade embryon 9, outre l’optimisation de ces paramètres peut être nécessaire. Toutes les solutions utilisées lors de la fixation et le tri doivent contenir des antiprotéases.
- Inverser le bac de ramassage et placez-le sur un tube de centrifugation conique de 15 mL. Rincer les embryons du panier dans le tube à l’aide d’une pipette Pasteur distribution PBS-T (PBS, 0,5 % Triton X-100).
- Laissez les embryons s’installer en bas et régler le volume total de 2 mL avec du PBS-T.
- Ajouter 6 mL d’heptane et 100 µL de 37 % de formaldéhyde dans l’eau.
ATTENTION : Heptane et le formaldéhyde sont toxiques lorsqu’ils sont inhalés ou après peau contacter. Porter un équipement de protection individuelle approprié et travailler sous une hotte. Déchets contenant de l’heptane ou formaldéhyde doit être traité séparément, conformément aux règlements de l’Institut de l’hôte. - Après l’addition de la formaldéhyde, démarrer une minuterie de 15 min et agiter vigoureusement le tube monte et descend pendant 1 min à la main. La phase aqueuse et organique combinera à consistance forme un shampooing-like.
- Agiter sur un mélangeur rotatoire jusqu'à 10 min après l’ajout de formaldéhyde.
- Centrifuger à 500 g pendant 1 min à température ambiante pour recueillir les embryons au fond du tube.
- Aspirer le liquide comme shampooing entière et jetez-le, prenant soin de ne pas pour aspirer les embryons. Petites quantités restantes de surnageant comme shampooing ne causent pas de problèmes.
- 15 min après l’ajout de formaldéhyde, remettre en suspension les embryons dans 5 mL de PBS-T avec glycine de 125 mM pour étancher le formaldéhyde. Mélanger vigoureusement en agitant de haut en bas pendant 1 min.
- Centrifuger à 500 g à température ambiante pendant 1 min et aspirer le surnageant.
- Lavez les embryons par eux resuspendant dans 5 mL de PBS glacee-T. Laissez les embryons settle et aspirer tout surnageant.
- Répéter le lavage à l’étape 2.10 deux fois plus.
- Garder les embryons sur la glace jusqu’au tri. Habituellement, c’est une bonne idée de recueillir 3 – 4 lots d’embryons mouches avant de procéder au tri. Cependant, les embryons doivent être triés le même jour. Un stockage prolongé sur la glace ou dans le réfrigérateur conduit à morphologie altérée embryon.
3. embryon tri
NOTE : Tri peut être fait sur n’importe quel microscope stéréo fluorescent équipé d’un filtre GFP au grossissement X 60 – 80.
- À l’aide d’une pipette 1 000 µL, transférer un lot d’environ 100 embryons dans un récipient de petit verre approprié pour le tri, de préférence d’une couleur foncée et placez-le sur la glace.
-
Embryons de tri par statut nucléaire densité et cycle cellulaire (Figure 1) en poussant les embryons souhaitables dans un tas séparé à l’aide d’une pointe d’aiguille ou une seringue.
- Supprimer tous les embryons avec distribution dispersée, non-nucléaire d’eGFP-PCNA (Figure 1E). Aussi, les embryons que partiellement montrent un signal GFP non nucléaires doivent être supprimés.
- Pour faciliter le tri, assembler un line-up d’embryons de référence au cycle nucléaire 12, 13 et 14 dans chaque lot en utilisant comme guide les photos dans la Figure 1 . Utiliser ce line-up pour faire correspondre les embryons d’un stade inconnu avec l’un des embryons référence afin de déterminer leur stade.
- Pour vérifier le stade de développement pour les embryons de référence, mesurer la densité nucléaire par imagerie de l’embryon et compter le nombre de noyaux à la surface de l’embryon dans une superficie de 2 500 µm2 à l’aide de logiciels d’imagerie qui fournit des informations à distance.
Remarque : Le nombre de noyaux pour une superficie de 2 500 µm2 est 12 à 16 noyaux au cycle nucléaire 12 et 20 à 30 noyaux au cycle nucléaire 1313.
- Une fois que tous les embryons au stade approprié sont séparées, prendre des photos des embryons de documentation et de contrôle de la qualité. Si le microscope stéréo n’est pas lui-même muni d’un module de caméra, un microscope à épifluorescence avec GFP filtres peut être utilisé.
- Pipetter vers le haut les embryons désirées en utilisant une pipette 1 000 µL, transfert vers un nouveau tube et le lieu sur glace.
- Continuez jusqu'à ce que suffisamment embryons sont triées pour l’expérience prévue. Pour les embryons âgés que de l’étape 9, généralement 20 embryons sont suffisants pour un in situ l’expérience Hi-C. Au cycle nucléaire 12, 80 embryons sont un bon point de départ. Lors des cycles précédents, le nombre d’embryons doit être pratiquement doublé pour chaque cycle.
- Piscine et split embryons dans des tubes de 1,5 mL de telle sorte qu’un tube contienne suffisamment embryons pour un seul in situ CIH experiment. Il est conseillé d’utiliser des tubes avec les caractéristiques de la liaison ADN faibles, puisque le même tube sera utilisé pour l’ensemble du protocole et l’adsorption de l’ADN peut conduire à des pertes importantes à de faibles concentrations d’ADN.
- Tourner les tubes brièvement à 100 g à la température ambiante et éliminer le surnageant. Les embryons devraient être aussi sèches que possible pour la congélation.
- Flash congeler les embryons en immergeant les tubes dans l’azote liquide et conserver à-80 ° C.
4. in Situ Salut-C
- Lyse
- Placer les tubes avec des embryons congelés sur la glace.
- Remettre en suspension les embryons dans 500 µL de tampon de lyse glacé (10 mM Tris-Cl pH 8,0, 10 mM NaCl, 0,2 % IGEPAL CA-630, les inhibiteurs de la protéase ; dissous dans l’eau). Ensuite, attendre 1 min pour laisser les embryons à s’installer au fond du tube.
- Embryons de mouture à l’aide d’un pilon micro métal, préalablement refroidi sur la glace, qui est conçu pour s’adapter étroitement à un tube de microtubes de 1,5 mL.
- Pour éviter d’agiter les embryons, insérer le pilon lentement jusqu'à ce qu’il touche le fond du tube, poussez vers le bas et puis moudre en tournant le pilon deux fois dans les deux sens.
- Soulever le pilon très légèrement, pousser jusqu’au fond du tube à nouveau et répéter meulage.
- 4.1.3.2 le répète 10 fois, ou jusqu'à ce que les embryons sont complètement lysées. La solution doit être homogène et sans gros morceaux résiduels d’embryons doit rester.
- Incuber la suspension homogénéisée sur la glace pendant 15 min. Spin à 1 000 x g, 4 ° C pendant 5 min et jeter le surnageant.
- Laver les granulés par resuspendant dans 500 µL de tampon de lyse glacee, pipetage de haut en bas.
- Tourner de nouveau comme 4.1.4 et jeter le surnageant.
- Resuspendre lavé culot dans 100 µL de 0,5 % dodécylsulfate de sodium (SDS), pipetage de haut en bas. Permeabilize noyaux en incubant pendant 10 min à 65 ° C dans un bloc chauffant. Étancher SDS en ajoutant 50 µL de 10 % Triton X-100 et 120 µL d’eau. Mélanger en effleurant le tube.
- Incuber à 37 ° C pendant 15 min dans le bloc chauffant.
- Enzyme de restriction digestion
- Ajouter 25 µL d’enzyme de restriction mémoire tampon 10 x et 20 U de 5 U/µL MboI. Mélanger en effleurant le tube.
- Digérer l’ADN en incubant pendant 90 min à 37 ° C dans le bloc chauffant sous agitation légère (750 tr/min).
- Ajouter un autre 20 U de MboI et continuer d’incubation pendant 90 min.
- Chaleur-inactiver MboI en incubant à 62 ° C pendant 20 min.
- Porte-à-faux Fill-in
NOTE : Remplissant le porte-à-faux avec biotinylé dATP permet de sélectionner des fragments ligaturés spécifiques. Biotine-dATP aux intersections de la ligature est protégé contre l’activité exonucléase de T4 ADN polymérase (section 4.6), tandis que la biotine-dATP à extrémités franches unligated est éliminée efficacement. Le menu déroulant utilisant streptavidine perles au point 4.7 donc spécifiquement enrichit de fragments d’ADN ligaturés, chimériques.- Ajouter 18 µL de 0,4 mM biotine-14-dATP, 2,25 µL d’un mélange de dCTP/dGTP/dTTP non modifié (3,3 mM) et 8 µL de 5 U/µL ADN polymérase I Fragment de Klenow.
- Mélanger en effleurant le tube et laisser incuber à 37 ° C pendant 90 min en bloc chauffant.
- LIGATURE
- Ajouter 657 µL d’eau, 120 µL de 10 x T4 DNA Ligase tampon, 100 µL de 10 % de X-100 Triton, 6 µL de 20 mg/mL d’albumine sérique bovine (BSA) et mélanger en effleurant le tube. Enfin ajouter 5 µL de 5 U/µL T4 DNA Ligase et mélanger en effleurant le tube.
- Tourner tube doucement (20 t/mn) à température ambiante pendant 2 h.
- Ajouter une deuxième tranche de 5 µL de 5 U/µL T4 DNA Ligase et continuer à tourner pour 2 h de plus.
- Tournez en bas des noyaux à 2 500 g pendant 5 min et jeter le surnageant.
- Extraction d’ADN
- Resuspendre le culot dans 500 µL de tampon d’extraction (50 mM Tris-Cl pH 8.0, 50 mM NaCl, 1 mM acide tétraacétique (EDTA), 1 % SDS ; dissous dans l’eau) et ajouter 20 µL de protéinase K. Mix de la 20 mg/mL en effleurant le tube.
- Digérer les protéines en incubant à 55 ° C pendant 30 min, secouant à 1 000 tr/min.
- De-crosslink, ajouter 130 µL de 5 M de NaCl et incuber une nuit à 68 ° C, secouant à 1 000 tr/min.
- Échantillon de la pipette dans un nouveau tube de 2 mL, préférentiellement avec faibles caractéristiques de liaison de l’ADN.
- Ajouter 0,1 x volumes (63 µL) de 3M sodium acétate pH 5,2 et 2 µL de 15 mg/mL GlycoBlue. Bien mélanger en retournant. Ajouter 1,6 x volumes (1 008 µL) d’éthanol absolu pur et mélanger par retournement.
- Incuber à-80 ° C pendant 15 min. Centrifuger à 20 000 x g à 4 ° C pendant au moins 30 min. Le culot d’ADN est souvent très faible, presque invisible et peut seulement être repéré en raison de la couleur bleue des GlycoBlue.
- Retirez surnageant très soigneusement, quittant l’embout de la pipette dans le tube le long du mur opposé où se trouve le granule d’ADN. Petites gouttelettes restantes sont souvent faciles à enlever au cours de cette étape et les lavages suivants en les poussant hors les tubes à l’aide d’une pointe de P10 plutôt que les pipetage.
- Pastille de lavage en ajoutant 800 µL d’éthanol 70 %. Mélanger en retournant et centrifuger à 20 000 x g à température ambiante pendant 5 min. répéter ce lavage au moins une fois.
- Éliminer toute trace d’éthanol et laissez le reposer tube lorsque le couvercle est ouvert jusqu'à 5 min à sécher à l’air. Une fois qu’aucun liquide n’est restant, ajouter 50 µL de pH 10 mM Tris-Cl 8.0. Pipette à plusieurs reprises la solution sur la zone sur la paroi du tube où se trouvait le culot de solubiliser l’ADN.
- Ajouter 1 µL de 20 mg/mL RNase A, mélangez en effleurant le tube et incuber à 37 ° C pendant 15 min à digérer RNA. L’échantillon peut maintenant être stocké dans le réfrigérateur pendant la nuit ou congelé à-20 ° C indéfiniment.
- Vérifier la concentration de l’ADN à un dosage de colorant fluorescent basé selon les instructions du fabricant. Le montant total de l’ADN dans l’échantillon doit être au moins 10 ng, sinon trop peu de matériel est disponible pour l’amplification et complexité de bibliothèque sera probablement faible. Lorsque cela se produit, la quantité de matériel de départ n’était probablement pas suffisante, ou matériel a été perdu en chemin, peut-être lors de la lyse et de précipitations.
- Enlèvement de la biotine et le cisaillement d’ADN
- Ajouter ensemble 12 µL de tampon de T4 ADN polymérase x 10, 3 µL de dATP 1 mM, 3 µL de 1 mM dGTP et 46 µL d’eau. Mélanger en effleurant le tube. Ajouter 5 µL de 3 U/mL T4 ADN polymérase, mélanger en effleurant le tube et laisser incuber à 20 ° C pendant 30 min.
- Ajouter 3 µL de 0,5 M EDTA pour arrêter la réaction et utiliser l’eau pour amener l’échantillon à un volume d’environ 120 µL.
- Cisaillement de l’ADN d’une taille de 200 – 400 bp à l’aide d’un dispositif de sonication conformément aux instructions du fabricant. Le sonicateur mentionné dans la Table des matières, le programme suivant est approprié : 2 cycles de chacun des 50 s, le droit de 10 %, intensité 5, 200 cycles/rafale.
- Menu déroulant biotine
- Distribuer 30 µL de billes magnétiques de streptavidine enduite 10 mg/mL dans un nouveau tube, séparez chacune d’elles sur un support magnétique et jeter le surnageant.
- Remettre en suspension les perles dans 1 x tampon B & W (5 mM Tris-Cl pH 7,4, EDTA de 0,5 mM, 1 M NaCl ; dissous dans l’eau) + 0,1 % Triton X-100 et mélanger au vortex. Placer le tube sur un support magnétique et attendez 1 – 5 min jusqu'à ce que les perles sont séparées, selon la marque et le modèle.
- Aspirer et éliminer le liquide surnageant tout en faisant glisser l’embout de la pipette le long du mur opposé où se trouvent les perles. Remettre en suspension les perles dans 120 µL de 2 x B & W un tampon (10 mM Tris-Cl pH 7,4, EDTA 1 mM et 2 M de NaCl). Mélanger au vortex.
- Transfert d’ADN cisaillé dans un nouveau tube de liaison ADN faible et mélanger avec 120 µL de la suspension de la perle dans un tampon 2 X B & W au vortex. Faire pivoter les perles avec l’échantillon d’ADN à 20 tr/min pendant 15 min.
- Séparer les perles sur un support magnétique et jeter le surnageant.
- Remettre perles de Triton X-100 dans 600 µL de 1 x B & W + 0,1 % et incuber à 55 ° C pendant 2 min, secouant à 1 000 tr/min. Après la séparation, éliminer le surnageant. Répéter une fois ce lavage.
- Laver les perles une fois avec 600 µL de pH 10 mM Tris-Cl 8.0 et éliminer le surnageant après la séparation.
- Remettre en suspension les perles dans 50 µL de pH 10 mM Tris-Cl 8.0.
5. séquençage bibliothèque préparation
Remarque : Toutes les étapes de bibliothèque sont faites en utilisant des composants d’une préparation commerciale de bibliothèque ADN nécessaire (voir la Table des matières). Toutefois, les kits de rechange ou d’autres peuvent être substitués. Précipitation tend à se former dans les agents de préparation de bibliothèque au cours de la conservation au congélateur. Il est donc important de s’assurer que toutes les précipitations se dissout avant d’utiliser des réactifs.
-
Réparation de fin
- Transférer la suspension de perle dans 50 µL de 10 mM Tris-Cl pH 8.0 dans un nouveau tube de PCR.
- Ajouter 3 µL du mélange de préparation enzymatique de fin et 7 µL de tampon de réaction prép de fin. La composition de pipetage de haut en bas.
- Tube de transfert à un thermocycleur et exécuter le programme suivant : 20 ° C pendant 30 min, 65 ° C pendant 30 min et maintenez à 4 ° C.
-
Ligature de l’adaptateur
- Ajouter 30 µL de ligature Master Mix, 2,5 µL de 1,5 µM séquençage adaptateur (diluer à 1,5 µM de stock) et 1 µL de renforceur de ligature à la suspension de la perle. La composition de pipetage de haut en bas.
- Incuber à 20 ° C pendant 15 min dans un thermocycleur.
- Ajouter 3 µL d’enzyme de l’utilisateur. La composition de pipetage de haut en bas.
- Incuber à 37 ° C pendant 15 min dans un thermocycleur.
- Séparer les perles sur un support magnétique et éliminer le surnageant.
- Aux perles de lavage, de remettre en suspension les perles dans 100 µL de 1 x tampon B & W + 0,1 % X-100 Triton. Mix de vortex et le transfert dans un nouveau tube de microcentrifuge. Séparer les perles sur un support magnétique et éliminer le surnageant.
- Répétez ce lavage une fois à l’aide de 600 µL de tampon de la même.
- Remettre les billes en suspension dans 600 µL de pH 10 mM Tris-Cl 8.0, mélanger au vortex et transférer des perles dans un nouveau tube.
- Séparer les perles sur un support magnétique, jeter le surnageant et remettre les billes en suspension dans 50 µL de 10 mM Tris-Cl pH 8.0.
-
Amplification par PCR
- Préparer deux tubes PCR et dans chacun, mélanger 25 µL du mélange réactionnel de la polymérase, 1,5 µL de 10 µM avant amorce PCR (non indexés) et 1,5 µL de 10 µM inverse amorce PCR (indexé).
Remarque : En avant (non indexé) amorce PCR :
5'-AATGATACGGCGACCACCGAGATCTACACTCTTTCCCTACACGACGCTCTTCCGATC * T-3´.
Amorce PCR (indexée) inverse :
5'-CAAGCAGAAGACGGCATACGAGATNNNNNNGTGACTGGAGTTCAGACGTGTGCTCTTCCGATC * T-3´. * indique les obligations phosphorothioate et Ns dans l’amorce PCR indexée. - Dans chaque tube, ajouter 22 µL de suspension de perles et mélanger par pipetage de haut en bas.
- Exécutez la PCR en utilisant le programme suivant : 98 ° C pendant 1 min, (98 ° C pendant 15 s, 65 ° C pendant 75 s, rampe de 1,5 ° C/s) répété 9 - 12 fois, 65 ° C pendant 5 min et maintenez à 4 ° C.
Remarque : Le nombre de cycles d’amplification doit être déterminée empiriquement. Cependant, nous avons trouvé que les bibliothèques nécessitant plus de 12 cycles étaient généralement de faible complexité et n’entraîne pas de haute qualité que HI-C cartes. En revanche, les bibliothèques nécessitant moins de 12 cycles ne sont pas affectées négativement en amplifiant pendant 12 cycles. Par conséquent, il est possible de par défaut à 12 cycles d’amplification. - Mettre en commun les deux réactions de PCR dans un tube de microcentrifuge unique, séparer les perles sur un support magnétique et transférer le surnageant contenant la bibliothèque dans un nouveau tube.
- Préparer deux tubes PCR et dans chacun, mélanger 25 µL du mélange réactionnel de la polymérase, 1,5 µL de 10 µM avant amorce PCR (non indexés) et 1,5 µL de 10 µM inverse amorce PCR (indexé).
-
Sélection de la taille
- Mettre XP Ampure suspension de perle à température ambiante et mélanger bien en secouant.
- Porter le volume de la réaction PCR regroupée à exactement 200 µL d’eau. Au cours de la PCR et la séparation magnétique, certains du volume initial est habituellement perdue. Vérifiez le volume en définissant la pipette à 200 µL et aspirer la totalité du volume de la réaction. Si l’air est aspiré, plus l’eau doit être ajouté. Si le volume dépasse 200 µL, régler le volume des perles ajoutées aux étapes 5.4.3 et 5.4.6 proportionnellement.
Remarque : Les volumes entre parenthèses sont valables si le volume total des réactions PCR commun est exactement 200 µL. - Ajouter 0,55 x volumes (110 µL) de suspension de perle de Ampure XP et mélanger en pipettant également, haut en bas, au moins 10 fois.
- Incuber à température ambiante pendant 5 min, séparé de perles sur un support magnétique pendant 5 min.
- Surnageant de déménagement dans un nouveau tube. Jeter le tube contenant les billes. Les perles ont lié l’ADN > 700 bp, ce qui est trop volumineux pour être séquencé.
- Pour le surnageant, ajouter 0,2 x volumes (40 µL, soit un total de 0.75 x Ampure tampon non contrôlé dans l’échantillon) de suspension de perle de Ampure XP et mélanger en pipettant également monter et descendre de 10 fois.
- Incuber à température ambiante pendant 5 min, séparé de perles sur un support magnétique pendant 5 min.
- Jeter le surnageant qui contient de l’ADN < 200 bp, qui comprend des amorces gratuits, dimères d’amorce et de fragments trop petits pour être séquencé.
- Maintenir le tube sur le support magnétique. Perles de lavage, ajouter 700 µL d’éthanol à 80 %, en prenant soin de ne pas déranger le culot de perle et incuber pendant 30 s.
- Jeter le surnageant, puis prenez le tuyau sur le support magnétique et remettre les billes en suspension dans 100 µL de pH 10 mM Tris-Cl 8.0. Mélanger en pipettant également, haut en bas 10 fois et incuber à température ambiante pendant 1 min.
- Ajouter 0,8 x volumes (80 µL) de suspension de perle de Ampure XP. Mélanger en pipettant également monter et descendre de 10 fois et incuber à température ambiante pendant 5 min. Cette deuxième ronde de sélection de la taille limite inférieure s’assure que la bibliothèque finale est complètement libre des amorces et des dimères d’amorce.
- Séparer les perles sur un support magnétique pendant 5 min et jeter le surnageant.
- Laver le culot de perle deux fois avec 700 µL d’éthanol à 80 % pendant 30 s chacune, tout en laissant le tube sur le support magnétique, comme indiqué ci-dessus.
- Avec le tube encore sur le support magnétique, éliminer toute trace de l’éthanol. Il permet de pousser les gouttelettes d’éthanol hors du tube à l’aide d’une pipette de P10. Laissez l’éthanol résiduel s’évaporer pendant un maximum de 5 min.
- Prendre le métro sur le support magnétique et remettre les billes en suspension dans 50 µL de pH 10 mM Tris-Cl 8.0. La composition de pipetage et descendre de 10 fois.
- Incuber à température ambiante pendant 5 min, puis séparé de perles sur un support magnétique.
- Transférer surnageant dans un nouveau tube. Il s’agit de la bibliothèque de CIH finale, prête à être quantifié et séquencés sur machines de séquençage prochaines génération, selon les instructions du fabricant.
Résultats
Tri des populations de l’embryon au cycle nucléaire, 12, 13 et 14 (correspondant à 01:30, 01:45 et 02:10 heures après la fécondation, respectivement12) et 3 à 4 h après la fécondation (hpf) ont été obtenus selon les procédures décrites dans le protocole. En prenant des photos du signal eGFP-PCNA de chaque lot d’embryon trié, il est possible de documenter le statut exact de scène et cycle cellulaire de chaque embryon unique qui est utilisé dans des expériences en aval. Photos d’exemple d’embryons provenant de populations triées sont indiquées dans la Figure 1 b-E. La sortie de la protocole de CIH in situ est une bibliothèque de nucléotides prête à séquencer sur des machines de séquençage de génération suivante. À cet effet, une concentration finale de bibliothèque d’au moins 2 à 4 nM est habituellement exigée. En utilisant les apports recommandés de matières entrantes, cette concentration est fiable obtenus (tableau 1).
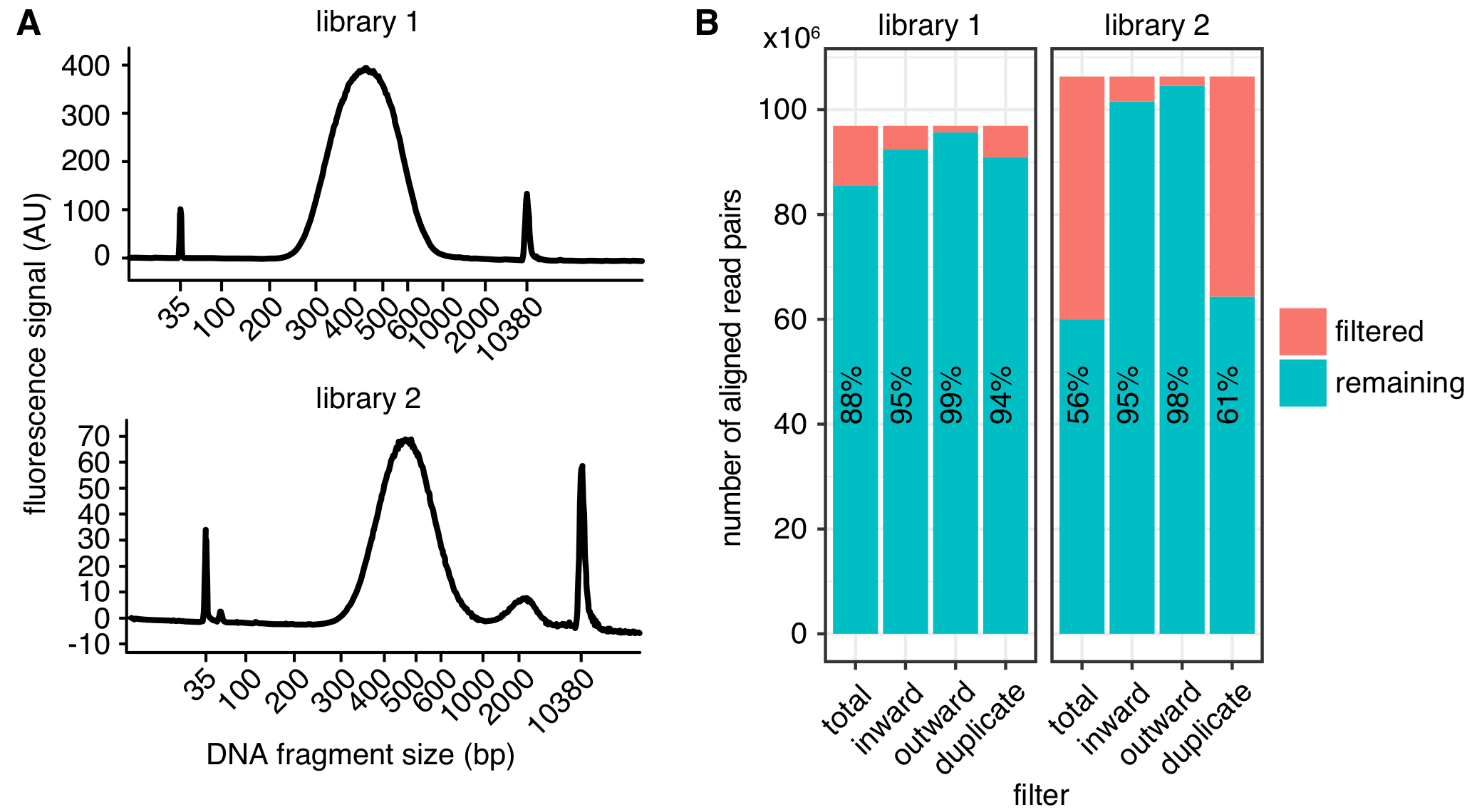
La distribution de la taille attendue de fragments d’ADN après que sélection de la taille est entre 300 à 600 ans bp, avec un maximum à environ 500 bp (Figure 2 a),) selon la tonte exacte et les paramètres de sélection de taille. Pour le séquençage, nous vous recommandons de lectures jumelé-fin d’au moins 75 longueur de bp pour réduire le nombre de fragments de restriction non mappable dans le génome. Des cartes à haute résolution avec 1 à 2 Ko bin taille peuvent provenir de 400 millions de lectures. Nous vous recommandons de séquençage des réplicats biologiques multiples à une profondeur inférieure de 150 millions lit chacune, au lieu de séquençage simple reprend à très grande profondeur. Cela permet d’évaluer la variation biologique et conduit à un nombre inférieur de lectures mis au rebut en raison de la duplication de la PCR. Représentation visuelle, les répétitions peuvent être combinées. Avant de s’engager pour le séquençage d’un échantillon à grande profondeur, nous vous recommandons d’exécuter des échantillons à l’aide de séquençage peu profond (quelques millions de lectures par exemple) afin de déterminer les paramètres de qualité de bibliothèque de base comme dans la Figure 2 b.
Analyse des données de la CIH requiert une grande compétence computationnelle de ressources et de la bioinformatique. Comme un aperçu approximatif, les lectures appariés sont mappés indépendamment sur le génome de référence, les alignements qui en résultent sont filtrés pour la qualité et l’orientation, puis une matrice de contacts à un niveau de résolution ou un fragment de donnée bin peut provenir de la filtrée alignements. La matrice de contact est la base de tout plus en aval analyse exploration dat, boucles et compartiments. Pour l’analyse initiale des lectures de séquençage, il existe plusieurs pipelines de bioinformatique qui permettent la transformation de lectures brutes en contact matrices sans beaucoup bioinformatique spécialisée connaissances18,19, 20,21,22,23. Comment une analyse plus approfondie est faite hors dépend en grande partie la question biologique exacte à l’étude et peuvent nécessiter une expérience significative dans la programmation et de script en Python ou en R. Toutefois, plusieurs outils et algorithmes d’appeler dat sont disponibles5,24,25,26,27,28, mais aussi des logiciels pour analyser et Explorez Hi-C données dans le navigateur web et des applications de bureau autonome29,30,31,32.
Une fois traitée, la qualité de la bibliothèque peut être déterminée à l’aide de différents paramètres (Figure 2 b). Tout d’abord, le taux de doublons PCR, qui correspond au nombre de paires de lecture séquencées découlant de la même molécule originale, doit être aussi faible que possible limiter la quantité de séquence gaspillé lectures. Cependant, même avec les bibliothèques > duplication PCR 40 % peut être transformée en qualité cartes contact si les doublons sont filtrés. Deuxièmement, les lectures filtrés en raison de leur orientation, tel que décrit dans4, constamment sera inférieur à 10 % de paires de lecture alignés.
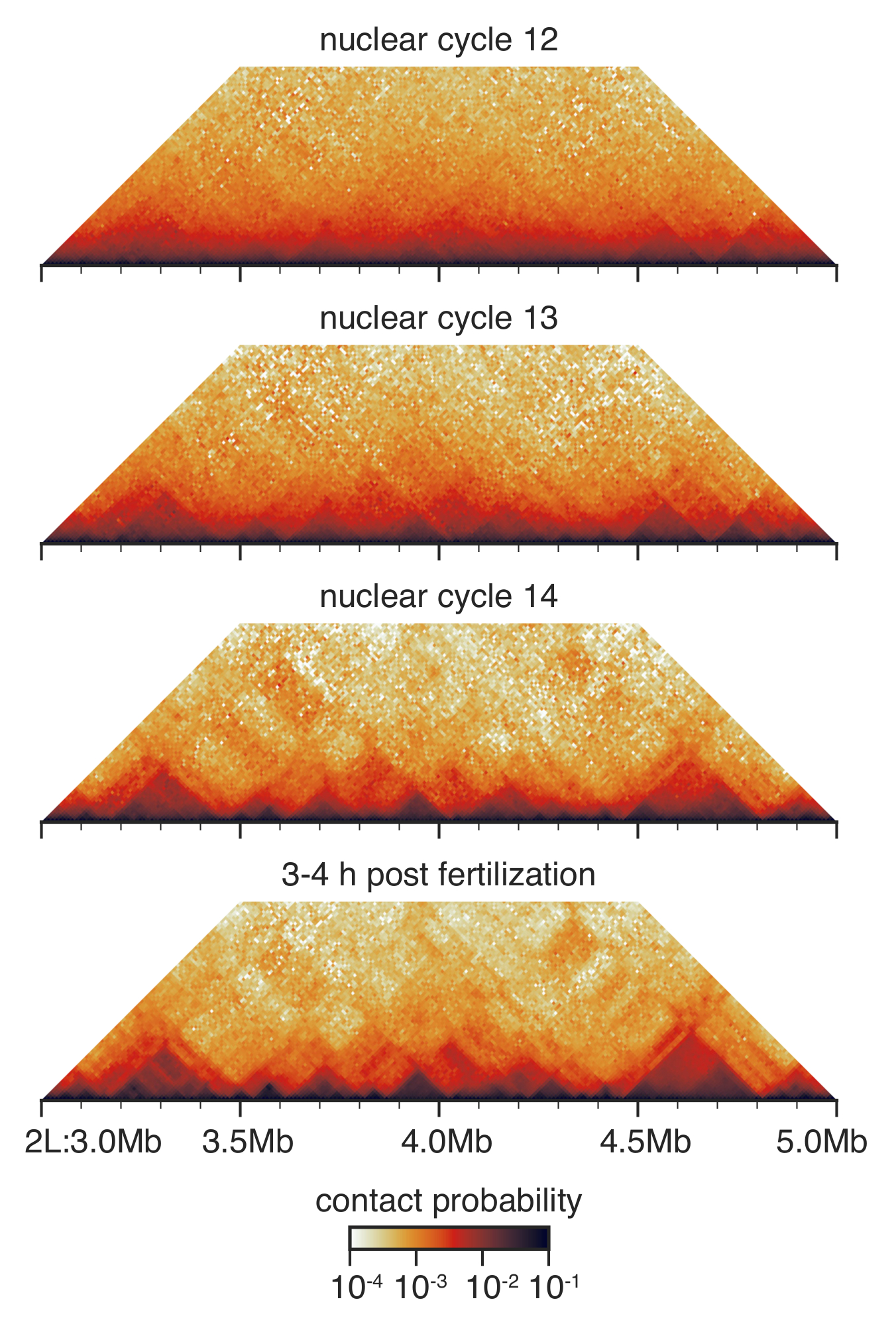
Au cours du développement pré gastrula de drosophile entre cycle nucléaire 12 et 14, l’architecture nucléaire est radicalement rénové33 (Figure 3). Au cycle nucléaire 12, quelques dat est détectés et la répartition générale des contacts est très lisse sans les nombreuses fonctionnalités discernables. C’est radicalement changé au cycle nucléaire 13 et 14, lorsque dat sont toujours plus importants et non-spécifique contacts à longue distance sont épuisés.

Figure 1 : images représentant des embryons eGFP-PCNA pendant tri. Signal eGFP-PCNA (A) d’une population non trié d’embryons après que collecte de 60 min et 2 h d’incubation à 25 ° C (B-E) exemples d’embryons provenant de populations triées au cycle nucléaire 12 (B), nuclear cycle 13 (C), (14) cycle nucléaire ( D) et des embryons en cours de mitose synchrone (E). Barreaux de l’échelle = 200 µm. s’il vous plaît cliquez ici pour visionner une version agrandie de cette figure.
{kind=link}

Figure 2 : Exemples de mesures de qualité in situ CIH bibliothèque. (A) Bioanalyzer traces montrant la distribution des tailles de fragment d’ADN auprès d’une bibliothèque de CIH réussie (bibliothèque 1, haut de la page) et d’une bibliothèque qui affiche un pic de fragments qui sont trop grands pour le séquençage (bibliothèque 2, bottom). Bibliothèque 2 a été séquencé avec succès, mais encore de plus grandes quantités de fragments d’ADN non désirés peuvent mener à une diminution de séquençage des rendements. (B) filtrage statistique des deux bibliothèques de CIH : affichée correspond au nombre de paires de lecture alignés qui sont exclus de l’analyse en raison de la lecture l’orientation et la distance (vers l’intérieur et l’extérieur)4 ou duplication de PCR (en double). Dans chaque bar, le nombre de lectures en passant le filtre (restant) et échec (filtré) est tracé. Le pourcentage de lectures en passant le filtre est en outre montré sous forme de texte. S’il vous plaît cliquez ici pour visionner une version agrandie de cette figure.
{kind=link}

Figure 3 : Salut-C interaction cartes embryons mise en scène de. Salut-C interaction cartes sont mis en cellule à résolution de 10KO et équilibrés comme décrit avant33. Montré est une région sur le chromosome 2 L. s’il vous plaît cliquez ici pour visionner une version agrandie de cette figure.
{kind=link}
| Bibliothèque | Stade | Nombre d’embryons | Montant ADN avant le cisaillement (ng) | Cycles PCR | Concentration finale de bibliothèque (nM) |
| 1 | cycle nucléaire 12 | 71 | 46 | 12 | 28.2 |
| 2 | cycle nucléaire 12 | 46 | 40 | 12 | 22.2 |
| 3 | cycle nucléaire 12 | 60 | 13 | 13 | 12.3 |
| 4 | cycle nucléaire 13 | 36 | 39 | 12 | 22.2 |
| 5 | cycle nucléaire 13 | 35 | 10 | 12 | 5.0 |
| 6 | cycle nucléaire 13 | 48 | 18 | 12 | 8.7 |
| 7 | Nuclear cycle 14 | 33 | 30 | 12 | 39,8 |
| 8 | Nuclear cycle 14 | 24 | 36 | 12 | 20,4 |
| 9 | Nuclear cycle 14 | 14 | 8 | 12 | 4.2 |
| 10 | 3-4 hpf | 17 | 30 | 12 | 24,0 |
| 11 | 3-4 hpf | 18 | 42 | 11 | 19.1 |
| 12 | 3-4 hpf | 22 | 63 | 11 | 48.4 |
Tableau 1 : liste des statistiques de bibliothèques représentatives séquençage. Pour chaque bibliothèque dans la liste, le nombre d’embryons qui ont été utilisés pour sa génération, la quantité d’ADN totale avant la biotine pulldown et cisaillement mesurée par Qubit, le nombre de PCR cycles utilisés pour l’amplification et la concentration finale de la bibliothèque de séquençage Après sélection de purification et de taille sont indiqués.
Discussion
Le protocole présenté ici est très efficace pour générer des cartes de qualité de l’architecture de la chromatine dans des embryons de drosophile au début. Par rapport à un précédent protocole34, l’approche décrite ici utilise un jour in situ CIH procédure5, résultant en un traitement plus rapide, une résolution plus élevée et moins son utilisation réactif. La procédure globale, y compris le protocole de CIH in situ est censée travailler sur un large éventail d’étapes et de systèmes expérimentaux en dehors de la drosophile. Étant donné que le protocole a une exigence d’entrée faible, il pourrait également être utilisé sur les populations de cellules isolées. Chez la drosophile, lors de l’utilisation du protocole pour les embryons en dehors de la plage décrite ici, certains paramètres, notamment la fixation de la matière, peut devoir être ajustée. Étant donné que les embryons plus âgés développent une cuticule hautement imperméable, augmentant la concentration de formaldéhyde et de prolonger la fixation peuvent être appropriés. Pour la collecte d’embryons aux stades autres que nucléaire cycle 14, les temps d’incubation des embryons à 25 ° C à l’étape 1.4 devant être modifiées comme suit : cycle nucléaire 12, 70 min ; cycle nucléaire 13, 90 min ; 3 – 4 hpf, 03:30 h.
Au cours des divisions de 13 clivage (étape 1-4), la densité des noyaux double à peu près à chaque division. Les noyaux peuvent facilement être identifiés par leur fluorescence GFP brillante. Au cours de la mitose, eGFP-PCNA ne se trouve pas dans le noyau, et son signal est dispersé tout au long de l’embryon. Cette caractéristique rend identification des embryons qui font l’objet d’une division de clivage synchrone possible. Pour étudier la conformation de la chromatine, ces embryons mitotiques ne sont généralement pas souhaitables, puisque l’organisation mitotique de la chromatine est radicalement différente de l’interphase organisation35. Il est possible d’adapter le protocole afin de sélectionner les embryons subissant une division mitotique synchrone. Dans ce cas, seuls les embryons avec distribution dispersée, non-nucléaire d’eGFP-PCNA doivent être conservés, et tous les autres embryons devraient être jetées. Puisque la densité nucléaire ne peut pas être déterminée, les méthodes alternatives d’embryons au stade de leur morphologie lus en microscopie photonique transmise doivent être employés. Présence de noyaux à la périphérie de l’embryon et les cellules de mât indiquent que l’embryon a complété au moins nucléaire cycle 9, alors que la cellularisation visible à la périphérie indique cycle nucléaire 1412.
Salut-C expériences peuvent être effectuées avec succès en utilisant une variété d’enzymes de restriction,5. Les approches actuelles utilisent généralement des enzymes qui reconnaissent soit une séquence de 4-base, tels que MboI, ou un site de reconnaissance 6-base, tels que HindIII. L’avantage de fraises 4-base sur base de 6 coupeurs est qu’ils offrent une meilleure résolution potentielle, donnée assez de profondeur séquençage et une couverture plus homogène de sites de restriction au sein du génome. Il n’y a aucun avantage évident en choisissant un couteau 4-base sur un autre5,23,36,37. Les deux enzymes les plus couramment utilisés, MboI et DpnII, tous deux reconnaissent le même site de reconnaissance GATC. DpnII est moins sensible à la méthylation de CpG, qui est d’aucun intérêt chez la drosophile. Le protocole présenté ici peut également être complété avec succès en utilisant DpnII comme une enzyme de restriction. Dans la section 4.2. enzyme de restriction et tampon doivent être ajustés pour la compatibilité DpnII, selon les recommandations du fabricant.
Si la taille du fragment de la bibliothèque de séquençage s’écarte sensiblement de la gamme illustrée à la Figure 2 a, la formation de cluster pendant le séquencement peut s’avérer moins efficace ou échouer complètement. Dans ce cas, la distribution de la taille après la tonte doit être vérifiée et cisaillement paramètres ajustés en conséquence. Pics dans la distribution des fragments d’ADN de très petites (< 100 bp) ou très grande taille (> 1 000 bp) tailles signale des problèmes avec sélection de taille, tels que porter plus de perles ou de surnageant qui sont censés être mis au rebut. Souvent, ces bibliothèques avec des petites pointes à ces tailles indésirables, tels que l’un sur la photo, sont encore séquencé avec succès avec seulement une diminution mineure efficacité de clustering.
Des taux élevés de duplication de PCR doivent être évitées car cela réduit considérablement le nombre de lectures de séquence utilisable. Le taux de PCR doublons est directement lié à la quantité de matières entrantes. En utilisant plus d’entrées donc habituellement atténue les problèmes de duplication de PCR.
Un nombre plus élevé de lectures filtrés en raison de l’orientation de lecture ()Figure 2 b) indique la digestion insuffisante, qui peut être le résultat de l’utilisation trop peu enzyme, trop d’entrée matériau ou incomplète homogénéisation des embryons.
Déclarations de divulgation
Les auteurs n’ont rien à divulguer.
Remerciements
Cette recherche a été financée par la Société Max Planck. C.B.H. a été soutenu par une bourse de l’International Max Planck Research School – biomédecine moléculaire. Nous remercions Shelby Blythe et Eric Wieschaus aimablement la ligne de melanogaster de drosophile eGFP-PCNA.
matériels
| Name | Company | Catalog Number | Comments |
| Biotin-14-dATP | Life Technologies | 19524016 | |
| MboI | New England Biolabs | R0147L | |
| DNA Polymerase I Klenow Fragment | New England Biolabs | M0210L | |
| T4 DNA Ligase | Thermo Fisher | EL0012 | T4 DNA Ligase Buffer included |
| T4 DNA Polymerase | New England Biolabs | M0203L | |
| Proteinase K | AppliChem | A4392 | |
| GlycoBlue | Life Technologies | AM9516 | |
| Complete Ultra EDTA-free protease inhibitors | Roche | 5892791001 | |
| NEBNext Multiplex Oligos for Illumina (Index Primers Set 1) | New England Biolabs | E7335 | Sequencing Adaptor, Forward (unindexed) PCR primer and Reverse (indexed) PCR primer and USER enzyme used in the Library preparation section are components of this kit |
| NEBNext Ultra II DNA Library Prep Kit | New England Biolabs | E7645 | End Prep Enzyme Mix, End Prep Reaction Buffer, Ligation Enhancer, Ligation Master Mix and Polymerase Master Mix used in the Library preparation section are components of this kit |
| Covaris S2 AFA System | Covaris | ||
| DNA LoBind Tubes, 1.5 mL | Eppendorf | 0030108051 | |
| Falcon cell strainer 100 µm | Corning | 352360 | Embryo collection baskets |
| 37% formaldehyde | VWR | 437536C | |
| Heptane | AppliChem | 122062.1612 | |
| M165 FC fluorescent stereo microscope | Leica | ||
| M165 FC DFC camera | Leica | ||
| Metal micro pestle | Carl Roth | P985.1 | Used to lyse embryos in step 4.1.4 |
| RNase A | AppliChem | A3832,0050 | |
| Dynabeads MyOne Streptavidin C1 | Life Technologies | 65002 | Streptavidin coated magnetic beads |
| Ampure XP beads | Beckman Coulter | A63881 | |
| Qubit 3.0 Fluorometer | Thermo Fisher Scientific | Q33216 | |
| Qubit assay tubes | Thermo Fisher Scientific | Q32856 | |
| Qubit dsDNA HS Assay Kit | Thermo Fisher Scientific | Q32854 | |
| Phosphate buffered saline (PBS) | Sigma-Aldrich | P4417 | |
| eGFP-PCNA flies | Gift from S. Blythe and E. Wieschaus | ||
| Sodium hypochlorite 13% | Thermo Fisher | AC219255000 | |
| Triton X-100 | AppliChem | A4975 | |
| Tris buffer pH 8.0 (1 M) for molecular biology | AppliChem | A4577 | |
| NaCl | AppliChem | A2942 | |
| IGEPAL CA-630 | Sigma-Aldrich | I8896 | |
| 1.5 mL microcentrifuge tubes | Greiner Bio-One | 616201 | |
| SDS for molecular biology | AppliChem | A2263 | |
| 10x CutSmart buffer | New England Biolabs | B7204S | Restriction enzyme buffer |
| PCR Nucleotide Mix | Sigma-Aldrich | 11814362001 | Unmodified dCTP, dGTP, dTTP |
| BSA, Molecular Biology Grade | New England Biolabs | B9000S | |
| EDTA 0.5 M solution for molecular biology | AppliChem | A4892 | |
| Sodium acetate 3 M pH 5.2 | Sigma-Aldrich | S7899 | |
| DynaMag-2 Magnet | Life Technologies | 12321D | Magnetic stand |
| Intelli-Mixer RM-2L | Omnilab | 5729802 | Rotator |
| ThermoMixer F1.5 | Eppendorf | 5384000012 | Mixer |
| Small Embryo Collection Cages | Flystuff.com | 59-100 | Egg collection cage |
| Centrifuge 5424 R | Eppendorf | 5404000413 | |
| C1000 Touch Thermal Cycler | Bio-Rad | 1851148 | |
| PCR tube strips | Greiner Bio-One | 673275 | |
| NEBuffer 2.1 | New England Biolabs | B7202S | T4 DNA Polymerase buffer |
Références
- Bonev, B., Cavalli, G. Organization and function of the 3D genome. Nat Rev Genet. 17 (11), 661-678 (2016).
- Lieberman-Aiden, E., et al. Comprehensive mapping of long-range interactions reveals folding principles of the human genome. Science. 326 (5950), 289-293 (2009).
- Dixon, J. R., et al. Topological domains in mammalian genomes identified by analysis of chromatin interactions. Nature. 485 (7398), 376-380 (2012).
- Jin, F., et al. A high-resolution map of the three-dimensional chromatin interactome in human cells. Nature. , (2013).
- Rao, S. S. P., et al. A 3D Map of the Human Genome at Kilobase Resolution Reveals Principles of Chromatin Looping. Cell. 159 (7), 1665-1680 (2014).
- Darbellay, F., Duboule, D. Topological Domains, Metagenes, and the Emergence of Pleiotropic Regulations at Hox Loci. Current topics in developmental biology. 116, 299-314 (2016).
- Beagan, J. A., et al. Local Genome Topology Can Exhibit an Incompletely Rewired 3D-Folding State during Somatic Cell Reprogramming. Cell stem cell. 18 (5), 611-624 (2016).
- Andrey, G., et al. Characterization of hundreds of regulatory landscapes in developing limbs reveals two regimes of chromatin folding. Genome Res. 27 (2), 223-233 (2017).
- Krijger, P. H. L., de Laat, W. Regulation of disease-associated gene expression in the 3D genome. Nature Reviews. Molecular Cell Biology. 17 (12), 771-782 (2016).
- Sexton, T., et al. Three-dimensional folding and functional organization principles of the Drosophila genome. Cell. 148 (3), 458-472 (2012).
- Ghavi-Helm, Y., et al. Enhancer loops appear stable during development and are associated with paused polymerase. Nature. 512 (7512), 96-100 (2014).
- Foe, V. E., Alberts, B. M. Studies of nuclear and cytoplasmic behaviour during the five mitotic cycles that precede gastrulation in Drosophila embryogenesis. J Cell Sci. 61, 31-70 (1983).
- Blythe, S. A., Wieschaus, E. F. Zygotic Genome Activation Triggers the DNA Replication Checkpoint at the Midblastula Transition. Cell. 160 (6), 1169-1181 (2015).
- Blythe, S. A., Wieschaus, E. F. Establishment and maintenance of heritable chromatin structure during early Drosophila embryogenesis. eLife. 5, e20148(2016).
- JoVE Science Education Database. Embryo and Larva Harvesting and Preparation. Biology I: yeast, Drosophila and C. elegans. Drosophila melanogaster. , JoVE, Cambridge, MA. (2017).
- Sicaeros, B., O'Dowd, D. K. Preparation of Neuronal Cultures from Midgastrula Stage Drosophila Embryos. Journal of Visualized Experiments. (5), (2007).
- Shermoen, A. W. Preparation of Baskets for Drosophila Egg Collections, Treatments, and Incubations. Cold Spring Harbor Protocols. (10), (2008).
- Ay, F., Noble, W. S. Analysis methods for studying the 3D architecture of the genome. Genome biology. 16 (1), 183(2015).
- Lazaris, C., Kelly, S., Ntziachristos, P., Aifantis, I., Tsirigos, A. HiC-bench: comprehensive and reproducible Hi-C data analysis designed for parameter exploration and benchmarking. BMC Genomics. 18 (1), (2017).
- Servant, N., et al. HiC-Pro: an optimized and flexible pipeline for Hi-C data processing. Genome Biology. 16 (1), (2015).
- Durand, N. C., et al. Juicer Provides a One-Click System for Analyzing Loop-Resolution Hi-C Experiments. Cell systems. 3 (1), 95-98 (2016).
- Lajoie, B. R., Dekker, J., Kaplan, N. The Hitchhiker's guide to Hi-C analysis: Practical guidelines. Methods. 72, 65-75 (2015).
- Schmitt, A. D., Hu, M., Ren, B. Genome-wide mapping and analysis of chromosome architecture. Nature Reviews. Molecular Cell Biology. 17 (12), 743-755 (2016).
- Shin, H., et al. TopDom: an efficient and deterministic method for identifying topological domains in genomes. Nucleic Acids Res. 44 (7), e70(2016).
- Kruse, K., Hug, C. B., Hernández-Rodríguez, B., Vaquerizas, J. M. TADtool: visual parameter identification for TAD-calling algorithms. Bioinformatics. 32 (20), 3190-3192 (2016).
- Lévy-Leduc, C., Delattre, M., Mary-Huard, T., Robin, S. Two-dimensional segmentation for analyzing Hi-C data. Bioinformatics. 30 (17), Oxford, England. i386-i392 (2014).
- Filippova, D., Patro, R., Duggal, G., Kingsford, C. Identification of alternative topological domains in chromatin. Algorithms for molecular biology: AMB. 9 (1), 14(2014).
- Crane, E., et al. Condensin-driven remodelling of X chromosome topology during dosage compensation. Nature. 523 (7559), 240-244 (2015).
- Durand, N. C., et al. Juicebox Provides a Visualization System for Hi-C Contact Maps with Unlimited Zoom. Cell systems. 3 (1), 99-101 (2016).
- Zhou, X., et al. Exploring long-range genome interactions using the WashU Epigenome Browser. Nature Methods. 10 (5), 375-376 (2013).
- Ramírez, F., et al. High-resolution TADs reveal DNA sequences underlying genome organization in flies. bioRxiv. , 115063(2017).
- Kerpedjiev, P., et al. HiGlass: Web-based Visual Comparison And Exploration Of Genome Interaction Maps. bioRxiv. , 121889(2017).
- Hug, C. B., Grimaldi, A. G., Kruse, K., Vaquerizas, J. M. Chromatin Architecture Emerges during Zygotic Genome Activation Independent of Transcription. Cell. 169 (2), (2017).
- Berkum, N. L., et al. Hi-C: a method to study the three-dimensional architecture of genomes. Journal of Visualized Experiments: JoVE. (39), (2010).
- Naumova, N., et al. Organization of the mitotic chromosome. Science. 342 (6161), 948-953 (2013).
- Denker, A., de Laat, W. The second decade of 3C technologies: detailed insights into nuclear organization. Genes & development. 30 (12), 1357-1382 (2016).
- Belaghzal, H., Dekker, J., Gibcus, J. H. Hi-C 2.0: An optimized Hi-C procedure for high-resolution genome-wide mapping of chromosome conformation. Methods (San Diego, Calif). 123, 56-65 (2017).
Réimpressions et Autorisations
Demande d’autorisation pour utiliser le texte ou les figures de cet article JoVE
Demande d’autorisationThis article has been published
Video Coming Soon
À PROPOS DE JoVE
Copyright © 2025 MyJoVE Corporation. Tous droits réservés.