
Method Article
הדור של הגנום כולו כרומטין קונפורמציה לכידת ספריות מעוברים דרוזופילה בחוזקה בשלבים המוקדמים
In This Article
Summary
עבודה זו מתאר פרוטוקול עבור הדור של רזולוציה גבוהה בחיי עיר ויטמינצ'יק ספריות מ מבוים בחוזקה gastrulation קדם דרוזופילה melanogaster עוברי.
Abstract
חוקרים את הארכיטקטורה תלת מימדי של כרומטין מציע תובנה מנגנוני הכונה שלא יסולא בפז. כאן, אנו מתארים עבור ביצוע ה כרומטין קונפורמציה לכידת טכניקה בחיי עיר ויטמינצ'יק על פרוטוקול מבוים דרוזופילה melanogaster העובר אוכלוסיות. התוצאה היא ספריה רצף המאפשר את המיפוי של כל האינטראקציות כרומטין שמתרחשות בגרעין בניסוי יחיד. עובר מיון נעשית באופן ידני באמצעות סטריאו מיקרוסקופ פלואורסצנטי, וקו לטוס הטרנסגניים המכילה סמן גרעינית. ניתן להשיג באמצעות טכניקה זו, העובר אוכלוסיות של כל מחזור החטיבה גרעינית, ועם מצב מוגדר מחזור התא, עם טוהר גבוהה מאוד. הפרוטוקול עשוי גם להיות מותאם כדי למיין בוגרים עוברי מעבר gastrulation. העוברים מיון משמשים כקלט בחיי עיר היי-סי כל הניסויים, לרבות קביעת רצף ספריית הכנה, יכולה להסתיים בעוד חמישה ימים. הפרוטוקול יש דרישות הקלט נמוך ועובד בצורה אמינה באמצעות 20 blastoderm שלב עוברי כחומר קלט. התוצאה הסופית היא ספריית רצף עבור רצף הדור הבא. לאחר רצף, ניתן לעבד את הנתונים לתוך הגנום כולו כרומטין אינטראקציה עם מפות מסוגל לנתח באמצעות מגוון רחב של כלים זמינים כדי לקבל מידע לגבי topologically שיוך מבנה קבוצת המחשבים (טד) כרומטין לולאות, כרומטין תאים במהלך הפיתוח דרוזופילה .
Introduction
לכידת קונפורמציה כרומטין (3 ג) התפתחה שיטה שימושית במיוחד כדי לחקור את הטופולוגיה של כרומטין גרעין1. הגרסה 3 ג ויטמינצ'יק מאפשרת מדידה התדרים קשר של כל האינטראקציות כרומטין שמתרחשות בגרעין ניסוי יחידה2. יישום של ויטמינצ'יק שיחק תפקיד חשוב הגילוי, אפיון רבים עקרונות היסוד של ארגון כרומטין, כגון TADs, תאים לולאות3,4,5.
לימודי האדריכלות כרומטין בהקשר של מעברים התפתחותיים ובידול תא משמשים יותר ויותר כדי לחשוף את המנגנונים של הכונה במהלך אלה תהליכים6,7,8, 9. אחד האורגניזמים דגם עניין רב הוא דרוזופילה melanogaster, הגנום ופיתוח אשר מאופיינים היטב. עם זאת, מחקרים מעטים לחקור אדריכלות כרומטין דרוזופילה מחוץ במבחנה תרביות רקמה הגדרות כבר ערכו10,11. עוברי 16 – 18 h שלאחר ההפריה, TADs, תאים מזכיר מבנים דומים אצל יונקים היו מזוהה10, אשר מעלה השאלה של איזה תפקיד הם משחקים בהכונה במהלך דרוזופילה העובר פיתוח. במיוחד בשלבים המוקדמים של פיתוח, לפני gastrulation, מחקרים כאלה הן מבחינה טכנית מאתגר. לפני gastrulation, עוברי דרוזופילה עוברים 13 דיביזיות גרעיני סינכרונית להמשיך בקצב מסחרר של 8 – 60 דקות לכל מחזור12,13. בנוסף לכך, חוסר של תכונות חזותיות כדי להבחין בין השלבים השונים להקשות להשיג חומר העובר בשלבים בחוזקה בכמויות מספיקות.
כדי לפתח פרוטוקול המאפשר ללמוד אדריכלות כרומטין בהתפתחות המוקדמת דרוזופילה ברזולוציה מחזור גרעינית, שילבנו בין שתי שיטות קיימות: בחיי עיר Hi-C, אשר מאפשר את הדור של רזולוציה גבוהה כל הגנום מפות קשר5ו- staging העובר באמצעות שורת דרוזופילה הטרנסגניים לבטא13,transgene14eGFP-PCNA. Transgene זו רגישה את הגרעין במהלך לאטמוספרה, והוא מפזר לאורך blastoderm syncytial במהלך מיטוזה. באמצעות מאפיין זה, אפשרי להבחין בקלות שלבים שונים על ידי צפיפות גרעינית שלהם ואת mitotic העוברים על ידי הפיזור של אות ה-GFP.
יחד, טכניקות אלה מאפשרות לימוד מבנה תלת ממדי של כרומטין ברזולוציה גבוהה מגם עוברי דרוזופילה 20 בהיקף. פרוטוקול זה כולל את ההוראות קציר ומיון עוברי דרוזופילה להשיג אוכלוסיות של העוברים של מחזור מחטיבה גרעיני. בהמשך מתאר כיצד העוברים שהושג משמשות לביצוע בחיי עיר היי-סי התוצאה הסופית היא ספריית נוקלאוטיד מתאים רצף על מכונות רצף ' הדור הבא '. ואז ניתן לעבד את קריאות רצף שנוצר לתוך מפות אינטראקציה עם כרומטין מפורטת המכסה את כל הגנום דרוזופילה .
Protocol
1. אוסף העובר דרוזופילה
הערה: אוסף המקביל העובר יכול להתבצע כפי שמוצג הפרסום הקודם15.
- העברת הזבובים eGFP צעיר-PCNA (< בן שבוע) לתוך ביצה כלובי איסוף עם אוסף yeasted לוחות16 (אתנול 1%, 1% חומצה אצטית, ו 4% אגר).
- להעביר אוסף הכלובים אינקובטור שוכן בגובה 25 º C. המודל הפסיכוסקסואלי # שלב 1 – 2 ימים לפני אוסף ביצי משפר באופן משמעותי את התשואה ביצה. שינוי אוסף צלחות פעמיים ביום.
- להסיר את הלוחות המכיל העוברים מן החדר אוסף במרווחים של 30-60 דקות. במרווחים קטנים יותר לגרום עוברי פחות, אך ההפצה הדוקה של שלבים התפתחותיים. לאסוף מן הכלובים מרובות במקביל אז זה אידיאלי > 200 הביצים מוטלות כל 30-60 דקות.
- חנות את הלוחיות 25 ° c עד העוברים מגיעים לגיל הרצוי. Blastoderm שלב עוברי (מחזור גרעיני 14), תקופת דגירה של 2 h.
- לאחר שעתיים של דגירה, להוסיף ברז מים מבקבוק שפריץ וספרייט כך השטח כולו מכוסה מים. להשעות את העוברים, שמרים בעזרת מברשת רכה.
- יוצקים resuspended העוברים מן וספרייט לסל אוסף העובר (strainers תא מסחרי עם גודל הנקבוביות מיקרומטר 100 או העבודה17 סלים תוצרת בית טוב), הוספת נוספים ברז מים מבקבוק מים, במידת הצורך. בשלב זה, לשלב העוברים מן כל הצלחות שנאספו במקביל. המדגם במאגר מייצג אצווה בודדת.
- לשטוף את העוברים על ידי שטיפה הסל עם ברז מים מבקבוק שפריץ ל 30 s עד כל שאריות שמרים נשטף הרחק.
- Dechorionate העוברים על-ידי הצבת סל איסוף לתוך פתרון 2.5% נתרן תת-כלורי במים. עצבנות קלה על ידי מתערבל מקלה על הסרת chorion. המשך עד עוברי הם מספיק הידרופובי כך הם צפים על פני השטח של הפתרון כאשר הסל הרים החוצה שקוע שוב, אשר צריך לקחת ~1.75–2 דקות.
זהירות: נתרן תת-כלורי היא מאכל. ללבוש ציוד מגן אישי המתאים. פתרונות המכיל < 10% נתרן תת-כלורי בדרך כלל יכול להיות מסולק בכיור, הקפד לבדוק את התקנות של המוסד המארח. - הסר את הסל של הפתרון, לשטוף היטב עם מים מהברז מבקבוק מים עד הריח של האקונומיקה כבר אינו מורגש.
2. עובר קיבוע
הערה: קיבוע אופטימלית תנאים, בעיקר את הריכוז של דטרגנט, פורמלדהיד, משך הזמן של קיבעון, צריך להיות נחוש מדעית להתאים את השלב של העוברים. על במות ברחבי blastoderm syncytial, ריכוז סופי של 0.5% טריטון X-100 ו- 1.8% פורמלדהיד בשלב מימית עובד טוב. עבור בשלבים מאוחרים יותר מעבר העובר בשלב 9, בהמשך אופטימיזציה של פרמטרים אלה ייתכן שיהיה צורך. כל הפתרונות בשימוש במהלך קיבוע ומיון צריך להכיל מעכבי פרוטאז.
- היפוך אוסף סל ומניחים אותו מעל צינור חרוטי צנטריפוגה 15 מ"ל. ריקון העוברים מן הסל לתוך הצינורית באמצעות פיפטה של פסטר dispensing PBS-T (PBS, 0.5% טריטון X-100).
- תן עוברי להתישב בתחת ולכוונן את העוצמה סה כ 2 מ עם PBS-טי
- מוסיפים 6 מ של heptane ב 100 µL של 37% פורמלדהיד במים.
התראה: heptane ב ופורמלדהיד רעילים כאשר בשאיפה או לאחר העור פנה. ללבוש ציוד מגן אישי המתאים ולעבוד בשכונה fume. פסולת המכילה heptane ב או פורמלדהיד צריך להיות מסולק בנפרד לפי התקנות של המכון המארח. - לאחר התוספת של הפורמלדהיד, להפעיל טיימר 15 דקות ומנערים נמרצות את הצינור למעלה ולמטה במשך 1 דקה בעבודת יד. השלב מימית ואורגניים ישלב לטופס שמפו-כמו עקביות.
- להתסיס על מערבל על כל ההזמנות עד 10 דקות לאחר התוספת של פורמלדהיד.
- צנטריפוגה ב x 500 g עבור 1 דקות בטמפרטורת החדר, כדי לאסוף את העוברים בחלק התחתון של הצינור.
- תשאף הנוזל כמו שמפו כולו וזורקים אותו, מטפל לא לרוקן כל העוברים. כמויות קטנות הנותרים של תגובת שיקוע כמו שמפו לא לגרום לבעיות.
- 15 דקות לאחר התוספת של פורמלדהיד, resuspend את העוברים ב 5 מ של PBS-T עם 125 מילימטר גליצין כדי להרוות את הפורמלדהייד. מערבבים היטב ע י ניעור למעלה ולמטה במשך 1 דקה.
- צנטריפוגה ב x 500 g בטמפרטורת החדר במשך 1 דקה, תשאף תגובת שיקוע.
- לשטוף את העוברים על-ידי resuspending אותם ב 5 מ של PBS קר כקרח-טי לתת ייחוס עוברי וביופסיה כל supernatant.
- חזור לשטוף את שלב 2.10 עוד פעמיים.
- לשמור העוברים על הקרח עד מיון. בדרך כלל, זה רעיון טוב כדי לאסוף 3-4 קבוצות של עוברי לטוס לפני שתמשיך למיון. עם זאת, העוברים מיון באותו היום. אחסון מורחב על קרח או במקרר מוביל העובר שינו מורפולוגיה.
3. עובר מיון
הערה: מיון יכול להיעשות על כל מיקרוסקופ סטריאו פלורסנט מצויד במסנן ה-GFP בהגדלה X 60 – 80.
- באמצעות פיפטה 1,000 µL, להעביר אוסף של העוברים כ-100 כלי זכוכית קטנים מתאימים למיון, רצוי של צבע כהה, ומניחים אותו על קרח.
-
העוברים מיון לפי גרעיני צפיפות, מחזור התא מצב (איור 1) על ידי דחיפת עוברי רצוי לתוך ערימה נפרדת באמצעות טיפ מחט או מזרק.
- הסר את כל העוברים עם מפוזר, לא גרעיני הפצה של eGFP-PCNA (איור 1E). בנוסף, העוברים המציגים חלקית אות GFP לא גרעיני להסירם.
- כדי לסייע המיון, להרכיב שורה של הפניה עוברי-גרעיני מחזור 12, 13 ו- 14 בכל אצווה באמצעות התמונות באיור 1 כמדריך. השתמש זה ההרכב תואמים העוברים של שלב לא ידוע עם אחד העוברים הפניה על מנת לקבוע את הבמה שלהם.
- כדי לוודא את הבמה התפתחותית עבור הפניה עוברי, למדוד צפיפות גרעינית על ידי הדמיה העובר וספירת מספר גרעינים על פני השטח של העובר שטח של 2,500 מיקרומטר2 באמצעות תוכנת הדמיה המספק מידע מרחק.
הערה: המספר הצפוי של גרעינים על שטח של 2,500 מיקרומטר2 הוא 12 עד 16 גרעינים-גרעין מחזור 12, 20-30 גרעינים-גרעין מחזור 1313.
- לאחר כל העוברים בשלב המתאים מופרדים, לוקחים תמונות של העוברים על תיעוד ובקרת איכות. אם סטריאו מיקרוסקופ אינו עצמו מאובזר עם מודול המצלמה, עשוי לשמש כל מיקרוסקופ epifluorescence עם מסנני ה-GFP.
- פיפטה את העוברים הרצוי באמצעות פיפטה של 1,000 µL, העברת צינור טריים, ו המקום על קרח.
- המשך עד עוברי מספיק מסודרים עבור הניסוי המתוכנן. עבור עוברי בוגרים מאשר שלב 9, בדרך כלל 20 עוברי מספיקים אחד בחיי עיר ויטמינצ'יק בניסוי. גרעיני מחזור 12, העוברים 80 הם נקודת התחלה טובה. במחזורים קודמות, מספר העוברים צריך כ להיות הכפילה של כל מחזור.
- בריכת ופיצול עוברי לתוך צינורות 1.5 mL כך כי שפופרת אחת מכילה מספיק עוברי עבור יישום יחיד בחיי עיר ויטמינצ'יק הניסוי. רצוי להשתמש צינורות עם מאפיינים איגוד ה-DNA נמוכה, שכן הצינור אותו ישמש עבור פרוטוקול כולו ספיחה של דנ א יכול להוביל הפסדים משמעותיים בריכוזים נמוכים של ה-DNA.
- לסובב את הצינורות בקצרה ב x 100 g בטמפרטורת החדר ולהסיר תגובת שיקוע. העוברים צריך להיות יבש ככל האפשר עבור לקפוא.
- פלאש להקפיא את העוברים על ידי השוקע צינורות חנקן נוזלי, חנות ב-80 מעלות צלזיוס.
4. בחיי עיר שלום-C
- פירוק
- במקום צינורות עם עוברי קפוא על קרח.
- Resuspend עוברי ב 500 µL של המאגר הקר פירוק (10 מ מ טריס-Cl pH 8.0, 10 מ מ NaCl, 0.2% IGEPAL CA-630, מעכבי פרוטאז; מים מומס ב). ואז לחכות 1 דקות לתת עוברי להתישב בתחת של הצינור.
- לטחון עוברי באמצעות המרוסקים מיקרו על מתכת, טרום מקורר על קרח, אשר נועד מתאימים בחוזקה צינור microcentrifuge 1.5 mL.
- כדי להימנע לזעזע את העוברים, הכנס איחדו את באיטיות עד שהוא נוגע בתחתית הצינורית לדחוף למטה, ואז לטחון על-ידי סיבוב איחדו את פעמיים, בשני הכיוונים.
- הרם את שהעקב טיפה, לדחוף לתחתית הצינור שוב, וחזור שחיקה.
- חזור על 4.1.3.2 10 פעמים, או עד העוברים הם לחלוטין lysed. הפתרון צריך להיות הומוגני, אין חתיכות גדולות שיורית של עוברי צריך להישאר.
- דגירה התליה homogenized על הקרח במשך 15 דקות ספין-x 1000 גרם, 4 מעלות צלזיוס למשך 5 דקות, ולמחוק את תגובת שיקוע.
- רחץ צניפה מאת resuspending במאגר 500 µL פירוק קר כקרח, pipetting למעלה ולמטה.
- ספין שוב כמו 4.1.4, ולמחוק את תגובת שיקוע.
- Resuspend צנפה שטף ב 100 µL של 0.5% נתרן dodecyl סולפט (מרחביות), pipetting למעלה ולמטה. Permeabilize גרעינים על ידי המקננת 10 דקות ב 65 מעלות צלזיוס בתוך גוש חימום. להרוות מרחביות על-ידי הוספת 50 µL של 10% טריטון µL X-100 ל-120 מים. מערבבים על ידי מצליף את הצינור.
- דגירה ב 37 מעלות צלזיוס למשך 15 דקות חום בלוק.
- אנזים הגבלה עיכול
- להוסיף 25 µL של 10 x אנזים הגבלה מאגר ו- 20 U MboI U/µL 5. מערבבים על ידי מצליף את הצינור.
- לעכל את ה-DNA על ידי המקננת במשך 90 דקות ב 37 מעלות צלזיוס בבלוק חום תחת עצבנות קלה (750 סל ד).
- להוסיף עוד 20 U של MboI והמשך הדגירה במשך 90 דקות.
- חום-בטל MboI על ידי המקננת ב 62 מעלות צלזיוס למשך 20 דקות.
- הסככה מילוי
הערה: מילוי הסככה עם biotinylated dATP מאפשר בחירה של קטעים מחוברים ספציפיים. ביוטין-dATP ובצמתים מצדו מוגן מפני הפעילות אקסונוקלאז של T4 DNA פולימראז (סעיף 4.6), ואילו ביוטין-dATP קצותיו בוטה unligated מוסר ביעילות. הנפתח באמצעות חרוזים מצופים streptavidin בסעיף 4.7 לכן במיוחד מעשיר על שברי DNA מחוברים, chimeric.- הוסף µL 18 של 0.4 מ מ ביוטין-14-dATP, µL 2.25 של תערובת dCTP/dGTP/dTTP שלא שונתה (3.3 מ מ כל אחד) ו- 8 µL של 5 U/µL DNA פולימראז I Klenow קטע.
- לערבב על ידי מצליף את הצינור, דגירה ב 37 מעלות צלזיוס במשך 90 דקות בבלוק חום.
- מצדו
- להוסיף 657 µL של מים, µL 120 10 x T4 DNA ליגאז מאגר, 100 µL של 10% טריטון X-100, 6 µL של 20 מ"ג/מ"ל אלבומין שור (BSA), מיקס על ידי מצליף את הצינור. לבסוף להוסיף 5 µL של 5 U/µL T4 DNA ליגאז, מיקס על ידי מצליף את הצינור.
- סובב צינור בעדינות (20 סל ד) בטמפרטורת החדר במשך שעתיים.
- הוסף את הפרק השני של 5 µL של 5 U/µL T4 DNA ליגאז ולהמשיך סיבוב 2 h יותר.
- ספין למטה גרעינים ב x 2,500 g למשך 5 דקות ולמחוק את תגובת שיקוע.
- הפקת דנ א
- Resuspend צנפה ב 500 µL מאגר החילוץ (50 מ מ טריס-Cl pH 8.0, 50 מ מ NaCl, 1 מ Ethylenediaminetetraacetic חומצה (EDTA), 1% מרחביות; מים מומס ב) ולהוסיף 20 µL של 20 מ"ג/מ"ל proteinase קיי מיקס על ידי מצליף את הצינור.
- לעכל פרוטאינים על ידי המקננת ב 55 מעלות צלזיוס למשך 30 דקות, רועדת-1000 סל ד.
- כדי דה-crosslink, להוסיף µL 130 של 5 M NaCl, דגירה בין לילה ב 68 מעלות צלזיוס, רועדת-1000 סל ד.
- פיפטה מדגם לתוך צינור 2 mL החדש, מעדיפים עם מאפייני איגוד ה-DNA נמוך.
- הוסף 0.1 x אמצעי אחסון (63 µL) של 3 מ' סודיום אצטט pH 5.2 ו- 2 µL של 15 מ"ג/מ"ל GlycoBlue. מערבבים היטב על ידי היפוך. להוסיף כרכים x 1.6 (1,008 µL) הטהור המוחלט אתנול וביו -מיקס על ידי היפוך.
- דגירה ב-80 מעלות צלזיוס במשך 15 דקות צנטריפוגה ב x 20,000 g ב 4 מעלות צלזיוס למשך 30 דקות לפחות. בגדר ה-DNA הוא לעתים קרובות מאוד קטן, כמעט בלתי נראים, ניתן הבחין רק בשל צבע כחול של GlycoBlue.
- הסר תגובת שיקוע בזהירות רבה, נע הטיפ פיפטה לתוך הצינור לאורך הקיר ההפוך שבו נמצא בגדר ה-DNA. תרסיס הנותרים מוסרות לעיתים קרובות בקלות במהלך שלב זה, שוטף את הבאים על ידי דוחף אותם מתוך הצינורות באמצעות טיפ P10 ולא pipetting אותם.
- רחץ צניפה על-ידי הוספת µL 800 של 70% אתנול. לערבב על-ידי היפוך, צנטריפוגה ב x 20,000 g בטמפרטורת החדר במשך 5 דק חוזר זה שטיפת לפחות פעם אחת.
- להסיר כל עקבות של אתנול ולהשאיר את הצינור עומד עם פתח מכסה עבור עד 5 דקות כדי מילה נהדרת... לאחר שהנוזל לא נשאר, להוסיף 50 µL של 10 מ מ טריס-Cl pH 8.0. שוב ושוב pipette הפתרון מעל האזור על הקיר של הצינור שבו היה ממוקם בגדר כדי solubilize את ה-DNA.
- להוסיף 1 µL של 20 מ"ג/מ"ל RNase A, מיקס על ידי מצליף ברכבת התחתית, דגירה ב 37 מעלות צלזיוס למשך 15 דקות כדי לעכל RNA. המדגם עכשיו ניתן לאחסן במקרר למשך הלילה או קפוא ב-20 ° C ללא הגבלת זמן.
- בדוק את ריכוז הדנ א באמצעות וזמינותו הפלורסנט מבוסס על פי הוראות היצרן. הסכום הכולל של ה-DNA המדגם צריך להיות לפחות 10 ng, אחרת גם קצת חומר זמין עבור הגברה, ספריית המורכבות צפויה להיות נמוכה. כאשר זה קורה, כמות החומר מתחיל כנראה לא היה מספיק, או חומר אבד לאורך הדרך, אולי במהלך פירוק, משקעים.
- הסרת ביוטין ו- DNA הטיה
- להוסיף יחד µL 12 10 x T4 DNA פולימראז המאגר, 3 µL של 1 מ מ dATP, 3 µL של 1 מ מ dGTP ו- µL 46 של מים. מערבבים על ידי מצליף את הצינור. להוסיף 5 µL של 3 U/mL T4 DNA פולימראז, לערבב על ידי מצליף את הצינור, דגירה ב 20 מעלות צלזיוס למשך 30 דקות.
- להוסיף 3 µL של 0.5 M EDTA לעצור את התגובה ולהשתמש במים כדי להביא את הדגימה נפח של-120 µL.
- להטות את הדנ א לגודל של 200-400 bp משתמש בהתקן sonication בהתאם להוראות היצרן. באמצעות את sonicator הזכיר את הטבלה של חומרים, התוכנית הבאה היא המתאימה: 2 מחזורים כל אחד 50 s, 10% חובה, בעוצמה 5, 200 מחזורים/פרץ.
- ביוטין הנפתח
- פיפטה 30 µL של 10 מ"ג/מ"ל streptavidin מצופה beads מגנטי לתוך צינור חדש, להפריד אותם על דוכן מגנטי, ולמחוק את תגובת שיקוע.
- Resuspend חרוזים 1 x מאגר B & W (5 מ מ טריס-Cl pH 7.4, 0.5 מ מ EDTA, 1 M NaCl; מים מומס ב) + 0.1% טריטון X-100 ו מיקס על ידי vortexing. במקום הצינור על דוכן מגנטי והמתן 1 – 5 דקות עד החרוזים מופרדים, בהתאם היצרן והדגם.
- האחות ולמחוק את תגובת שיקוע תוך החלקת הטיפ פיפטה לאורך גדר מול שבו ממוקמים החרוזים. Resuspend חרוזים ב 120 µL של 2 x מאגר B & W (10 מ מ טריס-Cl pH 7.4, EDTA 1 מ"מ, 2 M NaCl). מערבבים על-ידי vortexing.
- העברת DNA הוטו צינור חדש DNA נמוך מחייב, ולערבב עם 120 µL של ההשעיה חרוז במאגר X B & W 2 מאת vortexing. סובב חרוזים לדנ ב rpm 20 למשך 15 דקות.
- להפריד חרוזים על דוכן מגנטי ולמחוק את תגובת שיקוע.
- Resuspend חרוזים ב 600 µL של 1 x B & W + 0.1% X-100 טריטון, דגירה ב 55 מעלות צלזיוס למשך 2 דקות, רועדת-1000 סל ד. לאחר ההפרדה, למחוק את תגובת שיקוע. חזור על שטיפת הזה פעם אחת.
- לשטוף חרוזים פעם אחת עם 600 µL של 10 מ מ טריס-Cl pH 8.0, ולמחוק את תגובת שיקוע לאחר ההפרדה.
- Resuspend חרוזים ב- 50 µL של 10 מ מ טריס-Cl pH 8.0.
5. רצף ספריית הכנה
הערה: כל השלבים ספריית נעשים באמצעות רכיבים מתוך הכנה ספריית DNA מסחרי קיט (ראה טבלה של חומרים). עם זאת, ערכות חלופי או אחרים יכול להיות מוחלף. משקעים נוטה טופס בהסוכנים הכנה ספריית אחסון למקפיא. לכן חשוב לוודא כי כל המשקעים התפרקה לפני השימוש של נוגדנים.
-
סיום תיקון
- להעביר את המתלים חרוז ב- 50 µL של 10 מ מ טריס-Cl pH 8.0 לתוך צינור ה-PCR.
- להוסיף 3 µL של סוף אנזים הכנה מערבבים ו µL 7 בסוף התגובה הכנה המאגר. מערבבים על-ידי pipetting למעלה ולמטה.
- להעביר צינור הצנטרפוגה תרמי ולהפעיל את התוכנית הבאה: 20 ° C למשך 30 דקות, 65 מעלות צלזיוס במשך 30 דקות, והחזק ב 4 º C.
-
מתאם מצדו
- להוסיף 30 µL של מיקס מאסטר מצדו, 2.5 µL של 1.5 מיקרומטר רצף מתאם (שתדללו מיקרומטר 1.5 במלאי) ולאחר µL 1 של מצדו משפר את המתלה חרוז. מערבבים על-ידי pipetting למעלה ולמטה.
- דגירה ב 20 מעלות צלזיוס למשך 15 דקות הצנטרפוגה תרמי.
- להוסיף 3 µL של המשתמש האנזים. מערבבים על-ידי pipetting למעלה ולמטה.
- דגירה ב 37 מעלות צלזיוס למשך 15 דקות הצנטרפוגה תרמי.
- להפריד חרוזים על דוכן מגנטי ולהסיר תגובת שיקוע.
- כדי לשטוף חרוזים, resuspend חרוזים ב µL 100 1 x מאגר B & W + 0.1% טריטון X-100. מיקס על ידי vortexing, העברת צינור microcentrifuge. להפריד חרוזים על דוכן מגנטי ולהסיר תגובת שיקוע.
- חזור על זה לשטוף פעם אחת באמצעות 600 µL של המאגר אותו.
- Resuspend חרוזים ב 600 µL של 10 מ מ טריס-Cl pH 8.0, מיקס על ידי vortexing, ולהעביר חרוזי צינור חדש.
- נפרדים חרוזים על דוכן מגנטי, למחוק את תגובת שיקוע ולאחר resuspend חרוזים ב- 50 µL של 10 מ מ טריס-Cl pH 8.0.
-
PCR-הגברה
- להכין שתי המבחנות ומערבבים בכל, 25 µL של מיקס מאסטר פולימראז µL 1.5 של 10 מיקרומטר קדמי (מדד) PCR פריימר, µL 1.5 של 10 מיקרומטר הפוכה (אינדקס) PCR פריימר.
הערה: קדימה פריימר PCR (מדד):
5'-AATGATACGGCGACCACCGAGATCTACACTCTTTCCCTACACGACGCTCTTCCGATC * T-3´.
הפוך פריימר PCR (אינדקס):
5'-CAAGCAGAAGACGGCATACGAGATNNNNNNGTGACTGGAGTTCAGACGTGTGCTCTTCCGATC * T-3´. * מציין phosphorothioate חוב, Ns ב ומהצמיגים PCR באינדקס. - כל שפופרת, להוסיף µL 22 של הבולם חרוז, מיקס על ידי pipetting למעלה ולמטה.
- להפעיל את ה-PCR באמצעות התוכנית הבאה: 98 ° C עבור 1 דקות, (98 ° C 15 s, 65 מעלות צלזיוס במשך 75 s, ramping 1.5 ° C/s) חזר על 9 - 12 פעמים, 65 מעלות צלזיוס במשך 5 דקות, והחזק ב 4 º C.
הערה: מספר מחזורי הגברה צריך להיקבע מדעית. עם זאת, מצאנו כי ספריות נדרש יותר מ-12 מחזורים היו באופן כללי של מורכבות נמוכה ולא לגרום באיכות גבוהה ש-hi-C מפות. מצד שני, ספריות נדרש פחות מ-12 מחזורים היו לא מושפעים לרעה על ידי הגברה עבודה מלא 12 מחזורים. לכן, זה אפשרי כברירת מחדל, לביצוע 12 מחזורים של הגברה. - בריכת שתי תגובות PCR בשפופרת microcentrifuge יחיד, להפריד חרוזים על דוכן מגנטי, ולהעביר את תגובת שיקוע המכיל את הספריה שיש צינור.
- להכין שתי המבחנות ומערבבים בכל, 25 µL של מיקס מאסטר פולימראז µL 1.5 של 10 מיקרומטר קדמי (מדד) PCR פריימר, µL 1.5 של 10 מיקרומטר הפוכה (אינדקס) PCR פריימר.
-
בחירת גודל
- להביא Ampure XP חרוז ההשעיה בטמפרטורת החדר, לערבב היטב על ידי רועדת.
- להביא עוצמת התגובה PCR במאגר בדיוק 200 µL עם מים. במהלך ה-PCR, הפרדה מגנטית, חלק האחסון המקורי אובד בדרך כלל. ודא אמצעי אחסון על-ידי הגדרת את פיפטה 200 µL, תשאף בכל אמצעי האחסון של התגובה. אם האוויר הוא aspirated, יותר מים צריך להתווסף. אם אמצעי האחסון חורגת 200 µL, לכוונן את עוצמת הקול של חרוזים הוסיף בשלבים 5.4.3 ו 5.4.6 באופן פרופורציונלי.
הערה: אמצעי האחסון בסוגריים בתוקף, אם הנפח הכולל של תגובות PCR במאגר הוא בדיוק 200 µL. - מוסיפים כרכים x 0.55 (110 µL) Ampure XP חרוז השעיה ומערבבים על ידי pipetting למעלה ולמטה לפחות 10 פעמים.
- דגירה בטמפרטורת החדר במשך 5 דקות, חרוזים נפרד על דוכן מגנטי במשך 5 דקות.
- תגובת שיקוע להעביר צינור חדש. למחוק את הצינור המכיל את החרוזים. החרוזים אוחזים דנ א > 700 bp, אשר הוא גדול מדי כדי להיות רציף.
- תגובת שיקוע, להוסיף x 0.2 כרכים (40 µL, וכתוצאה מכך סך של מאגר x Ampure 0.75 במדגם) של הבולם חרוז Ampure XP, מיקס על ידי pipetting למעלה ולמטה 10 פעמים.
- דגירה בטמפרטורת החדר במשך 5 דקות, חרוזים נפרד על דוכן מגנטי במשך 5 דקות.
- למחוק את תגובת שיקוע אשר מכיל DNA < 200 bp, אשר כוללת תחל חינם, פריימר הדימרים שברי קטן מדי כדי להיות רציף.
- להשאיר את הצינורית על הדוכן מגנטי. כדי לשטוף חרוזים, להוסיף 700 µL של 80% אתנול, דואגת שלא להפריע בגדר חרוז ולאחר תקופת דגירה של 30 s.
- למחוק את תגובת שיקוע, לאחר מכן להשתמש ברכבת התחתית את הדוכן מגנטי, resuspend חרוזים ב 100 µL של 10 מ מ טריס-Cl pH 8.0. לערבב על ידי pipetting למעלה ולמטה 10 פעמים, דגירה בטמפרטורת החדר במשך 1 דקה.
- הוסף כרכים x 0.8 (80 µL) של Ampure XP חרוז ההשעיה. לערבב על-ידי pipetting למעלה ולמטה 10 פעמים, דגירה בטמפרטורת החדר במשך 5 דקות. זה השלב השני של בחירת גודל תחתון מבטיחה כי הספריה הסופית היא חופשית לחלוטין של תחל ו הדימרים פריימר.
- להפריד חרוזים על דוכן מגנטי עבור 5 דקות ולמחוק את תגובת שיקוע.
- לשטוף את צניפה חרוז פעמיים עם 700 µL של 80% אתנול ב-30 s כל אחד, תוך השארת הצינורית על הדוכן מגנטי, כמפורט לעיל.
- עם הרכבת התחתית עדיין על הדוכן מגנטי, להסיר כל עקבות של אתנול. היא מסייעת לדחוף טיפות של אתנול יצאה מהשפופרת באמצעות פיפטה של P10. תן אתנול שיורית להתאדות במשך 5 דקות לכל היותר.
- קח צינור את הדוכן מגנטי, resuspend חרוזים ב- 50 µL של 10 מ מ טריס-Cl pH 8.0. מערבבים על-ידי pipetting למעלה ולמטה 10 פעמים.
- דגירה בטמפרטורת החדר במשך 5 דקות, ואז נפרד חרוזים על דוכן מגנטי.
- תגובת שיקוע להעביר צינור טריים. זוהי הספרייה ויטמינצ'יק הסופי, מוכן להיות לכמת, וסודרו המכונות רצף של הדור הבא, על פי הוראות היצרן.
תוצאות
מיון אוכלוסיות העובר-גרעיני מחזור 12, 13, ו 14 (המקביל ל- 1:30, 1:45, 10:2 שעות פוסט הפריה, בהתאמה12) ו- 3-4 h פוסט הפריה (hpf) התקבלו על-פי ההליכים המתוארים בפרוטוקול. על ידי לקיחת תמונות של האות eGFP-PCNA של כל אצווה עובר מיון, זה ניתן לתעד את שלב ומצב מחזור התא מדויק של כל עובר יחיד המשמש בניסויים במורד הזרם. תמונות דוגמא של העוברים של אוכלוסיות ממוינים מוצגים איור 1B-E. הפלט של פרוטוקול ויטמינצ'יק בחיי עיר היא ספריית נוקלאוטיד מוכנים להיות וסודרו על מכונות רצף ' הדור הבא '. למטרה זו, ריכוז ספריית הסופי לפחות 2 – 4 nM נדרש בדרך כלל. באמצעות את הכמויות המומלצות של חומר קלט, הריכוז הזה היא אמינה מושגת (טבלה 1).
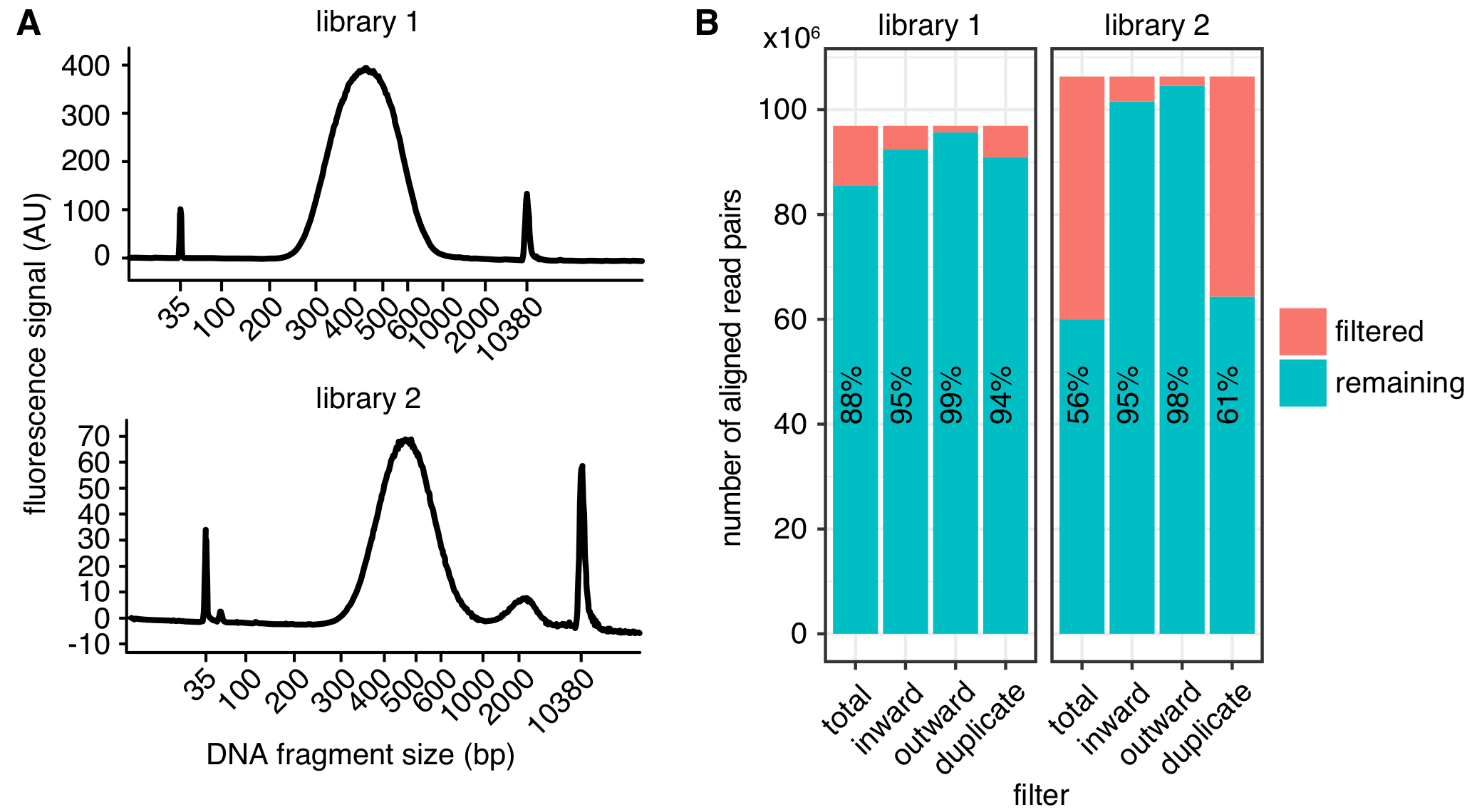
התפלגות גודל הצפוי של מקטעי דנ א לאחר גודל הבחירה היא בין 300-600 bp, עם מקסימום כ-500 bp (להבין 2A), בהתאם אמש המדויק ופרמטרים בחירת גודל. עבור רצף, אנו ממליצים לזווג-end קריאות לפחות 75 אורך bp כדי לצמצם את מספר קטעים הגבלת שאינו ניתן למיפוי הגנום. ניתן להשיג מפות ברזולוציה גבוהה עם 1-2 kb סל בגודל 400 מיליון קריאות. אנו ממליצים על קביעת רצף משכפל ביולוגי מרובים בעומק התחתון של 150 מיליון ~ קורא אחד, במקום רצף של שכפול יחיד בעומק גבוהה מאוד. זה מאפשר הערכה של וריאציית ביולוגי ומוביל מספר נמוך יותר של קריאות שנמחקו עקב כפילות PCR. לייצוג חזותי, משכפל יכול להיות משולב. לפני שתתחייב על רצף מדגם בעומק גבוהה, מומלץ לרוץ דגימות באמצעות רצף רדוד (כמה קריאות מיליון עבור דגימה) כדי לקבוע מדדי איכות ספריית בסיסיים כמו דמות 2B.
ניתוח של נתוני ויטמינצ'יק דורש מומחיות ומשאבים ביואינפורמטיקה חישובית משמעותית. כמו מבט קשוח, קריאות לזווג ממופים באופן עצמאי הגנום הפניה, במערכים וכתוצאה מכך הסינון של איכות, התמצאות, ואז מטריצה של אנשי קשר ברמה הרזולוציה, או קטע נתון סל יכול להיווצר מן מסוננים היישורים. המטריקס קשר היא הבסיס לניתוח כל עוד יותר במורד הזרם, חקר TADs, לולאות, תאים. עבור הניתוח הראשוני הקריאות רצף, מספר צינורות ביואינפורמטיקה זמינים המאפשרים עיבוד הקריאות גולמיים לתוך מטריצות קשר בלי הרבה ביואינפורמטיקה מיוחדים ידע18,19, 20,21,22,23. כיצד מתבצע ניתוח נוסף אאוט תלויה במידה רבה השאלה ביולוגי מדויק שנבחנה, וייתכן שיידרשו ניסיון משמעותי תכנות ו- scripting R או פיתון. עם זאת, מספר כלים ואלגוריתמים להתקשר TADs הם זמינים5,24,25,26,27,28, וכן תוכנה לניתוח, סקור נתונים ויטמינצ'יק בדפדפן האינטרנט וכן יישומי שולחן עבודה עצמאית29,30,31,32.
לאחר עיבוד, האיכות של הספרייה ניתן לקבוע באמצעות מדדים אחרים (איור 2B). ראשית, הקצב של PCR כפילויות, המהווה מספר זוגות קריאה ברצף הנובעים אותה המולקולה המקורי, צריך להיות נמוך ככל האפשר להגביל את כמות קריאות רצף מבוזבז. עם זאת, גם ספריות עם > שכפול PCR 40% יכולים להיות מעובדים לתוך מפות קשר איכותי אם הכפילויות מסוננים. שנית, קצב הקריאות מסוננים בשל האוריינטציה שלהם, כפי שמתואר4, באופן עקבי להיות נמוך יותר מ-10% של זוגות קריאה מיושר.
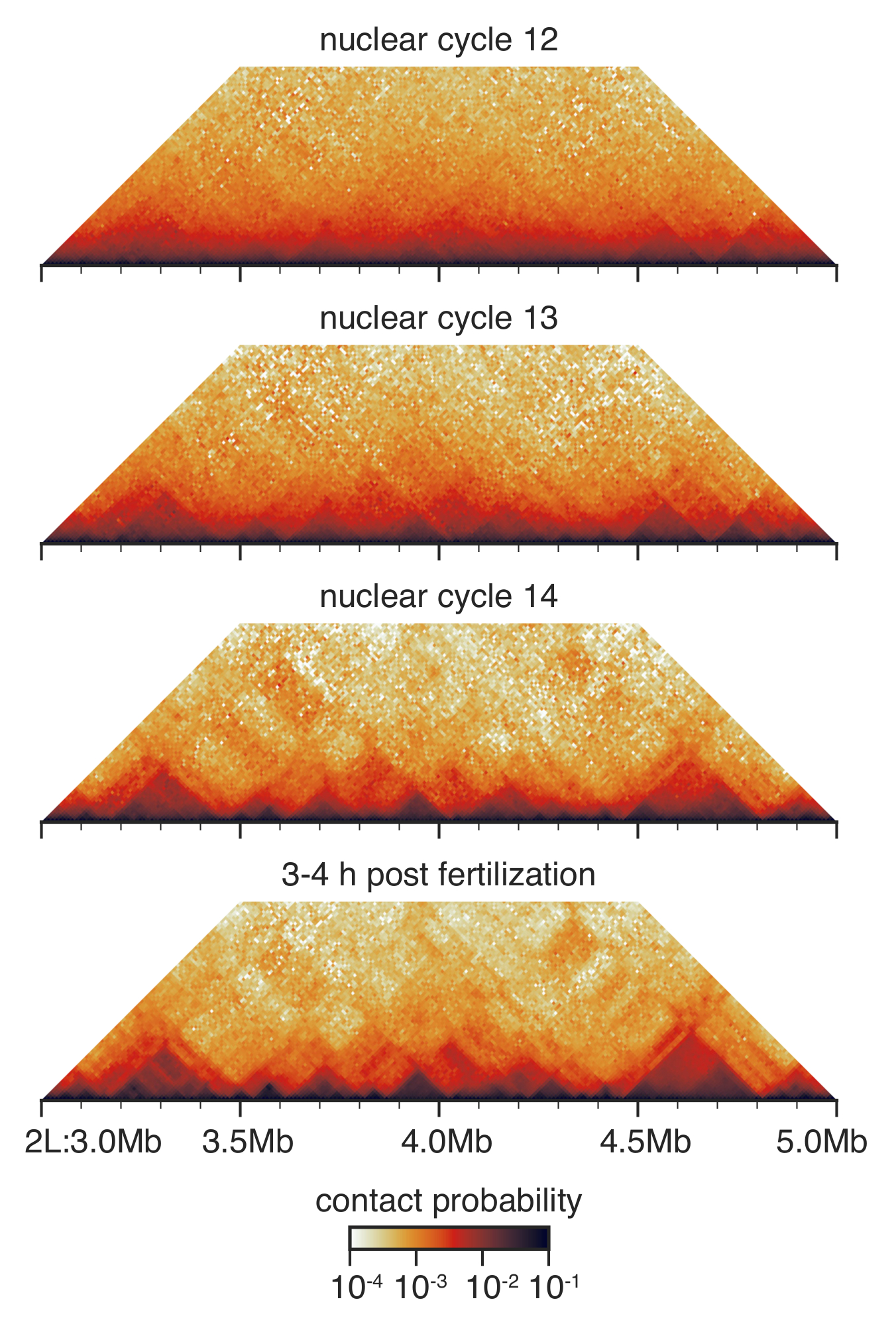
במהלך פיתוח מראש gastrular דרוזופילה בין מחזור גרעיני 12 ו 14, הארכיטקטורה גרעינית היא שופצה באופן דרסטי33 (איור 3). -גרעיני מחזור 12, TADs כמה מזוהים, ההתפלגות הכוללת של הקשר היא מאוד חלקה ללא תכונות רבות ניכרת. זה באופן דרמטי השתנה-מחזור גרעיני 13 ו-14, כאשר TADs בולטות יותר ויותר קשרים ארוכי טווח לא ספציפי מתרוקנים.

איור 1: נציג תמונות של eGFP-PCNA העוברים במהלך מיון. (א) eGFP-PCNA האיתות אוכלוסיה ממוינת של העוברים לאחר 60 דקות ואוסף הדגירה 2 h ב- 25 ° C (B-E) דוגמאות העוברים של אוכלוסיות ממוינות על הגרעין מחזור 12 (B), גרעיני מחזור 13 (ג), (14 מחזור גרעינית D), מן העוברים עוברים מיטוזה סינכרונית (E). גודל ברים = 200 מיקרומטר. אנא לחץ כאן כדי להציג גירסה גדולה יותר של הדמות הזאת.
{kind=link}

איור 2: דוגמאות בחיי עיר ויטמינצ'יק מדדי איכות הספרייה. Bioanalyzer (א) עקבות מציג את ההתפלגות בגדלים מקטע DNA מספריית ויטמינצ'יק מוצלחת (ספריית 1, העליון) וממנו ספריה מציגה לשיא של קטעים גדולים מדי עבור רצף (ספריית 2, למטה). ספריית 2 היה רציף בהצלחה, אך כמויות גדולות יותר ואפילו שברי DNA רצויה עלול להוביל רצף ירד התשואות. (B) סינון נתונים סטטיסטיים של שתי ספריות ויטמינצ'יק: המוצג הוא מספר זוגות קריאה מיושר שאינם נכללים בין ניתוח נוסף בשל קריאה כיוון ומרחק (פנימה, כלפי חוץ)4 או שכפול ה-PCR (כפולים). כל בר, מותווים מספר קריאות עובר את המסנן (שנותרו) ולהיכשל (מסונן). האחוז של קריאות עובר את המסנן בנוסף מוצג כטקסט. אנא לחץ כאן כדי להציג גירסה גדולה יותר של הדמות הזאת.
{kind=link}

איור 3: שלום-C אינטראקציה מפות מעוברים בשלבים. מפות-C אינטראקציה מנופה ברזולוציה 10 kb, מאוזן כפי שתואר לפני33. המוצג הוא אזור בכרומוזום 2 ל' אנא לחץ כאן כדי להציג גירסה גדולה יותר של הדמות הזאת.
{kind=link}
| הספרייה | הבמה | מספר העוברים | כמות ה-DNA לפני הטיה (ng) | PCR מחזורים | ספריית הסופי ריכוז (אן אם) |
| 1 | גרעיני מחזור 12 | 71 | 46 | 12 | 28.2 |
| 2 | גרעיני מחזור 12 | 46 | 40 | 12 | 22.2 |
| 3 | גרעיני מחזור 12 | 60 | 13 | 13 | 12.3 |
| 4 | גרעיני מחזור 13 | 36 | 39 | 12 | 22.2 |
| 5 | גרעיני מחזור 13 | 35 | 10 | 12 | 5.0 |
| 6 | גרעיני מחזור 13 | 48 | 18 | 12 | 8.7 |
| 7 | גרעיני מחזור 14 | 33 | 30 | 12 | 39.8 |
| 8 | גרעיני מחזור 14 | 24 | 36 | 12 | 20.4 |
| 9 | גרעיני מחזור 14 | 14 | 8 | 12 | 4.2 |
| 10 | 3-4 hpf | 17 | 30 | 12 | 24.0 |
| 11 | 3-4 hpf | 18 | 42 | 11 | 19.1 |
| 12 | 3-4 hpf | 22 | 63 | 11 | 48.4 |
טבלה 1: רשימת רצפי נציג ספריית סטטיסטיקה. עבור כל ספריה ברשימה, מספר העוברים ששימשו עבור הדור שלה, כמות ה-DNA הכולל לפני הנפתח ביוטין ו הטיה נמדדת קיוביט, המספר של ה-PCR מחזורי להשתמש הגברה של הריכוז הסופי של הספרייה רצף לאחר טיהור ובחירת גודל מצוינים.
Discussion
פרוטוקול המוצג כאן הוא יעיל מאוד יצירת מפות באיכות גבוהה של האדריכלות כרומטין ב מוקדם עוברי דרוזופילה . בהשוואה של פרוטוקול קודמות34, הגישה המתוארת כאן משתמש מעודכן בחיי עיר ויטמינצ'יק הליך5, וכתוצאה מכך עיבוד מהיר יותר, ברזולוציה גבוהה יותר, ופחות ריאגנט השימוש. ההליך הכולל כולל הפרוטוקול ויטמינצ'יק בחיי עיר צפוי לעבוד על מגוון רחב של שלבים ומערכות ניסיוני חוץ דרוזופילה. מאז הפרוטוקול יש דרישה קלט נמוך, זה יכול לשמש גם על אוכלוסיות תאים מבודדים. דרוזופילה, כאשר המשתמשים בפרוטוקול עבור עוברי מחוץ לטווח המתוארים כאן, מספר פרמטרים, בפרט הקיבעון של החומר, ייתכן שיהיה עליך להתאימם. מאז עוברי בוגרים לפתח לציפורן אטום מאוד, העלאת הריכוז של פורמלדהיד והארכת קיבוע עשוי להיות מתאים. אוסף של העוברים בשלבים שאינו גרעיני מחזור 14, בתקופת הדגירה עוברי 25 ° c בשלב 1.4 צריכים להיות מותאמים כדלקמן: גרעיני מחזור 12, 70 דקות; גרעיני מחזור 13, 90 דקות; 3-4 hpf, 3:30 שעות.
במהלך לחטיבות המחשוף 13 (שלב 1-4), צפיפות גרעינים מוכפל בערך עם כל חטיבה. הגרעינים יכול בקלות להיות מזוהה על-ידי קרינה פלואורסצנטית החיובי שלהם GFP. במהלך מיטוזה, eGFP-PCNA אינו נמצא בגרעין, האות מפוזרת בכל רחבי העובר. תכונה זו הופכת עוברי מזהים זה עוברים חטיבה המחשוף סינכרונית אפשרי. לימוד כרומטין קונפורמציה, אלה העוברים mitotic הם בדרך כלל לא רצוי, מאז הארגון mitotic של כרומטין שונה באופן דרסטי הארגון לאטמוספרה35. זה אפשרי להתאים הפרוטוקול לבחירת במיוחד העוברים העוברים חלוקה mitotic סינכרוני. במקרה זה, העוברים היחיד עם מפוזר, לא גרעיני הפצה של eGFP-PCNA צריך להישמר, כל העוברים האחרים צריכים להיות מושלך. מאז לא ניתן לקבוע את צפיפות גרעינית, חייב להיות מועסק שיטות אלטרנטיביות למניעת שלב עוברי מאת שלהם מורפולוגיה צפו ב המשודרת במיקרוסקופ אור. נוכחות של תאי מוט ואת הגרעינים בפריפריה העובר מציינים כי העובר סיים לפחות הגרעין מחזור 9, ואילו cellularization גלוי בפריפריה מציין גרעיני מחזור 1412.
שלום-C ניסויים יכול בהצלחה להתבצע באמצעות מבחר רחב של אנזימי הגבלה5. לגישות בדרך כלל להשתמש אנזימים מזהה גם 4-בסיס רצף של, כגון MboI, או אתר בסיס 6 זיהוי, כגון HindIII. היתרון של בסיס 4 חותכני על בסיס 6 חותכני הוא בכך שהם מציעים פוטנציאליים ברזולוציה גבוהה יותר, נתון מספיק עומק רצף, כיסוי אפילו יותר של הגבלת אתרים ברחבי הגנום. יש אין יתרון ברור בבחירת אחד חותך 4-בסיס מעל עוד5,23,36,37. שני אנזימים הנפוצות ביותר, MboI, DpnII, שניהם מזהה זיהוי GATC באותו האתר. DpnII הוא פחות רגישים CpG מתילציה, אשר לא יהווה בעיה בדרוזופילה. פרוטוקול המובאות כאן ניתן גם בהצלחה להשלים באמצעות DpnII כמו באנזים הגבלה. בסעיף 4.2. אנזים הגבלה מאגר יש יותאם לתאימות DpnII, על פי ההמלצות של היצרן.
אם גודל המקטע של הספרייה רצף סוטה משמעותית מהטווח המוצג באיור 2A, היווצרות אשכול במהלך רצף עשוי להיות פחות יעילה, או לכשל מוחלט. במקרה זה, יש לבדוק את התפלגות גודל לאחר הטיה, הטיית פרמטרים מותאמים בהתאם. פסגות בתחום ההפצה של שברי DNA של קטן מאוד (< 100 bp) או גדולים מאוד (> 1,000 bp) גדלים מעיד על בעיות עם בחירת גודל, כגון לשאת מעל של חרוזים או תגובת שיקוע שאמורים להיות מושלך. לעיתים קרובות ספריות אלה עם שיאים קטנים בגדלים האלה לא רצויים, כגון עדיין הם בתמונה, אחד רציף בהצלחה רק ירידה קטנה במספר אשכולות יעילות.
שיעור גבוה של שכפול PCR להימנע בגלל זה מפחית במידה רבה את מספר הקריאות רצף שמיש. הקצב של PCR כפילויות קשורה ישירות כמות החומר קלט. באמצעות קלט יותר ולכן בדרך כלל מקלה על בעיות עם שכפול ה-PCR.
המספרים גבוהים יותר הקריאות מסוננים בגלל כיוון קריאהלהבין 2B) (מציינים מספיקים לעיכול, אשר יכול להיות התוצאה של באמצעות אנזים מעט מדי, יותר מדי קלט חומר או המגון לא שלם של העוברים.
Disclosures
המחברים אין לחשוף.
Acknowledgements
מחקר זה מומן על ידי חברת מקס פלנק. C.B.H. נתמכה על ידי התחברות מאסכולת המחקר פלאנק מקס הבינלאומי – וההתערבות מולקולרית. אנו מודים שלבי בליית, אריק Wieschaus על הקו melanogaster דרוזופילה eGFP-PCNA מתן בחביבות.
Materials
| Name | Company | Catalog Number | Comments |
| Biotin-14-dATP | Life Technologies | 19524016 | |
| MboI | New England Biolabs | R0147L | |
| DNA Polymerase I Klenow Fragment | New England Biolabs | M0210L | |
| T4 DNA Ligase | Thermo Fisher | EL0012 | T4 DNA Ligase Buffer included |
| T4 DNA Polymerase | New England Biolabs | M0203L | |
| Proteinase K | AppliChem | A4392 | |
| GlycoBlue | Life Technologies | AM9516 | |
| Complete Ultra EDTA-free protease inhibitors | Roche | 5892791001 | |
| NEBNext Multiplex Oligos for Illumina (Index Primers Set 1) | New England Biolabs | E7335 | Sequencing Adaptor, Forward (unindexed) PCR primer and Reverse (indexed) PCR primer and USER enzyme used in the Library preparation section are components of this kit |
| NEBNext Ultra II DNA Library Prep Kit | New England Biolabs | E7645 | End Prep Enzyme Mix, End Prep Reaction Buffer, Ligation Enhancer, Ligation Master Mix and Polymerase Master Mix used in the Library preparation section are components of this kit |
| Covaris S2 AFA System | Covaris | ||
| DNA LoBind Tubes, 1.5 mL | Eppendorf | 0030108051 | |
| Falcon cell strainer 100 µm | Corning | 352360 | Embryo collection baskets |
| 37% formaldehyde | VWR | 437536C | |
| Heptane | AppliChem | 122062.1612 | |
| M165 FC fluorescent stereo microscope | Leica | ||
| M165 FC DFC camera | Leica | ||
| Metal micro pestle | Carl Roth | P985.1 | Used to lyse embryos in step 4.1.4 |
| RNase A | AppliChem | A3832,0050 | |
| Dynabeads MyOne Streptavidin C1 | Life Technologies | 65002 | Streptavidin coated magnetic beads |
| Ampure XP beads | Beckman Coulter | A63881 | |
| Qubit 3.0 Fluorometer | Thermo Fisher Scientific | Q33216 | |
| Qubit assay tubes | Thermo Fisher Scientific | Q32856 | |
| Qubit dsDNA HS Assay Kit | Thermo Fisher Scientific | Q32854 | |
| Phosphate buffered saline (PBS) | Sigma-Aldrich | P4417 | |
| eGFP-PCNA flies | Gift from S. Blythe and E. Wieschaus | ||
| Sodium hypochlorite 13% | Thermo Fisher | AC219255000 | |
| Triton X-100 | AppliChem | A4975 | |
| Tris buffer pH 8.0 (1 M) for molecular biology | AppliChem | A4577 | |
| NaCl | AppliChem | A2942 | |
| IGEPAL CA-630 | Sigma-Aldrich | I8896 | |
| 1.5 mL microcentrifuge tubes | Greiner Bio-One | 616201 | |
| SDS for molecular biology | AppliChem | A2263 | |
| 10x CutSmart buffer | New England Biolabs | B7204S | Restriction enzyme buffer |
| PCR Nucleotide Mix | Sigma-Aldrich | 11814362001 | Unmodified dCTP, dGTP, dTTP |
| BSA, Molecular Biology Grade | New England Biolabs | B9000S | |
| EDTA 0.5 M solution for molecular biology | AppliChem | A4892 | |
| Sodium acetate 3 M pH 5.2 | Sigma-Aldrich | S7899 | |
| DynaMag-2 Magnet | Life Technologies | 12321D | Magnetic stand |
| Intelli-Mixer RM-2L | Omnilab | 5729802 | Rotator |
| ThermoMixer F1.5 | Eppendorf | 5384000012 | Mixer |
| Small Embryo Collection Cages | Flystuff.com | 59-100 | Egg collection cage |
| Centrifuge 5424 R | Eppendorf | 5404000413 | |
| C1000 Touch Thermal Cycler | Bio-Rad | 1851148 | |
| PCR tube strips | Greiner Bio-One | 673275 | |
| NEBuffer 2.1 | New England Biolabs | B7202S | T4 DNA Polymerase buffer |
References
- Bonev, B., Cavalli, G. Organization and function of the 3D genome. Nat Rev Genet. 17 (11), 661-678 (2016).
- Lieberman-Aiden, E., et al. Comprehensive mapping of long-range interactions reveals folding principles of the human genome. Science. 326 (5950), 289-293 (2009).
- Dixon, J. R., et al. Topological domains in mammalian genomes identified by analysis of chromatin interactions. Nature. 485 (7398), 376-380 (2012).
- Jin, F., et al. A high-resolution map of the three-dimensional chromatin interactome in human cells. Nature. , (2013).
- Rao, S. S. P., et al. A 3D Map of the Human Genome at Kilobase Resolution Reveals Principles of Chromatin Looping. Cell. 159 (7), 1665-1680 (2014).
- Darbellay, F., Duboule, D. Topological Domains, Metagenes, and the Emergence of Pleiotropic Regulations at Hox Loci. Current topics in developmental biology. 116, 299-314 (2016).
- Beagan, J. A., et al. Local Genome Topology Can Exhibit an Incompletely Rewired 3D-Folding State during Somatic Cell Reprogramming. Cell stem cell. 18 (5), 611-624 (2016).
- Andrey, G., et al. Characterization of hundreds of regulatory landscapes in developing limbs reveals two regimes of chromatin folding. Genome Res. 27 (2), 223-233 (2017).
- Krijger, P. H. L., de Laat, W. Regulation of disease-associated gene expression in the 3D genome. Nature Reviews. Molecular Cell Biology. 17 (12), 771-782 (2016).
- Sexton, T., et al. Three-dimensional folding and functional organization principles of the Drosophila genome. Cell. 148 (3), 458-472 (2012).
- Ghavi-Helm, Y., et al. Enhancer loops appear stable during development and are associated with paused polymerase. Nature. 512 (7512), 96-100 (2014).
- Foe, V. E., Alberts, B. M. Studies of nuclear and cytoplasmic behaviour during the five mitotic cycles that precede gastrulation in Drosophila embryogenesis. J Cell Sci. 61, 31-70 (1983).
- Blythe, S. A., Wieschaus, E. F. Zygotic Genome Activation Triggers the DNA Replication Checkpoint at the Midblastula Transition. Cell. 160 (6), 1169-1181 (2015).
- Blythe, S. A., Wieschaus, E. F. Establishment and maintenance of heritable chromatin structure during early Drosophila embryogenesis. eLife. 5, e20148(2016).
- JoVE Science Education Database. Embryo and Larva Harvesting and Preparation. Biology I: yeast, Drosophila and C. elegans. Drosophila melanogaster. , JoVE, Cambridge, MA. (2017).
- Sicaeros, B., O'Dowd, D. K. Preparation of Neuronal Cultures from Midgastrula Stage Drosophila Embryos. Journal of Visualized Experiments. (5), (2007).
- Shermoen, A. W. Preparation of Baskets for Drosophila Egg Collections, Treatments, and Incubations. Cold Spring Harbor Protocols. (10), (2008).
- Ay, F., Noble, W. S. Analysis methods for studying the 3D architecture of the genome. Genome biology. 16 (1), 183(2015).
- Lazaris, C., Kelly, S., Ntziachristos, P., Aifantis, I., Tsirigos, A. HiC-bench: comprehensive and reproducible Hi-C data analysis designed for parameter exploration and benchmarking. BMC Genomics. 18 (1), (2017).
- Servant, N., et al. HiC-Pro: an optimized and flexible pipeline for Hi-C data processing. Genome Biology. 16 (1), (2015).
- Durand, N. C., et al. Juicer Provides a One-Click System for Analyzing Loop-Resolution Hi-C Experiments. Cell systems. 3 (1), 95-98 (2016).
- Lajoie, B. R., Dekker, J., Kaplan, N. The Hitchhiker's guide to Hi-C analysis: Practical guidelines. Methods. 72, 65-75 (2015).
- Schmitt, A. D., Hu, M., Ren, B. Genome-wide mapping and analysis of chromosome architecture. Nature Reviews. Molecular Cell Biology. 17 (12), 743-755 (2016).
- Shin, H., et al. TopDom: an efficient and deterministic method for identifying topological domains in genomes. Nucleic Acids Res. 44 (7), e70(2016).
- Kruse, K., Hug, C. B., Hernández-Rodríguez, B., Vaquerizas, J. M. TADtool: visual parameter identification for TAD-calling algorithms. Bioinformatics. 32 (20), 3190-3192 (2016).
- Lévy-Leduc, C., Delattre, M., Mary-Huard, T., Robin, S. Two-dimensional segmentation for analyzing Hi-C data. Bioinformatics. 30 (17), Oxford, England. i386-i392 (2014).
- Filippova, D., Patro, R., Duggal, G., Kingsford, C. Identification of alternative topological domains in chromatin. Algorithms for molecular biology: AMB. 9 (1), 14(2014).
- Crane, E., et al. Condensin-driven remodelling of X chromosome topology during dosage compensation. Nature. 523 (7559), 240-244 (2015).
- Durand, N. C., et al. Juicebox Provides a Visualization System for Hi-C Contact Maps with Unlimited Zoom. Cell systems. 3 (1), 99-101 (2016).
- Zhou, X., et al. Exploring long-range genome interactions using the WashU Epigenome Browser. Nature Methods. 10 (5), 375-376 (2013).
- Ramírez, F., et al. High-resolution TADs reveal DNA sequences underlying genome organization in flies. bioRxiv. , 115063(2017).
- Kerpedjiev, P., et al. HiGlass: Web-based Visual Comparison And Exploration Of Genome Interaction Maps. bioRxiv. , 121889(2017).
- Hug, C. B., Grimaldi, A. G., Kruse, K., Vaquerizas, J. M. Chromatin Architecture Emerges during Zygotic Genome Activation Independent of Transcription. Cell. 169 (2), (2017).
- Berkum, N. L., et al. Hi-C: a method to study the three-dimensional architecture of genomes. Journal of Visualized Experiments: JoVE. (39), (2010).
- Naumova, N., et al. Organization of the mitotic chromosome. Science. 342 (6161), 948-953 (2013).
- Denker, A., de Laat, W. The second decade of 3C technologies: detailed insights into nuclear organization. Genes & development. 30 (12), 1357-1382 (2016).
- Belaghzal, H., Dekker, J., Gibcus, J. H. Hi-C 2.0: An optimized Hi-C procedure for high-resolution genome-wide mapping of chromosome conformation. Methods (San Diego, Calif). 123, 56-65 (2017).
Reprints and Permissions
Request permission to reuse the text or figures of this JoVE article
Request PermissionExplore More Articles
This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. All rights reserved