
Method Article
단단히 준비 초기 초파리 배아에서 게놈 넓은 Chromatin 구조 캡처 라이브러리의 생성
요약
이 작품 설명 프로토콜 위한 고해상도 제자리에 이 C 라이브러리에서의 세대 단단히 준비 사전 gastrulation 초파리 melanogaster 태아.
초록
Chromatin의 3 차원 구조를 조사 하 고 유전자 규칙의 메커니즘에 대 한 귀중 한 통찰력을 제공 합니다. 여기, 우리는 염색 질 구조 캡처 기술에서 수행 제자리에 안녕-C에 대 한 프로토콜 설명 초파리 melanogaster 태아 인구를 벌였다. 결과 단 하나 실험에서 핵에서 발생 하는 모든 chromatin 상호 작용의 매핑을 허용 시퀀싱 라이브러리입니다. 배아 정렬 형광 스테레오 현미경 및 핵 마커를 포함 하는 유전자 변형 플라이 라인을 사용 하 여 수동으로 이루어집니다. 이 기술은, 배아 인구 정의 된 세포 주기 상태, 각 핵 사단 주기에서와 사용 하 여 매우 높은 순도로 얻을 수 있습니다. 프로토콜은 gastrulation 넘어 이전 배아 정렬에 적응 될 수 있습니다. 정렬 된 배아 제자리에 이-C에에 대 한 입력으로 사용 됩니다. 모든 실험, 도서관 준비, 시퀀싱 등 5 일 이내에 완료할 수 있습니다. 프로토콜은 낮은 입력된 요구 사항 입력된 자료로 20 blastoderm 단계 배아를 사용 하 여 안정적으로 작동 하며 최종 결과 다음 세대 시퀀싱 시퀀싱 라이브러리입니다. 시퀀싱, 후 데이터 도메인 (TAD) 구조, chromatin 루프, 및 chromatin 토폴로지가 연결에 대 한 정보를 얻기 위해 다양 한 사용할 수 있는 도구를 사용 하 여 분석할 수 있는 게놈 넓은 chromatin 상호 작용 지도로 처리할 수 있습니다. 초파리 개발 하는 동안 구획입니다.
서문
염색 질 구조 캡처 (3 C)는1핵에서 chromatin의 토폴로지를 공부 하는 매우 유용한 방법으로 떠오르고 있다. 3 C 변종 안녕 C 단일 실험2에서 핵에서 발생 하는 모든 chromatin 상호 작용의 연락처 주파수 측정 가능 안녕-C의 응용 프로그램은 chromatin 조직, TADs, 구획, 루프3,,45등의 많은 기본 원리의 발견에 중요 한 역할을 했다.
Chromatin 아키텍처 개발 및 세포 분화의 맥락에서의 연구는 점점 더 이러한 프로세스6,,78, 동안 유전자 규칙의 메커니즘을 해명 하는 데 사용 됩니다. 9. 하나 큰 관심의 모형 유기 체 초파리 melanogaster, 그 개발 및 게놈 잘 특징 이다. 그러나, 몇 가지 설정 되었습니다 조직 문화 체 외에 외부 초파리 에서 chromatin 아키텍처 조사 연구10,11. 배아에서 16-18 h 게시물 수정, TADs와 구획 포유류에서 비슷한 구조의 연상 했다 확인 된10, 그들은 초파리 배아 중 유전자 규칙에서 연주하는 역할의 질문을 제기 개발입니다. Gastrulation, 이전 개발의 초기 단계에서 특히 이러한 학문은 기술적으로 도전. Gastrulation, 전에 초파리 배아 주기12,13당 8-60 분의 매우 빠른 속도로 진행 13 동기 핵 분열을 겪는 다. 이것 이외에, 다른 단계를 구별 하기 시각적 특징의 부족 어렵게 충분 한 수량에 밀접 하 게 준비 된 배아 자료를 얻기 위해.
Chromatin 핵 주기 해상도에서 초기 초파리 개발에서 건축을 공부 하 고 수 있는 프로토콜을 개발 하기 위하여 우리는 기존 하는 두 가지 기술을 결합: 제자리에서 안녕-C를, 높은 해상도의 세대 전체를 수 있는 게놈 연락처 지도5, 그리고 배아 준비 eGFP PCNA transgene13,14표현 유전자 변형 초파리 라인을 사용 하 여. 이 transgene interphase 동안에 핵 localizes 및 유사 분열 동안 syncytial blastoderm 통해 분 광. 이 속성을 사용 하 여, 그들의 핵 밀도 의해 다른 단계와 mitotic 배아 GFP 신호의 분산에 의해 쉽게 구별이 가능 하다.
함께, 이러한 기술을 활성화 chromatin 적게 20 초파리 배아에서 고해상도 3 차원 구조를 공부 하 고. 이 프로토콜 단일 핵 사단 주기에서 배아의 인구를 수확 하 고 초파리 배아를 정렬에 대 한 지침을 포함 합니다. 더 얻은 배아에서 수행 현장에 이.를 사용 하는 방법 설명 최종 결과 다음 세대 시퀀싱 컴퓨터에서 시퀀싱을 위해 적당 한 뉴클레오티드 라이브러리입니다. 결과 연속 읽기 다음 자세한 chromatin 상호 작용 지도 전체 초파리 게놈을 다루는으로 처리할 수 있습니다.
프로토콜
1. 초파리 태아 컬렉션
참고: 동일한 배아 컬렉션 이전 게시15와 같이 수행할 수 있습니다.
- 젊은 eGFP PCNA 파리 전송 (< 1 주 이전) 계란 컬렉션 연습장 yeasted 컬렉션으로 접시16 (1% 에탄올, 1% 초 산, 및 4 %agar).
- 25 ° c.에 설정 인큐베이터 컬렉션 새 장 이동 부 화 달걀 수집 전에 1-2 일 동안 계란 생산량을 크게 향상 됩니다. 하루에 두 번 컬렉션 접시를 변경 합니다.
- 30-60 분 간격으로 수집 약 실에서 배아를 포함 하는 접시를 제거 합니다. 작은 간격 발달 단계의 엄격한 배포 하지만 적은 배아 발생. 수집 병렬로 여러 연습장에서 그렇게 하는 이상적으로 > 200 계란 30-60 분 마다 놓여 있다.
- 배아를 원하는 나이 도달할 때까지 25 ° C에서 접시를 저장. Blastoderm 단계에 대 한 배아 (핵 주기 14), 약 2 시간에 대 한 품 어.
- 외피의 2 시간 후 추가 수돗물 물 총 병에서 컬렉션 플레이트에 전체 표면을 물으로 덮여 있다. 배아를 일시 중단 하 고 부드러운 브러쉬를 사용 하 여 효 모.
- 붓고 컬렉션 접시에서 resuspended 배아 배아 컬렉션 바구니 (100 µ m 기 공 크기와 수 제 바구니17 일 잘 상업 세포 멤브레인), 물 총 병에서 추가 수돗물을 추가 필요한 경우. 이 단계에서 동시에 수집 된 모든 접시에서 배아를 결합 한다. 풀링된 샘플은 단일 일괄 처리를 나타냅니다.
- 잘 물 총 병 30에서 수돗물으로 바구니를 rinsing 하 여 태아를 씻어 s 모든 효 모 찌 꺼 기를 멀리 세척 될 때까지.
- 물에 2.5% 염소 솔루션으로 컬렉션 바구니를 배치 하 여는 Dechorionate 배아 소용돌이 의해 빛 동요는 chorion의 제거를 촉진 한다. 그들은 바구니 밖으로 해제 및 다시, 빠져들 때 솔루션의 표면에 떠 있도록 배아 충분히 소수 성까지 계속는 ~1.75–2 분 걸릴 한다.
주의: 염소 부식성입니다. 적절 한 개인 보호 장비를 착용 하십시오. 포함 하는 솔루션 < 10% 염소 일반적으로 싱크대에 삭제 수, 호스트 연구소의 규정을 확인 하시기 바랍니다. - 솔루션에서 바구니를 제거 하 고 표 백제 냄새는 더 이상 눈에 띄는 때까지 물 총 병에서 수돗물으로 깨끗이 씻어.
2. 배아 고정
참고: 최적의 고정 조건, 세제, 포름알데히드 및 고정, 기간의 주로 농도의 배아 단계에 맞게 실험적으로 결정 될 필요가. 단계 syncytial blastoderm 주위에 대 한 최종 농도 0.5% 트라이 톤 X-100 그리고 1.8% 포름알데히드의 수성 단계에서 잘 작동 합니다. 태아 단계 9 넘어 나중 단계에 대 한 추가 매개 변수 최적화 할 수 있습니다. 고정 및 정렬 하는 동안 사용 되는 모든 솔루션 protease 억제제를 포함 해야 합니다.
- 컬렉션 바구니를 반전 하 고 15 mL 원뿔 원심 튜브 위에 놓습니다. PBS-T를 분배 하는 파스퇴르 피 펫을 사용 하 여 튜브에 바구니에서 배아를 플러시 (PBS, 0.5% 트라이 톤 X-100).
- 배아 정착 하단에 볼륨을 조정 총 2 mL PBS T와 함께 하 게
- 물에 헵의 6 mL 및 37% 포름알데히드의 100 µ L를 추가 합니다.
주의: 헵 및 포름알데히드 독성 흡입 또는 피부 후 문의. 적절 한 개인 보호 장비를 착용 하 고 증기 두건에서 작동. 헵 또는 포 름 알 데히드를 포함 하는 폐기물 호스트 연구소의 규정에 따라 별도로 처리할 수 있다. - 포 름 알 데히드의 추가 후 15 분 타이머를 시작 하 고 적극적으로 흔들어 튜브 아래로 1 분 동안 직접. 수성 및 유기 단계는 폼 샴푸 같은 일관성을 결합할 것 이다.
- 교 반 하십시오 회전 믹서에 10 분까지 포름알데히드의 추가 후에.
- 튜브의 하단에 배아를 수집을 실 온에서 1 분 동안 500 x g 에서 원심.
- 전체 샴푸 같은 액체를 발음 하 고 어떤 배아를 발음 하지 않도록 주의 복용, 그것을 폐기. 샴푸 같은 상쾌한의 소량 남아 있는 문제가 발생 하지 않습니다.
- 포름알데히드, 추가 후 15 분은 포름알데히드를 끄다 125 mM 글리신과 PBS-T의 5 mL에 배아를 resuspend. 적극적으로 1 분 대를 위아래로 흔들어 섞는다.
- 1 분 동안 실 온에서 500 x g 에서 centrifuge 고 상쾌한 발음.
- 얼음 처럼 차가운 PBS-t 5 mL에 resuspending에 의해 태아를 씻어 배아 정착 및 aspirate 모든 표면에 뜨는 게.
- 세척 단계 2.10에서에서 두 번 더 반복 합니다.
- 정렬까지 얼음에 배아를 유지. 일반적으로, 그것은 정렬에 진행 하기 전에 비행 배아의 3-4 일괄 처리를 수집 하는 것이 좋습니다. 그러나, 같은 날 사용 하는 배아 정렬 되어야 한다. 얼음 이나 냉장고에 확장된 저장 변경 된 배아 형태 이끌어 낸다.
3. 배아 정렬
참고: 60-80 X 배율에서 GFP 필터를 갖춘 어떤 형광 스테레오 현미경에 수행할 수 있습니다 정렬.
- 1000 µ L 피 펫을 사용 하 여 정렬, 어두운 색의 선호에 적합 한 작은 유리 그릇에 약 100 배아의 일괄 처리를 전송 하 고 얼음에 그것을 배치.
-
핵 밀도 세포 주기 상태 (그림 1) 팁을 바늘 또는 주사기를 사용 하 여 별도 더미에 바람직한 배아를 밀어 여는 정렬 배아
- EGFP PCNA (그림 1E)의 분산, 비 핵 분포와 모든 배아를 제거 합니다. 또한, 부분적으로 비 핵 GFP 신호를 표시 하는 배아는 제거 되어야 한다.
- 정렬에 투입, 핵 주기 12, 13, 그리고 14 가이드로 그림 1 에서 그림을 사용 하 여 각 일괄 처리에서에서 참조 배아의 라인을 조립. 이 라인을 사용 하 여 그들의 단계를 결정 하기 위해 참조 배아 중 알 수 없는 단계의 배아를 일치.
- 참조 배아에 대 한 발달 단계를 확인 하려면 이미징 태아 및 거리 정보를 제공 하는 이미징 소프트웨어를 사용 하 여 2, 500 µ m2 의 영역에서 태아의 표면에 핵의 수를 계산 하 여 핵 밀도 측정 합니다.
참고: 2, 500 µ m2 의 영역에 대 한 핵의 예상된 번호는 핵 주기 12, 12 ~ 16 핵 및 핵 주기 131320 30 핵.
- 적절 한 단계에서 모든 배아 분리는 일단 설명서 및 품질 관리에 대 한 배아의 사진을 찍을. 스테레오 현미경 자체 카메라 모듈을 장착 하지 않으면, 어떤 epifluorescence 현미경 GFP 필터와 함께 사용할 수 있습니다.
- 얼음에 1000 µ L 피 펫, 신선한 관을 전송 사용 하 여 원하는 배아를 플라스틱.
- 충분 한 태아는 계획 된 실험에 대 한 정렬 될 때까지 계속 합니다. 오래 된 태아에 대 한 9, 무대 보다 일반적으로 20 배아는 안녕 C 실험 한 현장에 대 한 충분 한. 핵 주기 12, 80 배아는 좋은 출발점입니다. 이전 사이클에서 모든 사이클에 대 한 약 배아의 수를 두배로 한다.
- 1.5 mL 튜브 한 튜브는 단일 위치에 이-C에서에 대 한 충분 한 배아를 포함 하는 방식에서으로 및 분할 배아 실험. 그것은 동일한 관 전체 프로토콜 사용 됩니다 때문에 DNA의 흡착 낮은 DNA 농도에서 상당한 손실을 발생할 수 있습니다 낮은 DNA 바인딩 특성을 가진 튜브를 사용 하는 것이 좋습니다.
- 잠시 실 온에서 100 x g 에서 튜브를 회전 하 고 상쾌한을 제거 합니다. 배아를 건조 해야 가능한 동결에 대 한.
- 플래시는 액체 질소와-80 ° c.에 게 튜브를 잠수 하 여 배아를 동결
4. 제자리에서 안녕-C
- 세포의 용 해
- 얼음에 냉동된 배아로 튜브를 놓습니다.
- 얼음 처럼 차가운 세포의 용 해 버퍼 (10 mM Tris Cl pH 8.0, 10 mM NaCl, 0.2% IGEPAL CA-630, protease 억제제,에 녹은 물)의 500 µ L에 배아를 resuspend. 그런 다음 튜브의 하단에 정착 하는 배아를 1 분 기다립니다.
- 배아는 금속 마이크로 유 봉, 미리 단단히 1.5 mL microcentrifuge 튜브에 맞게 설계 된 얼음에 냉각을 사용 하 여 갈아.
- Agitating 배아를 방지 하려면는 유 봉 천천히 튜브의 바닥 접촉 될 때까지, 아래로 밀어 넣은 다음 두 방향에 (서) 두번 유 봉 회전 하 여 분쇄.
- 유 봉 아주 약간, 다시, 튜브의 하단에 밀어 들어올린 연 삭 반복.
- 4.1.3.2 10 번을 반복 하거나 배아는 완전히 lysed. 솔루션 균질 성, 그리고 배아의 잔여 더 큰 조각을 유지 해야 합니다.
- 1000 x g, 5 분, 4 ° C에서 15 분 회전을 위해 얼음에 무 균된 정지 품 어 고 상쾌한 삭제.
- 아래로 pipetting 500 µ L 얼음 세포의 용 해 버퍼에서 resuspending으로 펠 릿을 세척.
- 4.1.4에서 다시 회전 하 고 상쾌한 삭제.
- 씻어 펠 릿 100 µ L의 0.5% 나트륨 라우릴 황산 염 (SDS)에 위쪽 및 아래쪽 pipetting resuspend 열 블록에 65 ° C에서 10 분 동안 배양 하 여 핵 permeabilize. 10% 트라이 톤 X-100 그리고 120 µ L 물 50 µ L을 추가 하 여 SDS를 끄다. 튜브를 터치 하 여 혼합.
- 열 블록에 15 분 동안 37 ° C에서 품 어.
- 금지 효소 소화
- 제한 효소 버퍼 x 10와 20 U 5 U / µ L MboI의 25 µ L를 추가 합니다. 튜브를 터치 하 여 혼합.
- 약간의 동요 (750 rpm)에서 열 블록에 37 ° C에서 90 분 동안 배양 하 여 DNA를 소화.
- MboI의 또 다른 20 U를 추가 하 고 90 분 부 화를 계속.
- 20 분 동안 62 ° C에서 배양 하 여 열 비활성화 MboI
- 오버행 채우기
참고: 특정 합자 조각의 선택을 허용 biotinylated dATP와 오버행에 작성 됩니다. Biotin-dATP unligated 무뚝뚝한 끝에서 효율적으로 제거 하는 반면 결 찰 접속점에 biotin dATP exonuclease 활동의 T4 DNA 중 합 효소 (4.6 섹션)에서 보호 됩니다. 풀 다운 메뉴를 사용 하 여 streptavidin 입히는 구슬 섹션 4.7에에서 따라서 구체적으로 합자, 공상 DNA 파편에 대 한 풍요롭게.- Biotin-14-dATP 0.4 m m의 18 µ L, 2.25 µ L의 수정 되지 않은 dCTP/dGTP/dTTP 혼합 (3.3 m m 각), 그리고 5 U / µ L DNA 중 합 효소 I Klenow 파편의 8 µ L를 추가 합니다.
- 튜브를 터치 하 여 혼합 하 고 열 블록에 90 분 동안 37 ° C에서 품 어.
- 결 찰
- 추가 물 657 µ L, 10 x T4 DNA 리가 버퍼, 10%의 100 µ L의 120 µ L 트라이 톤 X-100, 6 µ L 20 mg/mL 소 혈 청 알 부 민 (BSA), 그리고 튜브를 터치 하 여 혼합의. 마지막으로 튜브를 터치 하 여 5 µ L 5 U / µ L T4 DNA 리가와 섞어를 추가 합니다.
- 회전 튜브 부드럽게 (20 rpm) 2 시간 실내 온도에.
- 5 U / µ L T4 DNA 리가의 5 µ L의 두 번째 할부를 추가 하 고 더 많은 h 2에 대 한 회전 계속.
- 5 분 동안 2500 x g 에서 핵 아래로 회전 시키십시오 그리고 상쾌한 삭제.
- DNA 추출
- 추출 버퍼 (50 mM Tris Cl pH 8.0, 50 m m NaCl, 1 mM (EDTA), Ethylenediaminetetraacetic 산 1 %SDS,에 녹은 물)의 500 µ L에 펠 릿을 resuspend 하 고 튜브를 터치 하 여 20 mg/mL 가수분해 K. 혼합의 20 µ L를 추가 합니다.
- 30 분, 1000 rpm에서 떨고 55 ° C에서 배양 하 여 단백질을 소화.
- 드-crosslink를 5 M NaCl의 130 µ L을 추가 하 고 1000 rpm에서 떨고 68 ° C에서 밤새 품 어.
- 우선적으로 낮은 DNA 바인딩 특성을 가진 새로운 2 mL 튜브에 피 펫 샘플.
- 3 M 15 mg/mL GlycoBlue의 나트륨 아세테이트 pH 5.2와 2 µ L의 0.1 x 볼륨 (63 µ L)를 추가 합니다. 반전 하 여 잘 혼합. 반전 하 여 순수한 절대 에탄올과 혼합의 1.6 x 볼륨 (1,008 µ L)를 추가 합니다.
- -80 ° C에 적어도 30 분 동안 4 ° C에서 20000 x g 15 분 원심 분리기에서 품 어. DNA 펠 릿은 종종 아주 작은, 거의 보이지 않는, 그리고 GlycoBlue의 파란 색깔 때문에 발견 될 수 있습니다.
- 제거 상쾌한 매우 신중 하 게, DNA 펠 릿이 있는 곳에서 반대쪽 벽을 따라 관으로 피 펫 팁을 이동 합니다. 작은 나머지 방울 P10 팁을 사용 하 여 튜브에서 그들을 밀고 하 여이 단계와 다음 세척 하는 동안 종종 쉽게 제거는 pipetting 그들을 밖으로 보다는 오히려.
- 70% 에탄올의 800 µ L을 추가 하 여 펠 릿을 세척. 반전 하 여 혼합 하 고 원심 20000 x g 5 분에 대 한 실 온에서 반복이 세척 한 번 이상.
- 에탄올의 모든 흔적을 제거 하 고 건조에 최대 5 분 동안 뚜껑 열고 관 서를 떠나. 일단 아무 액체, 잔여 10 mM Tris Cl pH 8.0의 50 µ L를 추가 합니다. 반복적으로 펠 릿을 solubilize DNA 위치 했다 튜브의 벽에 영역 솔루션 플라스틱.
- RNase A, 튜브, 터치 하 여 믹스 20 mg/mL의 1 µ L을 추가 하 고 요약 RNA에 15 분 동안 37 ° C에서 품 어. 샘플 지금 냉장고에서 밤새 껏 저장 또는-20 ° C에서 무기한 동결 수 있습니다.
- 제조업체의 지침에 따라 분석 결과 기반으로 하는 형광 염료를 사용 하 여 DNA의 농도 확인 하십시오. 샘플에서 DNA의 총 금액은 적어도 10 이어야 한다 ng, 그렇지 않으면 너무 작은 재료를 사용할 수 확대를 위한 라이브러리 복잡성 가능성이 낮을 것 이다. 이 경우, 시작 물자의 양을 하지 충분 했다, 또는 자료는 세포와 강 수 동안 아마도 도중에, 분실 되었다.
- Biotin 제거 및 DNA 전단
- 10 x T4 DNA 중 합 효소 버퍼의 12 µ L, 1 mM dATP의 3 µ L, 1 m m dGTP의 3 µ L 및 물 46 µ L 함께 추가 합니다. 튜브를 터치 하 여 혼합. 3 U/mL T4 DNA 중 합 효소의 5 µ L을 추가 하 고 튜브를 터치 하 여 섞어 30 분 동안 20 ° C에서 품 어.
- 반응을 중지 하 0.5 M EDTA의 3 µ L을 추가 하 고 사용 하 여 물 샘플 약 120 µ L의 볼륨을.
- DNA의 200-400 bp 쥡니다 장치 제조업체의 지침에 따라 사용 하 여 크기를 전단 테이블의 자료에 언급 된 sonicator 사용 하 여, 다음 프로그램은 적절 한: 2 주기 각 50 s, 10% 듀티, 강도 5, 200 사이클/버스트.
- Biotin 풀 다운
- 새로운 관으로 30 µ L 10 mg/mL 코팅 streptavidin 자석 구슬의 플라스틱, 마그네틱 스탠드에 그들을 분리 하 고 상쾌한 삭제.
- Vortexing에 의해 B & W 버퍼 (5 mM Tris Cl pH 7.4, 0.5 m m EDTA, 1 M NaCl,에 녹은 물) + 0.1% 트라이 톤 X-100 및 혼합 1 구슬 resuspend. 마그네틱 스탠드에 튜브를 놓고 제조사와 모델에 따라 구슬 구분 됩니다 때까지 1-5 분 대기 합니다.
- 발음 하 고 구슬이 있는 곳의 맞은편 벽을 따라 피 펫 팁을 슬라이딩 하면서 상쾌한 삭제. B & W 버퍼 (10 mM Tris Cl pH 7.4, 1 mM EDTA, 그리고 2 M NaCl) x 2의 120 µ L에 구슬 resuspend. Vortexing에 의해 혼합.
- 새로운 낮은 DNA 바인딩 튜브에 깎인된 DNA를 전송 하 고 120 µ L 2 X B & W 버퍼에 구슬의 서 스 펜 션의 vortexing에 의해 혼합. 15 분 20 rpm에서 DNA 샘플 구슬 회전.
- 마그네틱 스탠드에 구슬 하 고 상쾌한을 삭제 합니다.
- Resuspend 비즈 B & W + 0.1% x 1의 600 µ L에 Triton X-100 그리고 1000 rpm에서 떨고 2 분 동안 55 ° C에서 품 어. 분리 후 상쾌한을 삭제 합니다. 이 세척 한 번 반복 합니다.
- 비즈 10 mM Tris Cl pH 8.0의 600 µ L로 한번 세척 하 고 분리 후 상쾌한 삭제.
- 비즈 10 mM Tris Cl pH 8.0의 50 µ L에서 resuspend.
5. 시퀀싱 라이브러리 준비
참고: 모든 라이브러리 단계 완료 상업 DNA 도서관 준비에서 구성 요소를 사용 하 여 키트 ( 재료의 표참조). 그러나, 다른 장비 또는 다른 약 대체 수 있습니다. 강 수는 냉장고 보관 시 라이브러리 준비 요원에서 형성 하 경향이 있다. 그것은 따라서 모든 강 수는 시 약을 사용 하기 전에 녹는 다는 것을 확인 해야 합니다.
-
최종 수리
- 새로운 PCR 관으로 10 mM Tris Cl pH 8.0의 50 µ L에 구슬 정지를 전송 합니다.
- 최종 준비 효소 혼합의 3 µ L 및 최종 준비 반응 버퍼의 7 µ L를 추가 합니다. 아래로 pipetting으로 혼합.
- 튜브 열 cycler에 전송 하 고 다음 프로그램 실행: 20 ° C에서 30 분, 30 분, 및 4 ° c.에 보류에 대 한 65 ° C
-
어댑터 결 찰
- 결 찰 마스터 믹스, 1.5 µ M의 2.5 µ L의 30 µ L 추가 시퀀싱 어댑터 (주식에서 1.5 µ m 희석), 그리고 구슬 정지를 결 찰 증강의 1 µ L. 아래로 pipetting으로 혼합.
- 열 cycler에서 15 분 동안 20 ° C에서 품 어.
- 사용자 효소의 3 µ L를 추가 합니다. 아래로 pipetting으로 혼합.
- 열 cycler에 15 분 동안 37 ° C에서 품 어.
- 마그네틱 스탠드에 구슬 하 고 상쾌한을 제거 합니다.
- 씻어 구슬, resuspend 1 x B & W 버퍼 + 0.1%의 100 µ L에서 비즈 트라이 톤 X-100. Vortexing, 및 새로운 microcentrifuge 튜브에 의해 혼합. 마그네틱 스탠드에 구슬 하 고 상쾌한을 제거 합니다.
- 한 번 같은 버퍼의 600 µ L를 사용 하 여이 세척을 반복 합니다.
- 비즈 10 mM Tris Cl pH 8.0, vortexing에 의해 혼합의 600 µ L에서 resuspend 그리고 새로운 튜브에 비즈를 전송.
- 마그네틱 스탠드에 구슬, 상쾌한, 삭제 하 고 10 mM Tris Cl pH 8.0의 50 µ L에 비즈를 resuspend 합니다.
-
PCR 증폭
- 두 개의 PCR 튜브를 준비 하 고 각, 중 합 효소 마스터 믹스의 25 µ L, 1.5 µ L 10 µ M 앞으로 (인덱싱되지) PCR 뇌관의, 그리고 10 µ M 역 (인덱싱된) PCR 뇌관의 1.5 µ L를 혼합.
참고: 전달 (인덱싱되지) PCR 뇌관:
5'-AATGATACGGCGACCACCGAGATCTACACTCTTTCCCTACACGACGCTCTTCCGATC * T-3´.
역방향 (인덱싱된) PCR 뇌관:
5'-CAAGCAGAAGACGGCATACGAGATNNNNNNGTGACTGGAGTTCAGACGTGTGCTCTTCCGATC * T-3´. * 인덱싱된 PCR 뇌관에 phosphorothioate 채권과 Ns를 나타냅니다. - 각 관에서 아래로 pipetting으로 비드 펜션과 믹스의 22 µ L를 추가 합니다.
- 다음과 같은 프로그램을 사용 하 여 PCR을 실행: 1 분에 98 ° C (15 98 ° C s, 75 65 ° C s, 램프 1.5 ° C/s) 반복 9-12 시간, 5 분 및 4 ° c.에 보류에 대 한 65 ° C
참고: 확대 주기의 수는 경험적으로 확인할 수 있다. 그러나, 우리가 발견 이상 12 주기를 요구 하는 라이브러리 낮은 복잡성의 일반적으로 했다이 C 지도 하는 높은 품질을 초래 하지 않았다. 다른 한편으로, 라이브러리 미만 12 주기를 요구 했다 부정적인 영향을 받지 12 사이클 전체에 대 한 증폭. 따라서, 증폭의 12 사이클을 가능 하다. - 단일 microcentrifuge 튜브에 두 개의 PCR 반응 수영장, 마그네틱 스탠드에 구슬 고 상쾌한 새로운 튜브를 라이브러리에 포함 된 전송 합니다.
- 두 개의 PCR 튜브를 준비 하 고 각, 중 합 효소 마스터 믹스의 25 µ L, 1.5 µ L 10 µ M 앞으로 (인덱싱되지) PCR 뇌관의, 그리고 10 µ M 역 (인덱싱된) PCR 뇌관의 1.5 µ L를 혼합.
-
크기 선택
- Ampure XP 비드 정지를 가져올 실내 온도 믹스 떨고에 의해 잘.
- 물으로 정확 하 게 200 µ L 풀링된 PCR 반응의 볼륨을 가져와. PCR 및 자력 분리, 원래 볼륨의 일부는 일반적으로 손실 됩니다. 200 µ L를 피펫으로 설정 하 여 볼륨을 확인 하 고 반응의 전체 볼륨을 발음. 어 발음 하는 경우 더 추가할 필요가 물. 볼륨 200 µ L를 초과 하면 비례 5.4.3, 5.4.6 단계에 추가 하는 구슬의 볼륨을 조정 합니다.
참고: 괄호 안에 볼륨은 풀링된 PCR 반응의 총 볼륨은 정확 하 게 200 µ L 경우 유효 합니다. - Ampure XP 비드 현 탁 액의 0.55 x 볼륨 (110 µ L)를 추가 하 고 아래로 10 번 이상 pipetting으로 혼합.
- 5 분, 5 분 동안 마그네틱 스탠드에 별도 구슬에 대 한 실 온에서 품 어.
- 상쾌한 새 튜브를 이동 합니다. 구슬이 들어 있는 튜브를 삭제 합니다. 구슬 DNA 바인딩한 > 700 혈압이 너무 커서 시퀀싱 할 수 있는.
- 상쾌한, 추가 0.2 x 볼륨 (40 µ L, 샘플에서 0.75 x Ampure 버퍼의 총 결과) Ampure XP 비드 펜션과 10 번 위아래로 pipetting으로 혼합의.
- 5 분, 5 분 동안 마그네틱 스탠드에 별도 구슬에 대 한 실 온에서 품 어.
- DNA를 포함 하는 상쾌한 삭제 < 200 bp, 무료 뇌관, 뇌관 이합체, 및 너무 작은 조각 시퀀싱 할 게.
- 마그네틱 스탠드에 튜브를 둡니다. 구슬 세척, 700 µ L 80% 에탄올의 추가, 비드 펠 렛을 방해 하지 않도록 돌보는 및 30에 대 한 품 어 s.
- 상쾌한 삭제, 마그네틱 스탠드에서 튜브를 타고 그리고 비즈 10 mM Tris Cl pH 8.0의 100 µ L에서 resuspend. 10 번, 위아래로 pipetting으로 혼합 하 고 1 분 동안 실 온에서 품 어.
- Ampure XP 비드 현 탁 액의 0.8 x 볼륨 (80 µ L)를 추가 합니다. 10 번 위아래로 pipetting으로 혼합 하 고 5 분 동안 실 온에서 품 어. 하한값 크기 선택이 2 차 최종 라이브러리 뇌관 뇌관 이합체의 완전 무료입니다 보장 합니다.
- 5 분 동안 마그네틱 스탠드에 구슬 하 고 상쾌한을 삭제 합니다.
- 비드 펠 렛 30 80% 에탄올의 700 µ L로 두 번 씻어 s 각각 위와 같이 마그네틱 스탠드에 튜브를 떠나 있는 동안.
- 마그네틱 스탠드에 여전히 튜브, 에탄올의 모든 흔적을 제거 합니다. P10 피 펫을 사용 하 여 튜브 에탄올의 방울을 밀어 수 있습니다. 최대 5 분에 대 한 증발 잔여 에탄올을 하자.
- 마그네틱 스탠드 오프 튜브 고 비즈 10 mM Tris Cl pH 8.0의 50 µ L에서 resuspend. 10 번 위아래로 pipetting으로 혼합.
- 5 분, 다음 마그네틱 스탠드에 별도 구슬에 대 한 실 온에서 품 어.
- 신선한 튜브 상쾌한 전송. 이것은 마지막이 C 라이브러리, 계량 하 여 제조업체의 지침에 따라 다음 세대 시퀀싱 컴퓨터에서 시퀀싱 준비 이다.
결과
핵 주기 12, 13 및 14에 배아 인구를 정렬 (1:30, 해당 1:45, 그리고 2:10 시간 게시물 수정, 각각12)와 3-4 h 게시물 수정 (hpf) 프로토콜에서 설명 하는 절차에 따라 가져온. 각 정렬 된 배아의 eGFP PCNA 신호의 사진을 찍고, 여 다운스트림 실험에 사용 되는 모든 단일 배아의 정확한 단계와 세포 주기 상태를 가능 하다. 정렬 된 인구에서 배아의 예제 사진은 그림 1B-E에 표시 됩니다. 현장에 이 C 프로토콜의 출력은 뉴클레오티드 라이브러리 다음 세대 시퀀싱 컴퓨터에서 시퀀싱을 준비. 이 위해 적어도 2-4 nM의 최종 라이브러리 농도 일반적으로 필요 합니다. 입력된 자료의 권장된 금액을 사용 하 여,이 농도 안정적으로 (표 1)를달성.
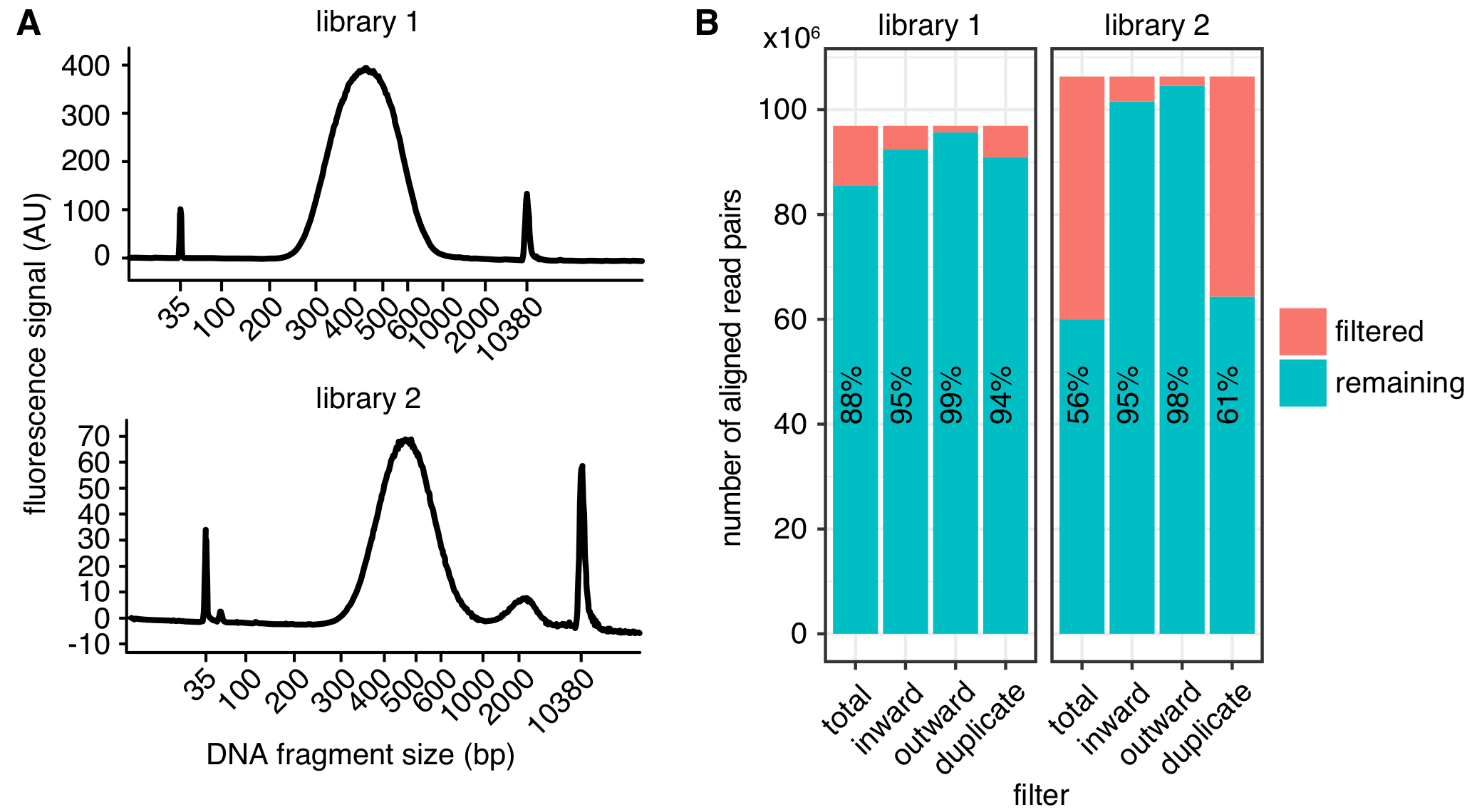
DNA 파편 크기 선택은 약 500 bp (그림 2A), 정확한 전단에 따라 크기 선택 매개 변수에서 최대 300-600 bp 사이 후의 예상된 크기 분포. 시퀀싱, 게놈에 매핑할 금지 파편의 수를 최소화 하기 위해 적어도 75 bp 길이의 쌍 간 읽기를 하는 것이 좋습니다. 1-2 kb 빈 크기와 고해상도 지도 400 백만 읽기에서 얻을 수 있습니다. 매우 높은 깊이에 단일 복제를 시퀀싱 하는 대신 각각 읽습니다 ~ 150 백만의 낮은 깊이에서 여러 생물 복제를 시퀀싱 하는 것이 좋습니다. 이 생물 학적 변이의 평가 허용 하 고 PCR 중복으로 인해 폐기 읽기의 낮은 수에 이르게. 시각적 표현에 대 한 복제는 결합할 수 있습니다. 높은 농도로 샘플 시퀀싱에 투입 하기 전에 그림 2B에서 같이 기본 라이브러리 품질 파라미터를 결정 하기 위해 얕은 시퀀싱 (샘플 당 몇 백만 읽기)을 사용 하 여 샘플을 실행 하는 것이 좋습니다.
안녕하세요 C 데이터의 분석 중요 한 전산 자원 및 생물 정보학 전문을 필요합니다. 거친 개요로 서 짝된 읽기 참조 게놈에 독립적으로 매핑되는 결과 정렬에 품질 필터링 됩니다 차례로 주어진된 빈 해상도 또는 조각 수준 연락처의 매트릭스를 생성할 수 있는 필터링에서 정렬입니다. 연락처 매트릭스 TADs, 루프, 및 구획을 탐험 하는 모든 추가 다운스트림 분석을 위한 기초 이다. 연속 읽기의 초기 분석을 위해 여러 생물 정보학 파이프라인은 사용할 수를 사용 하는 많은 전문된 생물 정보학 기술18,19, 없이 연락처 행렬에 원시 읽기의 처리 20,21,,2223. 추가 분석을 수행 하는 방법으로 연구에서 정확한 생물학 질문에 크게 의존 하 고 R 또는 파이썬 스크립팅 및 프로그래밍에서 중요 한 경험을 해야 할 수도 있습니다. 그러나, 여러 가지 도구와 알고리즘 TADs를 호출 하는 분석 하는 소프트웨어 뿐만 아니라 사용할 수 있는5,,2425,26,27,28및 웹 브라우저에서 독립 실행형 데스크톱 응용 프로그램29,30,,3132로-C 데이터를 탐험해 보세요.
일단 처리, 도서관의 품질 다른 통계 (그림 2B)를 사용 하 여 확인할 수 있습니다. 첫째, 동일한 원래 분자에서 발생 하는 시퀀스 읽기 쌍의 수는, PCR 중복의 속도 낭비 시퀀스 읽기의 양을 제한 하려면 가능한 한 낮게 이어야 한다. 그러나, 심지어와 함께 라이브러리 > 중복 필터링 하는 경우 높은 품질 연락처 지도로 40 %PCR 복제를 처리할 수 있습니다. 둘째, 그들의 방향으로 필터링 된 읽기 속도4에 설명 된 대로 일관 되 게 해야 합니다 정렬된 읽기 쌍의 10% 보다 낮은.
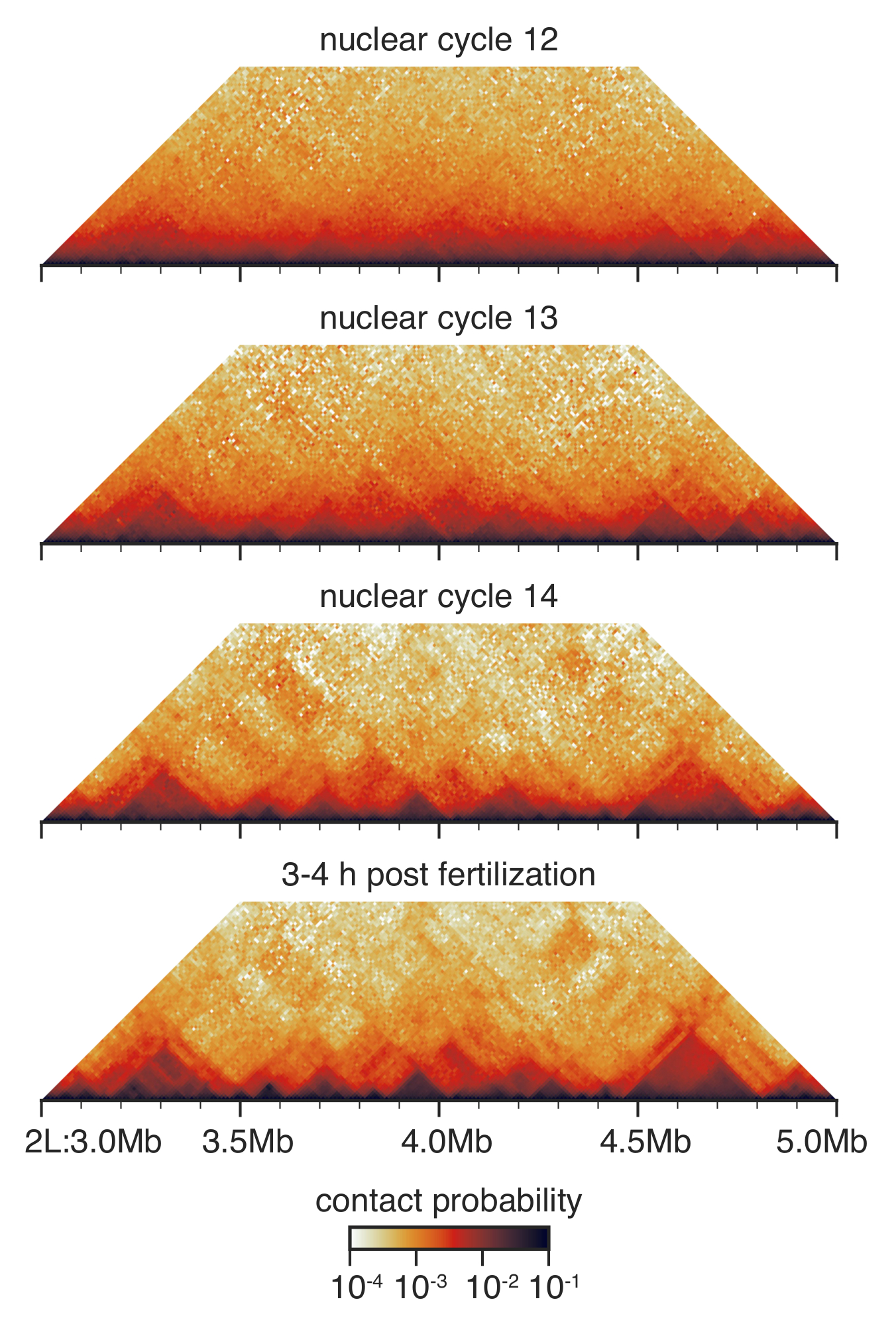
핵 주기 12와 14 사이 초파리 의 사전 gastrular 개발, 중 핵 건축은 크게 리 모델링된33 (그림 3). 핵 주기 12, 몇 TADs 감지 하 고 연락처의 전반적인 분포는 많은 뚜렷한 특징 없이 아주 부드러운. 이것은 극적으로 변경 핵 주기에서 13 및 14, TADs는 점점 저명 하 고 불특정 장거리 연락처는 고갈.

그림 1: 정렬 하는 동안 eGFP PCNA 배아의 대표 사진. (A) eGFP PCNA 신호 후 60 분 수집 및 핵 주기 12 (B), 핵에 정렬 된 인구에서 배아의 25 ° C (B-E) 예 2 h 인큐베이션 주기 13 (C), 핵 주기 14 (배아의 정렬 되지 않은 인구에서 D) 그리고 동기 유사 (E) 분열을 겪고 배아에서. 스케일 바 = 200 µ m. 이 그림의 더 큰 버전을 보려면 여기를 클릭 하십시오.
{kind=link}

그림 2: 제자리에 이 C 라이브러리 품질 통계의 예. (A) Bioanalyzer 추적 성공이 C 라이브러리 (도서관 1, 최고)와 시퀀싱 (도서관 2, 아래쪽)에 대 한 너무 큰 파편의 절정을 표시 하는 라이브러리에서 DNA 조각 크기의 분포를 보여 주는. 도서관 2는 시퀀스 성공적으로, 하지만 원치 않는 DNA 파편의 더 큰 양을 감소 시퀀싱 수율으로 이어질 수 있습니다. (B) 두이 C 라이브러리의 통계 필터링: 표시는 읽기 방향 및 거리 (안쪽, 바깥쪽)4 또는 PCR 이중 (중복) 추가 분석에서 제외 하는 정렬 된 읽기 쌍의 수입니다. 각 막대에서 읽기 (남은) 필터 통과 및 실패 (필터링 됨)의 수가 그려집니다. 읽기는 필터를 전달의 비율 또한 텍스트로 표시 됩니다. 이 그림의 더 큰 버전을 보려면 여기를 클릭 하십시오.
{kind=link}

그림 3: 안녕 C 상호 작용 지도 준비 된 배아에서. 안녕하세요 C 상호 작용 지도 10 kb 해상도 범주화 하 고33전에 설명한 대로 균형. 표시는 염색체 2에 지역 L. 이 그림의 더 큰 버전을 보려면 여기를 클릭 하십시오.
{kind=link}
| 라이브러리 | 단계 | 배아의 수 | (Ng)을 기울이기 전에 금액 DNA | PCR 주기 | 마지막 도서관 농도 (nM) |
| 1 | 핵 주기 12 | 71 | 46 | 12 | 28.2 |
| 2 | 핵 주기 12 | 46 | 40 | 12 | 22.2 |
| 3 | 핵 주기 12 | 60 | 13 | 13 | 12.3 |
| 4 | 핵 주기 13 | 36 | 39 | 12 | 22.2 |
| 5 | 핵 주기 13 | 35 | 10 | 12 | 5.0 |
| 6 | 핵 주기 13 | 48 | 18 | 12 | 8.7 |
| 7 | 핵 주기 14 | 33 | 30 | 12 | 39.8 |
| 8 | 핵 주기 14 | 24 | 36 | 12 | 20.4 |
| 9 | 핵 주기 14 | 14 | 8 | 12 | 4.2 |
| 10 | 3-4 hpf | 17 | 30 | 12 | 24.0 |
| 11 | 3-4 hpf | 18 | 42 | 11 | 19.1 |
| 12 | 3-4 hpf | 22 | 63 | 11 | 48.4 |
표 1: 대표 시퀀싱 도서관 통계 목록. 목록 생성, biotin 풀 다운 및 큐 비트에 의해 측정 기울이기 전에 총 DNA의 금액에 대 한 사용 된 배아의 수에서에서 각 라이브러리에 대 한 PCR의 수 확대, 그리고 시퀀싱 라이브러리의 최종 농도 대 한 사용 주기 정화 및 크기 선택 후 표시 됩니다.
토론
여기에 제시 된 프로토콜은 초기 초파리 배아에 높은-품질 지도 chromatin 아키텍처의 생성에 매우 효과적입니다. 이전 프로토콜34에 비해, 설명 하는 방법은 여기는 최신 제자리에 C 절차5, 빠른 처리, 높은 해상도, 그리고 시 약 사용에 더 적은 결과로 사용 합니다. 제자리에 이 C 프로토콜을 포함 하 여 전반적인 절차는 다양 한 단계와 외에 초파리실험 시스템에서 작동 하도록 예정입니다. 프로토콜은 낮은 입력된 요구, 이후는 고립 된 세포 인구에 또한 사용 될 수 있습니다. 초파리에서 범위 밖에 배아에 대 한 프로토콜을 사용 하 여 여기에 설명 된, 몇 가지 매개 변수 때 특히 소재, 고정 해야 조정 합니다. 이후 이전 배아 개발 높게 불 침투성 표 피, 포 름 알 데히드의 농도 제기 하 고 고정 연장 될 수 있습니다. 핵 주기 14 이외의 단계에서 배아의 컬렉션에 대 한 단계 1.4에서에서 25 ° C에서 배아의 부 화 시간이 다음과 같이 조정 될 필요가: 핵 주기 12, 70 분; 핵 주기 13, 90 분; 3-4 hpf, 3시 30분
13 분열 분열 (단계 1-4), 중 핵 밀도 대략 각 부서와 복식. 핵은 그들의 밝은 GFP 형광에 의해 쉽게 식별할 수 있습니다. 유사 분열, 동안 eGFP PCNA는 핵에 위치한 되지 않습니다 그리고 신호 태아에 걸쳐 분산 됩니다. 이 특징은 식별 배아 가능한 동기 분열 분열을 겪고 있다. 염색 질 구조를 공부에 대 한 이러한 mitotic 배아 없습니다 일반적으로 바람직한, chromatin의 mitotic 기구 interphase 조직35보다 크게 다르기 때문. 특히 동기 mitotic 사단을 겪고 배아를 선택 하는 프로토콜에 맞게 가능 하다. 이 경우에, 유일한 배아 eGFP PCNA의 분산, 비 핵 분포 유지 되어야 한다, 그리고 모든 다른 배아를 폐기 해야 합니다. 이후 핵 밀도 결정할 수 없는 단계 배아 전송된 가벼운 현미경 검사 법에서 그들의 형태에 의해 대체 방법은 사용 해야 합니다. 극 세포 및 배아 주변에서 핵의 존재 반면 주변에 보이는 cellularization 나타냅니다 핵 주기 1412태아가 적어도 핵 주기 9, 완료 되었음을 나타냅니다.
안녕하세요 C 실험은 다양 한 제한 효소5를 사용 하 여 성공적으로 수행 수 있습니다. 현재 접근은 일반적으로 어느 4 기반 시퀀스, MboI, HindIII 등 6 기반 인식 사이트 등을 인식 하는 효소를 사용 합니다. 6 자료 커터 4 기반 커터의 장점은 그들은 충분 한 시퀀싱 깊이 그리고 더 조차 범위 제한 사이트의 게놈에서 주어진 더 높은 잠재적인 해상도 제공 하는 이다. 또 다른5,23,,3637에 한 4 기반 커터를 선택에 명확한 이점이 있다. 두 개의 가장 일반적으로 사용 되 효소, MboI 및 DpnII, 둘 다 동일한 GATC 인식 사이트 인식. DpnII는 CpG 메 틸 화, 초파리에서 관심사의에 덜 민감합니다. 여기에 제시 된 프로토콜 수 있습니다 또한 성공적으로 완료 제한 효소로 DpnII를 사용 하 여. 섹션 4.2에. 제한 효소와 버퍼 제조 업체의 권장 사항에 따라 DpnII의 호환성을 위해 조정 될 수 있다.
시퀀싱 라이브러리의 조각 크기 그림 2A에 표시 된 범위에서 크게 벗어나는, 시퀀싱 하는 동안 클러스터 형성 덜 효율적일 수 있습니다 또는 완전히 실패. 이 경우에, 전단 후 크기 분포를 확인 한다 하 고 적절 하 게 조정 매개 변수를 기울이기. 매우 작은 DNA 파편의 분포에 봉우리 (< 100 bp) 또는 매우 큰 (> 1000 bp) 크기 구슬 또는 폐기 해야하는 상쾌한 이상 크기 선택, 수행 등으로 문제를 나타냅니다. 종종 이러한 바람직하지 않은 크기로 한 사진된는 여전히 같은 작은 봉우리 이러한 라이브러리 시퀀싱 클러스터링 효율에만 작은 감소와 성공적으로.
PCR 복제의 높은 속도이 획기적으로 사용 가능한 시퀀스 읽기 수를 줄일 수 있기 때문에 피해 야 한다. PCR 중복의 속도 입력된 자료의 양을 직접 관련이 있습니다. 더 많은 입력을 사용 하 여 따라서 일반적으로 완화 한다 PCR 중복 문제를.
읽기 때문에 읽기 방향그림 2B) (필터링 수가 부족 한 소화, 너무 작은 효소, 너무 많이 입력된 자료, 또는 태아의 불완전 한 균질을 사용 하 여 결과 될 수 있는 나타냅니다.
공개
저자는 공개 없다.
감사의 말
이 연구는 최대 Planck 학회에 의해 투자 되었다. C.B.H. 분자 의학 국제 최대 Planck 연구 학교-에서 장학금에 의해 지원 되었다. 우리 친절 하 게 제공 eGFP PCNA 초파리 melanogaster 라인에 대 한 셸 비 블 라와 에릭 Wieschaus 감사 합니다.
자료
| Name | Company | Catalog Number | Comments |
| Biotin-14-dATP | Life Technologies | 19524016 | |
| MboI | New England Biolabs | R0147L | |
| DNA Polymerase I Klenow Fragment | New England Biolabs | M0210L | |
| T4 DNA Ligase | Thermo Fisher | EL0012 | T4 DNA Ligase Buffer included |
| T4 DNA Polymerase | New England Biolabs | M0203L | |
| Proteinase K | AppliChem | A4392 | |
| GlycoBlue | Life Technologies | AM9516 | |
| Complete Ultra EDTA-free protease inhibitors | Roche | 5892791001 | |
| NEBNext Multiplex Oligos for Illumina (Index Primers Set 1) | New England Biolabs | E7335 | Sequencing Adaptor, Forward (unindexed) PCR primer and Reverse (indexed) PCR primer and USER enzyme used in the Library preparation section are components of this kit |
| NEBNext Ultra II DNA Library Prep Kit | New England Biolabs | E7645 | End Prep Enzyme Mix, End Prep Reaction Buffer, Ligation Enhancer, Ligation Master Mix and Polymerase Master Mix used in the Library preparation section are components of this kit |
| Covaris S2 AFA System | Covaris | ||
| DNA LoBind Tubes, 1.5 mL | Eppendorf | 0030108051 | |
| Falcon cell strainer 100 µm | Corning | 352360 | Embryo collection baskets |
| 37% formaldehyde | VWR | 437536C | |
| Heptane | AppliChem | 122062.1612 | |
| M165 FC fluorescent stereo microscope | Leica | ||
| M165 FC DFC camera | Leica | ||
| Metal micro pestle | Carl Roth | P985.1 | Used to lyse embryos in step 4.1.4 |
| RNase A | AppliChem | A3832,0050 | |
| Dynabeads MyOne Streptavidin C1 | Life Technologies | 65002 | Streptavidin coated magnetic beads |
| Ampure XP beads | Beckman Coulter | A63881 | |
| Qubit 3.0 Fluorometer | Thermo Fisher Scientific | Q33216 | |
| Qubit assay tubes | Thermo Fisher Scientific | Q32856 | |
| Qubit dsDNA HS Assay Kit | Thermo Fisher Scientific | Q32854 | |
| Phosphate buffered saline (PBS) | Sigma-Aldrich | P4417 | |
| eGFP-PCNA flies | Gift from S. Blythe and E. Wieschaus | ||
| Sodium hypochlorite 13% | Thermo Fisher | AC219255000 | |
| Triton X-100 | AppliChem | A4975 | |
| Tris buffer pH 8.0 (1 M) for molecular biology | AppliChem | A4577 | |
| NaCl | AppliChem | A2942 | |
| IGEPAL CA-630 | Sigma-Aldrich | I8896 | |
| 1.5 mL microcentrifuge tubes | Greiner Bio-One | 616201 | |
| SDS for molecular biology | AppliChem | A2263 | |
| 10x CutSmart buffer | New England Biolabs | B7204S | Restriction enzyme buffer |
| PCR Nucleotide Mix | Sigma-Aldrich | 11814362001 | Unmodified dCTP, dGTP, dTTP |
| BSA, Molecular Biology Grade | New England Biolabs | B9000S | |
| EDTA 0.5 M solution for molecular biology | AppliChem | A4892 | |
| Sodium acetate 3 M pH 5.2 | Sigma-Aldrich | S7899 | |
| DynaMag-2 Magnet | Life Technologies | 12321D | Magnetic stand |
| Intelli-Mixer RM-2L | Omnilab | 5729802 | Rotator |
| ThermoMixer F1.5 | Eppendorf | 5384000012 | Mixer |
| Small Embryo Collection Cages | Flystuff.com | 59-100 | Egg collection cage |
| Centrifuge 5424 R | Eppendorf | 5404000413 | |
| C1000 Touch Thermal Cycler | Bio-Rad | 1851148 | |
| PCR tube strips | Greiner Bio-One | 673275 | |
| NEBuffer 2.1 | New England Biolabs | B7202S | T4 DNA Polymerase buffer |
참고문헌
- Bonev, B., Cavalli, G. Organization and function of the 3D genome. Nat Rev Genet. 17 (11), 661-678 (2016).
- Lieberman-Aiden, E., et al. Comprehensive mapping of long-range interactions reveals folding principles of the human genome. Science. 326 (5950), 289-293 (2009).
- Dixon, J. R., et al. Topological domains in mammalian genomes identified by analysis of chromatin interactions. Nature. 485 (7398), 376-380 (2012).
- Jin, F., et al. A high-resolution map of the three-dimensional chromatin interactome in human cells. Nature. , (2013).
- Rao, S. S. P., et al. A 3D Map of the Human Genome at Kilobase Resolution Reveals Principles of Chromatin Looping. Cell. 159 (7), 1665-1680 (2014).
- Darbellay, F., Duboule, D. Topological Domains, Metagenes, and the Emergence of Pleiotropic Regulations at Hox Loci. Current topics in developmental biology. 116, 299-314 (2016).
- Beagan, J. A., et al. Local Genome Topology Can Exhibit an Incompletely Rewired 3D-Folding State during Somatic Cell Reprogramming. Cell stem cell. 18 (5), 611-624 (2016).
- Andrey, G., et al. Characterization of hundreds of regulatory landscapes in developing limbs reveals two regimes of chromatin folding. Genome Res. 27 (2), 223-233 (2017).
- Krijger, P. H. L., de Laat, W. Regulation of disease-associated gene expression in the 3D genome. Nature Reviews. Molecular Cell Biology. 17 (12), 771-782 (2016).
- Sexton, T., et al. Three-dimensional folding and functional organization principles of the Drosophila genome. Cell. 148 (3), 458-472 (2012).
- Ghavi-Helm, Y., et al. Enhancer loops appear stable during development and are associated with paused polymerase. Nature. 512 (7512), 96-100 (2014).
- Foe, V. E., Alberts, B. M. Studies of nuclear and cytoplasmic behaviour during the five mitotic cycles that precede gastrulation in Drosophila embryogenesis. J Cell Sci. 61, 31-70 (1983).
- Blythe, S. A., Wieschaus, E. F. Zygotic Genome Activation Triggers the DNA Replication Checkpoint at the Midblastula Transition. Cell. 160 (6), 1169-1181 (2015).
- Blythe, S. A., Wieschaus, E. F. Establishment and maintenance of heritable chromatin structure during early Drosophila embryogenesis. eLife. 5, e20148(2016).
- JoVE Science Education Database. Embryo and Larva Harvesting and Preparation. Biology I: yeast, Drosophila and C. elegans. Drosophila melanogaster. , JoVE, Cambridge, MA. (2017).
- Sicaeros, B., O'Dowd, D. K. Preparation of Neuronal Cultures from Midgastrula Stage Drosophila Embryos. Journal of Visualized Experiments. (5), (2007).
- Shermoen, A. W. Preparation of Baskets for Drosophila Egg Collections, Treatments, and Incubations. Cold Spring Harbor Protocols. (10), (2008).
- Ay, F., Noble, W. S. Analysis methods for studying the 3D architecture of the genome. Genome biology. 16 (1), 183(2015).
- Lazaris, C., Kelly, S., Ntziachristos, P., Aifantis, I., Tsirigos, A. HiC-bench: comprehensive and reproducible Hi-C data analysis designed for parameter exploration and benchmarking. BMC Genomics. 18 (1), (2017).
- Servant, N., et al. HiC-Pro: an optimized and flexible pipeline for Hi-C data processing. Genome Biology. 16 (1), (2015).
- Durand, N. C., et al. Juicer Provides a One-Click System for Analyzing Loop-Resolution Hi-C Experiments. Cell systems. 3 (1), 95-98 (2016).
- Lajoie, B. R., Dekker, J., Kaplan, N. The Hitchhiker's guide to Hi-C analysis: Practical guidelines. Methods. 72, 65-75 (2015).
- Schmitt, A. D., Hu, M., Ren, B. Genome-wide mapping and analysis of chromosome architecture. Nature Reviews. Molecular Cell Biology. 17 (12), 743-755 (2016).
- Shin, H., et al. TopDom: an efficient and deterministic method for identifying topological domains in genomes. Nucleic Acids Res. 44 (7), e70(2016).
- Kruse, K., Hug, C. B., Hernández-Rodríguez, B., Vaquerizas, J. M. TADtool: visual parameter identification for TAD-calling algorithms. Bioinformatics. 32 (20), 3190-3192 (2016).
- Lévy-Leduc, C., Delattre, M., Mary-Huard, T., Robin, S. Two-dimensional segmentation for analyzing Hi-C data. Bioinformatics. 30 (17), Oxford, England. i386-i392 (2014).
- Filippova, D., Patro, R., Duggal, G., Kingsford, C. Identification of alternative topological domains in chromatin. Algorithms for molecular biology: AMB. 9 (1), 14(2014).
- Crane, E., et al. Condensin-driven remodelling of X chromosome topology during dosage compensation. Nature. 523 (7559), 240-244 (2015).
- Durand, N. C., et al. Juicebox Provides a Visualization System for Hi-C Contact Maps with Unlimited Zoom. Cell systems. 3 (1), 99-101 (2016).
- Zhou, X., et al. Exploring long-range genome interactions using the WashU Epigenome Browser. Nature Methods. 10 (5), 375-376 (2013).
- Ramírez, F., et al. High-resolution TADs reveal DNA sequences underlying genome organization in flies. bioRxiv. , 115063(2017).
- Kerpedjiev, P., et al. HiGlass: Web-based Visual Comparison And Exploration Of Genome Interaction Maps. bioRxiv. , 121889(2017).
- Hug, C. B., Grimaldi, A. G., Kruse, K., Vaquerizas, J. M. Chromatin Architecture Emerges during Zygotic Genome Activation Independent of Transcription. Cell. 169 (2), (2017).
- Berkum, N. L., et al. Hi-C: a method to study the three-dimensional architecture of genomes. Journal of Visualized Experiments: JoVE. (39), (2010).
- Naumova, N., et al. Organization of the mitotic chromosome. Science. 342 (6161), 948-953 (2013).
- Denker, A., de Laat, W. The second decade of 3C technologies: detailed insights into nuclear organization. Genes & development. 30 (12), 1357-1382 (2016).
- Belaghzal, H., Dekker, J., Gibcus, J. H. Hi-C 2.0: An optimized Hi-C procedure for high-resolution genome-wide mapping of chromosome conformation. Methods (San Diego, Calif). 123, 56-65 (2017).
재인쇄 및 허가
JoVE'article의 텍스트 или 그림을 다시 사용하시려면 허가 살펴보기
허가 살펴보기더 많은 기사 탐색
This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. 판권 소유