
Method Article
2 في 1: انجذاب تطهير لتحليل مواز للبروتين-بروتين والبروتين-المستقلب المجمعات من خطوة واحدة
* These authors contributed equally
In This Article
Summary
تفاعلات البروتين البروتين والبروتين-المستقلب حاسمة بالنسبة لجميع الوظائف الخلوية. هنا، نحن وصف بروتوكول يسمح تحليل موازية لهذه التفاعلات مع بروتين في الاختيار. لدينا بروتوكول الأمثل للثقافات الخلية النباتية، ويجمع بين انجذاب تطهير مع البروتين الشامل القائم على قياس الطيف الكتلي وكشف المستقلب.
Abstract
وتخضع العمليات الخلوية بالتفاعلات بين الجزيئات البيولوجية مثل البروتينات ونواتج الأيض والأحماض النووية. بينما يتم التحقيق في تفاعلات البروتين البروتين (PPI) لا الجدة، نهج تجريبي يهدف إلى وصف التفاعلات الذاتية البروتين-المستقلب (PMI) تشكل تطورا الأخيرة بدلاً من ذلك. وهنا، نقدم بروتوكول يسمح لتوصيف المتزامنة PPI والصليب الأحمر من بروتين اختيارهم، ويشار إليها كطعم. لدينا بروتوكول كان الأمثل لخلية نبات الثقافات ويجمع بين انجذاب تطهير (AP) مع الطيف الكتلي (مللي ثانية)-على أساس الكشف عن البروتين والمستقلب. باختصار، أولاً يتم تفكيك خطوط نبات المحورة وراثيا، معربا عن البروتين الطعم تنصهر فيها علامة تقارب، للحصول على خلاصة خلوية أصلية. وتستخدم الأجسام المضادة العلامة لهدم البروتين والمستقلب الشركاء من البروتين الطعم. المجمعات تنقية تقارب يتم استخراج استخدام الميثيل خطوة واحدة ثالثي-بوتيل الاثير (مثيل ثالثي بوتيل الإيثر)/الميثانول/المياه الأسلوب. بينما فصل نواتج الأيض إلى القطبية أو في مرحلة مسعور، يمكن العثور على البروتينات في بيليه. ثم يتم تحليل نواتج الأيض والبروتينات بواسطة فائقة الأداء السائل اللوني-قياس الطيف الكتلي (أوبلك--مرض التصلب العصبي المتعدد أو أوبلك-MS/MS). متجه فارغة (EV) مراقبة خطوط تستخدم لاستبعاد المغلوطة. والميزة الرئيسية لأن البروتوكول أنه يمكن تحديد شركاء البروتين والمستقلب البروتين المستهدف بالتوازي في ظروف القرب الفسيولوجية (الخلوية ليستي). طريقة عرض واضحة وسريعة، ويمكن تكييفها بسهولة للنظم البيولوجية خلاف الثقافات الخلية النباتية.
Introduction
الأسلوب الموصوفة هنا يهدف إلى تحديد الشركاء المستقلب والبروتين بروتين الاختيار في ظروف شبهفي فيفو ليساتي الخلوية. كان قد تردد أن العديد من نواتج الأيض أكثر مما يتسم اليوم وظيفة تنظيمية هامة1. والايضات يمكن أن تعمل كمفاتيح البيولوجية، تغيير في النشاط ووظائف الترجمة على مستقبلات بروتينات2،،من34. وكانت أساليب اختراق عدة، مما يتيح تحديد الهوية من PMI في فيفو أو في ظروف شبهفي فيفو ، في العقد الماضي نمواً5. يمكن فصل النهج المتاحة إلى مجموعتين. وتضم المجموعة الأولى التقنيات التي تبدأ ب طعم المستقلب المعروفة بغية تعويض البروتين رواية الشركاء. وتشمل أساليب التقارب اللوني6والمخدرات تقارب الاستقرار المستهدفة تستجيب المقايسة7، البروتيوميات الكيماوي8والبروتين الحرارية التنميط9. تتكون المجموعة الثانية من أسلوب واحد يبدأ مع بروتين معروفة من أجل تحديد يغاندس جزيء صغير10،11.
AP يقترن المستندة إلى MS ليبيدوميكس استخدمت لتحليل البروتينات الدهنية المجمعات في. Saccharomyces cerevisiae12 كنقطة انطلاق، المؤلفين استخدام سلالات الخميرة معربا عن 21 الإنزيمات المشاركة في تخليق ارجوستيرول الحيوي وتنصهر مؤنزم 103 على تنقية جنبا إلى جنب-تقارب العلامة (الصنبور). تم العثور على 70% الإنزيمات و 20% من مؤنزم لربط مختلف يغاندس مسعور، تسليط الضوء في شبكة التفاعل المعقدة البروتينات الدهنية.
سابقا، يمكن أن نظهر أن، وبالمثل للدهون، المركبات القطبية وشبه القطبية أيضا تظل ملتزمة بمجمعات البروتين المعزولة من ليساتيس الخلوية13. وبناء على هذه النتائج، قررنا أن تحسين وكالة اسوشييتد برس الأسلوب سبق نشرة10،11 للخلايا النباتية ومجمعات ماء14. ولهذا الغرض، استخدمنا ناقلات الصنبور الموصوفة بلين فإن وآخرون 2010، استخدمت بنجاح في مصنع PPI الدراسات15. لتقصير الفترة الزمنية اللازمة للحصول على خطوط محوره وراثيا، قررنا نبات خلية الثقافات. نحن العاملين خطوة واحدة الميثيل ثالثي-بوتيل الاثير، (مثيل ثالثي بوتيل الإيثر)/الميثانول/المياه استخراج أسلوب، مما يتيح وصف البروتينات (بيليه)، والدهون (المرحلة العضوية)، وماء الأيض (المرحلة المائية)16 في واحد تنقية تقارب التجربة. وأدخلت خطوط التحكم EV استبعاد إيجابيات كاذبة، مثل البروتينات ملزمة للعلامة وحدها. وكدليل على مفهوم نحن معلم ثلاثة (من خمسة) نوكليوزيد diphosphate مؤنزم الموجودة في جينوم نبات (NDPK1-NDPK3). من بين النتائج الأخرى، يمكن أن نظهر أن NDPK1 يتفاعل مع الجلوتاثيون S-ترانسفيراز والجلوتاثيون. وبالتالي يمكن أن نثبت أن تعرض NDPK1 إلى جلوتاثيونيليشن14.
خلاصة القول، هو أداة هامة لوصف البروتين-بروتين وشبكات التفاعل جزيء صغير البروتين-البروتوكول المقدم ويشكل تقدما كبيرا على مدى الأساليب القائمة.
Protocol
يمكن العثور على إعداد من نبات المعدلة وراثيا خلية ثقافة الخطوط، بما في ذلك شروط الاستنساخ والتحول والتحديد، والنمو في17. لاحظ أن خطوط التحكم EV ينصح بتصحيح لايجابيات كاذبة. قبل التجربة، تؤكد overexpression البروتين الطعم بتحليل لطخة غربية، مثل استخدام الأجسام المضادة مفتش ضد الجزء زاي-البروتين من علامة تقارب جنبا إلى جنب. من المهم فصل وسائط النمو من المواد ثقافة الخلية النباتية.
1-إعداد المواد النباتية الخلية قبل التجربة
- تنمو ثقافة خط overexpressing البروتين الفائدة18خلية PSB-لام ألف-التمويل .
- إعداد مسمو المتوسطة، التي تتضمن 4.43 غ/لتر مسمو مختلطة مع السكروز 30 غرام/لتر. ضبط الأس الهيدروجيني للمخزن المؤقت إلى 5.7 مع 1 م كوه والاوتوكلاف الحل. قبل التجربة، الملحق المتوسطة مع حمض نافثالينيسيتيك α 0.5 ملغ/لتر والكينيتين 0.05 مغ/لتر 50 ميكروغرام/مل كاناميسين.
- زراعة النباتات تحول الثقافات الخلية في 50 مل من مسمو المتوسطة في قارورة 100 مل على شاكر منصة مداري مع الانفعالات لطيف (130 لفة في الدقيقة). تنمو الخلايا في قاعة ثقافة في 20 درجة مئوية وكثافة الضوء يساوي μmol 80 م-2 s-1.
- خلايا فرعية في وسائط جديدة كل 7 أيام، تمييع منهم 01:10.
- جمع الخلايا في مرحلة النمو لوغاريتمي استخدام زجاج قمع جنبا إلى جنب مع فراغ مضخة، استخدام شبكة نايلون كعامل تصفية. التفاف التغلغل في رقائق الألومنيوم وتجميدها في نيتروجين سائل.
تنبيه: تذكر أن النتروجين السائل باردة للغاية. معالجة غير صحيحة يمكن أن تسبب الحروق. ارتداء معدات الوقاية الشخصية المناسبة، بما في ذلك حرارياً معزول القفازات، ونظارات واقية، ومعطف معمل.
2. اضغط على البروتوكول
ملاحظة: الخطوة التالية هو مقتبس من مايدا et al. عام 201411 ولين Van et al. 201117.
- مجانسة المواد ثقافة الخلية النباتية المحصودة والمجمدة استخدام مطحنة خلاط (2 دقيقة في 20 هرتز) أو قذائف هاون ومدقة للحصول على مسحوق ناعم. الكوة على الجيل الثالث 3g من الأرض المادية (المقابلة لحوالي 90 ملغ بروتين الكلي) كل عينة. تجنب ذوبان العينة أثناء هذه الخطوة باستخدام النتروجين السائل المبرد مسبقاً المعدات.
ملاحظة: مخزن المواد النباتية البرية في أنبوب 50 مل في –80 درجة مئوية إلى بداية إجراء وكالة اسوشييتد برس. - تريتوراتي العينة في قذيفة هاون النيتروجين السائل-بريكوليد مع 3 مل من المخزن المؤقت لتحلل المثلج (0.025 م تريس – HCl pH 7.5; 0.5 "م كلوريد الصوديوم" 1.5 مم مجكل20.5 مم DTT؛ و 1 مم NaF؛ ومم 1 غ3فو4؛ 100 × مثبطات البروتياز التجارية المخفف كوكتيل; 1 مم بمسف) حتى يذوب بالمواد. عندما يذوب العينة، ينتقل فورا إلى الخطوة التالية.
ملاحظة: تعد الطازجة تحلل المخزن المؤقت. إدخال نماذج فارغة في هذه الخطوة. لا ينصح المنظفات كما أنها يمكن أن تسبب مشاكل في الكشف عن مرض التصلب العصبي المتعدد. - لإزالة الحطام الخلوية، تقسيم المواد إلى 2 مل ميكروسينتريفوجي الأنابيب وأجهزة الطرد المركزي في 20,817 س ز لمدة 10 دقائق في 4 درجات مئوية. جمع 3 مل من واضحة في أنبوب 15 مل مخروطية أجهزة الطرد مركزي.
- أثناء خطوة الطرد المركزي، حجته الخرز مفتش-سيفاروسي. اليكووت ميليلتر 100 من الخرز كل عينة وغسلها مع 1 مل من تحلل المخزن المؤقت. الدوامة ريسوسبيند الخرز وفلاش-سبين. تجاهل المخزن المؤقت تحلل وكرر الخطوة مرتين. ريسوسبيند الخرز في 400 ميليلتر من تحلل المخزن المؤقت.
- إضافة الخرز إلى المحطة التي تم جمعها ليساتي واحتضان المخلوط على عجلة دوارة ح 1 في 4 درجات مئوية.
- نقل الخليط في محقن مجتمعة عن طريق كاب اللوير لوك مع عمود دوران مع تصفية المسام حجم 35 ميكرون. تطبيق الضغط لتمرير من خلال. سوف تظل الخرز مع مجمعات المرفقة في عامل التصفية، في حين سوف تذهب من خلال.
ملاحظة: بشكل اختياري، استخدام نظام متشعبة فراغ. جعل معينة لتطبيق ضغط لطيف حتى لا يؤدي إلى إلحاق الضرر الخرز. - تغسل حبات في البداية مع 10 مل من الغسيل المخزن المؤقت (0.025 م تريس – HCl pH 7.5; 0.5 م كلوريد الصوديوم)، ومن ثم مع 1 مل من المخزن المؤقت شطف (10 ملم تريس – HCl درجة الحموضة 7.5؛ 150 مم كلوريد الصوديوم؛ يدتا 0.5 مم؛ 1000 x المخفف E64 و 1 مم بمسف). أداء الغسيل استخدام حقنه موصولة إلى عمود أو فراغ النظام المتعددة.
ملاحظة: عند استخدام نظام متشعبة فراغ جعل معينة لتطبيق ضغط لطيف حتى لا يؤدي إلى إلحاق الضرر الخرز. - احتضان الخرز مع 400 ميليلتر من شطف المخزن المؤقت الذي يحتوي على 50 ش من نسخة محسنة من التبغ أحفر حوزتي الفيروسات (أكتيف). استخدام الجدول شاكر 1,000 لفة في الدقيقة لمدة 30 دقيقة في 16 درجة مئوية.
ملاحظة: تذكر لاستخدام المكونات لإغلاق العمود أسفل إضافة شطف المخزن المؤقت. - إضافة جزء إضافي (50 ش) إنزيم في العمود واحتضانها الخليط مدة 30 دقيقة المقبلة تحت نفس، والشروط الموصوفة أعلاه،.
- جمع النذرة في أنبوب ميكروسينتريفوجي 2 مل بالطرد المركزي (1 دقيقة، 20,817 س ز) أو بفراغ المتشعبة. لإزالة المجمعات الباقية، يعرض خطوة شطف إضافي استخدام 200 ميليلتر من شطف المخزن المؤقت.
ملاحظة: تخزين العينة في –20 درجة مئوية أو –80 درجة مئوية، أو الانتقال إلى الخطوة استخراج البروتين والمستقلب فورا. ذوبان الجليد عينات مجمدة في الجليد.
3. تحليل لطخة غربية
- لتأكيد وجود الطعم استخدام البروتين في النذرة جمعت 10 ميليلتر من النذرة المحتوية على البروتين – المستقلب لإجراء تحليل لطخة غربية والحزب الديمقراطي الصربي – صفحة. لتحديد بروتين الفائدة، استخدم الماوس، الابتدائي أضداد البروتين ملزمة ستريبتافيدين (1: 200)، جزء من علامة الحنفية المتبقية بعد انشقاق حوزتي TEV، كما هو موضح في لين Van et al. 201117. بعد ذلك، استخدم الماعز الثانوي الماوس المضادة الأجسام المضادة مقترنة ببرنامج الصحة الإنجابية.
4-المستقلب واستخراج البروتين
ملاحظة: يتم تكييف هذا البروتوكول من جيافاليسكو et al. 201116.
ملاحظة: من هذه الخطوة فصاعدا استخدام الحلول UPLC – ماجستير – الصف.
- أضف 1 مل من الميثيل ثالثي-بوتيل الاثير (مثيل ثالثي بوتيل الإيثر)/الميثانول/الماء المذيب (3:1:1) إلى النذرة المجمعة وخلط العينة بانعكاس. تأكد من أن يتم تبريده المذيب إلى –20 درجة مئوية قبل الخطوة الاستخراج.
تنبيه: مثيل ثالثي بوتيل الإيثر والميثانول من المواد الضارة. نفذ الخطوة الاستخراج تحت غطاء الدخان وارتداء معدات الحماية الشخصية المناسبة، مثل القفازات. - إضافة حل 1:3 مل 0.4 من الميثانول: المياه لكل عينة ومزج محتوى العينة بانعكاس.
ملاحظة: النتائج مكملات المخلوط مع الماء: الميثانول الحل في المرحلة الفاصلة. المرحلة العليا يحتوي على الدهون والمرحلة السفلي يحتوي على نواتج الأيض القطبية وشبه القطبية، ويمكن العثور على البروتينات في بيليه. - إلى مراحل منفصلة، والطرد المركزي العينة في س 20,817 ز لمدة 2 دقيقة في درجة حرارة الغرفة، ثم جمع في المرحلة العليا لقياسات الدهون (لم تفعل في هذا البروتوكول) باستخدام ماصة معالجة سائل دليل سعة تخزين 1 مل.
- إضافة 0.2 مل من الميثانول ومزيج من انعكاس.
- الطرد المركزي العينة في 20,817 س ز 2 دقيقة في الرايت، ثم جمع في مرحلة القطبية للقياسات المستقلب (المركبات القطبية وشبه القطبية). لتفادي إزعاج بيليه البروتين، اترك حوالي 50 ميليلتر من الطور السائل في الجزء السفلي من الأنبوب.
- العينات التي تم جمعها الجافة للقياسات المستقلب بين عشية وضحاها في مبخر الطرد مركزي. تجنب الإفراط تجفيف حبيبات البروتين عن طريق إزالة عينات من المبخر بعد 30 – 60 دقيقة.
ملاحظة: تخزين العينات في –20 درجة مئوية أو –80 درجة مئوية، أو تشرع فورا في إعداد من البروتينات لتحليل LC – MS/MS.
5-إعداد عينات لتحليل البروتين
ملاحظة: هذه الخطوة مقتبسة من أولسن et al. 200419 والدليل التقني "مزيج" التربسين/Lys-ج (انظر الجدول للمواد).
- إجراء الهضم الأنزيمي للعينة.
تنبيه: المذيبات المستخدمة أثناء الهضم الأنزيمي والملحي للعينة الضارة. تعمل تحت غطاء الدخان وارتداء معدات الحماية الشخصية المناسبة، مثل القفازات.- حل بيليه البروتين في 30 ميليلتر من تمسخ طازجة المخزن المؤقت (40 مم بيكربونات الأمونيوم التي تحتوي على اليوريا ال/6 م م 2، درجة الحموضة 8). لتحقيق أفضل البروتين الذوبان، القيام بخطوة سونيكيشن 15 دقيقة. كرر الخطوة حتى يذوب بيليه.
- الطرد المركزي العينة في 20,817 س ز لمدة 10 دقائق في 4 درجات مئوية، ثم نقل المادة طافية إلى أنبوب ميكروسينتريفوجي جديدة.
- تحديد تركيز البروتين باستخدام مقايسة البروتين برادفورد.
- لمزيد من التحليل، الكوة وحدة تخزين ما يعادل 100 ميكروغرام من البروتين وملء نموذج ما يصل إلى 46 ميليلتر مع تمسخ المخزن المؤقت.
- إضافة إلى ميليلتر عينة 2 من المخزن المؤقت الحد الطازجة (DTT حله في ح2س 50 مم) واحتضانها لمدة 30 دقيقة في درجة حرارة الغرفة.
- علاج العينة مع 2 ميليلتر الألكلة الطازجة المخزن المؤقت (150 مم إيودواسيتاميدي حله في المخزن المؤقت بيكربونات الأمونيوم 40 مم) واحتضان المخلوط في الظلام لمدة 20 دقيقة في درجة حرارة الغرفة.
- تمييع العينة مع 30 ميليلتر من 40 مم بيكربونات الأمونيوم المخزن المؤقت وإضافة 20 ميليلتر من "مزيج" ليس/التربسين.
- بعد ح 4 الحضانة عند 37 درجة مئوية، يؤدي إلى تمييع العينة مع 300 ميليلتر من 40 مم بيكربونات الأمونيوم المخزن المؤقت.
- تواصل مع الحضانة بين عشية وضحاها في 37 درجة مئوية.
- أسيديفي العينة مع حوالي 20 ميليلتر من حمض تريفلورواسيتيك 10% (تفا) للحصول على درجة الحموضة < 2. فحص عينة الأس الهيدروجيني باستخدام شريط الأس الهيدروجيني.
ملاحظة: تخزين العينة في –20 درجة مئوية أو المضي قدما إلى الخطوة التالية.
- تحلية هضم البروتينات.
ملاحظة: يفضل استخدام نظام متشعبة فراغ. تجنب الإفراط في التجفيف العمود.- شطف عمود C18 جمعية مهندسي البترول (انظر الجدول للمواد) مع 1 مل من 100% ميوه وثم مع 1 مل من 80% الاسيتو الانيتريل (ACN) الذي يحتوي على 0.1% تفا المخفف في الماء. خطوات الاستخدام، هنا وفي زيادة البروتين الملحي، نظام متشعبة فراغ تسريع العملية. تجنب الإفراط في التجفيف العمود.
- حجته العمود بغسله مرتين مع 1 مل من 0.1% تفا المخفف في الماء.
- تحميل العينة على العمود. أنبوب شطف مع ميليلتر 200 إضافية من 0.1% تفا ونقل الحل في العمود. تشغيل الحلول من خلال العمود.
- أغسل العمود مرتين مع 1 مل من 0.1% تفا.
- الوت الببتيدات ديسالتيد من العمود مع 800 ميليلتر من 60% تزاول، حل تفا 0.1%. الجاف للكسر التي تم جمعها في مبخر الطرد مركزي، تجنب الإفراط تجفيف الجزء من البروتين عن طريق إزالة عينات من المبخر بعد 30 – 60 دقيقة.
ملاحظة: تخزين العينات في –20 درجة مئوية أو –80 درجة مئوية أو فورا للانتقال إلى الخطوة التالية. في الخطوات اللاحقة، الاحتفاظ بعينات على الجليد.
6-قياس إعداد عينات البروتين باستخدام أوبلك – MS/MS.
ملاحظة: قبل القياسات البروتين و metabolomic، تصفية (حجم المسام 0.2 ميكرون) وديغا كافة المخازن المؤقتة باستخدام مضخة فراغ ح 1.
- بيليه الببتيد ريسوسبيند المجففة المخزنة في أنبوب ميكروسينتريفوجي 2 مل في 50 ميليلتر من المخزن المؤقت ج (3% v/v تزاول، حمض الفورميك 0.1% v/v) باستخدام ماصة المناولة اليدوية السائل سعة تخزين 200 ميليلتر. Sonicate العينات عن 15 دقيقة في حمام الموجات فوق الصوتية مع 35 كيلو هرتز تردد الموجات فوق الصوتية.
تنبيه: تزاول وحمض الفورميك من المواد الضارة. تعمل تحت غطاء الدخان وارتداء معدات الحماية الشخصية المناسبة، مثل القفازات. - الطرد المركزي العينة في 20,817 س ز لمدة 10 دقائق في 4 درجات مئوية، ثم نقل 20 ميليلتر من المادة طافية على قنينة زجاج.
- الببتيدات هضمها منفصلة باستخدام عمود عكس مرحلة C18 متصلاً لوني سائل واكتساب الأطياف الشامل باستخدام مطياف كتلة.
- فصل في العمود 3 ميليلتر من العينة استخدام معدل تدفق 300-nl/دقيقة. لمرحلة الجوال باستخدام المخزن المؤقت جيم ودال (63% v/v تزاول، حمض الفورميك 0.1% v/v)، تشكيل تدرج تكثف من 3% تزاول على 15% تزاول أكثر من 20 دقيقة ومن ثم إلى 30% تزاول على مدى 10 دقيقة القادمة.
ملاحظة: تخزين بقية العينة في –20 درجة مئوية أو –80 درجة مئوية تصل إلى بضعة أشهر. قبل قياس البروتين، وتجميد العينة على الجليد. - تغسل الملوثات لمدة 10 دقائق استخدام 60% تزاول وحجته العمود مع 5 ميليلتر من المخزن المؤقت ج قبل القياس للعينة التالي.
- الحصول على أسلحة الأطياف باستخدام الأسلوب MS/MS تعتمد على البيانات مع مجموعة القرار مبلغ 000 70، الهدف AGC أيونات6 3e، وقت حقن الحد الأقصى من 100 مللي ثانية، وتتراوح بين 300 و 1600 m/z. الحصول على الحد الأقصى من 15 MS/MS مسح في القرار 17,500، الهدف AGC 1e5، وقت حقن الحد الأقصى من 100 مللي ثانية، أونديرفيل نسبة 20 في المائة، مع نافذة عزلة من 1.6 m/z و m/z تتراوح ما بين 200 إلى 2000. تمكين ابيكس الزناد (ق 6 – 20)، استبعاد دينامية مجموعة إلى 15 ثانية، واستبعاد تهمة 1 و > 5.
- فصل في العمود 3 ميليلتر من العينة استخدام معدل تدفق 300-nl/دقيقة. لمرحلة الجوال باستخدام المخزن المؤقت جيم ودال (63% v/v تزاول، حمض الفورميك 0.1% v/v)، تشكيل تدرج تكثف من 3% تزاول على 15% تزاول أكثر من 20 دقيقة ومن ثم إلى 30% تزاول على مدى 10 دقيقة القادمة.
7-تجهيز بيانات البروتين
- تحميل قاعدة بيانات البروتين التمويل نبات أحدث من http://www.uniprot.org/وتشمل قاعدة البيانات الملوث. تحليل البيانات الأولية التي تم الحصول عليها من LC – MS يتم تشغيلها باستخدام ماكسكوانت مع الببتيد أندروميدا متكاملة محرك البحث باستخدام الإعداد الافتراضي مع تمكين لفق تطبيع20،،من2122. العثور على معلومات مفصلة حول المعلمات المستخدمة في الجدول S1.
- فتح ملف الإخراج "البروتين groups.txt". لمزيد من التحليل، بتصفية المجموعات البروتين المحددة مع اثنين على الأقل من الببتيدات فريدة من نوعها. قم بإزالة مجموعات البروتين يحددها ماكسكوانت الملوثات المحتملة وتصفية التمويل (أ) البروتينات (أراث في فاستا رؤوس الأعمدة) موجود في قاعدة البيانات.
- لاختبار أهمية إثراء البروتين بين العينات، استخدام كثافات لفق تطبيع وأداء مزاوج، ثنائي الطرف الطالب لاختبار t متبوعاً بتصحيح مقارنة متعددة (علىسبيل المثال بينجاميني & هوشبيرغ اكتشاف خطأ سعر (فرانكلين روزفلت) تصحيح أو تصويب بونفيروني).
- حساب قيمة p بمقارنة كثافات لفق الحصول على مراقبة EV و NDPK1. تصفية كافة القيم غير محدد. فرز فالقيم في ترتيب تصاعدي، واستخدام البرنامج النصي للبحث والتطوير أو آلة حاسبة على الإنترنت (علىسبيل المثال، https://www.sdmproject.com/utilities/?show=FDR) لحساب تصحيح فرانكلين روزفلت. تصفية قيم فرانكلين روزفلت أقل من 0.1.
ملاحظة: النظر في شكل تحليل البيانات مناسبة للبحث. للدراسات الكمية (تحليل لإثراء البروتين بين العينات) استخدام قيمة "لفق الكثافة"، بينما للبحوث النوعية (وجود أو عدم وجود بروتين معين) اختر قيمة "الكثافة". - بتصفية المجموعات البروتين التي أكثر وفرة في NDPK1 مقارنة بمراقبة EV. تحديد ترجمة شركاء المحتملين من البروتين باستخدام قاعدة بيانات سوبا23 والصحيح للبروتين المشترك المترجمة مع NDPK1.
- حساب قيمة p بمقارنة كثافات لفق الحصول على مراقبة EV و NDPK1. تصفية كافة القيم غير محدد. فرز فالقيم في ترتيب تصاعدي، واستخدام البرنامج النصي للبحث والتطوير أو آلة حاسبة على الإنترنت (علىسبيل المثال، https://www.sdmproject.com/utilities/?show=FDR) لحساب تصحيح فرانكلين روزفلت. تصفية قيم فرانكلين روزفلت أقل من 0.1.
8-قياس العينات المحتوية على مرحلة القطبية باستخدام أوبلك--مرض التصلب العصبي المتعدد.
- ريسوسبيند المجففة المرحلة القطبية من الخطوة 4، 5 في 200 ميليلتر من الماء و sonicate العينة لمدة 5 دقائق.
- الطرد المركزي العينة في 20,817 س ز لمدة 10 دقائق في 4 درجات مئوية، ثم نقل المادة طافية على قنينة زجاج.
ملاحظة: تخزين بقية العينة في –20 درجة مئوية أو –80 درجة مئوية لتصل إلى عدة أشهر. قبل القياس metabolomic، تجميد العينة على الجليد. - القيام بخطوة انفصال باستخدام أوبلك بالإضافة إلى العمود عكس المرحلة C18 واكتساب الأطياف الشامل مع مرض التصلب العصبي المتعدد.
- تحميل على ميليلتر العمود 2 للعينة الواحدة وحقن لكل وضع التأين (الإيجابية والسلبية) وفصل الكسر باستخدام 400 معدل التدفق ميليلتر في الدقيقة. لإنشاء التدرج مطلوب لقياس المستقلب، إعداد حل المرحلة المتنقلة كما يلي: المخزن المؤقت من (حمض الفورميك 0.1% في ح2س) والمخزن المؤقت ب (0.1% حمض الفورميك في ACN).
- والايضات منفصلة في 400 ميليلتر/دقيقة والتدرج التالي: 1 دقيقة 99% من المخزن المؤقت A، 11-مين التدرج الخطي من 99 ٪ من المخزن المؤقت إلى 60 في المائة من المخزن المؤقت، 13-مين التدرج الخطي من 60 في المائة من المخزن المؤقت إلى 30% من المخزن المؤقت، 15-مين التدرج الخطي من 30% من المخزن المؤقت إلى 1% المخزن المؤقت A، اضغط تركيز 1% حتى 16 دقيقة ابتداء من 17 دقيقة، استخدام التدرج الخطي من 1% من المخزن المؤقت بأن 99 في المائة من المخزن المؤقت إعادة "توازن أ" العمود لمدة 3 دقائق مع تركيز 99% من المخزن المؤقت A قبل القياس للعينة التالي.
- الحصول على الأطياف الشامل الذي يغطي النطاق الشامل بين 100 و 1500 m/z مع قرار تعيين إلى 000 25 وتحميل وقت مقيد إلى 100 هدف AGC تعيين السيدة 1e6، الجهد الشعرية إلى 3kV مع غمد الغاز تدفق والغاز مساعدة قيمة من 60 و 20 ، على التوالي. تعيين درجة الحرارة الشعرية إلى 250 درجة مئوية والمصفاة الجهد إلى 25V.
9-تجهيز جميع البيانات
- عملية جمع تشروماتوجرامس المكتسبة من كلا أوضاع التأين. استخدم برنامج لاستخراج كتلة مقابل نسبة (m/z) والاحتفاظ بالوقت (RT) وكثافة قمم المرتبطة بها، مثل البرمجيات التجارية (انظر الجدول للمواد) أو بديلة24.
- بدء تجهيز البرمجيات بالنقر المزدوج فوق ملف.exe
- إنشاء سير عمل جديدة، والبحث عن نشاط "تحميل من ملف" ونقل هذا النشاط من "السحب والإفلات" إلى مساحة فارغة في سير العمل. اضغط على هذا النشاط بواسطة الزر الأيمن للماوس وفتح إعدادات لهذا النشاط.
- في علامة التبويب التي تحتوي على إعدادات "الجنرال"، تعيين اسماً للتجربة في حقل "الاسم" وبعد ذلك انقر فوق "تحديد الملفات والمجلدات" ووضع علامة تشروماتوجرامس الخام.
- في علامة التبويب التي تحتوي على إعدادات "خيارات متقدمة"، قم بتعيين "قطع البيانات الشخصية" لكثافة 0. انقر فوق "تطبيق" أو "موافق".
- البحث عن وإضافة نشاط "اكتساح البيانات". اضغط على هذا النشاط بواسطة الزر الأيمن للماوس وفتح إعدادات لهذا النشاط.
- في علامة التبويب التي تحتوي على إعدادات "الجنرال"، وضع علامة "Centroid البيانات" و "بيانات MS/MS". إزالة كافة البيانات MS/MS بتحديد "كل شيء" في لجنة الاختيار.
- البحث عن وإضافة نشاط "Chromatogram الكيميائية الضوضاء الطرح". اضغط على هذا النشاط بواسطة الزر الأيمن للماوس وفتح إعدادات لهذا النشاط.
- في علامة التبويب التي تحتوي على إعدادات "الجنرال"، وضع علامة "تجانس Chromatogram" وتعيين عدد من مسح إلى "3" و "مقدر" إلى "التحرك في المتوسط". تعيين "RT نافذة" لمسح 51، "قوانتيلي" إلى 50%، والطرح "أسلوب" وكثافة 750 "العتبة".
- في علامة التبويب التي تحتوي على إعدادات "خيارات متقدمة"، وضع علامة "إزالة هيكل RT" وتعيين "الحد الأدنى لطول RT" إلى 5 بالأشعة.
- في علامة التبويب التي تحتوي على إعدادات "خيارات متقدمة"، وضع علامة "m/z إزالة هيكل" وتعيين "الحد الأدنى m/z طول" إلى 3 نقاط.
- البحث عن وإضافة نشاط "محاذاة الرايت تشروماتوجرام". اضغط على هذا النشاط بواسطة الزر الأيمن للماوس وفتح إعدادات لهذا النشاط.
- في علامة التبويب التي تحتوي على إعدادات "عام"، تعيين "محاذاة مخطط" إلى "بيرويس محاذاة قاعدة الشجرة" و "RT البحث الفاصل" إلى 0.5 دقيقة.
- في علامة التبويب التي تحتوي على إعدادات "خيارات متقدمة"، استخدام المعلمات الافتراضية.
- البحث عن وإضافة نشاط "الكشف عن ذروة" في مجموعة "تشروماتوجرام" من أنشطة. اضغط على هذا النشاط بواسطة الزر الأيمن للماوس وفتح إعدادات لهذا النشاط.
- في علامة التبويب التي تحتوي على إعدادات "عام"، تعيين "خلاصة نافذة" إلى 0.09 دقيقة، "أقصى حجم" الحد الأدنى إلى 0.03 دقيقة، "دمج المسافة القصوى" إلى 5 نقاط و "دمج الاستراتيجية" إلى "مراكز". في "ذروة RT تقسيم" تعيين مربع "نسبة الفجوة/الذروة" إلى 50%.
- في علامة التبويب التي تحتوي على إعدادات "خيارات متقدمة"، تعيين "تجانس الإطار" إلى 5 نقاط، صقل "العتبة" إلى 80% والاتساق "العتبة" إلى 1. تعيين "حساب مركز" ك "مرجحة كثافة" مع "كثافة" العتبة المحددة في 70%.
- البحث عن وإضافة نشاط "تجميع النظائر" في مجموعة "تشروماتوجرام" من أنشطة. اضغط على هذا النشاط بواسطة الزر الأيمن للماوس وفتح إعدادات لهذا النشاط.
- في علامة التبويب التي تحتوي على إعدادات "عام"، تعيين "التسامح RT" إلى 0.015 دقيقة و "m/z التسامح" إلى 5 جزء في المليون.
- في علامة التبويب التي تحتوي على إعدادات "الظرف المناسب"، تعيين "أسلوب" كما "لا شكل القيد" و "التأين" ك "بروتونيشن (على طريقة إيجابية) و" ديبروتونيشن "(بالنسبة للوضع السلبي). تعيين "الحد الأدنى والحد الأقصى تهمة" إلى 1 و 4، على التوالي.
- في علامة التبويب التي تحتوي على إعدادات "خيارات متقدمة"، استخدام المعلمات الافتراضية.
- البحث عن وإضافة نشاط "عامل تصفية المفرد".
- لتصدير نتائج معالجة البيانات، البحث عن وإضافة نشاط "المحلل" في "تصدير" مجموعة من الأنشطة.
- في علامة التبويب التي تحتوي على إعدادات "الجنرال"، تعيين "نوع" كما "المجموعات" و "الملحوظ" وصفه "قد لخص كثافة". اختر "مخصص الوجهة" وحدد دليل ملف التصدير.
- في علامة التبويب التي تحتوي على إعدادات "خيارات متقدمة"، استخدام المعلمات الافتراضية.
- إضافة تعليق توضيحي ميزات الشامل باستخدام بيانات مرجعية-مجمع داخلية.
- تحليل واحد أو عدة MS الصف الإشارة-مجمع باستخدام "استخدام" أوبلك-السيدة نفس الأسلوب LC-مرض التصلب العصبي المتعدد لتحليل مركبات الإشارة ووالايضات تنقية يشترك مع البروتين للفائدة.
ملاحظة: لهذه الدراسة، تم تحليل والمستخدمة كمرجع-مجمع مكتبة مجموعة من ديبيبتيديس تقريبا 300. - استخدام تحليل البرمجيات (انظر الجدول للمواد) فتح ملف تشروماتوجرام الخام والبحث عن m/z محددة و RT المرتبطة بقياس مرجعية-مجمع (انظر دليل المستخدم).
ملاحظة: تختلف نواتج الأيض الثانوية في أنواع من التأين. التحقق من وجود المشتركة أدوكتس التي تبحث عن أيون يساوي كتلة M + 1.007276 و M-1.007276 و 18.033823 M + M + 22.989218 ل [م-ح]، [M + H]، [M + NH4] و [م + Na]، على التوالي. - تصدير جدول البيانات استخدام لفتح ملف "المحلل" التي تم الحصول عليها بعد تجهيز تشروماتوجرامس والبحث عن أيون محددة الشامل. قارن RT ميزة كتلة تقاس في التجربة و RT المجمع الإشارة. السماح بانحراف 0.005 دا لم/ض ودقيقة 0.1 الرايت
- تحليل واحد أو عدة MS الصف الإشارة-مجمع باستخدام "استخدام" أوبلك-السيدة نفس الأسلوب LC-مرض التصلب العصبي المتعدد لتحليل مركبات الإشارة ووالايضات تنقية يشترك مع البروتين للفائدة.
- لاختبار أهمية إثراء المستقلب تنقية يشترك مع البروتين خاصة بين العينات (خط مع البروتين أوفيريكسبريسيد للفائدة مقابل EV السيطرة)، قارن قيم الذروة باستخدام اختبار t-غير إقران التائي تليها متعددة تصحيح المقارنة (مثلاً بينجاميني & هوشبيرغ اكتشاف كاذبة معدل تصويب أو تصحيح بونفيروني).
النتائج
في الدراسة الأصلية، كانت overexpressed ثلاثة ندبك التمويل (أ) الجينات في الثقافات تعليق خلية PSB-L تحت سيطرة المروج 35S التأسيسي14 (الشكل 1). وكان تنصهر فيها علامة تقارب جنبا إلى جنب أما النهاية الطرفية كاربوكسي أو الأمينية بروتين الطعم. المجمعات تنقية تقارب تعرضوا ل استخراج الميثانول ومثيل ثالثي بوتيل الإيثر/الماء16. وحددت سحبت تقارب البروتينات والجزيئات الصغيرة باستخدام MS (الجداول S2 و S3).
لتصحيح المغلوطة، استخدمت عينات فارغة لاستبعاد الملوثات جزيء صغير من المواد الكيميائية والمواد الاستهلاكية المختبر. وعلاوة على ذلك، الأيض والبروتينات التي تربط إلى أما علامة تقارب أو الراتنج وحدها كانت استأثرت باستخدام خطوط التحكم EV. لاسترداد إيجابيات حقيقية، واختبار t-غير إقران التائي ومعدل اكتشاف كاذبة بينجاميني & هوشبيرغ تم تطبيق التصحيح لتحديد نواتج الأيض (S4 الجدول) والبروتينات (S5 الجدول) أثرت إلى حد كبير في "وكالة اسوشييتد برس ندبكس" تجارب (N-وج-الميؤوس من شفائهم معلم ندبكس) بالمقارنة مع خطوط التحكم EV (FDR < 0.1). لاحظ أن في الأعمال السابقة، استخدمنا غياب/وجود معايير لتحديد التمثيلات البروتين والجزيئات الصغيرة.
يعطي نتائج تمثيلية ل NDPK1، بينما المستقلب بيانات التركيز على ديبيبتيديس، فئة جديدة من المنظمين جزيء صغير درس في مجموعتنا. وكشف تحليل البروتين 26 شركاء البروتين المفترضة من NDPK1. عن طريق تصفية إضافية للبروتينات مترجمة شارك في نفس حجرة سوبسيلولار ك NDPK1 (cytosol)، القائمة ضاقت إلى 13 بروتين المفترضة التمثيلات. بين البروتينات التي تم تحديدها هي الجلوتاثيون S-ترانسفيراز، وعوامل بدء استطالة اثنين، وتوبولين وهيدراتاسي أكونيتاتي. وكشف تحليل Metabolomic أربعة ديبيبتيديس لوي فال وايل-غلو ولوي-إيل وايل-Phe أن الوتيد على وجه التحديد المشترك مع NDPK1 (الشكل 2). علما بأن حصة ديبيبتيديس الأربعة جميع بقايا مسعور في بهم ن-المحطة، مما يوحي بخصوصية الربط المشترك.
للبحث عن مجمعات البروتين-بروتين والبروتين-المستقلب المعروفة ونحن حددت 13 المستعلم عنها البروتينات وأربعة ديبيبتيديس ضد قاعدة الإبرة25 (الشكل 3). ويمكن إبداء الملاحظات عدة: (ط) أيا من التمثيلات وأبلغ سابقا عن NDPK1. (ثانيا) APX1 أورثولوج أفيد أن تتفاعل مع ألدهيد نازعة أفراد الأسرة ALDH7B4، بينما الترجمة بدء عامل FBR12 مع عامل آخر بدء الترجمة مرمزة بواسطة الجينات AT2G40290. (ثالثا وأفادت) ديبيبتيديس المحددة لا الشركاء البروتين. لم يتم الإبلاغ عنها ديبيبتيديس التيد شارك في وقت سابق كما المرتبطة بالبروتين المصنع استرجاع أي. ومع ذلك، أنها تلعب أدواراً هامة في الكائنات الحية الأخرى: لوي-إيل، مثلاً، له تأثير نيوروتروفين-تفعيل في خط خلية بشرية26. علما أن التجربة لا تسمح بتحديد الطبولوجيا الدقيق للنظام. على سبيل المثال، قد تتفاعل مباشرة مع NDPK1 ديبيبتيدي لكن جيدا قد تكون متصلة بأي من البروتينات المنقاة المشترك.
أخذت معا، النتائج التي توصلنا إليها تظهر أن الإجراءات المعمول بها، توظف AP جنبا إلى جنب مع الطيف الكتلي، يسهل تحديد التمثيلات البروتين-بروتين والبروتين-الصغيرة-جزيء ويساعد على توليد معلومات مستفيضة عن إينتيراكتومي البروتين الهدف.

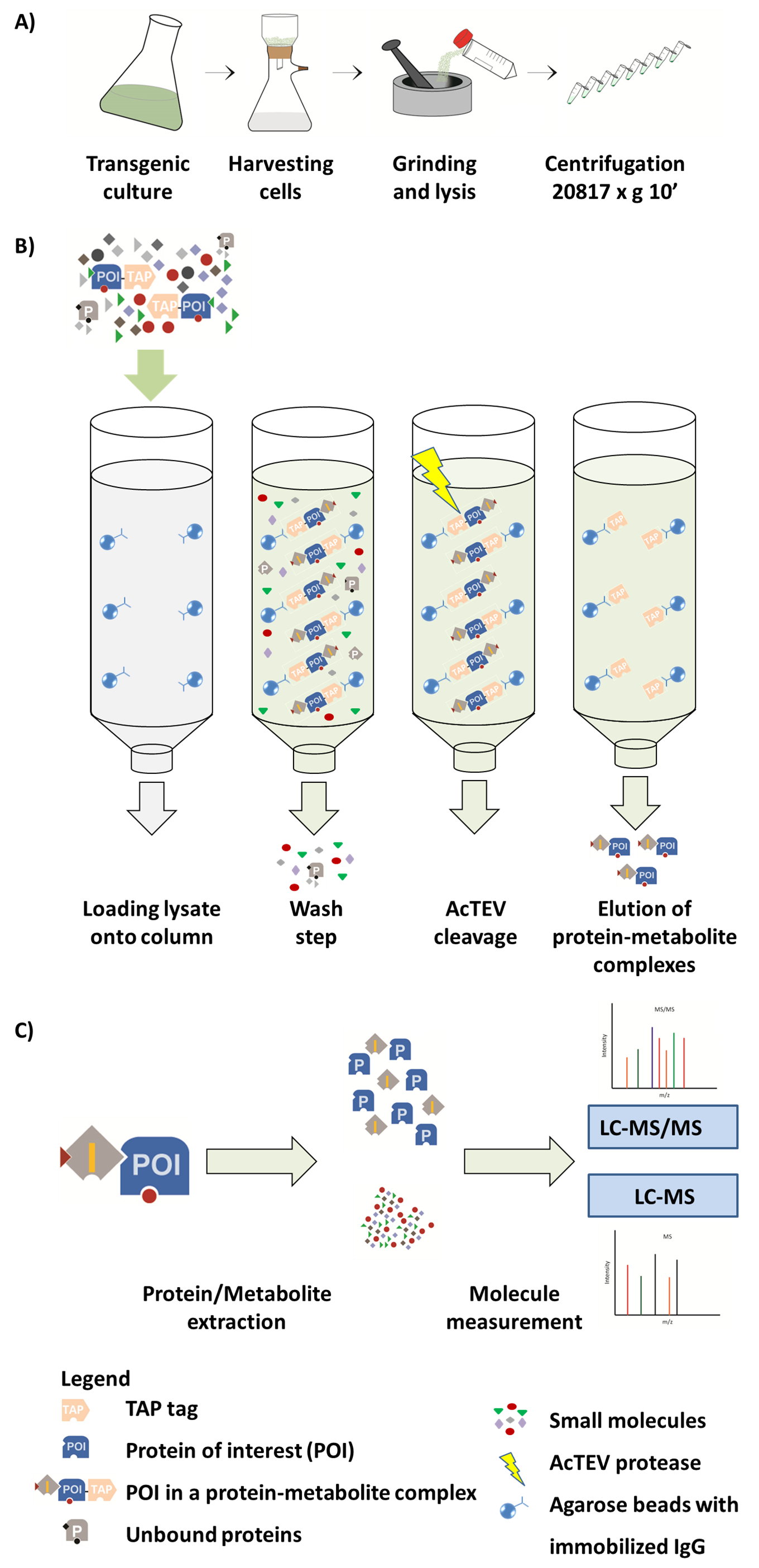
رقم 1. مخطط سير العمل AP-مرض التصلب العصبي المتعدد. (أ) إعداد جزء قابل للذوبان أصلي من زراعة الخلايا النباتية. (ب) الخطوات التالية في الإجراء AP. بعد تحميل العينة على العمود، يربط بروتين الاهتمام (POI) تنصهر فيها علامة حنفية إلى جسم مفتش معطلة على الخرز [اغروس]. غسالة عمود تسهل إزالة البروتينات غير منضم ونواتج الأيض. بعد أداء أكتيف الانقسام، هي التيد مجمعات البروتين-المستقلب البوي. (ج) فصل المجمعات إلى الكسر البروتين والمستقلب متبوعاً بالتحليل الكمي شبه لمرض التصلب العصبي المتعدد. ويرد جزء من هذا الرقم من لوزارووسكي et al. عام 201714. الرجاء انقر هنا لمشاهدة نسخة أكبر من هذا الرقم-
{kind=link}

رقم 2. ديبيبتيديس على وجه التحديد المشترك الوتينج مع NDPK1- تم رسم متوسط كثافة ديبيبتيديس أربعة Val-لوي (A)وايل-غلو (ب)ولوي-إيل (ج)وايل-الفنيل ألانين (د) تقاس في تجربة وكالة اسوشييتد برس. إظهار جميع ديبيبتيديس أربعة إثراء كبيرا في عينات NDPK1 مقارنة بمراقبة EV (العلامات النجمية تمثل فرانكلين روزفلت < 0.1). أشرطة الخطأ تمثل الخطأ المعياري للقياسات 6 (replicates 3 ن-و 3 من ج-شفاء معلم البروتينات). الرجاء انقر هنا لمشاهدة نسخة أكبر من هذا الرقم-
{kind=link}

الشكل 3. شبكة التفاعل من جميع الجزيئات التينج يشترك مع NDPK1، وتساءل ضد قاعدة الإبرة تنظر فقط في السابق التجريبية وقاعدة بيانات الأدلة (الثقة > 0.2). الثقة أعلى يشير إلى فرص أكبر للتفاعل ويحسب استناداً إلى البيانات المودعة. الرجاء انقر هنا لمشاهدة نسخة أكبر من هذا الرقم-
{kind=link}
الجدول S1. ماكسكوانت إخراج الجدول "parameters.txt". جدول يتضمن قيم العتبات للتعريف والتحديد الكمي، فضلا عن معلومات حول قواعد البيانات المستخدمة. اضغط هنا لتحميل هذا الملف.
الجدول S2. المعلومات الواردة من ماكسكوانت إخراج الجدول "proteinGroups.txt". يحتوي الجدول على قائمة بجميع المجموعات المحددة البروتين وكثافات ومعلومات إضافية مثل عدد من الببتيدات فريدة من نوعها ودرجة. اضغط هنا لتحميل هذا الملف.
الجدول S3. ملف الإخراج الذي يحتوي على تحليل لنواتج الأيض القطبية. الجدول يحتوي على قائمة من كافة الميزات الجماعية المحددة تميزت محددة m/z، الرايت وكثافة. اضغط هنا لتحميل هذا الملف.
الجدول S4. ديبيبتيديس وجدت في عينات AP في NDPK1، NDPK2 أو NDPK3 التي استخدمت كطعم. ديبيبتيديس الموجودة في عينات فارغة تم استبعادها من القائمة. تم تشغيل اثنين من خطوط مستقلة (معلم في أما ن-أو ج-تيرمينوس) لكل ندبك في ثلاث نسخ. الطالب t-اختبار ومواصلة تصحيح فالقيمه باستخدام بينجاميني & أسلوب هوشبيرغ واستخدمت لتحديد الشركاء إينتيراكتور المخصب بدرجة كبيرة من ندبكس (FDR < 0.1). ونظرا ΔRT تحسب بالنسبة للمركبات الإشارة و Δppm بالنسبة مونويسوتوبيك نظراً للدمار في ميتلين27. اضغط هنا لتحميل هذا الملف.
الجدول S5. تنقية البروتينات يشترك مع NDPK1. تم تشغيل اثنين من خطوط مستقلة (معلم في أما ن-أو ج-تيرمينوس) لكل ندبك في ثلاث نسخ. الطالب t-اختبار ومواصلة تصحيح فالقيمه باستخدام بينجاميني & أسلوب هوشبيرغ واستخدمت لتحديد الشركاء إينتيراكتور المخصب بدرجة كبيرة من ندبكس (FDR < 0.1). اضغط هنا لتحميل هذا الملف.
Discussion
يسمح البروتوكول قدم موازية الهوية مجمعات PP وبعد الظهر للبروتين الهدف. من الاستنساخ للنتائج النهائية، يمكن أن تكتمل التجربة في أقل من 8-12 أسبوعا. AP كاملة يأخذ حوالي 4-6 ح لمجموعة من العينات 12 إلى 24، مما يجعل لدينا بروتوكول مناسبة لتحليل الإنتاجية منتصف.
البروتوكول، على الرغم من كونها واضحة عموما، يحتوي على عدد من الخطوات الحاسمة. (ط) يكفي كمية البروتين الإدخال والخرز تقارب أمر حاسم للتوصل إلى مجموعة ديناميكية من كشف المستقلب. ولذلك تحلل الخلية فعالة خطوة حاسمة في الإجراء. يمكن أن تكون غلة بروتين الفقراء نتيجة لنضح غير كافية من المواد أو نسبة تحلل-المواد العازلة دون المستوى الأمثل. (ثانيا) ينبغي الحرص أن الكواشف المستخدمة هي صديقة لمرض التصلب العصبي المتعدد. وينبغي تجنب المنظفات القوية أو الجلسرين، أو كميات مفرطة من الملح كما أنها تتعارض مع الكشف عن مرض التصلب العصبي المتعدد. (ثالثا) الخرز [اغروس] ينبغي أن لا الإفراط المجفف أثناء الغسيل الخطوات، وعند استخدام مشعب فراغ من المهم تطبيق معدل تدفق بطيء حتى لا تدمير الخرز أو تؤثر على الاستقرار المعقدة.
وهناك بعض التعديلات المحتملة الهامة في البروتوكول المقدم: (ط) نستخدم المروج CaMV35S التأسيسي لزيادة كمية البروتين الطعم. Overexpression، كانت مفيدة جداً، يمكن أن يكون آثار خطيرة على التوازن خلية28 وأن يؤدي إلى تشكيل تفاعلات غير ذات صلة من الناحية الفسيولوجية. تعبير البروتينات المعلمة استخدام المروجين الأصلية والتي يعتبر فيها ممكن في خلفية فقدان لوظيفة متفوقة لاسترداد التمثيلات البيولوجية الحقيقية. للبروتينات عادة لا المعرب عنه في الثقافات الخلية النباتية، قد يثبت خلفية النباتية اللازمة لتحديد التمثيلات ذات الصلة. (ثانيا) عند العمل مع بروتينات الغشاء, المخزن المؤقت تحلل يحتاج إلى أن تستكمل بالمنظفات الصناعية متوافقة مع مرض التصلب العصبي المتعدد. (ثالثا) الأخذ بخطوة تقارب تنقية ثانية يمكن تحسين إيجابيات كاذبة بالنسبة الحقيقية من إيجابيات والقضاء على الحاجة إلى ضوابط EV29. علامة رواية جنبا إلى جنب مع موقعين حوزتي المستقلة-الانقسام ويقدم بديلاً جذاباً للخطوة حجم الاستبعاد اللوني يضيفها مايدا et al. عام 201411، وشاقة وتستغرق وقتاً طويلاً على حد سواء.
العائق أخطر من وكالة اسوشييتد برس هي المعدل المرتفع من إيجابيات كاذبة. أسباب عديدة. سبق أن ورد overexpression التأسيسي. مصدر آخر للتفاعلات غير ذات صلة من الناحية الفسيولوجية، ما لم يكن العمل مع العضيات معزولة، إعداد ليساتيس كامل الخلية التي تحتوي على خليط من البروتينات ونواتج الأيض من المقصورات سوبسيلولار مختلفة. وينبغي استخدام التعريب سوبسيلولار لتصفية للتمثيلات الحقيقية. ومع ذلك، نتيجة أغلبية المغلوطة ملزمة غير محدد بين البروتينات والراتنجات [اغروس]. مقدمة لتنقية الخطوة الثانية، كما هو موضح أعلاه، يقدم أفضل حل للمشكلة، لكن يأتي على حساب الوقت والإنتاجية. وعلاوة على ذلك، قد يتم فقدان التفاعل أضعف كما يطيل البروتوكول. تحذير آخر من وكالة اسوشييتد برس هو رغم شاملة المعلومات التي يقدمها حول إينتيراكتومي البروتين المستهدف، التفريق بين الأهداف المباشرة وغير المباشرة من البروتين اصطاد مستحيلة. النهج بيموليكولار المستهدفة ضرورية لتأكيد التفاعلات.
AP مقترنة بجميع المستندة إلى MS استخدمت لدراسة البروتين-المجمعات في S. cerevisiae12. هذا العمل، جنبا إلى جنب مع جهودنا الملاحظة السابقة13 ، وبالمثل للدهون، المركبات القطبية وشبه القطبية تظل ملتزمة بمجمعات البروتين المعزولة من ليساتيس الخلوية، وفرت الأساس المفاهيمي للبروتوكول المقدم. لدينا بروتوكول تتميز بثلاث نقاط فريدة من نوعها: (ط) وفي المقابل للخميرة العمل12، فإنه يوضح أن AP مناسبة لاسترجاع ليس فقط البروتين مسعور ولكن أيضا ماء يغاندس. (ثانيا) عن طريق إدخال بروتوكول استخراج ثلاثة في واحد، يمكن استخدام نقطة واحدة لدراسة البروتين والمستقلب التمثيلات من البروتين الطعم. (ثالثا) علينا تكييف البروتوكول الخلايا النباتية.
الجهود المقبلة ستركز على إنشاء علامة جديدة جنبا إلى جنب مع موقعين الانقسام حوزتي مستقلة. ونود أيضا لاستكشاف مدى ملاءمة البروتوكول إلى جزيئات صغيرة منخفضة-وفرة مثل الهرمونات النباتية.
Disclosures
الكتاب ليس لها علاقة بالكشف عن.
Acknowledgements
أننا نود أن يرجى نقر أ. د. لوثار ويلميتزير لمشاركته في المشروع، وإجراء مناقشات مثمرة، وإشراف كبير. ونحن ممتنون للدكتور دانييل Veyel لمساعدة مع قياسات MS البروتين. ونحن نقدر السيدة Änne ميكايليس الذين قدموا لنا المساعدة التقنية لا تقدر بثمن مع قياسات LC-مرض التصلب العصبي المتعدد. وعلاوة على ذلك، نود أن نشكر الدكتورة مونيكا كوسماكز والدكتور اويلينا Sokołowska على مساعدتها ومشاركتها في الأعمال على المخطوطة الأصلية، وإلى Jasińska ويرونيكا لتقديم الدعم التقني.
Materials
| Name | Company | Catalog Number | Comments |
| Murashige and Skoog Basal Salts with minimal organics | Sigma-Aldrich | M6899 | |
| 1-Naphthylacetic acid | Sigma-Aldrich | N1641 | |
| Kinetin solution | Sigma-Aldrich | K3253 | |
| Tris base | Sigma-Aldrich | 10708976001 | |
| NaCl | Sigma-Aldrich | S7653 | |
| MgCl2 | Carl Roth | 2189.1 | |
| EDTA | Sigma-Aldrich | 3609 | |
| NaF | Sigma-Aldrich | S6776 | |
| DTT | Sigma-Aldrich | D0632 | |
| PMSF | Sigma-Aldrich | P7626 | |
| E-64 protease inhibitor | Sigma-Aldrich | E3132 | |
| Protease Inhibitor Cocktail | Sigma-Aldrich | P9599 | |
| Na3VO4 | Sigma-Aldrich | S6508 | |
| AcTEV Protease | Thermo Fischer Scientific | 12575015 | |
| Rotiphorese Gel 30 (37,5:1) | Carl Roth | 3029.2 | |
| TEMED | Carl Roth | 2367.3 | |
| PageRuler Prestained Protein Ladder | Thermo Fischer Scientific | 26616 | |
| SBP Tag Antibody (SB19-C4) | Santa Cruz Biotechnology | sc-101595 | |
| Goat anti-mouse IgG-HRP | Santa Cruz Biotechnology | sc-2005 | |
| Bradford Reagent | Sigma-Aldrich | B6916 | |
| Trypsin/Lys-C Mix, Mass Spec Grade | Promega | V5071 | |
| Urea | Sigma-Aldrich | U5128 | |
| Thiourea | Sigma-Aldrich | T8656 | |
| Ammonium bicarbonate | Sigma-Aldrich | 9830 | |
| Iodoacetamide | Sigma-Aldrich | I1149 | |
| MTBE | Biosolve | 138906 | |
| Methanol | Biosolve | 136806 | |
| Water | Biosolve | 232106 | |
| Acetonitrile | Biosolve | 12006 | |
| Trifluoroacetic acid | Biosolve | 202341 | |
| Formic acid | Biosolve | 69141 | |
| Unimax 2010 Platform Shaker | Heidolph | 5421002000 | |
| Nylon Mesh (Wire diameter 34 µM, thickness 55 µM, open area 14%) | Prosepa | Custom order | |
| Glass Funnel, 47 mm, 300 ml | Restek | KT953751-0000 | |
| Filter Bottle Top 500 mL 0,2 µM Pes St | VWR International GmbH | 514-0340 | |
| Mixer Mill MM 400 | Retsch GmbH | 207450001 | |
| IgG Sepharose 6 Fast Flow | GE Healthcare Life Sciences | 17-0969-02 | |
| Mobicol ""Classic"" with 2 different screw caps without filters | MoBiTec GmbH | M1002 | |
| Filter (small) 35 µM pore size, for Mobicol M 1002, M1003, M1050 & M1053 | MoBiTec GmbH | M513515 | |
| Variable Speed Tube Rotator SB 3 | Carl Roth | Y550.1 | |
| Rotary dishes for rotators SB 3 | Carl Roth | Y555.1 | |
| Resprep 24-Port SPE Manifolds | Restek | 26080 | |
| Finisterre C18/17% SPE Columns 100mg / 1ml | Teknokroma | TR-F034000 | |
| Autosampler Vials | Klaus Trott Chromatographie-Zubehör | 40 11 01 740 | |
| Acclaim PepMap 100 C18 LC Column | Thermo Fischer Scientific | 164534 | |
| EASY-nLC 1000 Liquid Chromatograph | Thermo Fischer Scientific | LC120 | |
| Q Exactive Plus Hybrid Quadrupole-Orbitrap Mass Spectrometer | Thermo Fischer Scientific | IQLAAEGAAPFALGMBDK | |
| Acquity UPLC system | Waters | Custom order | |
| ACQUITY UPLC HSS C18 Column, 100A, 1.8 µM, 2.1 mM X 100 mM, 1/pkg | Waters | 186003533 | |
| High-power ultrasonic cleaning baths for aqueous cleaning solutions | Bandelin | RK 31 | |
| Genedata Expressionist | Genedata | NaN | |
| Xcalibur Software | Thermo Fischer Scientific | NaN | |
| MaxQuant | NaN | NaN |
References
- Li, X., Snyder, M. Metabolites as global regulators: A new view of protein regulation. Bioessays. 33 (7), 485-489 (2011).
- Jacob, F., Monod, J. Genetic regulatory mechanisms in the synthesis of proteins. Journal of Molecular Biology. 3 (3), 318-356 (1961).
- Schlattner, U., et al. Dual Function of Mitochondrial Nm23-H4 Protein in Phosphotransfer and Intermembrane Transfer a cardiolipin-dependent switch. Journal of Biological Chemistry. 288 (1), 111-121 (2013).
- Ramírez, M. B., et al. GTP binding regulates cellular localization of Parkinson's disease-associated LRRK2. Human Molecular Genetics. , ddx161 (2017).
- Jung, H. J., Kwon, H. J. Target deconvolution of bioactive small molecules: the heart of chemical biology and drug discovery. Archives of Pharmacal Research. 38 (9), 1627-1641 (2015).
- Harding, M. W., Galat, A., Uehling, D. E., Schreiber, S. L. A receptor for the immunosuppressant FK506 is a cis-trans peptidyl-prolyl isomerase. Nature. 341 (6244), 758-760 (1989).
- Lomenick, B., et al. Target identification using drug affinity responsive target stability (DARTS). Proceedings of the National Academy of Sciences of the United States of America. 106 (51), 21984-21989 (2009).
- Manabe, Y., Mukai, M., Ito, S., Kato, N., Ueda, M. FLAG tagging by CuAAC and nanogram-scale purification of the target protein for a bioactive metabolite involved in circadian rhythmic leaf movement in Leguminosae. Chemical Communications. 46 (3), 469-471 (2010).
- Pantoliano, M. W., et al. High-density miniaturized thermal shift assays as a general strategy for drug discovery. Journal of Biomolecular Screening. 6 (6), 429-440 (2001).
- Li, X., Snyder, M. Analyzing In vivo Metabolite-Protein Interactions by Large-Scale Systematic Analyses. Current Protocols in Chemical Biology. , 181-196 (2010).
- Maeda, K., Poletto, M., Chiapparino, A., Gavin, A. -. C. A generic protocol for the purification and characterization of water-soluble complexes of affinity-tagged proteins and lipids. Nature Protocols. 9 (9), 2256-2266 (2014).
- Li, X., Gianoulis, T. A., Yip, K. Y., Gerstein, M., Snyder, M. Extensive in vivo metabolite-protein interactions revealed by large-scale systematic analyses. Cell. 143 (4), 639-650 (2010).
- Veyel, D., et al. System-wide detection of protein-small molecule complexes suggests extensive metabolite regulation in plants. Scientific Reports. 7, (2017).
- Luzarowski, M., et al. Affinity purification with metabolomic and proteomic analysis unravels diverse roles of nucleoside diphosphate kinases. Journal of Experimental Botany. , (2017).
- Van Leene, J., et al. Targeted interactomics reveals a complex core cell cycle machinery in Arabidopsis thaliana. Molecular systems biology. 6 (1), 397 (2010).
- Giavalisco, P., et al. Elemental formula annotation of polar and lipophilic metabolites using 13C, 15N and 34S isotope labelling, in combination with high-resolution mass spectrometry. The Plant Journal. 68 (2), 364-376 (2011).
- Van Leene, J., et al. Isolation of transcription factor complexes from Arabidopsis cell suspension cultures by tandem affinity purification. Plant Transcription Factors: Methods and Protocols. , 195-218 (2011).
- Van Leene, J., et al. A tandem affinity purification-based technology platform to study the cell cycle interactome in Arabidopsis thaliana. Molecular & Cellular Proteomics. 6 (7), 1226-1238 (2007).
- Olsen, J. V., Ong, S. -. E., Mann, M. Trypsin cleaves exclusively C-terminal to arginine and lysine residues. Molecular & Cellular Proteomics. 3 (6), 608-614 (2004).
- Cox, J., Mann, M. MaxQuant enables high peptide identification rates, individualized p.p.b.-range mass accuracies and proteome-wide protein quantification. Nature Biotechnology. 26 (12), 1367-1372 (2008).
- Cox, J., et al. Andromeda: A peptide search engine integrated into the MaxQuant environment. Journal of Proteome Research. 10 (4), 1794-1805 (2011).
- Tyanova, S., Temu, T., Cox, J. The MaxQuant computational platform for mass spectrometry-based shotgun proteomics. Nature Protocols. 11 (12), 2301 (2016).
- Hooper, C. M., et al. SUBAcon: a consensus algorithm for unifying the subcellular localization data of the Arabidopsis proteome. Bioinformatics. 30 (23), 3356-3364 (2014).
- Katajamaa, M., Orešič, M. Data processing for mass spectrometry-based metabolomics. Journal of Chromatography A. 1158 (1-2), 318-328 (2007).
- Szklarczyk, D., et al. STITCH 5: augmenting protein-chemical interaction networks with tissue and affinity data. Nucleic Acids Research. 1277, (2015).
- Tanaka, K. -. i., et al. Dipeptidyl compounds ameliorate the serum-deprivation-induced reduction in cell viability via the neurotrophin-activating effect in SH-SY5Y cells. Neurological Research. 34 (6), 619-622 (2012).
- Smith, C. A., et al. METLIN: A metabolite mass spectral database. Therapeutic Drug Monitoring. 27, 747-751 (2005).
- Bhattacharyya, S., et al. Transient protein-protein interactions perturb E. coli metabolome and cause gene dosage toxicity. Elife. 5, (2016).
- Rigaut, G., et al. A generic protein purification method for protein complex characterization and proteome exploration. Nature Biotechnology. 17 (10), 1030-1032 (1999).
Reprints and Permissions
Request permission to reuse the text or figures of this JoVE article
Request PermissionExplore More Articles
This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. All rights reserved