
Method Article
2 в 1: один шаг очищение сродства для параллельного анализа протеин протеина и белка метаболит комплексов
* Эти авторы внесли равный вклад
В этой статье
Резюме
Взаимодействий протеин протеина и белка метаболит имеют решающее значение для всех клеточных функций. Здесь мы описываем протокол, который позволяет параллельный анализ этих взаимодействий с белка выбора. Наш протокол был оптимизирован для культур клеток растений и сочетает в себе очищение сродства с массы на основе спектрометрии белков и метаболитов обнаружения.
Аннотация
Клеточные процессы регулируются взаимодействия биологических молекул, таких как белки, метаболитов и нуклеиновых кислот. Хотя расследование белок белковых взаимодействий (PPI) не новинка, экспериментальные подходы, стремясь охарактеризовать эндогенного белка метаболит взаимодействий (PMI) составляют довольно недавно. Здесь мы представляем протокол, который позволяет одновременное характеристика ИЦП и PMI белка выбора, именуемые как приманку. Наш протокол был оптимизирован для клеток Arabidopsis культур и сочетает в себе очищение сродства (AP) с масс-спектрометрия (МС)-на основе обнаружения белков и метаболитов. Короче говоря трансгенных линий Arabidopsis, выражая приманки белка, сливается в тег сходства, сначала лизированы получить собственный сотовый экстракт. Анти тег антитела используются для тянуть вниз белков и метаболитов партнеров приманки белка. Близость очищенная комплексы извлекаются с помощью одношагового метил трет-бутилового эфира (МТБЭ) / метанол/воды методом. Хотя метаболитов разделить на полярных или этапа гидрофобная, белки можно найти в гранулы. Метаболиты и белки затем анализируются ультра производительность жидкого хроматография масс-спектрометрия (МС-UPLC или UPLC-МС/МС). Пустой вектор (EV) управления линии используются для исключения ложных срабатываний. Основное преимущество нашего протокола является, что он позволяет идентификации белков и метаболитов партнеров целевого белка параллельно в рядом физиологических условиях (сотовых lysate). Представленный метод простой, быстрый и может быть легко адаптирована для биологических систем, отличных от культуры клеток растений.
Введение
Метод здесь описано направлена на выявление партнеров метаболит и белка белка выбора в ближайшем -в vivo сотовой lysate условиях. Было предположение, что многие другие метаболиты чем характеризуется сегодня имеют важные нормативные функции1. Метаболиты могут действовать как биологические коммутаторы, изменяя активность, функциональность, и/или локализации их рецептор белков2,и3,4. В последнее десятилетие несколько прорыв методы, позволяя идентификации PMI в естественных условиях или в условиях Ближнего -в vivo , были развитые5. Доступные подходы могут быть разделены на две группы. Первая группа включает в себя методы, которые начинаются с известный метаболит приманку для улавливания романный протеин партнеров. Методы включают хромотографии сродства6, наркотиков близость реагировать целевой стабильность пробирного7, химио протеомики8и тепловых профилирование9протеома. Вторая группа состоит из одного метода, который начинается с известного белка для выявления мелкомолекулярных лигандов10,11.
AP, в сочетании с на основе MS lipidomics использовался для анализа белково липидные комплексов в Saccharomyces cerevisiae12. Как отправной точки авторы использовали штаммы дрожжей, выражая 21 ферментов, участвующих в биосинтезе эргостерол и 103 киназы сливается с тандем сродства очищения (TAP) тег. 70% ферментов и 20% киназ были найдены для привязки различных гидрофобные лигандов, проливая свет в сложный белково липидные сеть взаимодействия.
Ранее мы смогли продемонстрировать, что, аналогично в липиды, полярных и полу полярных соединений также оставаться связанными белковых комплексов, изолированных от сотовой лизатов13. Основываясь на этих выводах, мы решили оптимизировать AP метод ранее опубликованы11 10,растительных клеток и гидрофильные соединения14. Для этой цели мы использовали TAP векторов, описываемого Van Leene et al. 2010, успешно используется в завод PPI исследования15. Чтобы сократить время, необходимое для получения трансгенных линий, мы решили на культурах клеток арабидопсиса. Мы заняты одношаговый метил трет-бутиловый эфир, (МТБЭ) / метанол/вода извлечения метода, позволяя характеристика белков (гранулы), липиды (органические фазы) и гидрофильные метаболитов (водная фаза)16 в одном очищение сродства эксперимент. EV линии контроля были введены для исключения ложных срабатываний, например привязка к тегу только белки. Как доказательство концепции, мы с тегами три (из пяти) нуклеозидов дифосфат киназы присутствует в геном арабидопсиса (NDPK1-NDPK3). Среди других выводов, мы смогли продемонстрировать, что NDPK1 взаимодействует с глутатион S-трансферазы и глутатиона. Поэтому мы смогли доказать, что NDPK1 подвергается glutathionylation14.
Короче говоря, представленный протокол является важным инструментом для характеризующие протеин протеина и белка мелкомолекулярных взаимодействия сетей и представляет собой крупный шаг вперед над существующими методами.
протокол
Подготовка трансгенные Arabidopsis клетки культуры линий, включая клонирование, преобразование, отбора и роста условия могут быть найдены в17. Обратите внимание, что линии контроля EV рекомендуется исправить для ложных срабатываний. До эксперимента подтвердите гиперэкспрессия белка приманки, Западный анализ помаркой, например с использованием антител IgG против G-белок частью тега сродство тандем. Важно отделить роста СМИ от материала культуры клеток растений.
1. Подготовка материала клеток растений до эксперимента
- Растут ПСБ-L A. thaliana линия культуры клетки, экспрессирующих протеин интереса18.
- Подготовка MSMO среда, которая содержит 4.43 г/Л, которые MSMO смешанной с сахарозой 30 г/Л. Отрегулируйте pH буфера до 5,7 с 1 M Кох и автоклав решение. До эксперимента дополнение в среду с α-naphthaleneacetic кислота 0,5 мг/Л, кинетин 0,05 мг/Л и канамицин 50 мкг/мл.
- Культивируйте культуры клеток растений превращается в 50 мл MSMO среды в 100 мл флакон на орбитальной платформы шейкер с нежным агитации (130/мин). Рост клеток в комнате культуры при 20 ° C и интенсивности света равен 80 мкмоль m s-2 -1.
- Субкультура клетки в свежие средства каждые 7 дней, разбавляя их 1:10.
- Соберите клетки в фазе логарифмического роста, используя воронку стекла в сочетании с вакуумным насосом, используя нейлоновая сетка в качестве фильтра. Оберните проникнуть в алюминиевой фольги и замораживание в жидком азоте.
Внимание: Помните, что жидкий азот очень холодно. Некорректная обработка может вызвать ожоги. Носите соответствующие средства индивидуальной защиты, включая термически изолированные перчатки, защитные очки и халате.
2. Выберите протокол
Примечание: Следующий шаг заимствован из Маэда et al. 201411 и Van Leene et al. 201117.
- Однородный завод собранный и замороженных клеток культуры материала с помощью вибрационная мельница (2 мин при 20 Гц) или ступку и пестик для получения мелкодисперсного порошка. Аликвота 3 g земли материала (соответствует примерно 90 мг общего белка) на сэмпл. Избегайте оттаивания образца на этом шаге с помощью оборудования предварительно охлажденный жидкий азот.
Примечание: Магазин земли растительный материал в 50 мл трубки на – 80 ° C до начала процедуры AP. - Нарезанных образца в жидкий азот охладить раствора с 3 мл ледяной литического буфера (0,025 М трис-HCl рН 7,5; 0,5 М NaCl 1.5 мм MgCl20,5 мм DTT; 1 мм NaF; 1 мм Na3VO4; 100 x разреженных коммерческих протеазы ингибитор коктейль; 1 мм PMSF) до тех пор, пока материал таяет. Как только образец тает, сразу переходите к следующему шагу.
Примечание: Подготовьте свежие буфера lysis. Ввести пустой образцы на этот шаг. Моющие средства не рекомендуется, поскольку они могут вызвать проблемы в MS обнаружения. - Чтобы удалить сотовой мусора, разделите материал на 2 мл microcentrifuge трубы и центрифуги на 20817 g x 10 мин при 4 ° C. Соберите 3 мл ясно lysate в 15 мл конические пластиковых пробирок.
- Во время шага центрифугирования сбалансировать IgG-Sepharose бусины. Аликвота 100 мкл бусы на сэмпл и мыть их с 1 мл буфера lysis. Вихрь Ресуспензируйте бусы и флэш спина. Отменить литического буфера и повторите шаг дважды. Ресуспензируйте бусины в 400 мкл буфера lysis.
- Добавить бисер на собранных завод lysate и инкубировать смесь на вращающееся колесо за 1 час при 4 ° C.
- Передача смеси в шприц объединены через Luer-lock Кап со столбцом спин с фильтром поры размер 35 мкм. применить давление пройти lysate через. Бусины с прилагаемой комплексы останется на фильтр, в то время как lysate будет проходить через.
Примечание: При необходимости, используйте систему вакуумного коллектора. Убедитесь в том применить нежное давление, чтобы не нанести вред бисер. - Мыть бисер сначала с 10 мл отмывающего буфера (0,025 М трис-HCl рН 7,5; 0,5 М NaCl), а затем с 1 мл раствора Элюирующий буфер (10 мм трис-HCl рН 7,5; 150 мм NaCl; 0,5 мм ЭДТА 1000 x разреженных E64 и 1 мм PMSF). Выполните с помощью шприца вложенного к столбцу или вакуумного коллектора системы мойки.
Примечание: Когда с помощью вакуумного коллектора системы обязательно применять мягкое давление, чтобы не нанести вред бисер. - Инкубировать бусы с 400 мкл Элюирующий буфер, содержащий 50 U улучшенной версией табака etch протеазы вируса (AcTEV). Использование таблицы шейкер на 1000 об/мин за 30 мин при температуре 16 ° C.
Примечание: Помните, чтобы использовать плагин, чтобы закрыть столбец в нижней части, добавив Элюирующий буфер. - Добавьте дополнительную часть (50 U) фермента в столбце и инкубировать смесь для следующей 30 мин в тех же, описанных выше, условия.
- Соберите элюата в 2-мл пробирку microcentrifuge либо путем центрифугирования (1 мин, 20817 x g) вакуумный коллектор. Чтобы удалить оставшиеся комплексов, ввести дополнительные элюции шаг, используя 200 мкл буфера.
Примечание: Хранить образца при – 20 ° C или – 80 ° C, или немедленно приступить к этапу извлечения белков и метаболитов. Оттепель замороженных образцов на льду.
3. Западный анализ помаркой
- Чтобы подтвердить наличие приманка белка в собранных элюата использовать 10 мкл – метаболит белковые элюата для выполнения SDS-PAGE и Западный анализ помаркой. Чтобы определить протеина интереса, используйте мышь первичных антител против стрептавидина связывающий белок (1: 200), частью тега крана, оставшиеся после расщепления протеазы ТэВ, как описано в Van Leene et al. 201117. Далее использование вторичного коза против мышиных антител в сочетании с ПХ.
4. метаболита и экстракции белка
Примечание: Этот протокол заимствован из Giavalisco et al. 201116.
Примечание: Этот шаг начиная используйте UPLC – MS-класс решения.
- Добавьте 1 mL метил- трет-бутилового эфира (МТБЭ) / метанол/растворитель (3:1:1) для собранных элюата воды и перемешать образец путем инверсии. Убедитесь, что растворитель охлаждается до – 20 ° C до этапа добычи.
Предупреждение: МТБЭ и метанол являются вредными веществами. Выполните извлечение шаг под зонт и носить соответствующие личной защиты оборудования, например, перчатки. - Решение 1:3 добавить 0,4 мл метанола: воды каждый образец и смешать содержимое образца, инверсии.
Примечание: Добавок смеси метанола: водным раствором приводит разделение фаз. Верхний этап содержит липиды, Нижняя этап содержит полярного и полу полярных метаболитов и белки можно найти в гранулы. - Отдельные этапы, Центрифугуйте образцы на 20817 x g на 2 мин при комнатной температуре, а затем собирать Верхний этап для измерений липидов (не сделали в этом протоколе) с помощью ручной обработки жидкость пипеткой с 1 мл емкость.
- Добавьте 0,2 мл метанола и перемешать путем инверсии.
- Центрифугуйте образцы на 20817 x g на 2 мин на RT, а затем собирать полярных фазы для измерений метаболит (полярного и полу полярных соединений). Чтобы не мешать белка Пелле, оставьте около 50 мкл жидкой фазы в нижней части трубки.
- Сухой собранные образцы для измерения метаболит ночь в центробежных испарителя. Избегайте чрезмерной сушки гранул белка путем удаления образцов из испарителя после 30 – 60 мин.
Примечание: Хранить образцы при – 20 ° C или – 80 ° C, или немедленно приступить к подготовке белков для анализа LC-MS/MS.
5. подготовка проб для анализа Proteomic
Примечание: Этот шаг приспособлен от Олсен и др. 200419 и техническое руководство трипсина/Lys-C смеси (см. Таблицу материалы).
- Выполнение образца ферментативного пищеварения.
Предупреждение: Растворителей во время ферментативного пищеварения и обессоливания образца вредны. Работа под зонт и носить соответствующие личной защиты оборудования, например, перчатки.- Растворяют гранулы белка в 30 мкл буфера свежеприготовленные денатурация (40 мм бикарбонат аммония, содержащие 2 M Тиомочевина/6 M мочевины, рН 8). Для достижения лучшей растворимости белка, выполните шаг 15-мин sonication. Повторите шаг до полного растворения гранул.
- Центрифугуйте образцы на 20817 g x 10 мин при температуре 4 ° C, а затем передать новые пробки microcentrifuge супернатант.
- Определите концентрацию протеина используя assay протеина Брадфорд.
- Для дальнейшего анализа, аликвота тома эквивалентна 100 мкг белка и заполнить до 46 мкл пример с денатурации буфера.
- Добавить в образец 2 мкл свежеприготовленные уменьшения буфера (50 мм, которые DTT растворяют в H2O) и Инкубируйте 30 мин при комнатной температуре.
- Лечить образца с 2 мкл буфера свежеприготовленные алкилирование (150 мм iodoacetamide растворяется в буфере бикарбонат аммония 40 мм) и инкубировать смесь в темноте для 20 мин при комнатной температуре.
- Разбавить образца с 30 мкл буфера бикарбонат аммония 40 мм и 20 мкл LysC/трипсина смеси.
- После 4 ч инкубации при 37 ° C разбавляют образца с 300 мкл буфера бикарбонат аммония 40 мм.
- Продолжить с ночи инкубации при температуре 37 ° C.
- Подкислять образца с приблизительно 20 мкл 10% trifluoroacetic кислоты (ТФК) для получения рН < 2. Проверьте образец pH с помощью РН полосы.
Примечание: Хранить образца при – 20 ° C или перейти к следующему шагу.
- Опреснения переваренной белков.
Примечание: Желательно, используйте систему вакуумного коллектора. Избегайте чрезмерного высыхания столбца.- Промойте столбце C18 SPE (см. Таблицу материалы) с 1 мл раствора 100% метанола и затем с 1 мл раствора 80% Ацетонитрил (ACN) содержащий 0.1% TFA разводят в воде. Шаги использования, здесь и далее белок обессоливания, вакуумного коллектора системы ускорить этот процесс. Избегайте чрезмерного высыхания столбца.
- Сбалансировать столбце путем промывания его дважды с 1 мл 0,1%, которую TFA разводят в воде.
- Загрузите образец на столбец. Промойте трубку с дополнительные 200 мкл 0,1% TFA и передачи решения на столбце. Запуск решения через колонку.
- Промойте колонку дважды с 1 мл 0,1% TFA.
- Элюировать обессоленной пептиды из столбца с 800 мкл 60% ACN, 0,1% раствор TFA. Сухие собираемой фракции в центробежных испарителя, избегая чрезмерного высыхания белковые фракции путем удаления образцов из испарителя после 30 – 60 мин.
Примечание: Хранить образцы при – 20 ° C или – 80 ° C или сразу перейти к следующему шагу. В последующих шагах Храните пробы на льду.
6. Измерение подготовлены образцы протеина, с помощью UPLC – МС/МС.
Примечание: Перед протеомных и Метаболомные измерений, фильтр (размер пор 0.2 мкм) и Дега все буферы с помощью вакуумного насоса для 1 h.
- Ресуспензируйте сушеные пептид Пелле, хранятся в 2-мл пробирку microcentrifuge в 50 мкл буфера C (3% v/v ACN, муравьиная кислота 0,1% v/v) с использованием ручной обработки жидкость пипеткой с 200 мкл объём. Sonicate образцы для 15 мин в ультразвуковой ванне с ультразвуковой частотой 35 кГц.
Предупреждение: ACN и муравьиной кислоты являются вредными веществами. Работа под зонт и носить соответствующие личной защиты оборудования, например, перчатки. - Центрифугуйте образцы на 20817 g x 10 мин при температуре 4 ° C, а затем передать стеклянный флакон 20 мкл супернатант.
- Отдельные переваривается пептиды, с помощью столбца обратная фаза C18 подключены к жидкостной хроматографии и приобрести массового спектры с использованием масс-спектрометр.
- Отдельно в колонке 3 мкл пример с помощью скорость потока 300-нл/мин. Для мобильных фазы, используйте буфер C и D (63% v/v ACN, муравьиная кислота 0,1% v/v), образуя градиент наращивает от 3% ACN до 15% ACN свыше 20 мин и затем до 30% АКС в течение следующих 10 мин.
Примечание: Сохраните остальной части образца при – 20 ° C или – 80 ° C до нескольких месяцев. Перед измерением proteomic заморозить образца на льду. - Промыть загрязнителей за 10 мин, с использованием 60% АКС и сбалансировать столбец с 5 мкл буфера C перед измерением следующего образца.
- Получения массового спектры, зависящих от данных методом МС/МС с резолюции набор на 70000, AGC целевой 3e6 ионов, время максимальной впрыска 100 мс, и m/z от 300 до 1600. Приобрести максимум 15 МС/МС сканирование на резолюции 17500, СМЖЛ целевой 1e5, время максимальной впрыска 100 мс, underfill соотношение 20%, с окном изоляции 1.6 m/z и m/z от 200 до 2000 года. Включите Апекс триггера (6 – 20 s), Комплект динамического исключения 15 s, и исключить обвинения 1 и > 5.
- Отдельно в колонке 3 мкл пример с помощью скорость потока 300-нл/мин. Для мобильных фазы, используйте буфер C и D (63% v/v ACN, муравьиная кислота 0,1% v/v), образуя градиент наращивает от 3% ACN до 15% ACN свыше 20 мин и затем до 30% АКС в течение следующих 10 мин.
7. обработка протеомических данных
- Скачать новейшие базы данных протеома Arabidopsis thaliana от http://www.uniprot.org/ и включают загрязнителем базы данных. Анализировать исходные данные, полученные из LC-MS выполняется с использованием MaxQuant с интегрированной Андромеды пептид поисковой системы, используя настройки по умолчанию с поддержкой LFQ нормализации20,21,22. Найти подробную информацию о параметрах, используемых в Таблице S1.
- Откройте выходной файл «белка groups.txt». Для дальнейшего анализа фильтр для групп белков с по крайней мере два уникальных пептиды. Удалите группы белков, определяется MaxQuant как потенциальных загрязнителей и фильтр для A. thaliana белков (ARATH в Fasta заголовки столбцов), присутствующих в базе данных.
- Чтобы проверить значение белка обогащения между образцами, использовать LFQ нормированный интенсивности и выполнять студент непарных, двустороннее t тест следуют множественные сравнения коррекции (например Benjamini и Хёхберг ложных обнаружения ставка (ДРД) Коррекция или Бонферрони коррекции).
- Вычислить p значение путем сравнения интенсивности LFQ, полученные для управления EV и NDPK1. Отфильтровать все неопределенные значения. P значения сортируются в порядке возрастания и использовать сценарий R или онлайн калькулятор (например, https://www.sdmproject.com/utilities/?show=FDR) для вычисления ФДР коррекции. Фильтр по значениям ФДР ниже 0,1.
Примечание: Рассмотрим форму анализа данных подходит для исследований. Количественные исследования (анализ белков обогащения между выборками) использовать значение «LFQ света», тогда как для качественных исследований (наличие или отсутствие белка) выберите значение «Интенсивность». - Фильтр для групп белков, которые более обильны в NDPK1 по сравнению с EV управления. Определить локализацию потенциальных партнеров белка, с использованием базы данных SUBA23 и исправить белка локализован совместно с NDPK1.
- Вычислить p значение путем сравнения интенсивности LFQ, полученные для управления EV и NDPK1. Отфильтровать все неопределенные значения. P значения сортируются в порядке возрастания и использовать сценарий R или онлайн калькулятор (например, https://www.sdmproject.com/utilities/?show=FDR) для вычисления ФДР коррекции. Фильтр по значениям ФДР ниже 0,1.
8. Измерение образцов, содержащих полярных фазы с использованием UPLC – МС.
- Ресуспензируйте сушеные полярных фазы от шага 4.5 в 200 мкл воды и sonicate образец для 5 мин.
- Центрифугуйте образцы на 20817 g x 10 мин при температуре 4 ° C, а затем передать стеклянный флакон супернатант.
Примечание: Сохраните остальной части образца при – 20 ° C или – 80 ° C для до нескольких месяцев. Перед измерением Метаболомные заморозить образца на льду. - Выполните шаг разделения, используя UPLC сочетании C18 обратная фаза столбец и приобрести массового спектры с МС.
- Загрузить на колонке 2 мкл образца на инъекцию для каждого режима ионизации (положительные и отрицательные) и отдельные фракции, используя 400 мкл/мин скорость потока. Для создания требуемых градиента для измерения метаболит, приготовляют раствор мобильных этапа следующим образом: буфера (0,1% муравьиной кислоты в H2O) и буфера B (0,1% муравьиной кислоты в АКС).
- Отдельных метаболитов в 400 мкл/мин и следующий градиент: 1 мин 99% буфера A, 11-мин линейный градиент от 99% буфера A до 60% буфера A, 13-мин линейный градиент от 60% буфера A 30% буфера A, 15-мин линейный градиент от 30% буфера A до 1% буфера A удерживайте 1% концентрации до 16 мин, начиная с 17 мин, использования линейного градиента от 1% буфера A до 99% буфера, а. заново сбалансировать столбце 3 мин с 99% концентрации буфера A перед измерением следующего образца.
- Приобрести массового спектры, охватывающих область диапазона массы от 100 до 1500 m/z с резолюцией равным 25 000 и загрузки время ограничивается 100 г-жа целевой набор СМЖЛ 1e6, капиллярные напряжения в 3кв с оболочкой газа потока и значение вспомогательных газ 60 и 20 , соответственно. Установите капиллярного температуру до 250 ° C и скиммер напряжения 25V.
9. обработка данных Метаболомика
- Процесс собранные хроматограм, полученных от обоих режимов ионизации. Использовать программное обеспечение для извлечения массы заряда коэффициент (m/z), время удерживания (RT) и интенсивности пиков связанные, например, коммерческое программное обеспечение (см. Таблицу материалов) или альтернативных24.
- Запустите программное обеспечение для обработки, дважды щелкнув файл .exe
- Создание нового рабочего процесса, поиск для деятельности «Загрузки из файла» и переместить эту деятельность путем «перетаскивание» в пространство пустой рабочий процесс. Нажмите действия с правой кнопкой мыши и откройте параметры деятельности.
- В вкладку, содержащую параметры «Генерал» задайте имя эксперимента в поле «Имя» и затем нажмите «выбрать файлы и папки» и Марк сырье хроматограммы.
- В вкладки, содержащие параметры «Дополнительно» равным 0 интенсивности «Профиль данных отсечки». Нажмите кнопку «Применить» и «ОК».
- Поиск и добавить действие «Данные поручения». Нажмите действия с правой кнопкой мыши и откройте параметры деятельности.
- В закладке, содержащие настройки «Общие» Марк «Средневзвешенные данные» и «МС/МС данные». Удалите все данные MS/MS, выбрав «Все» в панели выбора.
- Поиск и добавить действие «Хроматограммы химический шум вычитание». Нажмите действия с правой кнопкой мыши и откройте параметры деятельности.
- В вкладку, содержащую параметры «Генерал» Марк, «Хроматограмма разглаживание» и задайте количество сканов на «3» и «Оценщика» до «Moving average». Установите «RT окна» 51 сканов, «Квантиль» до 50%, вычитание «Метод» и 750 интенсивности «Порог».
- В вкладки, содержащие параметры «Дополнительно» Марк «RT структуры удаление» и «Минимальная длина RT» присвоено 5 сканирования.
- В вкладку, содержащую параметры «Продвинутые» знак «m/z удаления структуры» и установить «Минимум m/z длина» 3 очка.
- Поиск и добавить действие «Хроматограмма RT выравнивание». Нажмите действия с правой кнопкой мыши и откройте параметры деятельности.
- В закладке, содержащие настройки «Общие» равным 0,5 мин «Выравнивание схема», «Попарно выравнивание база дерево» и «Интервал поиска RT».
- В вкладку «Дополнительно» настроек используйте параметры по умолчанию.
- Поиск и добавить действие «Пик обнаружения» в группе «Хроматограмма» деятельности. Нажмите действия с правой кнопкой мыши и откройте параметры деятельности.
- В вкладку, содержащую параметры «Общие» «Суммирование окна» присвоено 0,09 мин, «Минимальный размер пик» до 0,03 мин, «Максимальная слияния расстояние» до 5 баллов стратегия и «слияния» до «Центров». В «Пик расщепление RT» в поле Задайте «Разрыв/пиковое отношение» до 50%.
- В вкладку, содержащую параметры «Дополнительно» «Сглаживание окна» присвоено 5 очков, «Уточнение порог» до 80% и «Последовательности порог» до 1. Набор «Центр вычисления» как «Интенсивности взвешенный» с «Интенсивность порог» на 70%.
- Поиск и добавить действие «Изотоп кластеризации» в группе «Хроматограмма» деятельности. Нажмите действия с правой кнопкой мыши и откройте параметры деятельности.
- В вкладку «Общие» настройки установите «RT терпимости» 0,015 мин и «m/z терпимости» 5 ppm.
- В закладке, содержащие «Конверт установку» параметры установите «Метод», как «No форму ограничения» и «Ионизация» как «протонирование (для положительных режима) и «Deprotonation» (для отрицательных режима). Набор «минимальный и максимальный заряд» 1 и 4, соответственно.
- В вкладку «Дополнительно» настроек используйте параметры по умолчанию.
- Поиск и добавить действие «Синглтон фильтр».
- Чтобы экспортировать результаты обработки данных, поиска и добавить действие «Аналитик» в группе «Экспорт» деятельности.
- В вкладку «Общие» настройки установите «Тип» как «Группы» и «Наблюдаемый» как «Подвел интенсивности». Выберите «пользовательский» и укажите каталог файла экспорта.
- В вкладку «Дополнительно» настроек используйте параметры по умолчанию.
- Аннотируйте массового черт, с использованием внутренних ссылок соединения базы данных.
- Анализ одного или нескольких MS класс ссылки соединение с помощью использования UPLC-г-жа тот же метод LC-MS для анализа ссылок соединений и метаболитов, совместно очищенный с протеина интереса.
Примечание: Для этого исследования, набор почти 300 дипептиды был проанализирован и используется в качестве ссылки соединение библиотеки. - Анализ использования (см. Таблицу материалы) программного обеспечения, чтобы открыть файл Сырье Хроматограмма и поиск для конкретных m/z и RT, связанные с измеряется ссылка соединение (см. Руководство пользователя).
Примечание: Вторичные метаболиты отличаются в типах ионизации. Проверьте наличие общих аддукты, ища для ионной массы, равная M-1.007276, M + 1.007276, M + 18.033823 и M + 22.989218 [M-H], [M + H], [M + NH4] и [M + Na], соответственно. - Используйте электронную таблицу, чтобы открыть экспортируется файл «Аналитик», полученные после обработки хроматограммы и поиска конкретных Ион массы. Сравните RT функцию массового, измеряется в эксперименте и RT соединение ссылку. Разрешить отклонение 0,005 Да для m/z и 0.1 мин для RT.
- Анализ одного или нескольких MS класс ссылки соединение с помощью использования UPLC-г-жа тот же метод LC-MS для анализа ссылок соединений и метаболитов, совместно очищенный с протеина интереса.
- Чтобы проверить значение метаболит обогащения совместно очищенный с белка между выборками (линия с гиперэкспрессия белка интерес против EV управления), Сравните пиковые значения, с помощью двух паре Стьюдента t теста следуют несколько Сравнение коррекция (например Benjamini и Хёхберг ложных обнаружения курс коррекции или Бонферрони коррекции).
Результаты
В первоначальном исследовании были оверэкспрессировали три NDPK A. thaliana генов в ОВО-L подвеска культур клеток под контролем учредительного 35S промоутер14 (рис. 1). Тандем близости тег был снаряжен либо карбоксильную или амино терминал конец приманки белка. Близость очищенная комплексы были подвергнуты MTBE/метанол/вода извлечения16. Вытащил сродство белков и малые молекулы были определены с использованием MS (таблицы S2 и S3).
Для устранения ложных срабатываний, пустой образцы были использованы для исключения мелкомолекулярных загрязнителей из химикаты и лабораторные Расходные материалы. Кроме того метаболитов и белками, которые связывают либо тег сродство или смолы одиночку были учтены с помощью линий управления EV. Для получения истинных положительных результатов, два номера в паре Стьюдента t тест и Benjamini & Хёхберг ложных обнаружения курс коррекции был применен для выявления метаболитов (Таблица S4) и белков (Таблица S5) значительно обогатили в ап NDPKs эксперименты (N - и C-неизлечимо меткой NDPKs) по сравнению с линии управления EV (ФДР < 0.1). Обратите внимание, что в предыдущей работе, мы использовали присутствие/отсутствие критериев для определения белков и мелкомолекулярных посредники.
Представитель результаты даны для NDPK1, а метаболит данных занимаются дипептиды, Роман класса мелкомолекулярных регуляторов, учился в нашей группе. Протеомного анализа выявлено 26 партнеров предполагаемого белок NDPK1. Дальнейшей фильтрации для белков, совместно локализовано в одном отсеке субцеллюлярные как NDPK1 (цитозоль), список сократился до 13 предполагаемых белка посредники. Среди выявленных белки были глутатион S-трансферазы, два удлинение факторы инициации, тубулин и aconitate hydratase. Метаболомные анализ выявил четыре дипептиды Валь-лей, Иль-Glu, лей-Иль и Иль-Пхе, конкретно этого eluted совместно с NDPK1 (рис. 2). Обратите внимание, что все четыре дипептиды разделяют гидрофобные остатков в их N-отель terminus, предложив общие привязки специфичности.
Искать известных комплексов протеин протеина и белка метаболит мы запрашиваемые 13 определенных белков и четыре дипептиды против стежка базы данных25 (рис. 3). Несколько замечаний: (i) ни один из посредники сообщалось ранее для NDPK1. (ii) APX1 ortholog, как сообщается, взаимодействовать с членом семьи ALDH7B4, дегидрогеназа альдегид во время перевода фактор инициации FBR12 с другой фактор инициации перевода, кодируемых геном AT2G40290. (iii выявленных дипептиды не сообщили белка партнеров. Совместное eluted дипептиды не сообщалось ранее, как связанные с любой проверено растительного белка. Однако, они играют важную роль в других организмов: леи-Иль, например, имеет эффект Нейротрофин активации в клетки человека линии26. Обратите внимание, что эксперимент не позволяют определить точную топологию системы. К примеру дипептид могут взаимодействовать непосредственно с NDPK1, но также могут быть связаны с какой-либо совместной очищенную белков.
Взятые вместе, наши результаты показывают, что установленной процедуре, используя AP вместе с масс-спектрометрии, облегчает выявление протеин протеина и белка мелкомолекулярных посредники и помогает создавать обширную информацию о interactome целевого белка.

Рисунок 1. Схема рабочего процесса AP-MS. (A) подготовка родной растворимая фракция от культуры клеток растений. (B) последующие шаги в процедуре AP. После загрузки образца на столбце, протеина интереса (POI) сливается в тег TAP привязывается к IgG антитела, иммобилизованных на бусины агарозы. Стиральная столбце облегчает удаление несвязанных белков и метаболитов. После выполнения AcTEV расщепления, являются этого eluted комплексы белка метаболит POI. (C) разделение комплексов на долю белков и метаболитов, следуют полуколичественного анализа MS. Luzarowski et al. 201714воспроизводится часть этой фигуры. Пожалуйста, нажмите здесь, чтобы посмотреть большую версию этой фигуры.
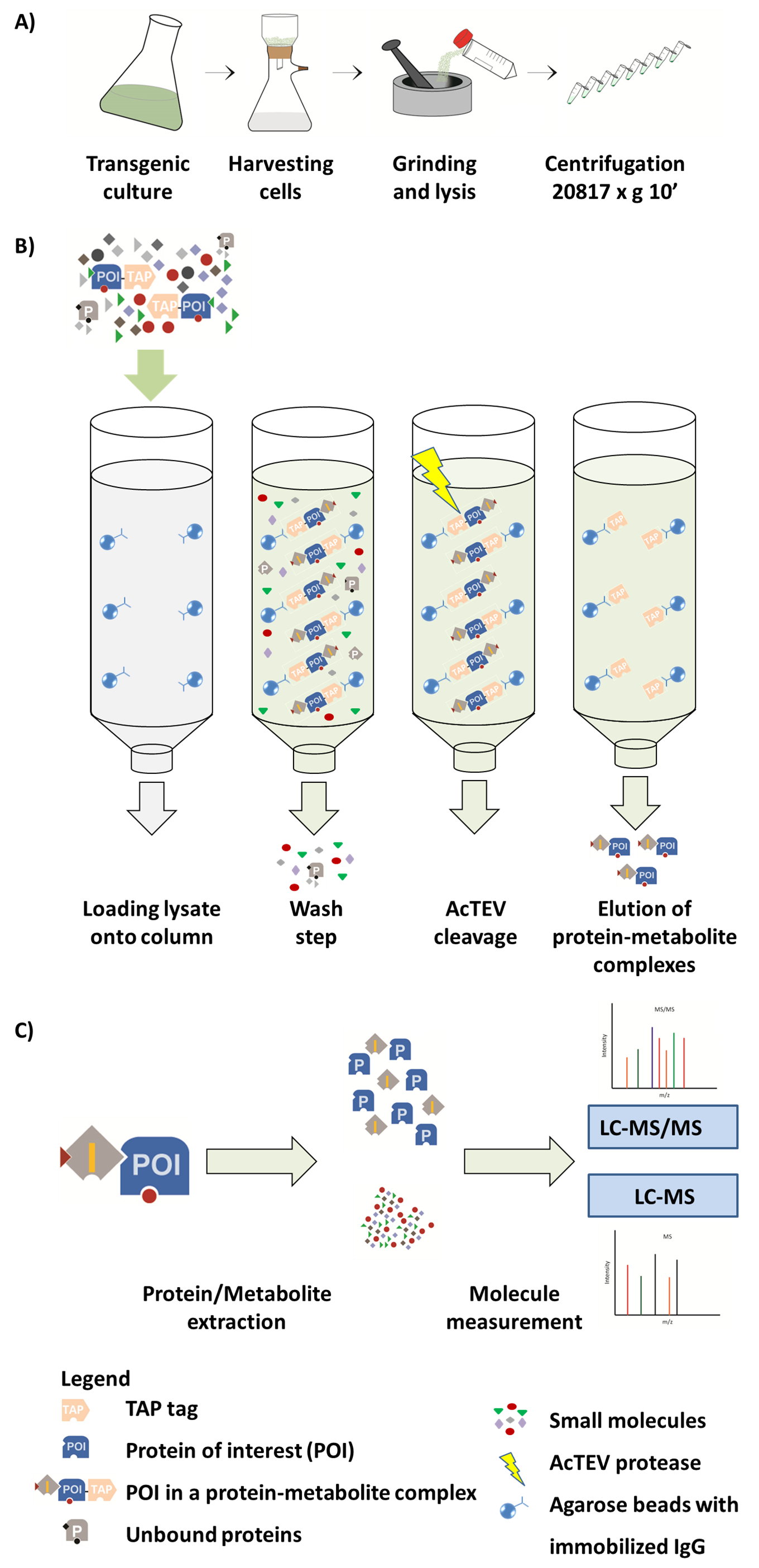{kind=link}

Рисунок 2. Дипептиды, специально совместно элюирующие с NDPK1. Средняя интенсивность четырех дипептиды Валь-леи (A), Иль-Glu (B), леи-Иль (C)и Иль-Пхе (D) измеряется в эксперименте на ЗС были нанесены. Все четыре дипептиды показывают значительные обогащения в NDPK1 образцах, по сравнению с контролем EV (звездочки представляют ФДР < 0.1). Представляют баров ошибка стандартная ошибка для 6 измерений (3 реплицирует N- и 3 C-неизлечимо меткой белки). Пожалуйста, нажмите здесь, чтобы посмотреть большую версию этой фигуры.
{kind=link}

Рисунок 3. Сети взаимодействия всех молекул элюирующие совместно с NDPK1, запросы стежка база данных рассматривает только предыдущих экспериментальных и базы данных свидетельства (доверие > 0.2). Больше доверия означает более высокие шансы взаимодействия и рассчитывается на основании хранение данных. Пожалуйста, нажмите здесь, чтобы посмотреть большую версию этой фигуры.
{kind=link}
Таблица S1. MaxQuant выходной таблицы «parameters.txt». Таблица включает пороговые значения для идентификации и количественной оценки, а также информация об используемых базах. Пожалуйста, нажмите здесь, чтобы загрузить этот файл.
Таблица S2. Информацию от MaxQuant выходной таблицы «proteinGroups.txt». Таблица содержит список всех выявленных белковых групп, интенсивности и дополнительной информации, например количество уникальных пептидов и оценка. Пожалуйста, нажмите здесь, чтобы загрузить этот файл.
Таблица S3. Выходной файл, содержащий анализ полярных метаболитов. Таблица содержит список всех выявленных массы возможностей, характеризуются конкретными m/z, RT и интенсивности. Пожалуйста, нажмите здесь, чтобы загрузить этот файл.
Таблица S4. Дипептиды обнаружен в пробах AP, в которых NDPK1, NDPK2 или NDPK3 были использованы в качестве приманки. Дипептиды, присутствующих в пустых проб были исключены из списка. Две независимые линии (отмеченных в N - или C-окончание) для каждого NDPK были запущены в трех экземплярах. Студента t-тест и дальнейшей коррекции p-значение с помощью Benjamini и метод Хёхберг использовались для определения значительно обогащенные interactor партнеров NDPKs (ФДР < 0.1). Учитывая ΔRT рассчитывается по отношению к ссылка соединений и Δppm по отношению к monoisotopic массы уступаны Метлин27. Пожалуйста, нажмите здесь, чтобы загрузить этот файл.
Таблица S5. Белки очищенные совместно с NDPK1. Были запущены две независимые линии (отмеченных в N - или C-окончание) для каждого NDPK в трех экземплярах. Студента t-тест и дальнейшей коррекции p-значение с помощью Benjamini и метод Хёхберг использовались для определения значительно обогащенные interactor партнеров NDPKs (ФДР < 0.1). Пожалуйста, нажмите здесь, чтобы загрузить этот файл.
Обсуждение
Представленные протокол позволяет параллельно идентификации PP и вечера комплексов целевого белка. От клонирования на окончательные результаты, эксперимент может быть завершена в качестве лишь 8-12 недель. Полная AP занимает около 4-6 ч для набора образцов 12-24, рендеринга нашей протокол для анализа средней пропускной способности.
Протокол, несмотря на в целом проста, имеет ряд важных шагов. (i) достаточное количество входных белка и близость бусины имеет решающее значение для достижения динамического диапазона обнаружения метаболит. Лизис эффективный клетки таким образом представляет собой решающий шаг в процедуре. Бедных белками урожайность может быть следствием недостаточной пульверизация материала или коэффициента субоптимальные лизис буфера/материал. (ii) следует позаботиться что используемые реагенты являются MS-чистые. Сильные моющие средства, глицерин или чрезмерного количества соли следует избегать, поскольку они мешают MS обнаружения. (iii) агарозы бусины не должно быть чрезмерно сухой во время стирки шаги, и при использовании вакуумного коллектора важно применять медленным потоком скоростью, так как не уничтожить бусы или повлиять на стабильность работы комплекса.
Есть некоторые важные возможные изменения, представленные протокол: (i) мы используем учредительного CaMV35S промоутер для максимального количества белка приманки. Гиперэкспрессия, в то время, как это очень полезно, может иметь серьезное воздействие на клетки гомеостаза28 и привести к образованию физиологически нерелевантных взаимодействий. Выражение тегами белков, с использованием собственных промоутеров и где возможно на фоне потери функции считается выше для получения истинной биологических посредники. Для белки, обычно не выражены в культуры клеток растений завод фона может оказаться необходимым определить соответствующие посредники. (ii) при работе с мембранных белков, литического буфера должна дополняться с MS-совместимых моющего средства. (iii) введение второй шаг очищение сродства может улучшить ложных срабатываний true срабатываний соотношение и устраняют необходимость управления EV29. Роман тандем тег с двумя независимыми протеаз расщепление сайтов представляет собой привлекательную альтернативу для размера-гель-проникающей хроматографии шаг добавил Маэда et al. 201411, который является трудоемким и требует много времени.
Наиболее серьезным недостатком AP является высокий уровень ложных срабатываний. Причины многочисленны. Учредительный гиперэкспрессия уже упоминалось. Еще одним источником физиологически нерелевантных взаимодействий, если работать с изолированной органеллы, является подготовка всего-lysates клетки содержащие смеси белков и метаболитов из различных внутриклеточных отсеков. Субцеллюлярные локализации должны использоваться для фильтрации для истинной посредники. Тем не менее большинство ложных срабатываний следствием неспецифической привязку между белками и агарозы смол. Введение второго шага очистки, как описано выше, предлагает лучшее решение для этой проблемы, однако, достигается за счет времени и пропускной способности. Кроме того слабые взаимодействия могут быть потеряны, поскольку протокол удлиняет. Еще один нюанс AP является то, что несмотря на всеобъемлющую информацию, которую он предоставляет о interactome целевого белка, различия между прямым и косвенным целями наживкой белка невозможно. Целевые bimolecular подходы необходимы для подтверждения взаимодействий.
AP, в сочетании с на основе MS метаболомики был использован для изучения белков комплексы в S. cerevisiae12. Эта работа, наряду с нашей ранее наблюдения13 , что аналогично в липиды, полярных и полу полярных соединений оставаться связанными белковых комплексов, изолированных от сотовой лизатов, предоставляет концептуальную основу представленные протокол. Наш протокол характеризуется три уникальных точек: (i) в отличие от дрожжей работают12, он демонстрирует, что AP подходит для извлечения не только гидрофобным но также гидрофильные белки лигандами. (ii) путем введения протокол извлечения три в одном, один AP может использоваться для изучения белков и метаболитов посредники приманки белка. (iii) мы адаптировали протокол к заводе клетки.
Будущие усилия будут сосредоточены на создание тега Роман тандем с двумя независимыми протеаз расщепление сайтов. Мы также хотели бы изучить пригодность протокола к низкой обилие небольших молекул, таких как растительные гормоны.
Раскрытие информации
Авторы не имеют ничего сообщать.
Благодарности
Мы хотели бы заранее признать профессор доктор Лотар Willmitzer за его участие в проекте, продуктивные дискуссии и большой надзор. Мы благодарны д-р Даниэль Veyel за помощь с proteomic MS измерений. Мы ценим миссис Änne Михаила, который предоставил нам неоценимую техническую помощь с измерениями LC-MS. Кроме того мы хотели бы поблагодарить д-р Моника Kosmacz и доктор Эвелина Sokołowska за их помощь и участие в работе на оригинальной рукописи и Вероника Jasińska для технической поддержки.
Материалы
| Name | Company | Catalog Number | Comments |
| Murashige and Skoog Basal Salts with minimal organics | Sigma-Aldrich | M6899 | |
| 1-Naphthylacetic acid | Sigma-Aldrich | N1641 | |
| Kinetin solution | Sigma-Aldrich | K3253 | |
| Tris base | Sigma-Aldrich | 10708976001 | |
| NaCl | Sigma-Aldrich | S7653 | |
| MgCl2 | Carl Roth | 2189.1 | |
| EDTA | Sigma-Aldrich | 3609 | |
| NaF | Sigma-Aldrich | S6776 | |
| DTT | Sigma-Aldrich | D0632 | |
| PMSF | Sigma-Aldrich | P7626 | |
| E-64 protease inhibitor | Sigma-Aldrich | E3132 | |
| Protease Inhibitor Cocktail | Sigma-Aldrich | P9599 | |
| Na3VO4 | Sigma-Aldrich | S6508 | |
| AcTEV Protease | Thermo Fischer Scientific | 12575015 | |
| Rotiphorese Gel 30 (37,5:1) | Carl Roth | 3029.2 | |
| TEMED | Carl Roth | 2367.3 | |
| PageRuler Prestained Protein Ladder | Thermo Fischer Scientific | 26616 | |
| SBP Tag Antibody (SB19-C4) | Santa Cruz Biotechnology | sc-101595 | |
| Goat anti-mouse IgG-HRP | Santa Cruz Biotechnology | sc-2005 | |
| Bradford Reagent | Sigma-Aldrich | B6916 | |
| Trypsin/Lys-C Mix, Mass Spec Grade | Promega | V5071 | |
| Urea | Sigma-Aldrich | U5128 | |
| Thiourea | Sigma-Aldrich | T8656 | |
| Ammonium bicarbonate | Sigma-Aldrich | 9830 | |
| Iodoacetamide | Sigma-Aldrich | I1149 | |
| MTBE | Biosolve | 138906 | |
| Methanol | Biosolve | 136806 | |
| Water | Biosolve | 232106 | |
| Acetonitrile | Biosolve | 12006 | |
| Trifluoroacetic acid | Biosolve | 202341 | |
| Formic acid | Biosolve | 69141 | |
| Unimax 2010 Platform Shaker | Heidolph | 5421002000 | |
| Nylon Mesh (Wire diameter 34 µM, thickness 55 µM, open area 14%) | Prosepa | Custom order | |
| Glass Funnel, 47 mm, 300 ml | Restek | KT953751-0000 | |
| Filter Bottle Top 500 mL 0,2 µM Pes St | VWR International GmbH | 514-0340 | |
| Mixer Mill MM 400 | Retsch GmbH | 207450001 | |
| IgG Sepharose 6 Fast Flow | GE Healthcare Life Sciences | 17-0969-02 | |
| Mobicol ""Classic"" with 2 different screw caps without filters | MoBiTec GmbH | M1002 | |
| Filter (small) 35 µM pore size, for Mobicol M 1002, M1003, M1050 & M1053 | MoBiTec GmbH | M513515 | |
| Variable Speed Tube Rotator SB 3 | Carl Roth | Y550.1 | |
| Rotary dishes for rotators SB 3 | Carl Roth | Y555.1 | |
| Resprep 24-Port SPE Manifolds | Restek | 26080 | |
| Finisterre C18/17% SPE Columns 100mg / 1ml | Teknokroma | TR-F034000 | |
| Autosampler Vials | Klaus Trott Chromatographie-Zubehör | 40 11 01 740 | |
| Acclaim PepMap 100 C18 LC Column | Thermo Fischer Scientific | 164534 | |
| EASY-nLC 1000 Liquid Chromatograph | Thermo Fischer Scientific | LC120 | |
| Q Exactive Plus Hybrid Quadrupole-Orbitrap Mass Spectrometer | Thermo Fischer Scientific | IQLAAEGAAPFALGMBDK | |
| Acquity UPLC system | Waters | Custom order | |
| ACQUITY UPLC HSS C18 Column, 100A, 1.8 µM, 2.1 mM X 100 mM, 1/pkg | Waters | 186003533 | |
| High-power ultrasonic cleaning baths for aqueous cleaning solutions | Bandelin | RK 31 | |
| Genedata Expressionist | Genedata | NaN | |
| Xcalibur Software | Thermo Fischer Scientific | NaN | |
| MaxQuant | NaN | NaN |
Ссылки
- Li, X., Snyder, M. Metabolites as global regulators: A new view of protein regulation. Bioessays. 33 (7), 485-489 (2011).
- Jacob, F., Monod, J. Genetic regulatory mechanisms in the synthesis of proteins. Journal of Molecular Biology. 3 (3), 318-356 (1961).
- Schlattner, U., et al. Dual Function of Mitochondrial Nm23-H4 Protein in Phosphotransfer and Intermembrane Transfer a cardiolipin-dependent switch. Journal of Biological Chemistry. 288 (1), 111-121 (2013).
- Ramírez, M. B., et al. GTP binding regulates cellular localization of Parkinson's disease-associated LRRK2. Human Molecular Genetics. , ddx161 (2017).
- Jung, H. J., Kwon, H. J. Target deconvolution of bioactive small molecules: the heart of chemical biology and drug discovery. Archives of Pharmacal Research. 38 (9), 1627-1641 (2015).
- Harding, M. W., Galat, A., Uehling, D. E., Schreiber, S. L. A receptor for the immunosuppressant FK506 is a cis-trans peptidyl-prolyl isomerase. Nature. 341 (6244), 758-760 (1989).
- Lomenick, B., et al. Target identification using drug affinity responsive target stability (DARTS). Proceedings of the National Academy of Sciences of the United States of America. 106 (51), 21984-21989 (2009).
- Manabe, Y., Mukai, M., Ito, S., Kato, N., Ueda, M. FLAG tagging by CuAAC and nanogram-scale purification of the target protein for a bioactive metabolite involved in circadian rhythmic leaf movement in Leguminosae. Chemical Communications. 46 (3), 469-471 (2010).
- Pantoliano, M. W., et al. High-density miniaturized thermal shift assays as a general strategy for drug discovery. Journal of Biomolecular Screening. 6 (6), 429-440 (2001).
- Li, X., Snyder, M. Analyzing In vivo Metabolite-Protein Interactions by Large-Scale Systematic Analyses. Current Protocols in Chemical Biology. , 181-196 (2010).
- Maeda, K., Poletto, M., Chiapparino, A., Gavin, A. -. C. A generic protocol for the purification and characterization of water-soluble complexes of affinity-tagged proteins and lipids. Nature Protocols. 9 (9), 2256-2266 (2014).
- Li, X., Gianoulis, T. A., Yip, K. Y., Gerstein, M., Snyder, M. Extensive in vivo metabolite-protein interactions revealed by large-scale systematic analyses. Cell. 143 (4), 639-650 (2010).
- Veyel, D., et al. System-wide detection of protein-small molecule complexes suggests extensive metabolite regulation in plants. Scientific Reports. 7, (2017).
- Luzarowski, M., et al. Affinity purification with metabolomic and proteomic analysis unravels diverse roles of nucleoside diphosphate kinases. Journal of Experimental Botany. , (2017).
- Van Leene, J., et al. Targeted interactomics reveals a complex core cell cycle machinery in Arabidopsis thaliana. Molecular systems biology. 6 (1), 397 (2010).
- Giavalisco, P., et al. Elemental formula annotation of polar and lipophilic metabolites using 13C, 15N and 34S isotope labelling, in combination with high-resolution mass spectrometry. The Plant Journal. 68 (2), 364-376 (2011).
- Van Leene, J., et al. Isolation of transcription factor complexes from Arabidopsis cell suspension cultures by tandem affinity purification. Plant Transcription Factors: Methods and Protocols. , 195-218 (2011).
- Van Leene, J., et al. A tandem affinity purification-based technology platform to study the cell cycle interactome in Arabidopsis thaliana. Molecular & Cellular Proteomics. 6 (7), 1226-1238 (2007).
- Olsen, J. V., Ong, S. -. E., Mann, M. Trypsin cleaves exclusively C-terminal to arginine and lysine residues. Molecular & Cellular Proteomics. 3 (6), 608-614 (2004).
- Cox, J., Mann, M. MaxQuant enables high peptide identification rates, individualized p.p.b.-range mass accuracies and proteome-wide protein quantification. Nature Biotechnology. 26 (12), 1367-1372 (2008).
- Cox, J., et al. Andromeda: A peptide search engine integrated into the MaxQuant environment. Journal of Proteome Research. 10 (4), 1794-1805 (2011).
- Tyanova, S., Temu, T., Cox, J. The MaxQuant computational platform for mass spectrometry-based shotgun proteomics. Nature Protocols. 11 (12), 2301 (2016).
- Hooper, C. M., et al. SUBAcon: a consensus algorithm for unifying the subcellular localization data of the Arabidopsis proteome. Bioinformatics. 30 (23), 3356-3364 (2014).
- Katajamaa, M., Orešič, M. Data processing for mass spectrometry-based metabolomics. Journal of Chromatography A. 1158 (1-2), 318-328 (2007).
- Szklarczyk, D., et al. STITCH 5: augmenting protein-chemical interaction networks with tissue and affinity data. Nucleic Acids Research. 1277, (2015).
- Tanaka, K. -. i., et al. Dipeptidyl compounds ameliorate the serum-deprivation-induced reduction in cell viability via the neurotrophin-activating effect in SH-SY5Y cells. Neurological Research. 34 (6), 619-622 (2012).
- Smith, C. A., et al. METLIN: A metabolite mass spectral database. Therapeutic Drug Monitoring. 27, 747-751 (2005).
- Bhattacharyya, S., et al. Transient protein-protein interactions perturb E. coli metabolome and cause gene dosage toxicity. Elife. 5, (2016).
- Rigaut, G., et al. A generic protein purification method for protein complex characterization and proteome exploration. Nature Biotechnology. 17 (10), 1030-1032 (1999).
Перепечатки и разрешения
Запросить разрешение на использование текста или рисунков этого JoVE статьи
Запросить разрешениеСмотреть дополнительные статьи
This article has been published
Video Coming Soon
Авторские права © 2025 MyJoVE Corporation. Все права защищены