
Method Article
2在 1: 一步亲和纯化为蛋白质蛋白质和蛋白质代谢物的平行的分析
摘要
蛋白质和蛋白质代谢物相互作用对所有细胞功能都至关重要。在这里, 我们描述了一个协议, 允许并行分析这些交互与蛋白质的选择。我们的协议优化了植物细胞培养, 并结合亲和纯化与质谱为基础的蛋白质和代谢物检测。
摘要
细胞过程由蛋白质、代谢物和核酸等生物分子之间的相互作用调节。虽然对蛋白质-蛋白质相互作用 (PPI) 的研究并不新奇, 但旨在表征内源性蛋白质代谢物相互作用 (PMI) 的实验方法构成了一个较近期的发展。在此, 我们提出一个协议, 允许同时描述 PPI 和 PMI 的蛋白质的选择, 称为诱饵。我们的协议优化了拟南芥细胞培养, 并结合亲和纯化 (AP) 与质谱 (MS) 为基础的蛋白质和代谢物检测。总之, 转基因拟南芥系, 表达诱饵蛋白融合到亲和标记, 首先裂解获得一个本地的细胞提取物。抗标记抗体是用来拉下蛋白质和代谢物的伙伴的诱饵蛋白。采用一步法甲基叔丁基醚 (MTBE)/甲醇/水法提取亲和纯化复合物。虽然代谢物分离成极性或疏水相, 蛋白质可以在颗粒中找到。然后用超效液相色谱-质谱 (UPLC-ms 或 UPLC-ms) 分析代谢产物和蛋白质。空矢量 (EV) 控制线用于排除误报。我们的协议的主要优点是, 它能够在近生理条件 (细胞裂解物) 中同时识别目标蛋白的蛋白质和代谢物伙伴。所提出的方法是直接的, 快速的, 可以很容易地适应生物系统以外的植物细胞培养。
引言
此处描述的方法旨在确定在体内细胞裂解条件下选择蛋白质的代谢物和蛋白质伙伴。据推测, 今天有许多比特征的代谢物具有重要的调节功能1。代谢物可以充当生物开关, 改变其受体蛋白2、3、4的活动、功能和/或定位。在过去十年中, 已经制定了一些突破性的方法, 能够在体内或近体条件下鉴定 PMI, 已经发展了5。可用方法可分为两组。第一组包括从已知代谢物诱饵开始的技术, 以诱捕新的蛋白质伙伴。方法包括亲和层析6、药物亲和反应靶-稳定性测定7、化疗蛋白质组学8和热蛋白质分析9。第二组由一个单一的方法, 开始与已知的蛋白质, 以确定小分子配体10,11。
采用 AP 结合 MS 为基础的 lipidomics 分析酵母蛋白脂复合物12.作为出发点, 作者使用酵母菌株表达21种酶参与麦角甾醇生物合成和103激酶融合到串联亲和纯化 (自来水) 标签。70% 的酶和20% 的激酶被发现绑定不同的疏水性配体, 脱落光进入复杂的蛋白质-脂质相互作用网络。
以前, 我们可以证明, 类似于脂质, 极性和半极性化合物也仍然绑定到从细胞裂解物13分离的蛋白质复合物。根据这些发现, 我们决定优化以前发表的10、11植物细胞和亲水性化合物14的 AP 方法。为此, 我们使用了 Van Leene等2010 所描述的抽头矢量, 成功地用于植物 PPI 研究15。为了缩短获得转基因品系所需的时间, 我们决定了拟南芥细胞培养。我们使用一步甲基叔丁基醚, (MTBE)/甲醇/水萃取方法, 允许蛋白质 (颗粒), 脂类 (有机相) 和亲水性代谢物 (水相)16在一个单一的亲和纯化实验。EV 控制线被引入, 以排除误报,例如蛋白质绑定到标签单独。作为概念的证明, 我们标记了三 (五) 核苷二磷酸激酶存在于拟南芥基因组 (NDPK1-NDPK3)。在其他发现中, 我们可以证明 NDPK1 与谷胱甘肽S转移酶和谷胱甘肽相互作用。因此, 我们可以证明 NDPK1 受 glutathionylation14。
综上所述, 所提出的协议是描述蛋白质和蛋白质-小分子相互作用网络的重要工具, 是对现有方法的重大进展。
研究方案
制备转基因拟南芥细胞培养线, 包括克隆、转化、选择和生长条件, 可在17中找到。请注意, EV 控制线建议纠正误报。在实验之前, 通过西方印迹分析确认诱饵蛋白的过度表达,例如对串联亲和标记的 g 蛋白部分使用 IgG 抗体。从植物细胞培养材料中分离生长培养基是很重要的。
1. 实验前制备植物细胞材料
- 生长一个 overexpressing 细胞培养线, 对18的蛋白质感兴趣。
- 制备 MSMO 培养基, 其中含有4.43 克/升 MSMO 混合30克/升蔗糖。将缓冲液的 pH 值调整为 5.7, 用1米 KOH 和高压釜解决。在实验之前, 补充培养基与0.5 毫克/升α萘乙酸酸, 0.05 毫克/升激动和50微克/毫升卡那霉素。
- 在一个100毫升的50毫升的 MSMO 培养基上, 用柔和的搅拌 (130 转每分钟), 在一个轨道平台上培养转化的植物细胞培养物。在20摄氏度的培养室中生长细胞, 光照强度等于80μmol m-2 s-1。
- 亚文化细胞每7天进入新鲜培养基, 稀释1:10。
- 在对数生长阶段收集细胞, 使用玻璃漏斗和真空泵结合使用尼龙网作为过滤器。包裹在铝箔中的渗透和冻结液氮。
注意: 请记住液氮非常冷。不正确的处理会导致烧伤。佩戴适当的个人防护设备, 包括隔热手套、防护眼镜和实验室大衣。
2. 点击协议
注: 以下步骤从 Maeda等201411和 Van Leene等201117适应。
- 融汇收获和冷冻植物细胞培养材料使用混合机 (2 分钟在20赫兹) 或砂浆和杵, 以获得细粉。整除3克的地面材料 (相当于大约90毫克的总蛋白) 每样。使用液氮预冷设备, 避免在这一步骤中解冻样品。
注: 贮存地面植物材料在50毫升管在–80°c 到 AP 程序的开始。 - Triturate 在液-氮-预换热器砂浆中, 用3毫升冰冷裂解缓冲剂 (0.025 米 Tris–HCl pH 7.5; 0.5 米氯化钠; 1.5 毫米氯化镁2; 0.5 毫米; 1 毫米 NaF; 1 毫米 Na3); 100x 稀释的商业蛋白酶抑制剂鸡尾酒; 4 毫米PMSF) 直到物质融化。一旦样品解冻, 立即进行下一步。
注: 准备溶解缓冲液新鲜。在此步骤中引入空白示例。不推荐使用洗涤剂, 因为它们会导致 MS 检测中出现问题。 - 要去除细胞碎片, 将材料分成2毫升离心管和离心机, 在 20817 x 克10分钟4摄氏度。在15毫升锥形离心管中收集3毫升的透明裂解液。
- 在离心过程中, 平衡 IgG-琼脂糖珠。整除100µL 的每样珠子, 并用1毫升的裂解缓冲液清洗。涡并用重悬珠和闪光旋转。放弃溶解缓冲, 重复步骤两次。并用重悬珠在400µL 的裂解缓冲液中。
- 在收集的植物裂解物中添加珠子, 在旋转轮上孵化混合物1小时, 在4摄氏度。
- 将混合物通过鲁尔锁帽与具有过滤孔径35µm 的自旋柱一起转移到注射器中. 施加压力通过裂解液。带附着配合物的珠子将留在过滤器上, 而裂解液会经过。
注: 可选, 使用真空歧管系统。一定要施加温和的压力, 以免伤害珠子。 - 先用10毫升洗涤缓冲器 (0.025 米 Tris–HCl ph 7.5; 0.5 米氯化钠), 再用1毫升洗脱缓冲器 (10 毫米 Tris–HCl pH 7.5; 150 毫米氯化钠; 0.5 毫米 EDTA; 1000x 稀释 E64 和1毫米 PMSF)。使用连接到柱或真空歧管系统的注射器进行洗涤。
注: 使用真空歧管系统时, 一定要施加温和的压力, 以免损害珠子。 - 孵化珠与400µL 洗脱缓冲器包含 50 U 的改进版本的烟草蚀刻病毒 (AcTEV) 蛋白酶。使用 1000 rpm 的表振动筛, 30 分钟16摄氏度。
注: 请记住使用插头关闭底部的列添加洗脱缓冲。 - 添加一个额外的部分 (50 U) 的酶入柱和孵化的混合物, 为未来30分钟在相同的, 上面描述的条件。
- 通过离心 (1 分钟, 20817 x g) 或真空歧管收集2毫升离心管中的洗脱液。要去除剩余的配合物, 引入一个额外的洗脱步骤使用200µL 洗脱缓冲。
注: 将样品贮存在–20°c 或–80°c, 或立即进行蛋白质和代谢物萃取步骤。在冰上解冻冰冻样品。
3. 西方污点分析
- 为了确认在收集的洗脱液中存在诱饵蛋白, 使用10µL 的 protein–metabolite 洗脱液进行 SDS–PAGE 和印迹分析。为了确定感兴趣的蛋白质, 使用小鼠主要抗体抗链亲和素结合蛋白 (1:200), TEV 蛋白酶解切后剩余的抽头标记的一部分, 如 Van Leene等201117所述。其次, 使用二次山羊抗鼠抗体与 HRP 结合。
4. 代谢物和蛋白质提取
注: 本协议适用于 Giavalisco等201116。
注意: 从这一步开始使用 UPLC–MS–grade 解决方案。
- 将1毫升的甲基叔丁基醚 (MTBE)/甲醇/水溶剂 (3:1: 1) 添加到所收集的洗脱液中, 并通过反转来混合样品。确保溶剂在萃取步骤之前冷却到–20°c。
注意: 甲基叔丁基醚和甲醇是有害物质。在油烟机下执行萃取步骤, 并佩戴适当的个人防护设备,如手套。 - 加入0.4 毫升甲醇: 水1:3 溶液对每个样品和混合样品的内容通过倒置。
注: 用甲醇补充混合物: 水溶液导致相分离。上部阶段包含脂质, 较低的阶段包含极性和半极性代谢物, 并且蛋白质在药丸能被发现。 - 分离阶段, 离心样品在 20817 x g 在室温下2分钟, 然后收集上阶段的脂质测量 (没有在本协议中) 使用手动液体处理吸管与1毫升容积容量。
- 加入0.2 毫升甲醇, 并通过反转混合。
- 离心样品在 20817 x g 在 RT 2 分钟, 然后收集极性阶段为代谢物测量 (极性和半极性化合物)。为了避免干扰蛋白颗粒, 在管的底部留下大约50µL 的液相。
- 在离心蒸发器中过夜的代谢物测量的干收集样品。避免在30–60分钟后从蒸发器中取出样品, 以免过度干燥蛋白颗粒。
注: 贮存样品在–20°c 或–80°c, 或立即着手准备蛋白质的 LC–MS/MS 分析。
5. 为蛋白质组分析准备样品
注意: 这一步是由奥尔森等。200419和胰蛋白酶/赖氨酸 C 混合的技术手册 (见材料表)。
- 对样品进行酶消化。
注意: 在酶消化和样品脱盐过程中使用的溶剂是有害的。在油烟机下工作, 佩戴适当的个人防护设备,如手套。- 在30µL 的新制备变性缓冲液中溶解蛋白颗粒 (40 毫米含2米 thiourea/6 尿素的碳酸铵, pH 为 8)。为了达到更好的蛋白质溶解度, 执行15分钟超声波步骤。重复步骤, 直到颗粒溶解。
- 离心样品在 20817 x g 10 分钟在4°c, 然后转移上清到一个新的离心管。
- 用布拉德福德蛋白测定法测定蛋白质浓度。
- 为进一步分析, 整除的体积相当于100µg 的蛋白质, 并填充样本多达46µL 变性缓冲。
- 加入2µL 的新制备的还原缓冲剂 (50 毫米在 H2O 中溶解), 室温下孵育30分钟。
- 用2µL 的新制备的烷基化反应缓冲剂 (150 毫米 iodoacetamide 溶解在40毫米的碳酸铵缓冲液中), 在室温下将混合物在黑暗中孵化20分钟。
- 稀释样品与30µL 40 毫米碳酸氢铵缓冲和增加20µL LysC/胰蛋白酶混合物。
- 4小时后孵化37摄氏度, 稀释样品与300µL 40 毫米碳酸氢铵缓冲。
- 继续在37摄氏度的夜间孵化。
- 酸化样品与大约20µL 10% 三氟乙酸酸 (TFA) 获得 pH 值 < 2。使用 ph 条检查样品 ph 值。
注: 将样品存储在–20°c 或继续下一步。
- 淡化被消化的蛋白质。
注: 最好使用真空歧管系统。避免列过度干燥。- 冲洗 C18 SPE 柱 (见材料表) 与1毫升100% 甲醇, 然后1毫升80% 乙腈 (ACN) 含 0.1% TFA 稀释在水中。使用, 在这里和进一步的蛋白质脱盐步骤, 一个真空流形系统, 以加快进程。避免列过度干燥。
- 用1毫升的 0.1% TFA 稀释在水中, 平衡该柱。
- 将该示例加载到该列上。冲洗管与额外的200µL 0.1% TFA 和转移的解决方案的列。通过该列运行解决方案。
- 用1毫升 0.1% TFA 洗两次柱。
- 洗脱淡化肽从柱800µL 60% ACN, 0.1% TFA 溶液。在离心蒸发器中干燥所收集的分数, 避免在30–60后从蒸发器中取出样品, 以免蛋白质组分过度干燥。
注: 在–20°c 或–80°c 储存样品或立即进行下一步。在随后的步骤中, 将样品放在冰上。
6. 使用 UPLC–MS/MS 测定制备的蛋白质样品。
注: 在蛋白质组和 metabolomic 测量之前, 过滤器 (0.2 µm 孔大小) 和德加德所有缓冲器使用真空泵为1小时。
- 并用重悬干肽颗粒储存在2毫升离心管在50µL 的缓冲 C (3% 伏/五 ACN, 0.1% v/v 甲酸) 使用手动液体处理吸管与200µL 容积容量。油脂实验样品15分钟, 在超声浴与35赫超声波频率。
注意: ACN 和甲酸是有害物质。在油烟机下工作, 佩戴适当的个人防护设备,如手套。 - 离心样品在 20817 x g 10 分钟在4°c, 然后转移20µL 的上清到玻璃瓶。
- 用 C18 反相柱连接到液相色谱分离消化肽, 利用质谱仪获得质谱。
- 使用 300-µL/分钟流量, 在样本的3列上分离。对于移动阶段, 使用缓冲 C 和 D (63% 伏/v ACN, 0.1% 伏/v 甲酸), 形成梯度从 3% ACN 到 15% ACN 超过20分钟, 然后到 30% ACN 在未来10分钟。
注: 将样品的其余部分保存在–20°c 或–80°c, 最多数月。在蛋白质组测量之前, 冻结在冰上的样品。 - 用 60% ACN 清洗污染物10分钟, 平衡5µL 的列, 然后再测量下一个样品。
- 利用数据依赖的 ms/毫秒方法获得质量谱, 分辨率设置为 7万, 3e6离子的 AGC 目标, 最大注入时间为100毫秒, 从300到1600不等的 m/z。获得最大15毫秒/毫秒扫描分辨率为 17500, AGC 目标为 1e5, 最大注入时间为100毫秒, 底充胶比率为 20%, 与隔离窗口1.6 米/z 和 m/z 不等, 从200到2000。启用顶点触发器 (6–20 s), 将动态排除设置为十五年代, 并排除1和 > 5 的费用。
- 使用 300-µL/分钟流量, 在样本的3列上分离。对于移动阶段, 使用缓冲 C 和 D (63% 伏/v ACN, 0.1% 伏/v 甲酸), 形成梯度从 3% ACN 到 15% ACN 超过20分钟, 然后到 30% ACN 在未来10分钟。
7. 蛋白质组数据的处理
- 从 http://www.uniprot.org/下载最新拟南芥蛋白质组数据库, 包括污染物数据库。分析从 LC–MS 运行获得的原始数据使用 MaxQuant 与集成的仙女座肽搜索引擎使用默认设置与启用 LFQ 正常化20,21,22。查找有关表 S1中使用的参数的详细信息。
- 打开 "蛋白质 groups.txt" 输出文件。为了进一步分析, 筛选出至少有两个独特的肽的蛋白质组。删除由 MaxQuant 定义的蛋白质组作为潜在的污染物和筛选的. 芥蛋白 (ARATH 在 Fasta 头专栏) 存在于数据库中。
- 为了检验样品间蛋白质富集的意义, 使用 LFQ 规范化强度, 进行配对, 两尾学生的 t 检验, 然后进行多重比较校正 (如Benjamini & 霍赫贝格假发现率 (罗斯福)更正或 Bonferroni 更正)。
- 通过比较获得的 EV 控制和 NDPK1 的 LFQ 强度计算 p 值。筛选出所有待定值。按升序对 p 值进行排序, 并使用 R 脚本或在线计算器 (如https://www.sdmproject.com/utilities/?show=FDR) 计算罗斯福更正。筛选罗斯福价值低于0.1。
注: 考虑数据分析的形式适合于研究。定量研究 (样品间蛋白质富集分析) 使用 "LFQ 强度" 值, 而对于定性研究 (存在或缺乏特定的蛋白质) 选择 "强度" 值。 - 筛选 NDPK1 比 EV 控制更丰富的蛋白质组。确定潜在的蛋白质合作伙伴的本地化使用 SUBA 数据库23和正确的蛋白质与 NDPK1 的共同本地化。
- 通过比较获得的 EV 控制和 NDPK1 的 LFQ 强度计算 p 值。筛选出所有待定值。按升序对 p 值进行排序, 并使用 R 脚本或在线计算器 (如https://www.sdmproject.com/utilities/?show=FDR) 计算罗斯福更正。筛选罗斯福价值低于0.1。
8. 使用 UPLC–MS 测量含有极性相的样品。
- 并用重悬干燥极性阶段从步骤4.5 在200µL 水和油脂实验样品为5分钟。
- 离心样品在 20817 x g 10 分钟在4°c, 然后转移上清到玻璃瓶。
注: 将样品的其余部分储存在–20°c 或–80°c, 可达数月。在 metabolomic 测量之前, 冻结冰上的样品。 - 使用 UPLC 耦合 C18 反相柱进行分离步骤, 并获得质谱与 MS。
- 将每个电离模式 (正和负) 上的每个注入样品的2µL 列加载到该柱上, 并使用400µL/最小流速分隔分数。要创建代谢物测量所需的梯度, 请准备移动相解, 如下所示: 缓冲 A (0.1% 甲酸在 H2O) 和缓冲 B (0.1% 甲酸在 ACN)。
- 分离代谢物在400µL/分钟和以下梯度: 1 分钟99% 缓冲 a, 11 分钟线性梯度从99% 缓冲 a 到60% 缓冲 a, 13 min 线性梯度从60% 缓冲 a 到30% 缓冲 a, 15 min 线性梯度从30% 缓冲 a 到1%缓冲 a, 保持1% 浓度, 直到16分钟从17分钟开始, 使用线性渐变从1% 缓冲区 a 到99% 缓冲区 a. 重新平衡列为3分钟与99% 集中缓冲 a 在测量下一个样品之前。
- 获得100至1500米/z 的质量谱, 分辨率设置为 2.5万, 加载时间限制为100毫秒. 设置 AGC 目标为 1e6, 毛细管电压为 3kV, 带鞘气流量和辅助气体值60和20分别。将毛细管温度设置为250°c 和撇嘴电压为25V。
9. 新陈代谢数据的处理
- 从两种电离模式中采集的图谱。使用软件提取质量的电荷比 (m/z), 保留时间 (RT) 和强度的相关峰值,例如,商业软件 (见材料表) 或备选案文24。
- 通过双击 .exe 文件开始处理软件
- 创建新工作流, 搜索活动 "从文件加载" 并将此活动通过 "拖放" 移动到空白工作流空间中。用鼠标右键按活动并打开活动设置。
- 在包含 "常规" 设置的选项卡中, 在 "名称" 字段中设置实验名称, 然后单击 "选择文件和文件夹" 并标记原始图谱。
- 在包含 "高级" 设置的选项卡中, 将 "配置文件数据截止" 设置为0强度。单击 "应用" 和 "确定"。
- 搜索并添加活动 "数据扫描"。用鼠标右键按活动并打开活动设置。
- 在包含 "常规" 设置的选项卡中, 标记 "质心数据" 和 "ms/ms 数据"。通过在 "选择" 面板中选择 "全部", 删除所有 ms/毫秒数据。
- 搜索并添加活动 "色谱化学噪声减法"。用鼠标右键按活动并打开活动设置。
- 在包含 "常规" 设置的选项卡中, 标记 "色谱平滑", 并将扫描次数设置为 "3" 和 "估计值" 到 "移动平均值"。将 "RT 窗口" 设置为51扫描, "分量" 为 50%, 减法 "方法" 和750强度 "阈值"。
- 在包含 "高级" 设置的选项卡中, 标记 "rt 结构删除", 并将 "最小 rt 长度" 设置为5扫描。
- 在包含 "高级" 设置的选项卡中, 标记 "m/z 结构删除", 并将 "最小 m-/z 长度" 设置为3磅。
- 搜索并添加活动 "色谱 RT 对齐"。用鼠标右键按活动并打开活动设置。
- 在包含 "常规" 设置的选项卡中, 将 "校准方案" 设置为 "对齐基准树" 和 "RT 搜索间隔", 以0.5 分钟为准。
- 在包含 "高级" 设置的选项卡中, 使用默认参数。
- 搜索并添加 "色谱" 组活动中的 "峰值检测" 活动。用鼠标右键按活动并打开活动设置。
- 在包含 "常规" 设置的选项卡中, 将 "求和窗口" 设置为0.09 分钟, "最小峰值大小" 为0.03 分钟, "最大合并距离" 为5磅, "合并策略" 为 "中心"。在 "峰值 RT 分裂" 框设置 "间隙/峰值比率" 到50%。
- 在包含 "高级" 设置的选项卡中, 将 "平滑窗口" 设置为5磅, "细化阈值" 为80% 和 "一致性阈值" 为1。设置 "中心计算" 为 "强度加权" 的 "强度阈值" 设置为70%。
- 在 "色谱" 活动组中搜索并添加活动 "同位素聚类"。用鼠标右键按活动并打开活动设置。
- 在包含 "常规" 设置的选项卡中, 将 "RT 公差" 设置为0.015 分钟和 "m/z 公差" 为 5 ppm。
- 在包含 "信封管接头" 设置的选项卡中, 将 "方法" 设置为 "无形状限制" 和 "电离" 为 "质子 (为正模式)" 和 "Deprotonation" (用于负模式)。分别将 "最小和最大电荷" 设置为1和4。
- 在包含 "高级" 设置的选项卡中, 使用默认参数。
- 搜索并添加活动 "单一筛选器"。
- 要导出数据处理的结果, 请在 "导出" 活动组中搜索并添加活动 "分析师"。
- 在包含 "常规" 设置的选项卡中, 将 "类型" 设置为 "簇", "可观察" 为 "汇总强度"。选择 "自定义目标" 并指定导出文件的目录。
- 在包含 "高级" 设置的选项卡中, 使用默认参数。
- 使用内部引用复合数据库对质量特征进行注释。
- 用 UPLC-ms 分析一个或多个 MS 级参考化合物. 使用相同的 LC-ms 方法分析参考化合物和代谢产物, 并与感兴趣的蛋白质进行共纯化。
注: 本研究将一组近300合成二肽进行分析, 作为参考复合库。 - 使用分析 (见材料表) 软件打开原始色谱文件, 并搜索特定的 m/z 和 RT 相关的测量参照化合物 (见用户指南)。
注: 次生代谢物在电离类型上有差异。通过搜索等于 M-1.007276、m + 1.007276、m + 18.033823 和 m + 22.989218 (m h)、[m + H]、[m + NH4] 和 [m + Na] 的离子质量, 检查常见加合物是否存在。 - 使用电子表格打开导出的 "分析" 文件, 在处理图谱后获得, 并搜索特定的离子质量。比较实验中测量的质量特征和参考化合物的 rt。允许偏差 0.005 Da 为 m/z 和0.1 分钟的 RT。
- 用 UPLC-ms 分析一个或多个 MS 级参考化合物. 使用相同的 LC-ms 方法分析参考化合物和代谢产物, 并与感兴趣的蛋白质进行共纯化。
- 为了检验在样品 (抗原蛋白与 EV 控制) 之间用特定蛋白质对代谢产物进行富集共纯化的意义, 用两尾非配对学生的 t 检验来比较峰值值, 后跟多个比较校正 (例如Benjamini & 霍赫贝格错误发现率更正或 Bonferroni 更正)。
结果
在原研究中, 三A. NDPK 基因在抗原细胞悬浮培养物的控制下, 在本构35S 启动子14 (图 1) 下进行。串联亲和标记被融合到一个诱饵蛋白的羧基或氨基端端。亲和纯化复合物受叔丁基醚/甲醇/水萃取16。用 MS (表 S2 和 S3) 识别亲和拉蛋白和小分子。
为了纠正误报, 用空白样本排除了化学物质和实验室消耗品中的小分子污染物。此外, 使用 EV 控制线来计算与亲和标记或树脂单独结合的代谢物和蛋白质。为了检索真正的阳性, 双尾非配对学生的 t 检验和 Benjamini & 霍赫贝格假发现率校正被用于识别代谢物 (表 S4) 和蛋白质 (表 S5) 显著丰富的 NDPKs AP实验 (N 和 C 末期标记 NDPKs) 与 EV 控制线 (罗斯福 < 0.1) 比较。请注意, 在以前的工作中, 我们使用缺勤/存在标准来描绘蛋白质和小分子 interactors。
给出了 NDPK1 的代表性结果, 而代谢物数据集中于合成二肽, 这是本组研究的一种新型小分子调节器。蛋白质组分析揭示了 26 NDPK1 的蛋白质合作伙伴。通过进一步过滤蛋白质与 NDPK1 (细胞质) 在同一个亚细胞室中的联合定位, 该清单缩小到13假定的蛋白质 interactors。在鉴定的蛋白质中有谷胱甘肽S转移酶、两个伸长起始因子、蛋白和 aconitate 水合。Metabolomic 分析揭示了四合成二肽, 即谷、伊、列和洗脱与 NDPK1 (图 2)。请注意, 所有四合成二肽在其 n-端共用疏水性残渣, 表明具有共同的结合特异性。
为了寻找已知的蛋白质-蛋白质和蛋白质代谢物络合物, 我们查询了13识别的蛋白质和四合成二肽对针数据库25 (图 3)。可以提出几点意见: (一) interactors 以前没有 NDPK1 的报告。(ii) 报告 APX1 直系同源与醛脱氢酶家族成员 ALDH7B4 相互作用, 而翻译起始因子 FBR12 以基因 AT2G40290 编码的另一种转化起始因子。(三) 确定的合成二肽没有报告的蛋白质伙伴。洗脱合成二肽未被报告为与任何检索到的植物蛋白相关联。然而, 它们在其他生物体中扮演着重要的角色: 列伊岛,例如, 在人类细胞26行中具有神经营养因子激活作用。请注意, 该实验不允许识别系统的精确拓扑。例如, 二肽可能直接与 NDPK1 相互作用, 但很可能与任何共同纯化的蛋白质有关。
结合起来, 我们的研究结果表明, 建立的程序, 采用 AP 和质谱, 促进蛋白质蛋白和蛋白质-小分子 interactors 的鉴定, 并有助于产生广泛的信息,目标蛋白的 interactome。

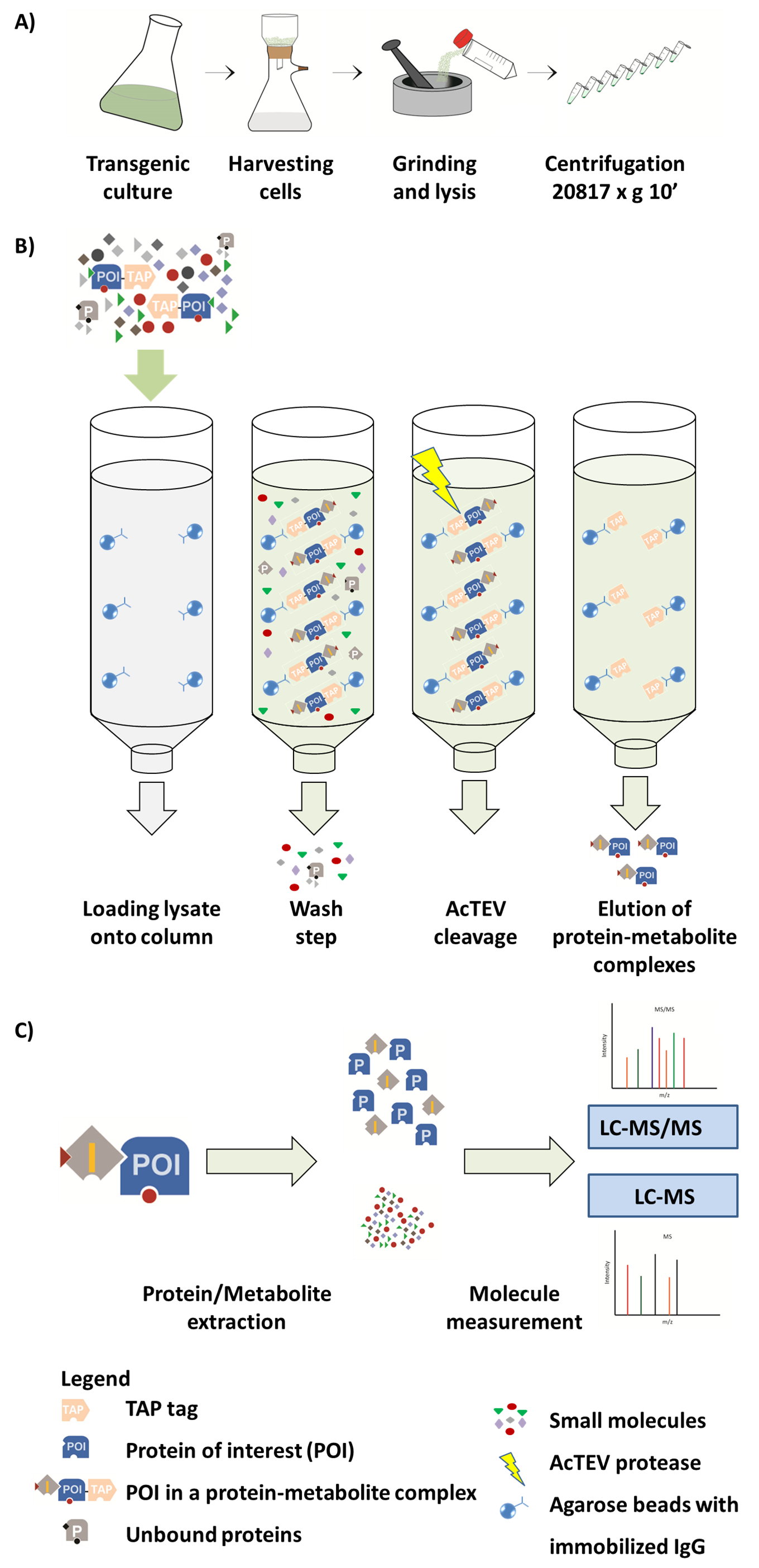
图1。AP-MS 工作流方案。(a)从植物细胞培养中制备本族可溶性分数。(B) AP 程序中的下一步步骤。将样品加载到柱上后, 将所附的兴趣蛋白 (点) 与一个分路标记结合, 并与固定在琼脂糖珠上的 IgG 抗体结合。洗涤柱有助于去除未绑定的蛋白质和代谢物。AcTEV 卵裂后, 洗脱蛋白代谢物复合体。(C)将络合物分离成蛋白质和代谢物分数, 然后进行半定量 MS 分析。这个数字的一部分是从 Luzarowski等201714转载。请单击此处查看此图的较大版本.
{kind=link}

图2。合成二肽专门与 NDPK1 共同洗脱.绘制了 AP 试验中测定的四合成二肽(A)、谷 ( B)、列伊岛(C)和(D)的平均强度。与 EV 控制 (星号代表罗斯福 < 0.1) 相比, 所有四合成二肽在 NDPK1 样品中均显着丰富。误差线表示6测量的标准错误 (3 复制 N 和3的 C 末期标记蛋白)。请单击此处查看此图的较大版本.
{kind=link}

图3。所有分子的相互作用网络与 NDPK1 共洗脱, 查询对缝合数据库仅考虑以前的实验和数据库证据 (信心 > 0.2).较高的置信度表明了较高的交互几率, 并根据沉积数据进行了计算。请单击此处查看此图的较大版本.
{kind=link}
表 S1。MaxQuant 输出表 "parameters.txt"。表包括用于标识和量化的阈值, 以及有关所用数据库的信息。请单击此处下载此文件.
表 S2。来自 MaxQuant 输出表 "proteinGroups.txt" 的信息。表包含所有已识别的蛋白质组、强度和其他信息 (如独特的多肽和分数) 的列表。请单击此处下载此文件.
表 S3。含有极性代谢物分析的输出文件。表包含所有已识别的质量特征的列表, 其特征是特定的 m-/z、RT 和强度。请单击此处下载此文件.
表 S4。合成二肽发现的 AP 样品中, NDPK1, NDPK2 或 NDPK3 被用作诱饵。合成二肽在空白样品中的显示被排除在名单之外。两条独立的线 (标记在 N 或 C 总站) 为每个 NDPK 的运行三个三。采用 Benjamini & 霍赫贝格法对学生进行t检验并对p值进行进一步修正, 以确定 NDPKs (罗斯福 < 0.1) 显著丰富的 interactor 合作伙伴。给出的是ΔRT 计算与参考化合物和Δppm 有关的 monoisotopic 质量在 Metlin27。请单击此处下载此文件.
表 S5。蛋白质与 NDPK1 共纯化。两条独立的线 (标记在 N 或 C 总站) 为每个 NDPK 的运行三个三。采用 Benjamini & 霍赫贝格法对学生进行t检验并对p值进行进一步修正, 以确定 NDPKs (罗斯福 < 0.1) 显著丰富的 interactor 合作伙伴。请单击此处下载此文件.
讨论
所提出的协议允许平行识别靶蛋白的 PP 和 PM 复合物。从克隆到最终结果, 实验可以在8-12 周内完成。完整 AP 需要大约4-6 小时的12到24个样本, 使我们的协议适用于中吞吐量分析。
尽管该议定书总体上直截了当, 但仍有一些关键步骤。(i) 足够数量的输入蛋白和亲和珠对于达到动态的代谢产物检测至关重要。因此, 有效的细胞裂解是过程中的关键步骤。不良的蛋白质产量可能是由于材料的粉碎不足或次优裂解-缓冲/物质比的结果。(二) 应注意使用的试剂是 MS 友好的。在干扰 MS 检测时, 应避免使用强力洗涤剂、甘油或过量食盐。(iii) 在洗涤步骤中, 琼脂糖珠不应过度干燥, 当使用真空歧管时, 应用慢速流速, 以免破坏珠子或影响复杂的稳定性是很重要的。
对所提出的协议有一些重要的可能的修改: (i) 我们使用本构 CaMV35S 启动子来最大化诱饵蛋白的数量。过度表达虽然非常有用, 但会对细胞稳态28产生严重影响, 导致生理上无关的相互作用的形成。标记蛋白的表达使用本地启动器, 并在可能的功能损失的背景被认为优于检索真正的生物 interactors。对于通常没有在植物细胞培养中表达的蛋白质, 植物的背景可能证明有必要确定相关的 interactors。(ii) 在使用膜蛋白时, 溶解缓冲液需要辅以 MS 兼容洗涤剂。(iii) 引入第二亲和纯化步骤可提高误报率, 并消除对 EV 控制29的需要。一个新的串联标签与两个独立的蛋白酶解的网站提出了一个有吸引力的替代的大小排除色谱步骤添加的 Maeda等201411, 这既费力又费时。
AP 最严重的缺点是误报率高。原因很多。本构表达已被提及。另一种生理上无关的相互作用的来源, 除非与孤立的细胞器, 是制备全细胞裂解物包含蛋白质和代谢物的不同亚细胞间的混合物。亚细胞定位应用于筛选真正的 interactors。然而, 大多数假阳性是由蛋白质和琼脂糖树脂之间不特异的结合引起的。如上文所述, 引入第二个净化步骤提供了解决问题的最佳方法, 但代价是时间和吞吐量。此外, 随着协议的延长, 较弱的交互可能会丢失。AP 的另一个告诫是, 尽管它提供了关于目标蛋白 interactome 的全面信息, 但在诱饵蛋白的直接和间接目标之间进行区分是不可能的。需要有针对性的分子方法来确认交互。
采用 AP 结合 MS 为基础的新陈代谢用于研究12的酵母蛋白复合物.这项工作, 连同我们早先的观察13 , 类似于脂质, 极性和半极性化合物仍然绑定到从细胞裂解物分离的蛋白质复合物, 为提出的议定书提供了概念基础。我们的协议的特点是三独特的点: (i) 与酵母工作12相比, 它表明, AP 是适合检索不仅疏水性, 而且亲水蛋白配体。(ii) 通过引入一个三入一萃取协议, 单一 AP 可以用来研究蛋白和代谢物 interactors 的诱饵蛋白。(iii) 我们修改了该议定书以种植细胞。
未来的努力将集中在创建一个新的串联标签与两个独立的蛋白酶解的网站。我们还想探讨该议定书对低丰度小分子如植物激素的适用性。
披露声明
作者没有什么可透露的。
致谢
我们谨向 Willmitzer 博士表示感谢, 感谢他参与了该项目, 进行了富有成效的讨论, 并进行了重大监督。我们感谢丹尼尔 Veyel 博士对蛋白质组 MS 测量的帮助。我们感谢Änne 米氏夫人, 他为我们提供了宝贵的技术帮助与 LC MS 测量。此外, 我们还要感谢莫妮卡 Kosmacz 博士和 Ewelina 博士 Sokołowska 对原手稿工作的帮助和参与, 并向维罗尼卡 Jasińska 提供技术支持。
材料
| Name | Company | Catalog Number | Comments |
| Murashige and Skoog Basal Salts with minimal organics | Sigma-Aldrich | M6899 | |
| 1-Naphthylacetic acid | Sigma-Aldrich | N1641 | |
| Kinetin solution | Sigma-Aldrich | K3253 | |
| Tris base | Sigma-Aldrich | 10708976001 | |
| NaCl | Sigma-Aldrich | S7653 | |
| MgCl2 | Carl Roth | 2189.1 | |
| EDTA | Sigma-Aldrich | 3609 | |
| NaF | Sigma-Aldrich | S6776 | |
| DTT | Sigma-Aldrich | D0632 | |
| PMSF | Sigma-Aldrich | P7626 | |
| E-64 protease inhibitor | Sigma-Aldrich | E3132 | |
| Protease Inhibitor Cocktail | Sigma-Aldrich | P9599 | |
| Na3VO4 | Sigma-Aldrich | S6508 | |
| AcTEV Protease | Thermo Fischer Scientific | 12575015 | |
| Rotiphorese Gel 30 (37,5:1) | Carl Roth | 3029.2 | |
| TEMED | Carl Roth | 2367.3 | |
| PageRuler Prestained Protein Ladder | Thermo Fischer Scientific | 26616 | |
| SBP Tag Antibody (SB19-C4) | Santa Cruz Biotechnology | sc-101595 | |
| Goat anti-mouse IgG-HRP | Santa Cruz Biotechnology | sc-2005 | |
| Bradford Reagent | Sigma-Aldrich | B6916 | |
| Trypsin/Lys-C Mix, Mass Spec Grade | Promega | V5071 | |
| Urea | Sigma-Aldrich | U5128 | |
| Thiourea | Sigma-Aldrich | T8656 | |
| Ammonium bicarbonate | Sigma-Aldrich | 9830 | |
| Iodoacetamide | Sigma-Aldrich | I1149 | |
| MTBE | Biosolve | 138906 | |
| Methanol | Biosolve | 136806 | |
| Water | Biosolve | 232106 | |
| Acetonitrile | Biosolve | 12006 | |
| Trifluoroacetic acid | Biosolve | 202341 | |
| Formic acid | Biosolve | 69141 | |
| Unimax 2010 Platform Shaker | Heidolph | 5421002000 | |
| Nylon Mesh (Wire diameter 34 µM, thickness 55 µM, open area 14%) | Prosepa | Custom order | |
| Glass Funnel, 47 mm, 300 ml | Restek | KT953751-0000 | |
| Filter Bottle Top 500 mL 0,2 µM Pes St | VWR International GmbH | 514-0340 | |
| Mixer Mill MM 400 | Retsch GmbH | 207450001 | |
| IgG Sepharose 6 Fast Flow | GE Healthcare Life Sciences | 17-0969-02 | |
| Mobicol ""Classic"" with 2 different screw caps without filters | MoBiTec GmbH | M1002 | |
| Filter (small) 35 µM pore size, for Mobicol M 1002, M1003, M1050 & M1053 | MoBiTec GmbH | M513515 | |
| Variable Speed Tube Rotator SB 3 | Carl Roth | Y550.1 | |
| Rotary dishes for rotators SB 3 | Carl Roth | Y555.1 | |
| Resprep 24-Port SPE Manifolds | Restek | 26080 | |
| Finisterre C18/17% SPE Columns 100mg / 1ml | Teknokroma | TR-F034000 | |
| Autosampler Vials | Klaus Trott Chromatographie-Zubehör | 40 11 01 740 | |
| Acclaim PepMap 100 C18 LC Column | Thermo Fischer Scientific | 164534 | |
| EASY-nLC 1000 Liquid Chromatograph | Thermo Fischer Scientific | LC120 | |
| Q Exactive Plus Hybrid Quadrupole-Orbitrap Mass Spectrometer | Thermo Fischer Scientific | IQLAAEGAAPFALGMBDK | |
| Acquity UPLC system | Waters | Custom order | |
| ACQUITY UPLC HSS C18 Column, 100A, 1.8 µM, 2.1 mM X 100 mM, 1/pkg | Waters | 186003533 | |
| High-power ultrasonic cleaning baths for aqueous cleaning solutions | Bandelin | RK 31 | |
| Genedata Expressionist | Genedata | NaN | |
| Xcalibur Software | Thermo Fischer Scientific | NaN | |
| MaxQuant | NaN | NaN |
参考文献
- Li, X., Snyder, M. Metabolites as global regulators: A new view of protein regulation. Bioessays. 33 (7), 485-489 (2011).
- Jacob, F., Monod, J. Genetic regulatory mechanisms in the synthesis of proteins. Journal of Molecular Biology. 3 (3), 318-356 (1961).
- Schlattner, U., et al. Dual Function of Mitochondrial Nm23-H4 Protein in Phosphotransfer and Intermembrane Transfer a cardiolipin-dependent switch. Journal of Biological Chemistry. 288 (1), 111-121 (2013).
- Ramírez, M. B., et al. GTP binding regulates cellular localization of Parkinson's disease-associated LRRK2. Human Molecular Genetics. , ddx161 (2017).
- Jung, H. J., Kwon, H. J. Target deconvolution of bioactive small molecules: the heart of chemical biology and drug discovery. Archives of Pharmacal Research. 38 (9), 1627-1641 (2015).
- Harding, M. W., Galat, A., Uehling, D. E., Schreiber, S. L. A receptor for the immunosuppressant FK506 is a cis-trans peptidyl-prolyl isomerase. Nature. 341 (6244), 758-760 (1989).
- Lomenick, B., et al. Target identification using drug affinity responsive target stability (DARTS). Proceedings of the National Academy of Sciences of the United States of America. 106 (51), 21984-21989 (2009).
- Manabe, Y., Mukai, M., Ito, S., Kato, N., Ueda, M. FLAG tagging by CuAAC and nanogram-scale purification of the target protein for a bioactive metabolite involved in circadian rhythmic leaf movement in Leguminosae. Chemical Communications. 46 (3), 469-471 (2010).
- Pantoliano, M. W., et al. High-density miniaturized thermal shift assays as a general strategy for drug discovery. Journal of Biomolecular Screening. 6 (6), 429-440 (2001).
- Li, X., Snyder, M. Analyzing In vivo Metabolite-Protein Interactions by Large-Scale Systematic Analyses. Current Protocols in Chemical Biology. , 181-196 (2010).
- Maeda, K., Poletto, M., Chiapparino, A., Gavin, A. -. C. A generic protocol for the purification and characterization of water-soluble complexes of affinity-tagged proteins and lipids. Nature Protocols. 9 (9), 2256-2266 (2014).
- Li, X., Gianoulis, T. A., Yip, K. Y., Gerstein, M., Snyder, M. Extensive in vivo metabolite-protein interactions revealed by large-scale systematic analyses. Cell. 143 (4), 639-650 (2010).
- Veyel, D., et al. System-wide detection of protein-small molecule complexes suggests extensive metabolite regulation in plants. Scientific Reports. 7, (2017).
- Luzarowski, M., et al. Affinity purification with metabolomic and proteomic analysis unravels diverse roles of nucleoside diphosphate kinases. Journal of Experimental Botany. , (2017).
- Van Leene, J., et al. Targeted interactomics reveals a complex core cell cycle machinery in Arabidopsis thaliana. Molecular systems biology. 6 (1), 397 (2010).
- Giavalisco, P., et al. Elemental formula annotation of polar and lipophilic metabolites using 13C, 15N and 34S isotope labelling, in combination with high-resolution mass spectrometry. The Plant Journal. 68 (2), 364-376 (2011).
- Van Leene, J., et al. Isolation of transcription factor complexes from Arabidopsis cell suspension cultures by tandem affinity purification. Plant Transcription Factors: Methods and Protocols. , 195-218 (2011).
- Van Leene, J., et al. A tandem affinity purification-based technology platform to study the cell cycle interactome in Arabidopsis thaliana. Molecular & Cellular Proteomics. 6 (7), 1226-1238 (2007).
- Olsen, J. V., Ong, S. -. E., Mann, M. Trypsin cleaves exclusively C-terminal to arginine and lysine residues. Molecular & Cellular Proteomics. 3 (6), 608-614 (2004).
- Cox, J., Mann, M. MaxQuant enables high peptide identification rates, individualized p.p.b.-range mass accuracies and proteome-wide protein quantification. Nature Biotechnology. 26 (12), 1367-1372 (2008).
- Cox, J., et al. Andromeda: A peptide search engine integrated into the MaxQuant environment. Journal of Proteome Research. 10 (4), 1794-1805 (2011).
- Tyanova, S., Temu, T., Cox, J. The MaxQuant computational platform for mass spectrometry-based shotgun proteomics. Nature Protocols. 11 (12), 2301 (2016).
- Hooper, C. M., et al. SUBAcon: a consensus algorithm for unifying the subcellular localization data of the Arabidopsis proteome. Bioinformatics. 30 (23), 3356-3364 (2014).
- Katajamaa, M., Orešič, M. Data processing for mass spectrometry-based metabolomics. Journal of Chromatography A. 1158 (1-2), 318-328 (2007).
- Szklarczyk, D., et al. STITCH 5: augmenting protein-chemical interaction networks with tissue and affinity data. Nucleic Acids Research. 1277, (2015).
- Tanaka, K. -. i., et al. Dipeptidyl compounds ameliorate the serum-deprivation-induced reduction in cell viability via the neurotrophin-activating effect in SH-SY5Y cells. Neurological Research. 34 (6), 619-622 (2012).
- Smith, C. A., et al. METLIN: A metabolite mass spectral database. Therapeutic Drug Monitoring. 27, 747-751 (2005).
- Bhattacharyya, S., et al. Transient protein-protein interactions perturb E. coli metabolome and cause gene dosage toxicity. Elife. 5, (2016).
- Rigaut, G., et al. A generic protein purification method for protein complex characterization and proteome exploration. Nature Biotechnology. 17 (10), 1030-1032 (1999).
转载和许可
请求许可使用此 JoVE 文章的文本或图形
请求许可探索更多文章
This article has been published
Video Coming Soon
版权所属 © 2025 MyJoVE 公司版权所有,本公司不涉及任何医疗业务和医疗服务。