
Method Article
2 1: 원스텝 단백질 단백질과 단백질 대사 산물 단지의 병렬 분석에 대 한 친 화력 정화
요약
단백질 대사 산물 및 단백질-단백질 상호 작용은 모든 세포 기능에 중요 한. 여기, 우리는 선택의 이러한 상호 작용 단백질으로의 병렬 분석을 허용 하는 프로토콜을 설명 합니다. 우리의 프로토콜 식물 세포 배양에 대 한 최적화 되었습니다 및 질량 분석 기반 단백질 및 대사 산물 검출 친 화력 정화를 결합 했다.
초록
세포질 과정은 단백질, 대사, 핵 산 등과 같은 생체 분자 간의 상호 작용에 의해 통제 된다. 단백질 단백질 상호 작용 (PPI)의 조사는 없는 참신, 실험적인 접근 생 단백질 대사 산물 상호 작용 (PMI)을 특성화 목표로 오히려 최근 개발을 구성 합니다. 여기, 우리는 PPI와 미끼 라고 선택의 단백질의 PMI의 동시 분석을 허용 하는 프로토콜을 제시. 우리의 프로토콜은 최적화 된 애기 셀 문화 및 친 화력 정화 (AP)를 결합 한 질량 분석 (MS)와 함께-기반 단백질 및 대사 산물 검출. 즉, 미끼 단백질 선호도 태그 융합을 표현 하는 유전자 변형 애기 라인 기본 세포 추출을 lysed 처음은. 방지 태그 항 체는 풀 미끼 단백질의 단백질 및 대사 산물 파트너 다운 하는 데 사용 됩니다. 친 화력 정화 단지 1 단계 메 틸 tert를 사용 하 여 추출 됩니다-부 틸 에테르 (티비) / 메탄올/물 방법. 대사는 극 지 또는 소수 성 위상, 분리 하는 동안 단백질 펠 릿에서 찾을 수 있습니다. 대사 산물과 단백질 다음 울트라 고성능 액체 크로마토그래피-질량 분석 (UPLC MS 또는 UPLC-MS/MS)에 의해 분석 된다. 빈 벡터 (EV) 제어 라인 가양성을 제외 하는 데 사용 됩니다. 우리의 프로토콜의 주요 장점은 근처 생리 적 조건 (세포 lysate)에서 동시에 대상 단백질의 단백질 및 대사 산물 파트너의 식별 수 있습니다. 제시 방법 간단, 신속, 그리고 식물 세포 배양 이외의 생물 학적 시스템에 쉽게 적용할 수 있습니다.
서문
방법을 설명 여기 근처-vivo에서 세포 lysate 조건에서 선택의 단백질의 대사 산물과 단백질 파트너의 식별에 목표. 그것은 많은 더 많은 대사 산물 오늘 특징 보다는 중요 한 규제 기능1는 추측 하고있다. 대사 산물은 생물학 스위치, 활동, 기능, 및 그들의 수용 체 단백질의2,,34의 지역화 변경으로 작동할 수 있다. 지난 10 년간에서 근처-vivo 조건, 또는 vivo에서 PMI의 식별 사용 여러 가지 획기적인 방법이 개발된5되었습니다. 사용할 수 있는 접근은 두 그룹으로 분리 수 있습니다. 첫 번째 그룹에는 소설 단백질 파트너를 트랩 하려면 알려진 대사 산물 미끼와 함께 시작 하는 기술 구성 되어 있습니다. 방법은 선호도 크로마토그래피6, 마약 선호도 응답 대상-안정성 분석 결과7, 항 암 치료-proteomics8및9프로 파일링 열 프로테옴 포함 됩니다. 알려진된 단백질 작은 분자 ligands10,11을 식별 하기 위해 시작 하는 단일 메서드 두 번째 그룹에 의하여 이루어져 있다.
MS 기반 lipidomics 함께 AP Saccharomyces cerevisiae12. 단백질 지질 단지 분석 하는 데 사용 했다 시작 지점으로 저자 표현 21 효소 ergosterol 생 합성에 관여 하는 효 모 종자를 사용 하 고 103 kinases 융합 협동 친 화력 정화 (꼭지) 태그. 효소의 70% 및 20%는 kinases의 복잡 한 단백질 지질 상호 작용 네트워크로 빛을 발산 하는 다른 소수 성 ligands를 바인딩할 발견 됐다.
이전에, 마찬가지로 지질, 극 지와 반 극 지 화합물 또한 유지 세포 lysates13에서 고립 된 단백질 복합물에 바인딩된 수 설명 합니다. 이러한 결과에 따라, 우리는 AP를 최적화 하기로 메서드 출판 이전 식물 세포 및 친수성 화합물1410,11 . 이 목적으로, 밴 Leene 외. 2010, PPI 연구15공장에서 성공적으로 사용 하 여 설명 탭 벡터 사용 했습니다. 유전자 변형 라인을 얻는 데 필요한 시간을 단축, 우리 애기 세포 배양에 결정 했다. 우리 고용 원스텝 메 틸 tert-부 틸 에테르, (티비) / 메탄올/물 추출 방법, 단일에서 단백질 (펠 릿), 지질 (유기 단계), 그리고 친수성 대사 (수성 단계)16 의 특성을 허용 친 화력 정화 실험입니다. EV 컨트롤 라인 가양성, 혼자 태그에 바인딩 단백질 예 제외에 도입 되었다. 우리는 3 (5) 태그 개념의 증거로 서 nucleoside diphosphate kinases 애기 게놈 (NDPK1-NDPK3)에 존재. NDPK1 티 S상호 작용 설명 수 다른 결과, 중-전이 효소와 티. 따라서 우리는 NDPK1 glutathionylation14받게 됩니다 증명할 수 있습니다.
정리해 보면, 제시 프로토콜 단백질-단백질 및 단백질 작은 분자 상호 작용 네트워크 특성화를 위한 중요 한 도구 이며 기존 방법에 비해 주요 사전 구성.
프로토콜
선의 유전자 변형 애기 셀 문화, 복제, 변환, 선택, 및 성장 조건을 포함 한 준비17에서 찾을 수 있습니다. 참고 EV 컨트롤 라인 판정에 대 한 해결 하기 위해 권장 됩니다. 실험, 이전 서쪽 오 점 분석, 예를 들면 협동 선호도 태그의 G-단백질 부분에 대 한 IgG 항 체를 사용 하 여 미끼 단백질의 overexpression를 확인 합니다. 그것은 식물 세포 문화 소재로 성장 매체를 분리 해야 합니다.
1. 실험 전에 식물 셀 재료 준비 하기
- PSB L A. thaliana 셀 문화 선 관심18의 단백질 overexpressing 성장.
- MSMO 매체, 4.43 g/L MSMO 혼합 30 g/L 자당을 포함 준비. 1 m 코 5.7 버퍼의 pH 및 고압 솔루션을 조정 합니다. 실험 전에 0.5 mg/L α-naphthaleneacetic 산, 0.05 mg/L kinetin와 50 μ g/mL 대 매체를 보충 한다.
- 부드러운 동요 (130 rpm)와 궤도 플랫폼 통에 100 mL 플라스 크에 MSMO 매체의 50 mL에 변형 된 식물 세포 배양을 배양. 20 ° C와 빛의 강도에 문화 방에 셀 성장 80 μmol m-2의 -1와 동일 합니다.
- 신선한 미디어 7 일 마다 그들에 게 1시 10분 희석으로 subculture 셀.
- 진공 펌프, 나일론 메쉬를 사용 하 여 필터와 결합 하 여 유리 깔때기를 사용 하 여 로그 성장 단계에서 세포를 수집 합니다. 알루미늄 호 일에는 침투를 포장 하 고 액체 질소에서 동결.
주의: 액체 질소는 매우 추운 기억. 잘못 된 처리는 화상을 일으킬 수 있습니다. 적절 한 개인 보호 장비를 착용, 장갑, 보호 안경, 그리고 실험실 코트 절연 열 등.
2. 프로토콜 탭
참고: 다음 단계는 마에 다 외. 201411 및 반 Leene 외. 201117에서 적응.
- 수확 및 냉동 공장 셀 문화 자료 정밀한 분말을 얻기 위해 믹서 밀 (2 분 20 Hz에서) 또는 박격포 및 방 앗 공이 사용 하 여 균질 Aliquot 3 g 샘플 당 (전체 단백질의 약 90 밀리 그램에 해당) 지상 물자의. 이 단계 동안 액체 질소 미리 냉장된 장비를 사용 하 여 샘플의 해빙 하지 마십시오.
참고: 저장소 지상 식물 소재 AP 프로시저의 시작 부분에 –80 ° C에서 50 mL 튜브에. - 얼음 처럼 차가운 세포의 용 해 버퍼의 3 mL와 함께 액체 질소 precooled 박격포에 샘플을 triturate (0.025 M Tris-HCl pH 7.5; 0.5 M NaCl, 1.5 m m MgCl2, 0.5 m m DTT; 1mm NaF; 1 m m 나3보4; 100 희석된 상업 프로 테아 제 억제 물 칵테일 x, 1 m m PMSF) 소재 녹은 때까지 일단 샘플 녹아서, 즉시 다음 단계를 진행 합니다.
참고: 세포의 용 해 버퍼 신선한 준비. 이 단계에서 빈 샘플을 소개 합니다. 세제는 그들은 MS 검색에서 문제를 발생할 수 있습니다 권장 하지 않습니다. - 세포질 파편을 제거 하려면 나눌 자료 2 mL microcentrifuge 튜브와 4 ° c.에서 10 분 동안 20,817 x g에서 원심 분리기 분명 15 mL 원뿔 원심 분리기 튜브에서 lysate의 3 mL를 수집 합니다.
- 원심 분리 단계 IgG Sepharose 구슬 equilibrate. Aliquot 100 µ L 샘플 당 구슬의 세포의 용 해 버퍼의 1 mL와 함께 그들을 씻어. Resuspend 구슬 및 플래시 스핀에 소용돌이입니다. 세포의 용 해 버퍼를 삭제 하 고 두 번 단계를 반복 합니다. 세포의 용 해 버퍼의 400 µ L에 구슬 resuspend.
- 구슬 lysate 수집 된 식물을 추가 하 고 1 h 4 ° c.에 대 한 회전 바퀴에 혼합물을 품 어
- 전송 혼합물에 주사기 Luer 잠금 캡 스핀 열 필터 기 공 크기 35 µ m. 통해 lysate 통과 하 적용 압력을 통해 결합. 연결 된 단지와 구슬 lysate를 통해 갈 것입니다 하는 동안 필터에 유지 됩니다.
참고: 필요에 따라 진공 매니폴드 시스템을 사용 합니다. 확인을 구슬 해 부드러운 압력을 적용 합니다. - 버퍼를 세척의 10 mL와 함께 처음에 비즈를 세척 (0.025 M Tris-HCl pH 7.5; 0.5 M NaCl), 그리고 차입 버퍼의 1 mL에 다음 (10 mM Tris-HCl pH 7.5; 150 mM NaCl, 0.5 m m EDTA, 희석 E64 및 1 밀리미터 PMSF x 1000). 열 또는 진공 매니폴드 시스템에 부착 된 주사기를 사용 하 여 세척을 수행 합니다.
참고: 때 진공 매니폴드 시스템을 사용 하 여 확인을 구슬 해 부드러운 압력을 적용 합니다. - 400와 구슬을 품 어 차입 버퍼를 포함 하는 담배의 향상된 된 버전의 U 50 µ L 엣지 바이러스 (AcTEV) 효소. 테이블 통 1000 rpm에서 16 ° c.에 30 분 사용
참고: 플러그를 사용 하 여 추가 차입 버퍼 하단에 열을 기억 해요. - 열에는 효소의 추가 부분 (50 U)를 추가 하 고, 위에서 설명한 조건, 동일에서 다음 30 분 동안 혼합물을 품 어.
- 원심 (1 분, 20,817 x g) 또는 진공 매니폴드 2 mL microcentrifuge 튜브에서 eluate를 수집 합니다. 나머지 단지 제거, 차입 버퍼의 200 µ L를 사용 하 여 추가 차입 단계를 소개 합니다.
참고: 저장 – 20 ° C 또는 –80 ° C, 샘플 또는 즉시 단백질 및 대사 산물 추출 단계와 진행. 얼음에 냉동된 샘플 녹여
3. 서쪽 오 점 분석
- 미끼의 존재를 확인 수집된 eluate에 단백질 SDS-페이지 및 서쪽 오 점 분석을 수행할 단백질-대사 산물을 포함 하 eluate의 10 µ L을 사용 합니다. 관심사의 단백질을 식별 하려면 마우스 streptavidin 의무적인 단백질 (1: 200), 밴 Leene 외. 201117에 설명 된 대로 TEV protease 분열 후 남은 꼭지 태그의 부분에 대 한 1 차 항 체를 사용 합니다. 다음, 보조 염소 반대로 마우스 항 체 HRP와 결합 하 여 사용 합니다.
4. 대사 산물 및 단백질 추출
참고:이 프로토콜은 Giavalisco 외. 201116에서 적응.
참고:이 단계 이후부터 UPLC-MS-학년 솔루션 사용.
- 추가 1 mL의 메 틸 tert-부 틸 에테르 (티비) / 메탄올/수집된 eluate 용 (3:1:1) 매 물 및 반전에 의해 샘플을 혼합. 용 매 추출 단계 전에 – 20 ° C에 냉각은 다는 것을 확인 하십시오.
주의: 티비와 메탄올은 유해 물질입니다. 증기 두건에서 추출 단계를 수행 하 고 적절 한 개인 보호 장비, 예를 들어, 장갑 착용. - 각 샘플을 0.4 mL 메탄올: 물 1:3 솔루션을 추가 하 고 반전 하 여 샘플의 내용을 혼합.
참고: 메탄올: 물 솔루션 혼합물의 보완 단계 구분에서 결과. 지질을 포함 하는 상위 단계, 낮은 단계 극 지와 반 극 지 대사 산물을 포함 하 고 단백질 펠 릿에서 찾을 수 있습니다. - 단계를 구분, 원심, 실내 온도에 2 분 동안 20,817 x g에서 샘플 다음 지질 측정 (하지에이 프로토콜)에 대 한 위 단계를 수집 1 mL 볼륨 용량 수동 액체 처리 피 펫을 사용 하 여.
- 반전 하 여 메탄올과 혼합의 0.2 mL를 추가 합니다.
- RT, 2 분에 대 한 20,817 x g에서 샘플 원심 다음 대사 산물 측정 (극 지와 반 극 지 화합물)에 대 한 극 지 단계를 수집 합니다. 단백질 펠 렛을 방해 하지 않도록, 튜브의 바닥에서 액체 위상의 약 50 µ L를 둡니다.
- 대사 산물 측정을 위한 건조 수집 된 샘플은 원심 증발 기에서 하룻밤. -30-60 분 후 증발에서 샘플을 제거 하 여 단백질 pellet을 건조 하지 마십시오.
참고: 저장 – 20 ° C 또는 –80 ° C, 샘플 또는 즉시 LC-MS/MS 분석을 위한 단백질의 준비와 진행.
5. Proteomic 분석에 대 한 샘플 준비
참고:이 단계는 올 슨 외. 에서 적응 200419 와 트립 신/리스-C 혼합의 기술 설명서 ( 재료의 표참조).
- 샘플의 효소 소화를 수행 합니다.
주의: 용 제 효소 소화 하는 동안 사용 하 고 유해는 샘플의 담. 증기 두건에서 작업과 적절 한 개인 보호 장비, 예를 들어, 장갑 착용.- 갓된 변성 버퍼 (40 m m 염화 중 탄산염 2 M thiourea/6 M 요소, pH 8 포함)의 30 µ L 펠 릿 단백질 분해. 더 나은 단백질 용 해도 위해 15 분 쥡니다 단계를 수행 합니다. 펠 릿 녹이 고 때까지 단계를 반복 합니다.
- 샘플에서 20,817 x g 4 ° C에서 10 분 원심 다음 새로운 microcentrifuge 튜브에는 상쾌한을 전송 합니다.
- Bradford 단백질 분석 실험을 사용 하 여 단백질 농도 결정 합니다.
- 에 대 한 추가 분석, aliquot 단백질의 100 µ g에 해당 볼륨 하 고 변성 버퍼 46 µ L까지 샘플을 입력 합니다.
- 갓된 감소 버퍼 (50 m m H2O에에서 녹아 있는 DTT)의 샘플 2 µ L을 추가 하 고 실 온에서 30 분 동안 품 어.
- 갓된 알 버퍼 (150 mM iodoacetamide 40 mM 염화 중 탄산염 버퍼에 녹아 있는)의 2 µ L로 샘플을 취급 하 고 실 온에서 20 분 동안 어둠 속에서 혼합물을 품 어.
- 40 m m 염화 중 탄산염 버퍼의 30 µ L로 샘플을 희석 하 고 LysC/트립 신 혼합의 20 µ L를 추가 합니다.
- 37 ° C에서 4 h 인큐베이션, 후 40 m m 염화 중 탄산염 버퍼의 300 µ L로 샘플을 희석.
- 37 ° c.에 야간 보육을 계속
- 10 %trifluoroacetic 산 (TFA) pH < 2를 약 20 µ L로 샘플을 시 어 지 다. 샘플 pH 산도 스트립을 사용 하 여 확인 하십시오.
참고: – 20 ° C에서 샘플을 저장 하거나 다음 단계를 진행 합니다.
- 소화 단백질 desalt
참고: 선호, 진공 매니폴드 시스템을 사용 합니다. -열 건조 하지 마십시오.- C18 SPE 열 린스 ( 재료의 표참조) 100%의 1 mL로 MeOH 다음 1 mL의 0.1%를 포함 하는 80% 이기 (ACN)와 함께 TFA 희석 물에. 사용, 여기 및 추가 단백질 담 단계, 프로세스 속도를 하는 진공 매니폴드 시스템. -열 건조 하지 마십시오.
- 0.1 %TFA 물에 희석의 1 mL로 두 번 세척 하 여 열을 equilibrate.
- 열에 샘플을 로드 합니다. 0.1 %TFA 전송 솔루션 열에 추가 200 µ L로 린스 튜브. 열을 통해 솔루션을 실행 합니다.
- 0.1%의 1 mL로 두 번 열 세척 TFA.
- 60%의 800 µ L 열에서 desalted 펩 티 드 elute ACN, 0.1 %TFA 솔루션. -단백질 분수의 30-60 분 후 증발에서 샘플을 제거 하 여 건조 방지, 원심 증발 기에서 수집 된 분수를 건조.
참고: 샘플 – 20 ° C 또는 –80 ° C에 저장 하거나 즉시 다음 단계를 진행 합니다. 이후 단계에서 얼음 샘플을 유지 합니다.
6. 측정 UPLC-MS/MS를 사용 하 여 단백질 샘플 준비.
참고: proteomic 및 metabolomic 측정, 이전 (0.2 µ m 기 공 크기)를 필터링 하 고 진공 펌프를 사용 하 여 1 시간에 대 한 모든 버퍼를 드.
- 건조 resuspend 펩 티 드 펠 릿 버퍼 C (3 %v / v ACN, 0.1 %v / v 포 름 산)를 사용 하 여 50 µ L에 2 mL microcentrifuge 튜브에 저장 된 200 µ L 볼륨 용량의 피 펫 수동 액체 처리. 35 kHz 초음파 주파수 초음파 목욕에서 15 분 샘플을 sonicate.
주의: ACN과 개미의 산 성 유해 물질입니다. 증기 두건에서 작업과 적절 한 개인 보호 장비, 예를 들어, 장갑 착용. - 샘플에서 20,817 x g 4 ° C에서 10 분 원심 다음 유리 유리병에는 상쾌한의 20 µ L를 전송 합니다.
- C18 반전 단계 열을 사용 하 여 별도 소화 펩 티 드 액체 크로마토그래피에 연결 하 고 질량 분석기를 사용 하 여 질량 스펙트럼을 취득.
- 3 µ L 300-nl/min 흐름 속도 사용 하 여 샘플의 열에 별도. 모바일 단계에 대 한 C와 D (63 %v / v ACN, 0.1 %v / v 포 름 산), 버퍼를 사용 하 여 그라데이션 3%에서 램프를 형성 15% ACN ACN 20 분 이상 30%를 다음 다음 10 분 이상 = 뉴스.
참고: – 20 ° C 또는 몇 개월 –80 ° C에서 샘플의 나머지 부분을 저장 합니다. Proteomic 측정 전에 얼음 샘플을 refreeze. - 60%를 사용 하 여 10 분 동안 오염 물질을 씻어 ACN 다음 샘플의 측정 전에 버퍼 C의 5 µ L 열 equilibrate 고.
- 70000 해상도 설정, 3e6 이온의 AGC 대상, 100 ms의 최대 주입 시간 데이터 종속 MS/MS 방법 및 m/z 300에서 1600까지를 사용 하 여 질량 스펙트럼을 얻을. 최대 17500의 해상도, 1 e5의 AGC 대상, 최대 주입 시간 100 ms의 언더 비율 20%의 1.6 m/z 및 m/z 200에서 2000에 이르기까지의 격리 창에서 15 MS/MS 검색의 취득. 에이펙스 트리거 (6-20 s) 15 s, 및 제외 요금 1과 > 5 설정된 동적 제외 가능
- 3 µ L 300-nl/min 흐름 속도 사용 하 여 샘플의 열에 별도. 모바일 단계에 대 한 C와 D (63 %v / v ACN, 0.1 %v / v 포 름 산), 버퍼를 사용 하 여 그라데이션 3%에서 램프를 형성 15% ACN ACN 20 분 이상 30%를 다음 다음 10 분 이상 = 뉴스.
7. Proteomic 데이터의 처리
- Http://www.uniprot.org/에서 최신 애기 thaliana 프로테옴 데이터베이스를 다운로드 하 고 오염 물질 데이터베이스 포함. LC-MS에서 얻은 원시 데이터 분석으로 기본 설정을 사용 하 여 통합 안드로메다 펩 티 드 검색 엔진 MaxQuant 사용 LFQ 정규화20,,2122사용 하 여 실행. 테이블 s 1에서 사용 하는 매개 변수에 대 한 자세한 정보를 찾을.
- "단백질 groups.txt" 출력 파일을 엽니다. 추가 분석을 위해 두 개 이상의 독특한 펩 티 드를 동일시 하는 단백질 그룹에 대 한 필터링. 잠재적인 오염 물질 및 A. thaliana 단백질 (Fasta 헤더 열에서 ARATH) 데이터베이스에 대 한 필터 MaxQuant에 의해 정의 된 단백질 그룹을 제거 합니다.
- 샘플 사이의 단백질 농축의 중요성을 테스트 하려면 정규화 LFQ 농도 사용 하 고 여러 비교 수정 (예: Benjamini & Hochberg 틀린 발견 비율 (루즈벨트) 다음 홀, 양측 스튜던트 t-검정을 수행 수정 또는 Bonferroni 보정)입니다.
- EV 제어 및 NDPK1에 대 한 얻은 LFQ 농도 비교 하 여 p-값을 계산 합니다. 모든 알 수 없는 값을 필터링. P-값을 오름차순으로 정렬 하 고 R 스크립트 또는 온라인 계산기 (예: https://www.sdmproject.com/utilities/?show=FDR)를 사용 하 여 계산 루즈벨트 교정. 0.1 아래 루즈벨트 값에 대 한 필터입니다.
참고: 데이터 분석의 형태를 연구에 대 한 적합 한 고려 하십시오. 양적 연구 (샘플 사이 단백질 농축의 분석) "LFQ 강도" 값을 사용 하는 반면 질적 연구 (특정 단백질의 존재 유무)에 대 한 "강도" 값을 선택 하십시오. - NDPK1 EV 컨트롤에 비교에서 더 풍부한 단백질 그룹에 대 한 필터입니다. 수바 데이터베이스23 을 사용 하 여 잠재적인 단백질 파트너의 지역화가 결정 하 고 NDPK1와 공동 화 된 단백질에 대 한 수정.
- EV 제어 및 NDPK1에 대 한 얻은 LFQ 농도 비교 하 여 p-값을 계산 합니다. 모든 알 수 없는 값을 필터링. P-값을 오름차순으로 정렬 하 고 R 스크립트 또는 온라인 계산기 (예: https://www.sdmproject.com/utilities/?show=FDR)를 사용 하 여 계산 루즈벨트 교정. 0.1 아래 루즈벨트 값에 대 한 필터입니다.
8. 폴라 단계 UPLC-MS를 사용 하 여 포함 된 샘플의 측량.
- Resuspend 단계 4.5에서 200 µ L의 물에서 극 단계 건조 하 고 5 분 동안 샘플을 sonicate.
- 샘플에서 20,817 x g 4 ° C에서 10 분 원심 다음 유리 유리병에는 상쾌한을 전송 합니다.
참고: – 20 ° C 또는 몇 개월에 대 한 –80 ° C에서 샘플의 나머지 부분을 저장 합니다. Metabolomic 측정 전에 얼음 샘플을 refreeze. - UPLC C18 반전 단계 열 결합을 사용 하 여 분리 단계를 수행 하 고 MS와 질량 스펙트럼을 취득.
- 각 이온화 모드 (긍정적 및 부정적) 주입 당 샘플의 열 2 µ L에 로드 하 고 400 µ L/min 흐름 속도 사용 하 여 분수를 분리. 대사 산물 측정을 위한 필요한 그라디언트를 만들려면 다음과 같이 준비 모바일 단계 솔루션: 버퍼는 (0.1% 개미 산 H2O)와 B 버퍼 (= 0.1% 포 름 산).
- 400 µ L/min 및 다음 그라데이션에 별도 대사 산물: 1 분 99% 버퍼, 버퍼의 버퍼, 버퍼 A 버퍼 A, 30%에서 15 분 선형 그라데이션의 30% ~ 60%에서 13 분 선형 그라데이션의 60%를 A의 99%에서 11 분 선형 그라데이션의 버퍼 A 1%를 버퍼 A의 16 분의 99% 농도와 십 분에 17 분, 버퍼 A의 1%에서 A. 다시 equilibrate 버퍼의 99%를 사용 하 여 선형 그라데이션에서에서 시작 다음 샘플의 측정 전에 A 버퍼 때까지 1% 농도 개최.
- 대량 범위 질량 스펙트럼을 취득 100과 1500 사이의 25000 m/z 해상도 설정 및 로딩 시간 1e6100 양 AGC 설정 대상 제한, 모 세관 전압 3kV 칼 집과 가스 흐름과 60 및 20의 보조 가스 값 각각. 모 세관 온도 250 ° C와 스키 머 전압 25V 설정 합니다.
9. 대사체학 데이터의 처리
- 프로세스 chromatograms 이온화 모드 모두에서 인수를 수집. 소프트웨어를 사용 하 여 전 비 (m/z), 보존 시간 (RT)와 연결 된 봉우리, 예를 들면, 상용 소프트웨어의 강도를 대량 추출 ( 재료의 표참조) 또는 대체24.
- .Exe 파일을 두 번 클릭 하 여 프로세싱 소프트웨어를 시작
- 새 워크플로 만들기, 활동 "파일에서 로드"에 대 한 검색 그리고 빈 워크플로 공간으로 "끌어서 놓기"이이 활동 이동 합니다. 활동을 마우스 오른쪽 버튼으로 누르고 활동의 설정을 엽니다.
- 포함 하는 "일반" 설정 탭에서 "이름" 필드에 실험의 이름을 설정 하 고 "선택 파일 및 폴더"를 클릭 한 원시 chromatograms 표시.
- 포함 하는 "고급" 설정 탭에서 "프로필 데이터 구분" 0 강도를 설정 합니다. "적용" 및 "확인"을 클릭 합니다.
- 에 대 한 검색 및 "데이터 청소" 활동을 추가. 활동을 마우스 오른쪽 버튼으로 누르고 활동의 설정을 엽니다.
- 포함 하는 "일반" 설정 탭에서 "중심 데이터"와 "MS/MS 데이터"를 표시 합니다. 선택 패널에서 "모든 것"을 선택 하 여 모든 MS/MS 데이터를 제거 합니다.
- 검색 하 고 추가 활동 "크로마 화학 잡음 빼기". 활동을 마우스 오른쪽 버튼으로 누르고 활동의 설정을 엽니다.
- 포함 하는 "일반" 설정 탭에서 "크로마 스무 딩"을 표시 하 고 "3" 검색 및 "이동 평균"에 "견적"의 수를 설정 합니다. 51 검색, 50%, 빼기 "방법" 및 750 강도 "임계값"을 "논집" 창 "RT"를 설정 합니다.
- 포함 하는 "고급" 설정 탭에서 "실시간 구조 제거"를 표시 하 고 5 검색 "최소 RT 길이"를 설정 합니다.
- 포함 하는 "고급" 설정 탭에서 "m/z 구조 제거"를 표시 하 고 3 포인트 "최소 m/z 길이"를 설정 합니다.
- 검색 하 고 추가 활동 "크로마 실시간 맞춤". 활동을 마우스 오른쪽 버튼으로 누르고 활동의 설정을 엽니다.
- 포함 하는 "일반" 설정 탭에서-0.5 분 "맞춤" "쌍 정렬 기본 트리에" 및 "실시간 검색 간격"을 설정 합니다.
- 포함 된 "고급" 탭에서 설정, 기본 매개 변수를 사용 합니다.
- 검색 하 고 "크로마" 그룹 활동의 활동 "피크 탐지"를 추가. 활동을 마우스 오른쪽 버튼으로 누르고 활동의 설정을 엽니다.
- 포함 하는 "일반" 설정 탭에서 0.09 분, 0.03 분, "최대 병합 거리" 5 점 및 "병합 전략" "센터"를 "최소 피크 크기"를 "변론 창"을 설정 합니다. "피크 RT 분할"에서 상자 50% "차이/피크 비율"를 설정합니다.
- 포함 하는 "고급" 설정 탭에서 5 점, 80% "구체화 임계값" 및 "일관성 임계값" 1로 "스무 딩 창"을 설정 합니다. 70%에서 설정 "강도 임계값"와 "강도 가중치"로 "센터 계산"를 설정 합니다.
- 검색 하 고 활동의 "크로마" 그룹에서 "동위 원소 클러스터링" 활동을 추가. 활동을 마우스 오른쪽 버튼으로 누르고 활동의 설정을 엽니다.
- 포함 하는 "일반" 설정 탭에서 "실시간 공차" 0.015 min을 "m/z 허용" 5 ppm을 설정 합니다.
- 탭에 포함 된 "봉투 피팅", 설정 "방법" "아니 모양 제한" 및 "이온화" "Protonation (대 한 긍정적인 모드) 및"Deprotonation"(부정적인 모드)에 대 한. 설정 "최소 및 최대 충전" 1, 4, 각각.
- 포함 된 "고급" 탭에서 설정, 기본 매개 변수를 사용 합니다.
- 검색 하 고 추가 활동 "단일 필터".
- 데이터 처리의 결과 내보내려면, 검색 하 고 "내보내기" 그룹 활동의 활동 "분석가"를 추가 합니다.
- 포함 하는 "일반" 설정 탭에서 "클러스터"와 "관찰" "표현 강도"으로 "유형"을 설정 합니다. "대상 사용자 지정"을 선택 하 고 내보내기 파일의 디렉터리를 지정 합니다.
- 포함 된 "고급" 탭에서 설정, 기본 매개 변수를 사용 합니다.
- 주석을 사내 기준 화합물 데이터베이스를 사용 하 여 대량 기능.
- 하나 또는 여러 개의 MS 학년 참조-화합물 UPLC 양 사용을 사용 하 여 참조 화합물 및 공동 관심사의 단백질을 순화 하는 대사 산물의 분석에 대 한 동일한 LC MS 방법 분석.
참고:이 연구를 위해 거의 300 dipeptides 집합 분석 되었고 참조-화합물 라이브러리 사용. - 분석을 사용 하 여 (참조 테이블의 재료) 소프트웨어와 관련 된 특정 m/z와 RT에 대 한 원시 크로마 파일 및 검색을 열려면 측정 참조 화합물 (사용자 설명서 참조).
참고: 보조 metabolites 이온화의 종류에 차이가 있습니다. 검사 일반의 존재는 이온 질량 M + 22.989218, M + 18.033823, M + 1.007276, M-1.007276와 같은 [M-H]에 대 한 검색 하 여 adducts [M + H], [M + NH4]와 [M + 나], 각각. - 사용 스프레드시트 열을 내보낸 "분석가" 파일 chromatograms 및 특정 이온에 대 한 검색의 처리 후 얻은 질량. 실시간 대량 기능 실험에 참조 화합물의 실시간 측정의 비교. M/z 0.005 다와 실시간에 대 일 분의 편차를 허용
- 하나 또는 여러 개의 MS 학년 참조-화합물 UPLC 양 사용을 사용 하 여 참조 화합물 및 공동 관심사의 단백질을 순화 하는 대사 산물의 분석에 대 한 동일한 LC MS 방법 분석.
- 대사 산물 농축 공동 샘플 (overexpressed 단백질 관심 대 EV 통제의 선) 사이 특정 단백질 정화의 의미를 테스트 하려면 여러 뒤 양측 쌍이 아닌 학생의 t-검정을 사용 하 여 피크 값 비교 비교 수정 (예: Benjamini 및 Hochberg 거짓 발견 속도 보정 또는 Bonferroni 보정).
결과
원래 연구에 3 개의 A. thaliana NDPK 유전자 PSB L 셀 서 스 펜 션 문화 제정 35S 발기인14 (그림 1)의 통제에 overexpressed 했다. 탠덤 선호도 태그 미끼 단백질의 카 또는 아미노-터미널 끝을 융합 했다. 친 화력 정화 단지 티비/메탄올/물 추출16를 받게 했다. 선호도 뽑아 단백질과 작은 분자는 MS (테이블 S2 및 S3)를 사용 하 여 확인 되었다.
판정에 대 한 수정, 빈 샘플 작은 분자 오염 물질 화학 제품 및 실험실 소모품에서 제외 사용 되었다. 또한, 대사 산물 선호도 태그 중 하나에 바인딩 또는 혼자 수 지 단백질은 고려 EV 컨트롤 라인을 사용 하 여. 참 긍정 양측 쌍이 아닌 학생의 t-검정, Benjamini & Hochberg 틀린 발견 비율을 검색 하 보정 대사 (테이블 S4) 및 단백질 (테이블 S5) 크게 NDPKs AP에 농축 식별에 적용 된 EV 컨트롤 라인에 비해 (N-와 C-말기 태그 NDPKs) 실험 (루즈벨트 < 0.1). Note는 이전 작품에서 우리 사용 부재/존재 조건을 단백질과 작은 분자 인터 윤곽을 그리 다.
대표 결과 NDPK1를 위한 dipeptides, 우리의 그룹에 작은 분자 레 귤 레이 터의 새로운 클래스에 대사 산물 데이터 초점 동안 받는다. Proteomic 분석 NDPK1의 26 상 상속 단백질 파트너를 공개 했다. 공동 NDPK1으로 같은 subcellular 구획에서 화 된 단백질에 대 한 추가 필터링 (cytosol) 목록 13 상 상속 단백질 인터 한정. 확인 된 단백질 가운데 티 S-전이 효소, 두 개의 신장 개시 요인, tubulin, 및 aconitate hydratase. Metabolomic 분석 4 dipeptides 발 레이, Ile Glu, 레이-한 일-NDPK1와는 특별히 공동 eluted Phe 밝혀 (그림 2). 참고 모든 4 개의 dipeptides 그들의 N-말단, 공유 바인딩 특이성 제안에 소수 성 잔류물을 공유 합니다.
알려진된 단백질 단백질과 단백질 대사 산물 단지 우리가 찾을 쿼리 13 단백질과 스티치 데이터베이스25 (그림 3)에 대 한 4 개의 dipeptides 식별. 여러 가지 관측을 만들 수 있다: (i) 이전 NDPK1에 대 한 보고는 없는 인터. (ii) APX1 ortholog 알데하이드 효소 가족 ALDH7B4, 번역 개시 인자 FBR12 다른 번역 개시 인자 유전자 AT2G40290에 의해 동안에 상호 작용을 보고 되었다. (iii) 확인 된 dipeptides 더 단백질 파트너를 보고 있다. 공동 eluted dipeptides 이전 모든 검색 된 식물 단백질에 관련 된 보도 했다. 그러나, 그들은 다른 유기 체에서 중요 한 역할 놀이: 레이-일, 예를 들면, 인간 세포 선26neurotrophin 활성화 효과가 있다. 참고 실험 시스템의 정확한 토폴로지 확인 허용 하지 않습니다. 예를 들어 한 dipeptide NDPK1와 직접 상호 작용할 수 있지만 잘 공동 순화 된 단백질의 어떤 관련이 있을 수 있습니다.
함께 찍은, 우리의 결과 보여준다 질량 분석, 함께 AP를 고용 설립된 절차, 단백질-단백질 및 단백질 작은 분자 인터의 식별을 용이 하 게 하 고 도움에 대 한 광범위 한 정보를 생성 합니다 위하여 대상 단백질의.

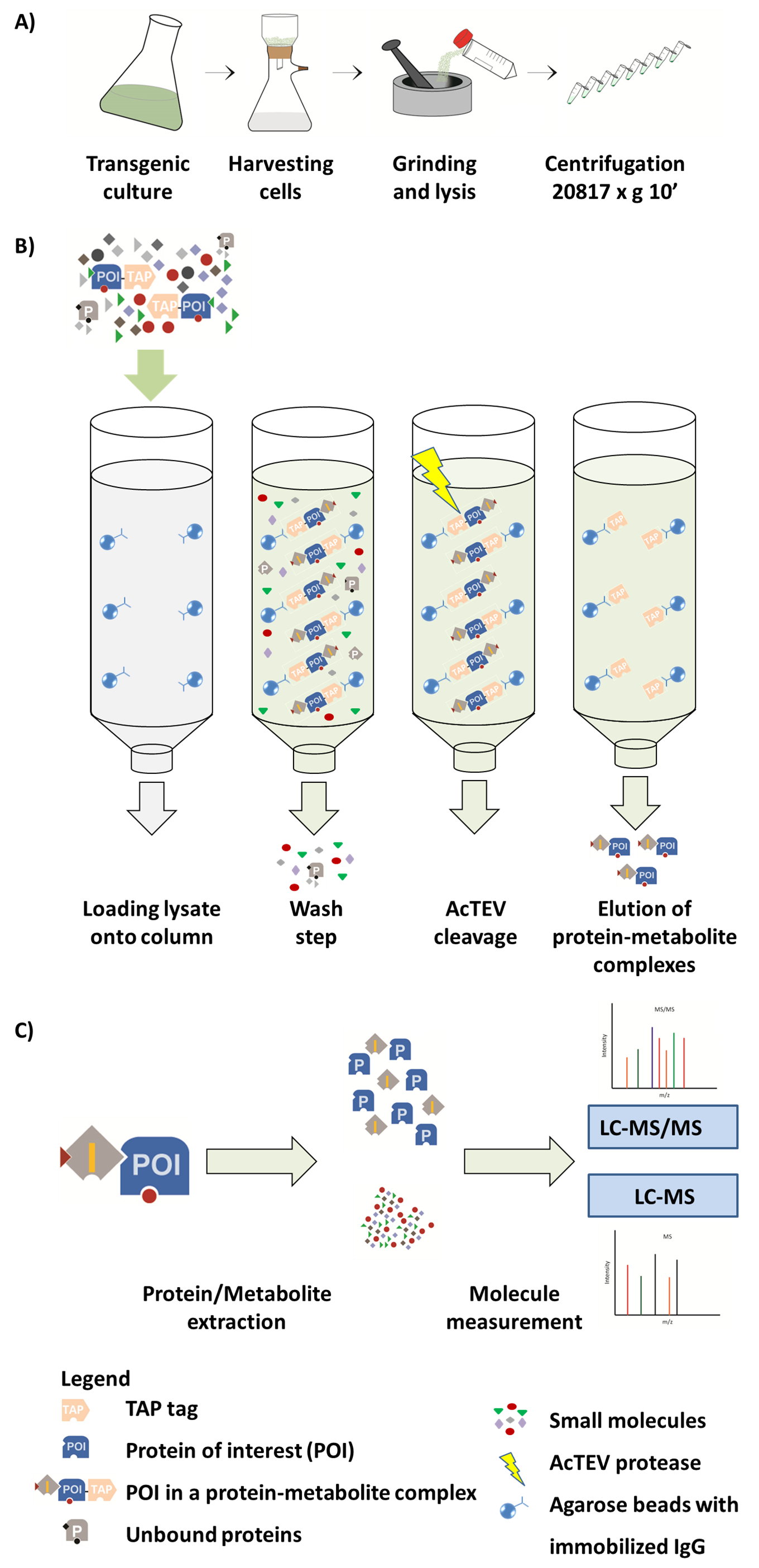
그림 1입니다. AP-MS 워크플로의 체계입니다. (A) 식물 세포 배양에서 네이티브 녹는 분수의 준비. (B) AP 절차의 다음 단계. 열에 샘플을 로드 한 후 관심 (POI) 탭 태그 융합 단백질 agarose 구슬에 움직일 IgG 항 체에 바인딩합니다. 열 세척 언바운드 단백질 및 대사 산물의 제거를 촉진 한다. AcTEV 분열을 수행한 후 포 단백질 대사 산물 단지는 eluted. (C) 단백질 및 대사 산물 분수 반 양적 MS 분석 뒤에 단지의 분리. 이 그림의 일부는 Luzarowski 외. 201714에서 재현 됩니다. 이 그림의 더 큰 버전을 보려면 여기를 클릭 하십시오.
{kind=link}

그림 2입니다. 특히 공동 NDPK1와 방출 dipeptides. 4 dipeptides 발 레이 (A), Ile Glu (B), 레이 일 (C), 및 일 페 (D) AP 실험에서 측정의 평균 농도 구성 했다. 모든 4 개의 dipeptides NDPK1 샘플 EV 제어에 비해 상당한 농축 표시 (별표 나타냅니다 루즈벨트 < 0.1). 오차 막대 대표 6 측정 표준 오차 (N-3 복제와 C 말기의 3 태그 단백질). 이 그림의 더 큰 버전을 보려면 여기를 클릭 하십시오.
{kind=link}

그림 3입니다. 공동으로 NDPK1, 방출 하는 모든 분자의 상호 작용 네트워크 이전만 고려 하는 스티치 데이터베이스에 대해 쿼리 실험 및 데이터베이스 증거 (신뢰 > 0.2). 높은 신뢰 상호 작용의 더 높은 기회를 나타냅니다 하 고 예금 된 데이터에 따라 계산 됩니다. 이 그림의 더 큰 버전을 보려면 여기를 클릭 하십시오.
{kind=link}
테이블 S1입니다. MaxQuant 출력 테이블 "parameters.txt". 테이블 식별 및 정량화, 뿐만 아니라 사용 하는 데이터베이스에 대 한 정보에 대 한 임계값이 포함 됩니다. 이 파일을 다운로드 하려면 여기를 클릭 하십시오.
테이블의 S2 MaxQuant에서 정보 출력 테이블 "proteinGroups.txt". 테이블에는 모든 확인 된 단백질 그룹, 농도, 및 고유 펩 티 드 및 점수 번호 등 추가 정보 목록이 포함 되어 있습니다. 이 파일을 다운로드 하려면 여기를 클릭 하십시오.
테이블의 S3 극 지 대사 산물의 분석을 포함 하는 출력 파일입니다. 테이블에 모든 확인 된 대량 기능 특정 m/z, RT 및 강도 특징의 목록이 포함 되어 있습니다. 이 파일을 다운로드 하려면 여기를 클릭 하십시오.
테이블 S4입니다. Dipeptides는 NDPK1, NDPK2 또는 NDPK3 미끼로 사용 되었다 AP 샘플에서 발견. Dipeptides 빈 샘플에는 목록에서 제외 했다. 두 개의 독립적인 라인 (태그에 어느 N-또는 C-말단) 각 NDPK는 3 중에서 실행 했다. 학생의 t-테스트 및 p의 수정 추가-Benjamini를 사용 하 여 값 및 Hochberg 메서드 NDPKs의 크게 농축된 끌어와서 파트너를 결정 하는 데 사용 되었다 (루즈벨트 < 0.1). 주어진 ΔRT 계산 됩니다 참조 화합물과 monoisotopic 질량 주어진 Metlin27에 관하여 Δppm. 이 파일을 다운로드 하려면 여기를 클릭 하십시오.
테이블 S5입니다. NDPK1와 공동 순화 하는 단백질. 3 중에서 2 개의 독립적인 라인 (태그에 어느 N-또는 C-말단) 각 NDPK 실행 했다. 학생의 t-테스트 및 p의 수정 추가-Benjamini를 사용 하 여 값 및 Hochberg 메서드 NDPKs의 크게 농축된 끌어와서 파트너를 결정 하는 데 사용 되었다 (루즈벨트 < 0.1). 이 파일을 다운로드 하려면 여기를 클릭 하십시오.
토론
제시 프로토콜 대상 단백질의 PP와 오후 단지의 병렬 식별 수 있습니다. 최종 결과를 복제에서 실험 8-12 주 만큼 적게 완료할 수 있습니다. 완전 한 AP 걸립니다 약 4-6 h 12 ~ 24 샘플 집합에 대 한 우리의 프로토콜 중간 처리량 분석을 위한 적합 한 렌더링.
프로토콜, 되 고 전반적으로 간단에 불구 하 고 중요 한 단계의 수가 있다. (i) 충분 한 양의 입력된 단백질 및 선호도 구슬 대사 산물 탐지의 동적 범위를 도달 하는 중요 한 이다. 효율적인 셀 세포 따라서 절차에서 중요 한 단계입니다. 불 쌍 한 단백질 수율 차선 세포의 용 해 버퍼/재료 비율 또는 자료의 부족 분쇄의 결과 될 수 있습니다. (ii) 해야 주의 사용된 시 약 MS-친화입니다. 강한 세제, 글리세롤, 또는 소금의 과도 한 양의 그들은 MS 검색을 방해 피해 야 한다. (iii) Agarose 구슬 해서는 안됩니다-건조 단계, 세척 하는 동안 그리고 진공 매니폴드를 사용 하는 경우 그것은 구슬 파괴 또는 복잡 한 안정성에 영향을 미칠에 그렇게 하지 않도록 느린 흐름 속도 적용 하는 것이 중요.
제시 프로토콜에 중요 한 몇 가지 가능한 수정이 있다: (i) 제정 CaMV35S 모터를 사용 하 여 미끼 단백질의 양을 최대화 하는 우리. Overexpression, 매우 유용한, 고 수 있습니다 세포 항상성28 에 심각한 영향을 미칠 생리 적으로 관련이 없는 상호 작용의 형성으로 이어질. 식 사용 하 여 네이티브 발기인 기능 손실 배경에서 가능한 진정한 생물학 인터 검색에 대 한 우수한 여겨진다 태그 단백질입니다. 일반적으로 식물 세포 배양에 표현 하는 단백질에 대 한 식물 배경 관련 인터를 식별 하는 데 필요한 증명할 수 있습니다. (ii) 작업할 때 막 단백질, 세포의 용 해 버퍼는 MS 호환 세제로 보완 될 필요가 있다. (iii) 두 번째 친 화력 정화 단계 소개 가양성 true 반응 비율을 개선 하 고 EV 컨트롤29필요 수 있습니다. 두 개의 독립적인 효소 분열 사이트 소설 탠덤 태그 모두 힘들고 시간이 소모 되는 마에 다 외. 201411, 추가 크기 배제 크로마토그래피 단계에 매력적인 대안을 선물 한다.
AP의 가장 심각한 결점은 잘못 된 반응의 높은 속도. 이유는 수많은. 제정 overexpression 이미 언급 했다. 순수 없는 상호 작용의 다른 소스 고립 된 세포를 사용 하지 않는 한 단백질과 다른 subcellular 구획에서 대사 산물의 혼합물을 포함 하는 전체 세포 lysates의 준비 이다. Subcellular 지 방화는 진정한 인터에 대 한 필터링을 사용 해야 합니다. 그럼에도 불구 하 고, 잘못 된 반응의 대부분 난다 바인딩 단백질과 agarose 수 지 사이에서 유래한 다. 그러나 두 번째 정화 단계의 소개, 위에서 설명한 문제에 최고의 솔루션을 제공 합니다, 그리고 시간과 처리량 비용 온다. 또한, 약한 상호 작용 프로토콜 확장으로 손실 될 수 있습니다. AP의 또 다른 경고는 포괄적인 정보 제공 하는 위하여 대상 단백질의,에 불구 하 고 baited 단백질의 직접 및 간접 목표 구별 불가능 이다. 타겟된 분자 접근 상호 작용을 확인 필요 합니다.
MS 기반 대사체학 함께 AP S. cerevisiae12. 단백질 복합물 공부 하는 사용 했다 이 작품은, 마찬가지로 지질, 극 지와 반 극 지 화합물 유지 세포 lysates에서 고립 된 단백질 복합물에 바인딩된 우리의 이전 관찰13 함께 제시 프로토콜에 대 한 개념적 기초를 제공 합니다. 우리의 프로토콜 3 독특한 포인트 특징 이다: (i) 반면에 누 룩을 작동12, AP 검색 뿐만 아니라 소수 하지만 또한 친수성 단백질 ligands에 적합 하다는 설명. (2) 3-에서-하나의 추출 프로토콜을 도입 하 여 미끼 단백질의 단백질 및 대사 산물 인터를 공부 하는 단일 AP은 사용할 수 있습니다. (iii) 우리는 식물 세포 프로토콜을 적응.
향후 두 개의 독립적인 효소 분열 사이트 소설 탠덤 태그를 만드는 방법에 집중할 것 이다. 우리 또한 낮은 풍부한 식물 호르몬과 같은 작은 분자에 프로토콜의 적합성을 탐험 하 고 싶습니다.
공개
저자는 공개 없다.
감사의 말
우리는 친절 하 게 그의 참여 프로젝트, 생산적인 토론과 좋은 감독에 대 한 교수 박사 로타어 Willmitzer를 인정 하 고 싶습니다. 우리는 proteomic MS 측정 도움 박사 다니엘 Veyel에 감사입니다. 우리 부인 Änne Michaelis LC MS 측정 우리 귀중 한 기술적인 도움을 제공 주셔서 감사 합니다. 또한, 우리는 닥터 모니카 Kosmacz와 박사 Ewelina Sokołowska Weronika Jasińska를 그들의 도움과 원래 원고에서 작업에 참여에 대 한 기술 지원을 감사 하 고 싶습니다.
자료
| Name | Company | Catalog Number | Comments |
| Murashige and Skoog Basal Salts with minimal organics | Sigma-Aldrich | M6899 | |
| 1-Naphthylacetic acid | Sigma-Aldrich | N1641 | |
| Kinetin solution | Sigma-Aldrich | K3253 | |
| Tris base | Sigma-Aldrich | 10708976001 | |
| NaCl | Sigma-Aldrich | S7653 | |
| MgCl2 | Carl Roth | 2189.1 | |
| EDTA | Sigma-Aldrich | 3609 | |
| NaF | Sigma-Aldrich | S6776 | |
| DTT | Sigma-Aldrich | D0632 | |
| PMSF | Sigma-Aldrich | P7626 | |
| E-64 protease inhibitor | Sigma-Aldrich | E3132 | |
| Protease Inhibitor Cocktail | Sigma-Aldrich | P9599 | |
| Na3VO4 | Sigma-Aldrich | S6508 | |
| AcTEV Protease | Thermo Fischer Scientific | 12575015 | |
| Rotiphorese Gel 30 (37,5:1) | Carl Roth | 3029.2 | |
| TEMED | Carl Roth | 2367.3 | |
| PageRuler Prestained Protein Ladder | Thermo Fischer Scientific | 26616 | |
| SBP Tag Antibody (SB19-C4) | Santa Cruz Biotechnology | sc-101595 | |
| Goat anti-mouse IgG-HRP | Santa Cruz Biotechnology | sc-2005 | |
| Bradford Reagent | Sigma-Aldrich | B6916 | |
| Trypsin/Lys-C Mix, Mass Spec Grade | Promega | V5071 | |
| Urea | Sigma-Aldrich | U5128 | |
| Thiourea | Sigma-Aldrich | T8656 | |
| Ammonium bicarbonate | Sigma-Aldrich | 9830 | |
| Iodoacetamide | Sigma-Aldrich | I1149 | |
| MTBE | Biosolve | 138906 | |
| Methanol | Biosolve | 136806 | |
| Water | Biosolve | 232106 | |
| Acetonitrile | Biosolve | 12006 | |
| Trifluoroacetic acid | Biosolve | 202341 | |
| Formic acid | Biosolve | 69141 | |
| Unimax 2010 Platform Shaker | Heidolph | 5421002000 | |
| Nylon Mesh (Wire diameter 34 µM, thickness 55 µM, open area 14%) | Prosepa | Custom order | |
| Glass Funnel, 47 mm, 300 ml | Restek | KT953751-0000 | |
| Filter Bottle Top 500 mL 0,2 µM Pes St | VWR International GmbH | 514-0340 | |
| Mixer Mill MM 400 | Retsch GmbH | 207450001 | |
| IgG Sepharose 6 Fast Flow | GE Healthcare Life Sciences | 17-0969-02 | |
| Mobicol ""Classic"" with 2 different screw caps without filters | MoBiTec GmbH | M1002 | |
| Filter (small) 35 µM pore size, for Mobicol M 1002, M1003, M1050 & M1053 | MoBiTec GmbH | M513515 | |
| Variable Speed Tube Rotator SB 3 | Carl Roth | Y550.1 | |
| Rotary dishes for rotators SB 3 | Carl Roth | Y555.1 | |
| Resprep 24-Port SPE Manifolds | Restek | 26080 | |
| Finisterre C18/17% SPE Columns 100mg / 1ml | Teknokroma | TR-F034000 | |
| Autosampler Vials | Klaus Trott Chromatographie-Zubehör | 40 11 01 740 | |
| Acclaim PepMap 100 C18 LC Column | Thermo Fischer Scientific | 164534 | |
| EASY-nLC 1000 Liquid Chromatograph | Thermo Fischer Scientific | LC120 | |
| Q Exactive Plus Hybrid Quadrupole-Orbitrap Mass Spectrometer | Thermo Fischer Scientific | IQLAAEGAAPFALGMBDK | |
| Acquity UPLC system | Waters | Custom order | |
| ACQUITY UPLC HSS C18 Column, 100A, 1.8 µM, 2.1 mM X 100 mM, 1/pkg | Waters | 186003533 | |
| High-power ultrasonic cleaning baths for aqueous cleaning solutions | Bandelin | RK 31 | |
| Genedata Expressionist | Genedata | NaN | |
| Xcalibur Software | Thermo Fischer Scientific | NaN | |
| MaxQuant | NaN | NaN |
참고문헌
- Li, X., Snyder, M. Metabolites as global regulators: A new view of protein regulation. Bioessays. 33 (7), 485-489 (2011).
- Jacob, F., Monod, J. Genetic regulatory mechanisms in the synthesis of proteins. Journal of Molecular Biology. 3 (3), 318-356 (1961).
- Schlattner, U., et al. Dual Function of Mitochondrial Nm23-H4 Protein in Phosphotransfer and Intermembrane Transfer a cardiolipin-dependent switch. Journal of Biological Chemistry. 288 (1), 111-121 (2013).
- Ramírez, M. B., et al. GTP binding regulates cellular localization of Parkinson's disease-associated LRRK2. Human Molecular Genetics. , ddx161 (2017).
- Jung, H. J., Kwon, H. J. Target deconvolution of bioactive small molecules: the heart of chemical biology and drug discovery. Archives of Pharmacal Research. 38 (9), 1627-1641 (2015).
- Harding, M. W., Galat, A., Uehling, D. E., Schreiber, S. L. A receptor for the immunosuppressant FK506 is a cis-trans peptidyl-prolyl isomerase. Nature. 341 (6244), 758-760 (1989).
- Lomenick, B., et al. Target identification using drug affinity responsive target stability (DARTS). Proceedings of the National Academy of Sciences of the United States of America. 106 (51), 21984-21989 (2009).
- Manabe, Y., Mukai, M., Ito, S., Kato, N., Ueda, M. FLAG tagging by CuAAC and nanogram-scale purification of the target protein for a bioactive metabolite involved in circadian rhythmic leaf movement in Leguminosae. Chemical Communications. 46 (3), 469-471 (2010).
- Pantoliano, M. W., et al. High-density miniaturized thermal shift assays as a general strategy for drug discovery. Journal of Biomolecular Screening. 6 (6), 429-440 (2001).
- Li, X., Snyder, M. Analyzing In vivo Metabolite-Protein Interactions by Large-Scale Systematic Analyses. Current Protocols in Chemical Biology. , 181-196 (2010).
- Maeda, K., Poletto, M., Chiapparino, A., Gavin, A. -. C. A generic protocol for the purification and characterization of water-soluble complexes of affinity-tagged proteins and lipids. Nature Protocols. 9 (9), 2256-2266 (2014).
- Li, X., Gianoulis, T. A., Yip, K. Y., Gerstein, M., Snyder, M. Extensive in vivo metabolite-protein interactions revealed by large-scale systematic analyses. Cell. 143 (4), 639-650 (2010).
- Veyel, D., et al. System-wide detection of protein-small molecule complexes suggests extensive metabolite regulation in plants. Scientific Reports. 7, (2017).
- Luzarowski, M., et al. Affinity purification with metabolomic and proteomic analysis unravels diverse roles of nucleoside diphosphate kinases. Journal of Experimental Botany. , (2017).
- Van Leene, J., et al. Targeted interactomics reveals a complex core cell cycle machinery in Arabidopsis thaliana. Molecular systems biology. 6 (1), 397 (2010).
- Giavalisco, P., et al. Elemental formula annotation of polar and lipophilic metabolites using 13C, 15N and 34S isotope labelling, in combination with high-resolution mass spectrometry. The Plant Journal. 68 (2), 364-376 (2011).
- Van Leene, J., et al. Isolation of transcription factor complexes from Arabidopsis cell suspension cultures by tandem affinity purification. Plant Transcription Factors: Methods and Protocols. , 195-218 (2011).
- Van Leene, J., et al. A tandem affinity purification-based technology platform to study the cell cycle interactome in Arabidopsis thaliana. Molecular & Cellular Proteomics. 6 (7), 1226-1238 (2007).
- Olsen, J. V., Ong, S. -. E., Mann, M. Trypsin cleaves exclusively C-terminal to arginine and lysine residues. Molecular & Cellular Proteomics. 3 (6), 608-614 (2004).
- Cox, J., Mann, M. MaxQuant enables high peptide identification rates, individualized p.p.b.-range mass accuracies and proteome-wide protein quantification. Nature Biotechnology. 26 (12), 1367-1372 (2008).
- Cox, J., et al. Andromeda: A peptide search engine integrated into the MaxQuant environment. Journal of Proteome Research. 10 (4), 1794-1805 (2011).
- Tyanova, S., Temu, T., Cox, J. The MaxQuant computational platform for mass spectrometry-based shotgun proteomics. Nature Protocols. 11 (12), 2301 (2016).
- Hooper, C. M., et al. SUBAcon: a consensus algorithm for unifying the subcellular localization data of the Arabidopsis proteome. Bioinformatics. 30 (23), 3356-3364 (2014).
- Katajamaa, M., Orešič, M. Data processing for mass spectrometry-based metabolomics. Journal of Chromatography A. 1158 (1-2), 318-328 (2007).
- Szklarczyk, D., et al. STITCH 5: augmenting protein-chemical interaction networks with tissue and affinity data. Nucleic Acids Research. 1277, (2015).
- Tanaka, K. -. i., et al. Dipeptidyl compounds ameliorate the serum-deprivation-induced reduction in cell viability via the neurotrophin-activating effect in SH-SY5Y cells. Neurological Research. 34 (6), 619-622 (2012).
- Smith, C. A., et al. METLIN: A metabolite mass spectral database. Therapeutic Drug Monitoring. 27, 747-751 (2005).
- Bhattacharyya, S., et al. Transient protein-protein interactions perturb E. coli metabolome and cause gene dosage toxicity. Elife. 5, (2016).
- Rigaut, G., et al. A generic protein purification method for protein complex characterization and proteome exploration. Nature Biotechnology. 17 (10), 1030-1032 (1999).
재인쇄 및 허가
JoVE'article의 텍스트 или 그림을 다시 사용하시려면 허가 살펴보기
허가 살펴보기더 많은 기사 탐색
This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. 판권 소유