
Method Article
2 in 1: tek adımlı benzeşme arıtma paralel analizi için Protein-Protein ve Protein-metaboliti kompleksleri
* Bu yazarlar eşit katkıda bulunmuştur
Bu Makalede
Özet
Protein-protein ve protein-metaboliti etkileşimler tüm hücresel fonksiyonları için önemlidir. Burada, biz seçim bir protein ile bu etkileşimlerin paralel analiz sağlayan bir protokol tanımlamak. Bizim iletişim kuralı bitki hücre kültürleri için optimize edildi ve benzeşme arıtma kütle spektrometresi tabanlı protein ve metaboliti algılama ile birleştirir.
Özet
Hücresel proteinler, metabolitleri ve nükleik asitler gibi biyolojik moleküller arasındaki etkileşimler tarafından düzenlenmektedir. Protein-protein etkileşimleri (PPI) soruşturma yok yenilik olmakla birlikte, endojen protein-metaboliti etkileşimleri (PMI) karakterize amaçlayan deneysel yaklaşımlar oldukça yeni bir gelişme oluşturmaktadır. Burada, biz aynı anda karakterizasyonu PPI ve PMI yem olarak anılacaktır seçtikleri bir proteinin sağlayan bir protokol mevcut. Arabidopsis hücre kültürleri ve benzeşme arıtma (AP) birleştirir bizim iletişim kuralı (MS) kütle spektrometresi ile optimize edildi-protein ve metaboliti algılama dayalı. Kısacası, bir benzeşme etiketi için erimiş yem protein ifade transgenik Arabidopsis satırları ilk yerel hücresel özü elde etmek için lysed. Anti-etiket antikorlar yem protein protein ve metaboliti ortakları çekmek için kullanılır. Benzeşme saf kompleksleri bir tek adımlı metil tertkullanarak ayıklanır-bütil eter (MTBE) / metanol/su yöntemi. Kutup veya hidrofobik faz metabolitleri ayrı iken, proteinler Pelet içinde bulunabilir. Metabolitleri ve proteinler ultra performanslı sıvı kromatografi-kütle spektrometresi tarafından (UPLC-MS veya UPLC-MS/MS) analiz edilir. Boş-vektör (EV) kontrol şeritleri yanlış pozitif dışlamak için kullanılır. Bizim Protokolü en büyük avantajı fizyolojik şartlarda (hücresel lysate) paralel bir hedef protein protein ve metaboliti ortakların kimlik sağlar olduğunu. Sunulan yöntemi basit, hızlı ve kolayca bitki hücre kültürleri dışındaki biyolojik sistemleri için adapte edilebilir.
Giriş
Yöntem tanımlamak burada amaçları çevre -In-vivo hücresel lysate koşullar seçtiğiniz bir proteinin metaboliti ve protein ortak tanımlaması. Bu bugün karakterize daha çok daha fazla metabolitleri önemli düzenleyici fonksiyonu1var spekülasyonlar. Metabolitleri etkinlik, işlevsellik ve/veya onların reseptör proteinler2,3,4yerelleştirme değiştirme biyolojik anahtarları hareket edebilir. Son on yılda birkaç atılım Yöntem, kimlik PMI vivo içinde veya çevre -In-vivo koşullarda etkinleştirme Gelişmiş5olmuştur. Mevcut yaklaşımları iki gruba ayrılabilir. İlk grup roman protein ortakları tuzak için bilinen-metaboliti yem ile başlatmanız teknikleri kapsar. Benzeşme Kromatografi6, uyuşturucu benzeşme duyarlı hedef-istikrar tahlil7, kemoterapi-proteomik8ve termal Proteom9profil oluşturma yöntemleri içerir. İkinci grup küçük molekül ligandlar10,11tanımlamak için bilinen bir protein ile başlayan tek bir yöntemi içerir.
MS tabanlı lipidomics ile birleştiğinde AP protein-lipid kompleksleri Saccharomyces cerevisiae12. çözümlemeye Bir başlangıç noktası olarak maya suşları 21 enzimler içinde biyosentezi ergosterol dahil ifade yazarlar kullanılan ve 103 kinaz bir tandem-benzeşme saflaştırma (TAP) etiket erimiş. enzimlerin % 70 ve kinazlar % 20'si ışık karmaşık protein-lipid etkileşim ağı akıtan farklı hidrofobik ligandlar bağlamak için bulunamadı.
Daha önce benzer şekilde lipidler için polar ve yarı polar bileşikleri de hücresel lysates13izole protein kompleksleri bağlı kalmasını, göstermek olabilir. Bu bulgulara dayanarak, AP optimize etmek karar yöntemi daha önce bitki hücreleri ve hidrofilik bileşikler14için10,11 yayınlandı. Bu amaçla, biz dokunun vektörel çizimler başarıyla kullanılan bitki PPI çalışmaları15Van Leene ve ark. 2010, tarafından açıklanan kullanılır. Transgenik hatlarını edinmeniz gereken süreyi kısaltmak için Arabidopsis hücre kültürleri üzerinde karar verdi. Çalıştırmaya başladık bir tek adımlı metil tert-butil eter, tek bir protein (Pelet), lipitler (Organik faz) ve hidrofilik metabolitleri (sulu faz)16 karakterizasyonu izin (MTBE) / metanol/su ekstraksiyon yöntemi, benzeşme arıtma deney. EV kontrol şeritleri yanlış pozitif, Örneğin tek başına etiket bağlayıcı proteinler dışlamak için kullanılmaya başlanmıştır. Biz üç (beş) öğesini kavram kanıtı olarak nükleozit nükleotittir kinaz mevcut Arabidopsis genom (NDPK1-NDPK3). Diğer bulgular arasında biz NDPK1 glutatyon Sile etkileşim göstermek olabilir-transferaz ve glutatyon. Sonuç olarak NDPK1 glutathionylation14' e tabi olduğunu kanıtlayabilir.
Özetlemek gerekirse, sunulan Protokolü protein-protein ve protein-küçük-molekül etkileşim ağları karakterize için önemli bir araçtır ve varolan yöntemleri üzerinde büyük bir ilerleme oluşturmaktadır.
Protokol
Klonlama, dönüştürme, seçim ve büyüme koşulları dahil olmak üzere transgenik Arabidopsis hücre kültür satırları hazırlanması17' bulunabilir. Not EV kontrol şeritleri yanlış pozitif olarak düzeltmek için tavsiye edilir. Deney öncesinde, western blot analizi, tandem benzeşme etiketi G-protein parçası karşı IgG antikorları kullanarak Örneğin tarafından overexpression yem protein onaylayın. Bitki hücre kültür malzemeden büyüme medya ayırmak önemlidir.
1. bitki hücre malzemesi deney öncesinde hazırlanması
- Faiz18protein overexpressing bir PSB-L A. thaliana hücre kültür çizgi büyümek.
- 4,43 g/L MSMO 30 g/L sükroz ile karışık içerir MSMO orta hazırlayın. 1 M KOH ile 5,7 arabelleğe pH ve otoklav çözüm ayarlayın. Deney öncesinde 0,5 mg/L α-naphthaleneacetic asit, 0,05 mg/L kinetin ve 50 μg/mL sefaloridin ile orta ek.
- Dönüştürülmüş bitki hücre kültürleri 50 ml 100 mL şişe nazik ajitasyon (130 devir/dakika) bir yörüngesel platformu shaker üzerinde MSMO ortamda yetiştirmek. 20 ° C ve ışık şiddeti Kültür Salonu'nda hücrelerde 80 μmol m-2 s-1eşit büyümek.
- Alt kültür hücrelere taze medya her 7 günde 1:10 sulandrarak.
- Naylon mesh filtre olarak kullanarak bir vakum pompası ile birlikte bir cam huni kullanarak Logaritmik büyüme faz hücreleri toplamak. Alüminyum folyo infiltrat sarın ve sıvı azot içinde dondurmak.
Dikkat: sıvı nitrojen aşırı soğuk olduğunu unutmayın. Hatalı işlenmesi yanıklara neden. Uygun kişisel koruyucu donanımları, izoleli, eldiven, koruyucu gözlük ve önlük termal dahil olmak üzere.
2. musluk Protokolü
Not: Aşağıdaki adımı Maeda vd. 201411 ve Van Leene ve ark. 201117uyarlanmıştır.
- Hasat ve donmuş bitki hücre kültür malzemesi bir mikser değirmen (2 dk 20 Hz) veya harç ve havaneli ince toz elde etmek için kullanarak lunaparkçı. Aliquot 3 g (Toplam protein yaklaşık 90 mg için karşılık gelen) yerden malzeme örnek başına. Sıvı azot önceden soğutulmuş ekipman kullanarak bu adımı sırasında örnek çözdürme kaçının.
Not: Mağaza zemin bitki materyali 50 mL tüp bir AP yordamı başına –80 ° C'de. - Buz gibi lizis arabellek 3 mL sıvı azot precooled harçla örnekte triturate (0,025 M Tris-HCl pH 7.5; 0.5 M NaCl 1.5 mM MgCl20,5 mM DTT; 1 mM NaF; 1 mM Na3VO4; 100 x seyreltilmiş ticari proteaz inhibitörü kokteyl; 1 mM PMSF) malzeme Tammy'nin kadar. Örnek Tammy'nin sonra hemen bir sonraki adıma geçin.
Not: lizis arabellek taze hazırlayın. Bu adımda boş örnekleri tanıtmak. Onlar MS algılama sorunları neden olabilir deterjanlar için tavsiye edilmez. - Hücresel enkaz kaldırmak için malzeme 2 mL microcentrifuge tüpler ve 4 ° C'de 10 dakika 20,817 x g, santrifüj bölün 3 mL açık lysate 15 mL konik santrifüj tüpü içinde toplamak.
- Santrifüjü adımı sırasında IgG-Sepharose boncuk equilibrate. Aliquot 100 µL örnek başına boncuk ve 1 mL lizis arabellek ile yıkayın. Girdap boncuk ve flash-spin resuspend. Lizis arabellek atın ve iki kez tekrarlayın. Boncuk lizis arabellek 400 µL içinde resuspend.
- Boncuk toplanan bitki lysate ekleyin ve karışımı 4 ° C'de 1 h için dönen bir tekerlek üzerinde kuluçkaya
- Transfer karışımı içine bir şırınga radarı-lock kapağı ile bir spin sütun ile filtre gözenek boyutu 35 µm. lysate üzerinden geçmek için Uygula basınç ile birlikte. Lysate üzerinden gidecek iken boncuk ekli kompleksleri ile filtre üzerinde kalır.
Not: İsteğe bağlı olarak, bir vakum manifold sistemi kullanın. Boncuk zarar vermeyecek hafif basınç uygulamak emin olun. - Boncuk, arabellek yıkama 10 mL ile başta yıkayın (0,025 M Tris-HCl pH 7.5; 0.5 M NaCl) ve o zaman 1 mL elüsyon arabelleği ile (10 mM Tris-HCl pH 7.5; 150 mM NaCl; 1000 seyreltilmiş E64 ve 1 mM PMSF x 0,5 mM EDTA). Çamaşır sütun veya vakum manifold sistemi bağlı bir Ģırınga kullanarak gerçekleştirin.
Not: Otelde bir vakum manifold sistemi kullanarak yaptığınız boncuk zarar vermeyecek hafif basınç uygulamak belirli. - Boncuk 400 ile kuluçkaya µL 50 U tütün geliştirilmiş bir sürümü içeren elüsyon arabelleği etch virüs (AcTEV) proteaz. Tablo shaker 1000 devir / dakikada 16 ° C'de 30 dk için kullanın
Not: sütun elüsyon arabellek ekleme alt kapatmak için bir eklenti kullanmayı unutmayın. - İlave bir bölümüne (50 U) enzim sütuna ekleyin ve kuluçkaya karışımı 30 dk altında aynı için yukarıda açıklanan koşullar.
- Eluate 2 mL microcentrifuge tüp içinde Santrifüjü (1 dk, 20,817 x g) veya vakum manifold toplamak. Kalan kompleksleri kaldırmak için elüsyon arabelleği 200 µL kullanarak bir ek elüsyon adım tanıtmak.
Not: –20 ° C veya –80 ° C da örneğine depolamak veya hemen protein ve metaboliti ayıklama adımla devam edin. Donmuş numuneler üzerinde buz çözme.
3. Western Blot Analizi
- Yem varlığını doğrulamak için toplanan eluate protein protein – metaboliti içeren eluate 10 µL SDS-sayfa ve western blot analizi gerçekleştirmek için kullanın. Protein ilgi tanımlamak için fare birincil antikorlar karşı streptavidin-bağlayıcı protein (1: 200), Van Leene ve ark. 201117' açıklandığı gibi TEV proteaz bölünme sonra kalan musluk etiketi bir parçası kullanın. Daha sonra ikincil keçi Anti-fare antikorlar HRP ile birleştiğinde kullanın.
4. metaboliti ve Protein çıkarma
Not: Bu protokol Giavalisco vd. 201116' dan uyarlanmıştır.
Not: Bu adımından itibaren UPLC – MS-sınıf çözümleri kullanır.
- Eklemek 1 mL metil tert-butil eter (MTBE) / metanol/su çözücü (3:1:1) toplanan eluate ve örnek INVERSION tarafından karıştırın. Çözücü –20 ° c ayıklama adım önce soğutulur emin olun.
Dikkat: MTBE ve metanol zararlı maddelerdir. Duman başlık altında ayıklama adımıysa ve uygun bireysel koruma araçları, Örneğin, eldiven giymek. - 0.4 mL metanol: su 1:3 çözüm için her örnek ve örnek içeriği INVERSION tarafından karıştırın.
Not: Faz ayrılmaya takviyesi karışım metanol: su solüsyonu ile sonuçlanır. Lipidler üst aşama içerir, daha düşük Faz kutupsal ve yarı polar metabolitleri içerir ve proteinler Pelet içinde bulunabilir. - Aşama ayrı, örneği ' 20,817 x g için oda sıcaklığında 2 dk santrifüj kapasitesi sonra üst aşama (Bu protokol için bitmedi) lipid ölçümleri için toplamak için bir el ile sıvı işleme pipet 1 mL hacim kapasitesi ile kullanma.
- INVERSION tarafından 0.2 mL metanol ve karışımı ekleyin.
- 20,817 x g RT 2 min için de örnek santrifüj kapasitesi sonra kutup aşama metaboliti ölçümleri (kutupsal ve yarı polar bileşikleri) için toplamak. Protein Pelet rahatsız edici önlemek için sıvı faz tüpün dibinde yaklaşık 50 µL bırakın.
- Metaboliti ölçümler için kuru toplanan örnekleri bir santrifüj evaporatör gecede. Protein Pelet kurutma örnekleri Buharlaştırıcı 30-60 dakika sonra kaldırarak aşırı kaçının.
Not: –20 ° C veya –80 ° C örnekleri depolamak veya hemen LC-MS/MS Analizi proteinler hazırlanması ile devam.
5. proteomik analiz için numune hazırlanması
Not: Bu adımı Olsen ve ark. adapte 200419 ve tripsin/Lys-C Mix teknik kılavuz ( Tablo malzemelerigörmek).
- Örnek enzimatik sindirim gerçekleştirin.
Dikkat: Çözücüler enzimatik sindirim sırasında kullanılan ve örnek desalting zararlı. Duman başlık altında iş ve uygun bireysel koruma araçları, Örneğin, eldiven giymek.- Taze hazırlanmış denatürasyon arabelleği (40 mM amonyum bikarbonat içeren 2 M Tioüre/6 M üre, pH 8) 30 µL protein Pelet geçiyoruz. Daha iyi protein çözünürlük elde etmek için bir 15dk sonication adımı gerçekleştirin. Pelet eriyene kadar yineleyin.
- Örneği ' 20,817 x g 4 ° C'de 10 dakika santrifüj kapasitesi, sonra yeni bir microcentrifuge tüp süpernatant aktarmak.
- Bradford protein tahlil kullanarak protein konsantrasyonu belirlemek.
- İçin bir birim için protein 100 µg eşdeğer analizi, aliquot daha fazla ve 46 µL kadar örnek denatürasyon arabellek ile doldurun.
- Örnek 2 µL taze hazırlanmış azaltma arabelleği (50 mM DTT H2O çözünmüş) eklemek ve oda sıcaklığında 30 dk için kuluçkaya.
- Örnek 2 µL taze hazırlanmış alkillenme arabelleği (40 mM Amonyum Bikarbonat arabellekte çözünmüş 150 mM iodoacetamide) ile tedavi ve karışımı oda sıcaklığında 20 dakika karanlıkta kuluçkaya.
- 40 mM Amonyum Bikarbonat arabelleği 30 µL örnekle sulandırmak ve 20 µL LysC/tripsin karışımı ekleyin.
- 37 ° C'de 4 h kuluçka sonra 40 mM Amonyum Bikarbonat arabelleği 300 µL örnekle sulandırmak.
- 37 ° C'de gecede kuluçka ile devam
- Yaklaşık 20 µL % 10 trifluoroacetic asit pH < 2'yi edinmek için (TFA) ile örnek acidify. Bir pH şerit kullanarak örnek pH kontrol edin.
Not: örnek –20 ° C'de depolayın veya sonraki adıma geçin.
- Sindirilir protein desalt.
Not: Tercihen, bir vakum manifold sistemi kullanın. Sütun aşırı kurutma önlemek.- C18 SPE sütun durulama ( Tablo malzemelerigörmek) 1 mL % 100 ile MeOH ve sonra % 80 Asetonitril (ACN) %0.1 içeren 1 mL ile TFA suda seyreltilmiş. Kullanım, burada ve daha fazla protein-desalting içinde adımları, süreci hızlandırmak için bir vakum manifold sistemi. Sütun aşırı kurutma önlemek.
- Sütun 1 mL TFA suda seyreltilmiş % 0.1 ile iki kez yıkayarak equilibrate.
- Örnek sütuna yerleştirin. Durulama tüp ek 200 µL %0.1 TFA ve transfer çözüm sütun ile. Çözümler sütunu boyunca çalıştırın.
- Sütun 1 mL % 0.1 ile iki kez yıkayın TFA.
- 800 µL % 60 ile sütundan desalted peptidler elute ACN, % 0,1 TFA çözüm. Protein fraksiyonu örnekleri Buharlaştırıcı 30-60 dakika sonra kaldırarak aşırı kurutma kaçınarak bir santrifüj evaporatör toplanan kesir kuru.
Not: –20 ° C veya –80 ° C örnekleri depolamak veya hemen sonraki adıma geçin. Sonraki adımda, buz üzerinde örnekleri tutmak.
6. ölçüm UPLC-MS/MS kullanarak Protein örnekleri hazır.
Not: Proteomik ve metabolomic ölçüleri önce (0.2 µm gözenek boyutu) filtre ve bir vakum pompası için 1 h kullanarak tüm arabellekleri degas.
- Kurutulmuş resuspend peptid Pelet 2 mL microcentrifuge tüp arabellek C (% 3 v/v ACN, % 0,1 v/v formik asit) kullanarak 50 µL içinde depolanan el ile sıvı işleme pipet 200 µL birim kapasitesi ile. 15 dk ultrasonik banyo için örnekleri ile 35 kHz ultrasonik frekans solüsyon içeren temizleyicide.
Dikkat: Afrika Kupası ve formik asit zararlı maddelerdir. Duman başlık altında iş ve uygun bireysel koruma araçları, Örneğin, eldiven giymek. - 20,817 x g 4 ° C'de 10 dakika için de örnek santrifüj kapasitesi, sonra 20 µL süpernatant ile cam şişe aktarmak.
- C18 ters faz sütun kullanarak ayrı sindirilir peptidler sıvı kromatografi için bağlı ve kitle spectra bir Kütle Spektrometre kullanılarak elde etmek.
- Sütun üzerinde bir 300-nl/dk akış hızı kullanarak örneği 3 µL ayırın. Arabellek C ve D (% 63 v/v ACN, % 0,1 v/v formik asit), bir mobil faz için kullanın % 3 ramping gradyan oluşturan %15 ACN ACN 20 dk ve sonra % 30 için ACN sonraki 10 dakika içinde.
Not: –20 ° C veya –80 ° C birkaç ay kadar da örneğine geri kalanını saklayın. Önce proteomik ölçümü, buz üzerinde örnek refreeze. - % 60'ı kullanarak 10 dakikadır kirleri yıkayın ACN ve sütun 5 µL arabelleği C önce sonraki örnek ölçümü ile equilibrate.
- Kitle spectra kullanarak veri bağımlı MS/MS yöntemiyle 70.000 çözünürlük set, AGC hedef 3e6 iyonlar, en büyük enjeksiyon zaman 100 ms ve m/z 1600 için 300 den değişen kazanç. En fazla 15 MS/MS taramalar 17.500 çözünürlüğe, AGC hedef 1e5, en fazla enjeksiyon zaman 1,6 m/z ve m/z 200 den 2000'e kadar bir yalıtım penceresi ile % 20, underfill oranı 100 ms, kazanmak. Apex tetikleyici (6-20 s), 15 s ve hariç tutma ücretleri > 5 ve 1 ayarlama dinamik dışlama etkinleştirmek.
- Sütun üzerinde bir 300-nl/dk akış hızı kullanarak örneği 3 µL ayırın. Arabellek C ve D (% 63 v/v ACN, % 0,1 v/v formik asit), bir mobil faz için kullanın % 3 ramping gradyan oluşturan %15 ACN ACN 20 dk ve sonra % 30 için ACN sonraki 10 dakika içinde.
7. proteomik bilgilerin işlenmesi
- En yeni Arabidopsis thaliana Proteom veritabanı http://www.uniprot.org/ download ve kirletici veritabanı içerir. LC-MS elde edilen ham veri analiz kullanarak varsayılan ayarı ile kullanarak entegre Andromeda peptid arama motoru ile MaxQuant LFQ normalleştirme20,21,22etkin çalışır. Tablo S1içinde kullanılan parametreler hakkında ayrıntılı bilgi bulabilirsiniz.
- "Protein groups.txt" çıktı dosyasını açın. Daha fazla çözümleme için en az iki benzersiz peptidler ile tanımlanan protein grupları için filtre. MaxQuant tarafından tanımlanan potansiyel kirletici ve A. thaliana proteinleri (ARATH Fasta üstbilgileri sütun) veritabanında için filtre protein grupları kaldırın.
- Protein zenginleştirme örnekleri arasında önemini sınamak için normalleştirilmiş LFQ yoğunluklarda kullanın ve unpaired, iki kuyruklu öğrenci t-testi birden çok karşılaştırma düzeltme (Örneğin Benjamini & Hochberg yanlış bulma oranı (FDR) tarafından takip gerçekleştirmek Düzeltme veya Bonferroni düzeltmesi).
- P-değeri için EV hakim olmak ve NDPK1 elde edilen LFQ yoğunluklarda karşılaştırarak hesaplayın. Tüm belirsiz değerleri dışarıda filtre. P-değerleri artan sırada sıralamak ve FDR düzeltme hesaplamak için R komut dosyası veya online hesap makinesi (Örneğin, https://www.sdmproject.com/utilities/?show=FDR) kullanın. 0,1 altındaki FDR değerler için filtre.
Not: veri analizi formu araştırmaları için uygun göz önünde bulundurun. Nicel çalışmalar (protein zenginleştirme örnekleri arasında Analizi) "LFQ yoğunluk" değeri kullanır için nitel araştırma (varlığı ya da yokluğu belirli protein) "Yoğunluk" değeri seçin. - EV denetimine NDPK1 karşılaştırarak daha bol protein grupları için filtre. Yerelleştirme SUBA veritabanı23 kullanarak potansiyel protein ortakları belirlemek ve NDPK1 ile birlikte lokalize protein için düzeltmek.
- P-değeri için EV hakim olmak ve NDPK1 elde edilen LFQ yoğunluklarda karşılaştırarak hesaplayın. Tüm belirsiz değerleri dışarıda filtre. P-değerleri artan sırada sıralamak ve FDR düzeltme hesaplamak için R komut dosyası veya online hesap makinesi (Örneğin, https://www.sdmproject.com/utilities/?show=FDR) kullanın. 0,1 altındaki FDR değerler için filtre.
8. kutup faz UPLC – MS kullanarak içeren örnekleri ölçümü.
- Resuspend adım 4.5 200 µL su kutup fazından ve örnek 5 min için solüsyon içeren temizleyicide.
- 20,817 x g 4 ° C'de 10 dakika için de örnek santrifüj kapasitesi sonra süpernatant bir cam şişe aktarmak.
Not: –20 ° C veya –80 ° C için birkaç ay kadar da örneğine geri kalanını saklayın. Metabolomic ölçüm öncesinde örnek buz üzerinde refreeze. - UPLC C18 ters faz sütun için birleştiğinde kullanarak bir ayrılık adımıysa ve kitle spectra MS ile kazanmak.
- Enjeksiyon her iyonlaşma modu (pozitif ve negatif) başına örnek sütun 2 µL üzerine yük ve 400 µL/dk akış hızı kullanarak kesir ayrı. Metaboliti ölçüm için gerekli degrade oluşturmak için aşağıdaki gibi mobil faz çözüm hazırlamak: bir (%0,1 formik asit H2O) tampon ve B tampon (ACN %0,1 formik asit).
- 400 µL/dak ve aşağıdaki degrade ayrı metabolitleri: 1 min % 99'a, arabellek arabellek arabellek A, 13-min doğrusal gradyan arabellek arabellek A, 15dk doğrusal gradyan % 30 dan % 30'u A % 60'dan % 60'a % 99'u üzerinden 11-min doğrusal gradyan tampon A-% 1 kadar 16 dk. 17 dk, kullanım arabellek A. yeniden equilibrate % 99'u doğrusal gradyan arabellek A % 1'den 3 min için sütun % 99 konsantrasyon ile başlayarak A sonraki örnek ölçü önce arabellek arabellek A, %1 konsantrasyon tutun.
- Yığın aralığı kapsayan toplu spectra elde 100 ve 1500 m/z ile kararlılık ayarlamak için 25.000 arasında yükleme süresi 100 Bayan ayarla AGC hedef 1e6için sınırlı, 3kV bir kılıf ile kılcal gerilim gaz akışı ve yardımcı gaz değeri 60 ve 20 , sırasıyla. Kapiller sıcaklık 250 ° C ile kepçe gerilim 25V için ayarlayın.
9. Metabolomics bilgilerin işlenmesi
- İşlemi her iki iyonlaşma modları arasından elde chromatograms toplanan. Ücret oranı (m/z), tutma zamanı (RT) ve ilişkili tepeler, Örneğin, ticari yazılım yoğunluğunu ayine ayıklamak için yazılım kullanın (bkz. Tablo reçetesi) veya alternatif24.
- .Exe dosyasını çift tıklatarak işleme yazılımı başlatmak
- Yeni iş akışı, etkinlik "Yük--dan eğe" için arama oluşturup bu etkinliği "sürükle ve bırak" tarafından boş iş akışı uzaya. Sağ fare tuşu ile faaliyet basın ve aktivitesinin ayarlarını açın.
- "Genel" ayarları içeren sekmeyi, bir deneme "Ad" alanına ayarlayın ve sonraki "Select dosya ve klasörleri" tıklatın ve ham chromatograms işaretleyin.
- "Gelişmiş" ayarları içeren sekmeyi, "Profil veri kesme" için 0 yoğunluğunu ayarlayın. "Uygula" ve "OK"'ı tıklatın.
- Arayın ve etkinlik "Veri süpürme" ekleyin. Sağ fare tuşu ile faaliyet basın ve aktivitesinin ayarlarını açın.
- "Genel" ayarları içeren sekmeyi, mark "Centroid veri" ve "MS/MS veri". Tüm MS/MS veri "Her şeyi" seçim panelinde seçerek kaldırabilirsiniz.
- Arayın ve etkinlik "Kromatografik kimyasal gürültü çıkarma" ekleyin. Sağ fare tuşu ile faaliyet basın ve aktivitesinin ayarlarını açın.
- "Genel" ayarları içeren sekmeyi "Kromatografik yumuşatma" işaretleyin ve inceden inceye gözden geçirmek "3" ve "Hareketli ortalama" için "Tahmincisi" sayısını belirleyin. "RT pencere" 51 inceden inceye gözden geçirmek, % 50, çıkarma "Yöntemi" ve 750 yoğunluğu "Eşik" "parametreli" için ayarlayın.
- "Gelişmiş" ayarları içeren sekmeyi "RT yapısı kaldırma" işaretle ve "En az RT uzunluğu" 5 taramalarla ayarlayın.
- "Gelişmiş" ayarları içeren sekmeyi, "m/z yapısı kaldırma" işaretleyin ve "En az m/z uzunluğu" 3 noktaya ayarla.
- Arayın ve etkinlik "Kromatografik RT hizalama" ekleyin. Sağ fare tuşu ile faaliyet basın ve aktivitesinin ayarlarını açın.
- "Genel" ayarları içeren sekmesinde "İkili hizalama Bankası ağaç" için "hizalama düzeni" ve "RT arama aralığı" 0,5 dk için ayarlayın.
- "Gelişmiş" ayarları içeren sekmede varsayılan parametrelerini kullanın.
- Aramak ve etkinlik "Peak algılama" Etkinlikler "Kromatografik" grubuna ekleyin. Sağ fare tuşu ile faaliyet basın ve aktivitesinin ayarlarını açın.
- "Genel" ayarları içeren sekmesinde "Toplamı pencere" 0.09 dk, "En az en yüksek boyutu" 0,03 dk, 5 puan "en fazla birleştirme uzaklık" ve "Birleştirme stratejisi" "Merkezlerine" ayarlayın. "Tepe RT bölme içinde" kutu "Gap/tepe oranı" % 50'ye ayarlayın.
- "Gelişmiş" ayarları içeren sekmeyi, 5 puan, % 80 oranında "iyileştirme eşiği" ve "Tutarlılık eşiği" 1 "Yumuşatma pencere" kurmak. "Merkezi hesaplama" "Yoğunluk ağırlıklı" ile "Yoğunluk %70 ayarlamak eşik" ayarlayın.
- Aramak ve etkinlik "İzotop kümeleme" Etkinlikler "Kromatografik" grubuna ekleyin. Sağ fare tuşu ile faaliyet basın ve aktivitesinin ayarlarını açın.
- "Genel" ayarları içeren sekmesinde, "RT hoşgörü" 0,015 min ve "m/z hoşgörü" için 5 ppm için ayarlayın.
- Sekmede içeren "Zarf uydurma" ayarları, grubu "Yöntemi" olarak "Hayır şekil kısıtlama" ve "İyonlaşma" olarak "Protonation (için olumlu modu) ve"Deprotonation"(için negatif modu). "Minimum ve maksimum ücret" 1 ve 4, anılan sıraya göre ayarlayın.
- "Gelişmiş" ayarları içeren sekmede varsayılan parametrelerini kullanın.
- Arayın ve etkinlik "Aynı cinsten tek adet filtre" ekleyin.
- Veri işleme sonuçlarını vermek için aramak ve faaliyetleri "Dışa aktar" grubundaki "Analist" aktivitesi ekleyin.
- "Genel" ayarları içeren sekmesinde "Türü" "Kümeler" ve "Gözlemlenebilir" olarak "Özetlenebilir yoğunluğu" olarak ayarlayın. "Özel hedef" seçin ve dizin verme dosyasının belirtin.
- "Gelişmiş" ayarları içeren sekmede varsayılan parametrelerini kullanın.
- Kurum içi başvuru-bileşik veritabanı kullanarak toplu özellikleri açıklama ekleyin.
- Bir veya birden çok MS sınıf başvuru-bileşik kullanarak UPLC-Bayan LC-MS yöntemin referans-birleştirici karışımlar ve faiz protein ile birlikte saf metabolitleri analizi için analiz.
Not: Bu çalışma için yaklaşık 300 dipeptides bir dizi analiz ve başvuru-bileşik Kütüphane olarak kullanılan. - Analiz kullanmak için belirli m/z ve RT ile ilişkili ham kromatografik dosya ve arama açmak için (bkz: Malzemeler tablo) yazılım ölçülen referans-bileşik (kullanım kılavuzuna bakın).
Not: Sekonder metabolitler iyonlaşma türlerinde farklı. Ortak varlığı [M-H], iyon M-1.007276, M + 1.007276, M + 18.033823 ve M + 22.989218 kitle eşit için arayarak adducts adlı [M + H], [M + NH4] ve [M + Na], anılan sıraya göre. - Kullanım elektronik tablosunu açmak üzere verilen "Analist" dosyayı chromatograms ve arama için belirli iyon işleme sonra elde edilen kitle. RT deney ve RT başvuru bahçedeki ölçülen kitle özelliği karşılaştırın. Bir sapma 0.005 Da m/z için ve 0.1 min RT. için izin
- Bir veya birden çok MS sınıf başvuru-bileşik kullanarak UPLC-Bayan LC-MS yöntemin referans-birleştirici karışımlar ve faiz protein ile birlikte saf metabolitleri analizi için analiz.
- Örnek (faiz vs EV denetiminin overexpressed protein satır) arasında belirli protein ile birlikte saf metaboliti zenginleştirme önemini sınamak için kullanarak iki kuyruklu sigara eşleştirilmiş öğrenci t-test çoklu tarafından takip en yüksek değerlerini karşılaştırma Karşılaştırma düzeltme (Örneğin Benjamini & Hochberg yanlış bulma oranı düzeltme veya Bonferroni düzeltmesi).
Sonuçlar
Orijinal çalışmada, üç A. thaliana NDPK genler PSB-L hücre süspansiyon kültürlerinde bünye 35S organizatörü14 (Şekil 1) kontrol altına overexpressed. Tandem benzeşme etiket carboxy - veya amino-terminal bir ucunu bir yem protein için erimiş. Benzeşme saf kompleksleri MTBE/metanol/su ekstraksiyon16için tabi tutuldu. Proteinler ilgi çekti ve küçük moleküller MS (Tablolar S2 ve S3) kullanarak tespit edilmiştir.
Yanlış pozitif olarak düzeltmek için küçük molekül kirletici kimyasallar ve laboratuvar sarf malzemeleri dışlamak için boş örnekleri kullanılmıştır. Ayrıca, metabolitleri ve ya bir benzeşme etiketi bağlamak veya yalnız reçine proteinler EV kontrol şeritleri kullanarak sorumluydu. Gerçek pozitif durumlar, iki kuyruklu sigara eşleştirilmiş öğrenci t-testi ve Benjamini & Hochberg yanlış bulma oranı almak için düzeltme metabolitleri (Tablo S4) ve önemli ölçüde NDPKs AP zenginleştirilmiş proteinler (Tablo S5) tanımlamak için uygulanan (N - ve C-ölümcül NDPKs öğesini) deneyler ile karşılaştırıldığında EV kontrol şeritleri (FDR < 0,1). Önceki çalışma, biz devamsızlık/varlığı ölçüt protein ve küçük molekül interactors betimlemek için kullandığını unutmayın.
Temsilcisi sonuçları için NDPK1, dipeptides, bizim grupta okudu küçük molekül düzenleyiciler roman bir sınıfın metaboliti veri odaklanmak süre verilir. Proteomik çözümleme NDPK1 26 sözde protein ortaklarının ortaya. Co NDPK1 aynı hücre altı yerde lokalize proteinler için daha fazla filtreleme tarafından (sitozol), listenin 13 sözde protein interactors aşağı daralmış. Glutatyon Sarasında saptanan proteinler vardı-transferaz, iki uzama başlama faktörleri, tübülin ve aconitate hidrataz. Metabolomic çözümleme ortaya dört dipeptides Val-Leu, Ile-Glu, Leyi-Ile ve Ile-bu özellikle birlikte eluted Phe NDPK1 ile (Şekil 2). Tüm dört dipeptides onların N-terminus paylaşılan bağlama özgüllük düşündüren, hidrofobik bir kalıntı paylaşmak unutmayın.
Bakmak için bilinen protein-protein ve protein-metaboliti kompleksleri için biz sorgulanan 13 proteinler ve dikiş veritabanı25 (Şekil 3) karşı dört dipeptides tespit edilmiştir. Birkaç gözlemler yapılmış olabilir: (i) interactors hiçbiri daha önce NDPK1 için bildirildi. (II) APX1 ortholog aldehid dehidrojenaz aile üyesi ALDH7B4, çeviri başlatma faktörü FBR12 faktörle gen AT2G40290 tarafından kodlanmış başka bir çeviri başlatma sırasında etkileşimli olarak bildirildi. (iii) tanımlanan dipeptides Hayır protein ortakları bildirdi. Ortak eluted dipeptides daha önce herhangi bir erişim tarihi bitki proteini ilişkili rapor değil. Ancak, diğer canlılar arasında önemli rol oynarlar: Leu-Ile, Örneğin, bir insan hücre satırı26' neurotrophin harekete geçirmek bir etkisi vardır. Not deneme sistemi tam topolojisi tanımlama izin vermez. Örneğin, bir dipeptid NDPK1 ile doğrudan etkileşim olabilir ama herhangi bir ortak saf protein de ilgili olabilir.
Birlikte ele alındığında, bizim sonuçları AP kütle spektrometresi ile birlikte istihdam kurulan yordamı protein-protein ve protein-küçük-molekül interactors tanımlaması kolaylaştırır ve yardımcı olur hakkında geniş bilgi oluşturmak göster hedef protein interactome.

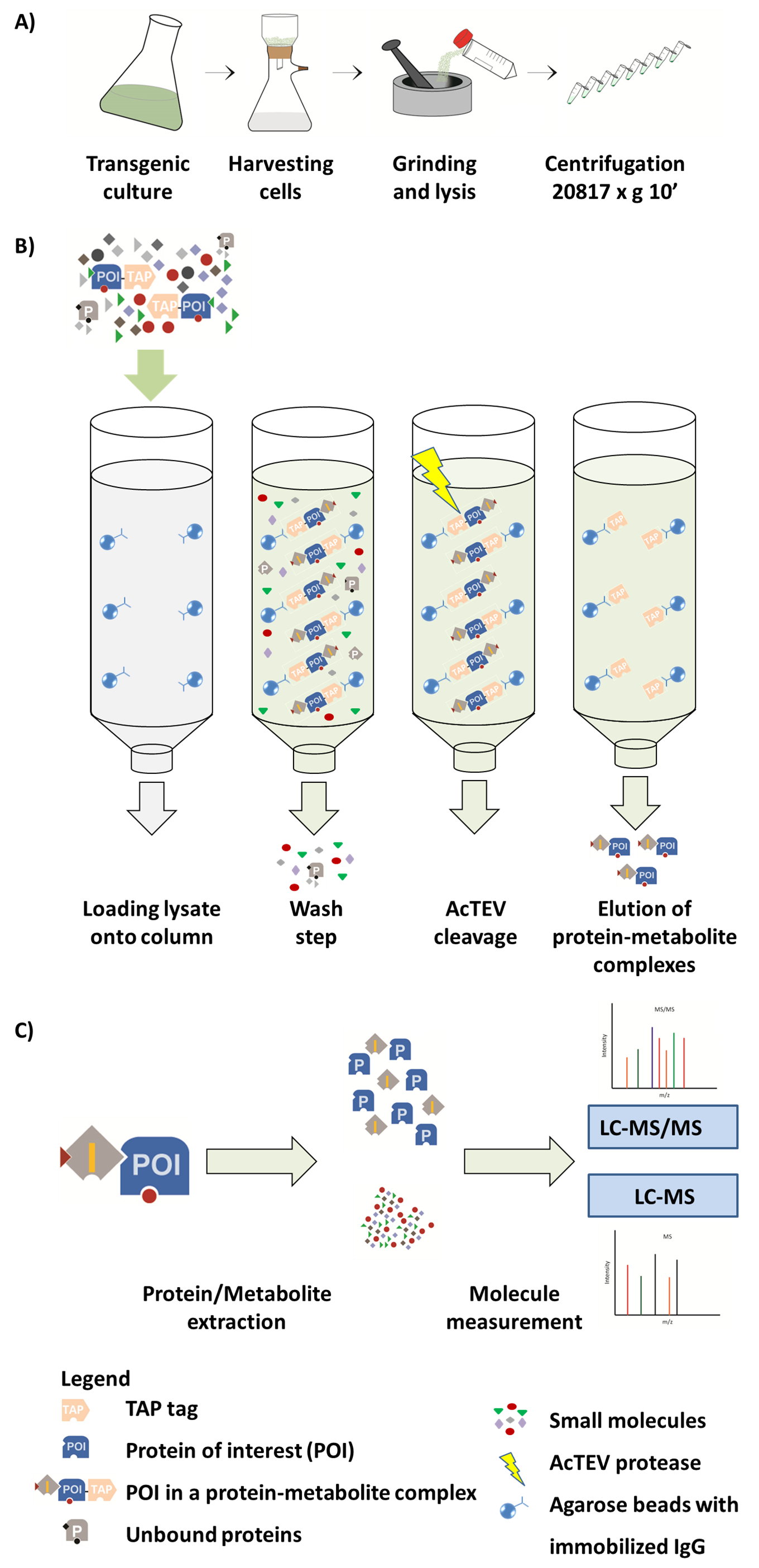
Şekil 1. AP-MS iş akışı şeması. (A) bir yerel çözünür kesir bitki hücre kültürü üzerinden hazırlanması. (B) AP yordam sonraki adımda. Örnek sütuna yükledikten sonra bir dokunun etiketi erimiş faiz (POI) protein üzerinde özel boncuk immobilize IgG antikor bağlar. Çamaşır sütun ilişkisiz proteinler ve metabolitleri kaldırılmasını kolaylaştırır. AcTEV bölünme gerçekleştirdikten sonra POI protein-metaboliti kompleksleri eluted. Kompleksleri (C) ayrılması protein ve metaboliti kesir yarı kantitatif MS Analizi tarafından takip içine. Bu rakam bir parçası tekrar Luzarowski ve ark. 201714oluşturulur. Bu rakam daha büyük bir versiyonunu görüntülemek için buraya tıklayınız.
{kind=link}

Şekil 2. Özellikle NDPK1 ile birlikte eluting dipeptides. Dört dipeptides Val-Leyi (A), Ile-Glu (B), Leyi-Ile (C)ve Ile-Phe (D) AP deneyde ölçülen ortalama yoğunluklarda çizildi. Tüm dört dipeptides NDPK1 örnekleri EV denetimine göre önemli zenginleştirme göstermek (yıldız işareti temsil FDR < 0,1). Hata çubukları temsil eden 6 ölçümler için standart hata (N-3 çoğaltır ve 3 / C-ölümcül proteinler öğesini). Bu rakam daha büyük bir versiyonunu görüntülemek için buraya tıklayınız.
{kind=link}

Şekil 3. Tüm molekülleri NDPK1 ile birlikte eluting etkileşim ağı yalnızca önceki göz önünde bulundurarak dikiş veritabanında sorgulanan deneysel ve veritabanı delilleri (güven > 0,2). Daha yüksek güven daha yüksek etkileşim olasılığını gösterir ve yatırılan veriler temel alınarak hesaplanır. Bu rakam daha büyük bir versiyonunu görüntülemek için buraya tıklayınız.
{kind=link}
Tablo S1. MaxQuant tablo "parameters.txt" çıktı. Tablo tanımı ve miktar gibi kullanılan veritabanları hakkında bilgi için eşik değerlerini içerir. Bu dosyayı indirmek için buraya tıklayınız.
Tablo S2. Bilgi MaxQuant dan çıkış tablo "proteinGroups.txt". Tablo tüm saptanan protein grupları, yoğunluklarda ve benzersiz peptidler ve puan sayısı gibi ek bilgileri listesini içerir. Bu dosyayı indirmek için buraya tıklayınız.
Tablo S3. Kutup metabolitleri analizini içeren çıkış dosyası. Tablo tüm tanımlanan kitle özellikleri belirli m/z, RT ve yoğunluğu ile karakterize bir listesini içerir. Bu dosyayı indirmek için buraya tıklayınız.
Tablo S4. Dipeptides hangi NDPK1, NDPK2 veya NDPK3 yem olarak kullanılan AP örnekleri buldum. Dipeptides boş örnekleri mevcut listeden dışlanan. Her NDPK için iki bağımsız çizgi (ya da N - veya C-terminus öğesini) nüsha işletilmiştir. Öğrenci t-test ve daha fazla pdüzeltilmesi-Benjamini kullanarak değer & Hochberg yöntemi NDPKs önemli ölçüde zenginleştirilmiş interactor ortakları belirlemek için kullanılmıştır (FDR < 0,1). Verilen ΔRT başvuru bileşikleri ve Δppm kitle Metlin27' verilen monoisotopic ile ilgili olarak ilgili olarak hesaplanır. Bu dosyayı indirmek için buraya tıklayınız.
Tablo S5. NDPK1 ile birlikte saf protein. Her NDPK için iki bağımsız çizgi (ya da N - veya C-terminus öğesini) nüsha çalıştırmak. Öğrenci t-test ve daha fazla pdüzeltilmesi-Benjamini kullanarak değer & Hochberg yöntemi NDPKs önemli ölçüde zenginleştirilmiş interactor ortakları belirlemek için kullanılmıştır (FDR < 0,1). Bu dosyayı indirmek için buraya tıklayınız.
Tartışmalar
Sunulan Protokolü hedef protein s ve PM kompleksleri paralel kimliği sağlar. Sonuçlar için klonlama üzerinden deneme az 8-12 hafta olarak tamamlanabilir. Tam AP yaklaşık 4-6 h 12-24 örnekleri, bir dizi için bizim protokol orta üretilen iş analizi için uygun işleme alır.
İletişim kuralının rağmen varlık genel olarak basit bir dizi kritik adım vardır. (i) yeterli miktarda giriş protein ve benzeşme boncuk metaboliti algılama dinamik bir dizi ulaşmak çok önemlidir. Etkin hücre lizis bu nedenle yordamı çok önemli bir adımdır. Zavallı protein verimleri yetersiz pulverizasyon malzeme veya suboptimal lizis-tampon/malzeme oranı bir sonucu olabilir. (ii) kullanılan reaktifler MS-dostu olduğunu özen gösterilmelidir. MS algılama ile müdahale güçlü deterjanlar, gliserol veya aşırı miktarda tuz kaçınılmalıdır. (iii) özel boncuk adımları yıkama sırasında aşırı kuru olmamalı, ve bir vakum manifold kullanırken bu şekilde değil boncuk yok veya karmaşık istikrar etkilemek için bir yavaş akış hızı uygulamak önemlidir.
Bazı önemli olası değişiklikler için sunulan Protokolü vardır: (i) kurucu CaMV35S organizatörü yem protein miktarı en üst düzeye çıkarmak için kullanıyoruz. Overexpression, çok yararlı olsa da, hücre homeostazı28 üzerinde ciddi etkiler ve fizyolojik alakasız etkileşimleri oluşumuna yol. İfade tagged proteinlerin kullanarak yerel rehberleri ve nerede mümkün bir işlev kaybı arka planda gerçek biyolojik interactors almak için üstün olarak kabul edilir. Bitki hücre kültürleri içinde normalde proteinlerin için bitki arka plan ilgili interactors tanımlamak gerekli olabilir. (ii) zar proteinleri ile çalışırken, bir MS uyumlu deterjan ile takıma girecek lizis arabelleği gerektiriyor. (iii) ikinci bir benzeşme arıtma adım getirilmesi yanlış-mutlak gerçek pozitif oranı geliştirmek ve EV denetimleri29ihtiyacını ortadan kaldırmak. Bir roman tandem etiketi iki bağımsız proteaz-bölünme site ile zahmetli ve zaman alıcı tarafından Maeda vd. 201411, ekledi boyut-dışlama Kromatografi adım için cazip bir alternatif sunar.
AP'nin en ciddi sıkıntıları yanlış pozitif yüksek oranıdır. Nedenleri çoktur. Bünye overexpression zaten belirtilmiştir. Başka bir kaynak fizyolojik alakasız etkileşimlerin izole organelleri ile çalışma sürece bütün hücreli lysates karışımları proteinler ve metabolitleri üzerinden farklı hücre altı bölmeleri içeren hazırlıktır. Hücre altı yerelleştirme için doğru interactors süzmek için kullanılır. Yine de, yanlış mutlak çoğunluğu özel reçineler ve proteinler arasında belirsiz bağ neden. İkinci bir arıtma adım giriş yukarıda açıklandığı gibi sorunun en iyi çözümü sunmaktadır, ancak zaman ve işlem hacmi pahasına geliyor. Ayrıca, protokol uzatır gibi zayıf etkileşim kaybolabilir. AP'nin başka bir uyarı sağladığı hakkında kapsamlı bilgi hedef protein rağmen interactome, baited protein doğrudan ve dolaylı hedefler arasında differentiating mümkün olmadığı. Hedeflenen bimolecular yaklaşımlar etkileşimleri onaylamak için ihtiyaç vardır.
MS tabanlı metabolomics ile birleştiğinde AP protein-kompleksleri S. cerevisiae12. çalışırdım Benzer şekilde lipidler için polar ve yarı polar bileşikleri hücresel lysates izole protein kompleksleri bağlı kalmasını, bizim daha önceki gözlem13 ile birlikte bu iş için sunulan iletişim kuralı kavramsal zemin sağladı. Bizim iletişim kuralı tarafından üç benzersiz noktaları ile karakterize: (i) buna karşılık12Maya için iş, AP sadece hidrofobik ama aynı zamanda hidrofilik protein ligandlar almak için uygun olduğunu gösterir. (ii) üç tek ayıklama Protokolü tanıtarak, tek bir AP protein ve metaboliti interactors yem protein eğitim için kullanılabilir. (iii) Biz hücreleri bitki Protokolü uyarlanmış.
Gelecekteki çabalar bir roman tandem etiketi ile iki bağımsız proteaz-bölünme site oluşturmaya özen gösterin. Ayrıca bitki hormonları gibi düşük-bereket küçük moleküllere protokol uygunluğu keşfetmek istiyorsunuz.
Açıklamalar
Yazarlar ifşa gerek yok.
Teşekkürler
Lütfen proje, verimli tartışmalar ve büyük denetleme oynadığı için Prof. Dr. Lothar Willmitzer kabul etmek istiyoruz. Bize proteomik MS ölçümleri ile yardımcı olduğunuz için Dr. Daniel Veyel için minnettarız. Bayan Änne Michaelis kim bize çok değerli teknik yardım LC-MS ölçümleri ile sağladığı için teşekkür ederiz. Ayrıca, Dr Monika Kosmacz ve Dr. Ewelina Sokołowska Weronika Jasińska için onların yardım ve orijinal el yazması ve çalışma katılımı için teknik destek için teşekkür etmek istiyorum.
Malzemeler
| Name | Company | Catalog Number | Comments |
| Murashige and Skoog Basal Salts with minimal organics | Sigma-Aldrich | M6899 | |
| 1-Naphthylacetic acid | Sigma-Aldrich | N1641 | |
| Kinetin solution | Sigma-Aldrich | K3253 | |
| Tris base | Sigma-Aldrich | 10708976001 | |
| NaCl | Sigma-Aldrich | S7653 | |
| MgCl2 | Carl Roth | 2189.1 | |
| EDTA | Sigma-Aldrich | 3609 | |
| NaF | Sigma-Aldrich | S6776 | |
| DTT | Sigma-Aldrich | D0632 | |
| PMSF | Sigma-Aldrich | P7626 | |
| E-64 protease inhibitor | Sigma-Aldrich | E3132 | |
| Protease Inhibitor Cocktail | Sigma-Aldrich | P9599 | |
| Na3VO4 | Sigma-Aldrich | S6508 | |
| AcTEV Protease | Thermo Fischer Scientific | 12575015 | |
| Rotiphorese Gel 30 (37,5:1) | Carl Roth | 3029.2 | |
| TEMED | Carl Roth | 2367.3 | |
| PageRuler Prestained Protein Ladder | Thermo Fischer Scientific | 26616 | |
| SBP Tag Antibody (SB19-C4) | Santa Cruz Biotechnology | sc-101595 | |
| Goat anti-mouse IgG-HRP | Santa Cruz Biotechnology | sc-2005 | |
| Bradford Reagent | Sigma-Aldrich | B6916 | |
| Trypsin/Lys-C Mix, Mass Spec Grade | Promega | V5071 | |
| Urea | Sigma-Aldrich | U5128 | |
| Thiourea | Sigma-Aldrich | T8656 | |
| Ammonium bicarbonate | Sigma-Aldrich | 9830 | |
| Iodoacetamide | Sigma-Aldrich | I1149 | |
| MTBE | Biosolve | 138906 | |
| Methanol | Biosolve | 136806 | |
| Water | Biosolve | 232106 | |
| Acetonitrile | Biosolve | 12006 | |
| Trifluoroacetic acid | Biosolve | 202341 | |
| Formic acid | Biosolve | 69141 | |
| Unimax 2010 Platform Shaker | Heidolph | 5421002000 | |
| Nylon Mesh (Wire diameter 34 µM, thickness 55 µM, open area 14%) | Prosepa | Custom order | |
| Glass Funnel, 47 mm, 300 ml | Restek | KT953751-0000 | |
| Filter Bottle Top 500 mL 0,2 µM Pes St | VWR International GmbH | 514-0340 | |
| Mixer Mill MM 400 | Retsch GmbH | 207450001 | |
| IgG Sepharose 6 Fast Flow | GE Healthcare Life Sciences | 17-0969-02 | |
| Mobicol ""Classic"" with 2 different screw caps without filters | MoBiTec GmbH | M1002 | |
| Filter (small) 35 µM pore size, for Mobicol M 1002, M1003, M1050 & M1053 | MoBiTec GmbH | M513515 | |
| Variable Speed Tube Rotator SB 3 | Carl Roth | Y550.1 | |
| Rotary dishes for rotators SB 3 | Carl Roth | Y555.1 | |
| Resprep 24-Port SPE Manifolds | Restek | 26080 | |
| Finisterre C18/17% SPE Columns 100mg / 1ml | Teknokroma | TR-F034000 | |
| Autosampler Vials | Klaus Trott Chromatographie-Zubehör | 40 11 01 740 | |
| Acclaim PepMap 100 C18 LC Column | Thermo Fischer Scientific | 164534 | |
| EASY-nLC 1000 Liquid Chromatograph | Thermo Fischer Scientific | LC120 | |
| Q Exactive Plus Hybrid Quadrupole-Orbitrap Mass Spectrometer | Thermo Fischer Scientific | IQLAAEGAAPFALGMBDK | |
| Acquity UPLC system | Waters | Custom order | |
| ACQUITY UPLC HSS C18 Column, 100A, 1.8 µM, 2.1 mM X 100 mM, 1/pkg | Waters | 186003533 | |
| High-power ultrasonic cleaning baths for aqueous cleaning solutions | Bandelin | RK 31 | |
| Genedata Expressionist | Genedata | NaN | |
| Xcalibur Software | Thermo Fischer Scientific | NaN | |
| MaxQuant | NaN | NaN |
Referanslar
- Li, X., Snyder, M. Metabolites as global regulators: A new view of protein regulation. Bioessays. 33 (7), 485-489 (2011).
- Jacob, F., Monod, J. Genetic regulatory mechanisms in the synthesis of proteins. Journal of Molecular Biology. 3 (3), 318-356 (1961).
- Schlattner, U., et al. Dual Function of Mitochondrial Nm23-H4 Protein in Phosphotransfer and Intermembrane Transfer a cardiolipin-dependent switch. Journal of Biological Chemistry. 288 (1), 111-121 (2013).
- Ramírez, M. B., et al. GTP binding regulates cellular localization of Parkinson's disease-associated LRRK2. Human Molecular Genetics. , ddx161 (2017).
- Jung, H. J., Kwon, H. J. Target deconvolution of bioactive small molecules: the heart of chemical biology and drug discovery. Archives of Pharmacal Research. 38 (9), 1627-1641 (2015).
- Harding, M. W., Galat, A., Uehling, D. E., Schreiber, S. L. A receptor for the immunosuppressant FK506 is a cis-trans peptidyl-prolyl isomerase. Nature. 341 (6244), 758-760 (1989).
- Lomenick, B., et al. Target identification using drug affinity responsive target stability (DARTS). Proceedings of the National Academy of Sciences of the United States of America. 106 (51), 21984-21989 (2009).
- Manabe, Y., Mukai, M., Ito, S., Kato, N., Ueda, M. FLAG tagging by CuAAC and nanogram-scale purification of the target protein for a bioactive metabolite involved in circadian rhythmic leaf movement in Leguminosae. Chemical Communications. 46 (3), 469-471 (2010).
- Pantoliano, M. W., et al. High-density miniaturized thermal shift assays as a general strategy for drug discovery. Journal of Biomolecular Screening. 6 (6), 429-440 (2001).
- Li, X., Snyder, M. Analyzing In vivo Metabolite-Protein Interactions by Large-Scale Systematic Analyses. Current Protocols in Chemical Biology. , 181-196 (2010).
- Maeda, K., Poletto, M., Chiapparino, A., Gavin, A. -. C. A generic protocol for the purification and characterization of water-soluble complexes of affinity-tagged proteins and lipids. Nature Protocols. 9 (9), 2256-2266 (2014).
- Li, X., Gianoulis, T. A., Yip, K. Y., Gerstein, M., Snyder, M. Extensive in vivo metabolite-protein interactions revealed by large-scale systematic analyses. Cell. 143 (4), 639-650 (2010).
- Veyel, D., et al. System-wide detection of protein-small molecule complexes suggests extensive metabolite regulation in plants. Scientific Reports. 7, (2017).
- Luzarowski, M., et al. Affinity purification with metabolomic and proteomic analysis unravels diverse roles of nucleoside diphosphate kinases. Journal of Experimental Botany. , (2017).
- Van Leene, J., et al. Targeted interactomics reveals a complex core cell cycle machinery in Arabidopsis thaliana. Molecular systems biology. 6 (1), 397 (2010).
- Giavalisco, P., et al. Elemental formula annotation of polar and lipophilic metabolites using 13C, 15N and 34S isotope labelling, in combination with high-resolution mass spectrometry. The Plant Journal. 68 (2), 364-376 (2011).
- Van Leene, J., et al. Isolation of transcription factor complexes from Arabidopsis cell suspension cultures by tandem affinity purification. Plant Transcription Factors: Methods and Protocols. , 195-218 (2011).
- Van Leene, J., et al. A tandem affinity purification-based technology platform to study the cell cycle interactome in Arabidopsis thaliana. Molecular & Cellular Proteomics. 6 (7), 1226-1238 (2007).
- Olsen, J. V., Ong, S. -. E., Mann, M. Trypsin cleaves exclusively C-terminal to arginine and lysine residues. Molecular & Cellular Proteomics. 3 (6), 608-614 (2004).
- Cox, J., Mann, M. MaxQuant enables high peptide identification rates, individualized p.p.b.-range mass accuracies and proteome-wide protein quantification. Nature Biotechnology. 26 (12), 1367-1372 (2008).
- Cox, J., et al. Andromeda: A peptide search engine integrated into the MaxQuant environment. Journal of Proteome Research. 10 (4), 1794-1805 (2011).
- Tyanova, S., Temu, T., Cox, J. The MaxQuant computational platform for mass spectrometry-based shotgun proteomics. Nature Protocols. 11 (12), 2301 (2016).
- Hooper, C. M., et al. SUBAcon: a consensus algorithm for unifying the subcellular localization data of the Arabidopsis proteome. Bioinformatics. 30 (23), 3356-3364 (2014).
- Katajamaa, M., Orešič, M. Data processing for mass spectrometry-based metabolomics. Journal of Chromatography A. 1158 (1-2), 318-328 (2007).
- Szklarczyk, D., et al. STITCH 5: augmenting protein-chemical interaction networks with tissue and affinity data. Nucleic Acids Research. 1277, (2015).
- Tanaka, K. -. i., et al. Dipeptidyl compounds ameliorate the serum-deprivation-induced reduction in cell viability via the neurotrophin-activating effect in SH-SY5Y cells. Neurological Research. 34 (6), 619-622 (2012).
- Smith, C. A., et al. METLIN: A metabolite mass spectral database. Therapeutic Drug Monitoring. 27, 747-751 (2005).
- Bhattacharyya, S., et al. Transient protein-protein interactions perturb E. coli metabolome and cause gene dosage toxicity. Elife. 5, (2016).
- Rigaut, G., et al. A generic protein purification method for protein complex characterization and proteome exploration. Nature Biotechnology. 17 (10), 1030-1032 (1999).
Yeniden Basımlar ve İzinler
Bu JoVE makalesinin metnini veya resimlerini yeniden kullanma izni talebi
Izin talebiThis article has been published
Video Coming Soon
JoVE Hakkında
Telif Hakkı © 2020 MyJove Corporation. Tüm hakları saklıdır