
需要订阅 JoVE 才能查看此. 登录或开始免费试用。
Method Article
猪全血表达编码和非编码 rna 类的鉴定
摘要
在这里, 我们提出了一个协议优化的处理编码 (mrna) 和非编码 (ncrna) 全球减少 rna-seq 库从一个单一的全血样本。
摘要
创新且日益强大的下一代测序技术的出现为研究与所感兴趣的生物过程相关的潜在基因表达的能力开辟了新的途径。这些创新不仅使研究人员能够从编码影响细胞功能的基因的 mrna 序列中观察表达, 而且还可以观察到仍未翻译但仍具有调控功能的非编码 rna (ncRNA) 分子。尽管研究人员有能力观察 mrna 和 ncrna 的表达, 但研究的习惯是关注其中的一种。然而, 当研究对 mrna 和 ncrna 表达感兴趣时, 由于图书馆准备工作的差异, 很多时候它们使用单独的样本来检查编码或非编码 rna。这可能会导致需要更多的样品, 这可能会增加时间、消耗品和动物压力。此外, 它可能会导致研究人员决定只为一种分析准备样本, 通常是 mrna, 从而限制可以调查的生物问题的数量。然而, ncrna 跨越多个类, 具有影响 mrna 表达的调节作用。由于 ncrna 对基本的生物过程和感染过程中的紊乱很重要, 因此, 它们可能会成为有吸引力的治疗目标。这份手稿演示了一个改进的协议, 用于从一个完整的血液样本生成 mrna 和非编码 rna 表达库, 包括病毒 rna。优化该协议, 提高 rna 纯度, 增加结扎恢复甲基化 rna, 并省略大小选择, 以允许捕获更多的 rna 物种。
引言
下一代测序 (ngs) 已成为研究生物有机体基因组水平变化的有力工具。ngs 方法的样品制备可以根据生物体、组织类型以及更重要的是研究人员渴望解决的问题而有所不同。许多研究转向 ngs 作为研究健康和患病个体 1、2、3、4等国家之间基因表达差异的一种手段。测序是在整个基因组的基础上进行的, 研究人员可以在某个时间点捕获特定遗传标记的大部分 (如果不是全部的话) 基因组信息。
观察到的最常见的表达标记是信使 rna (mrna)。rna-seq 准备库的最常用的过程是通过使用一系列纯化、碎裂和结扎5、6 来优化 mrna 分子恢复的。但是, 有关如何执行协议的决定在很大程度上取决于样本类型和对所述示例提出的问题。在大多数情况下, 总 rna 被提取;然而, 并不是所有的 rRNA 分子都有兴趣, 在 mrna 表达研究等情况下, 过度丰富的 rRNA 物种, 如核糖体 rRNA (rrna) 需要删除, 以增加可检测的转录与 mrna 相关的数量。去除丰富的 rrna 分子最流行和最广泛使用的方法是减少被称为多 a 消耗7的多腺苷酸 rRNA转录。这种方法很好地用于 mrna 表达的分析, 因为它不影响 mrna 转录。然而, 在对非编码或病毒 rna 感兴趣的研究中, 多 a 消耗也会去除这些分子。
许多研究选择专注于 rna 序列库的准备, 以检查 mrna 表达 (编码) 或特定类别的小或大的非编码 rna。虽然还有其他程序8像我们这样的程序, 允许双重样品制备, 许多研究准备图书馆从单独的样本为单独的研究, 如果可用。对于我们这样的研究, 这通常需要多个血液样本来增加时间、消耗品和动物压力。我们的研究目标是能够使用动物的全血来识别和量化健康和高致病性猪繁殖和呼吸综合征病毒 (hp-prrsv) 之间表达的不同类别的 mrna 和非编码 rna挑战猪9,10 尽管只有一个全血样本 (2.5 毫升) 从每头猪。为了做到这一点, 我们需要优化典型的提取和库创建协议, 以生成适当的数据, 以便从单个样本中分析 mrna 和非编码 rna (ncrna) 表达 11.
这就需要制定一项协议, 允许 mrna 和非编码 rna 分析, 因为用于 rna 提取和库创建的现有标准试剂盒和方法主要用于 mrna, 并使用多 a 消耗步骤12。这一步将使其无法从样本中恢复非编码 rna 或病毒转录。因此, 需要一种优化的方法, 允许在不消耗样品多 a 消耗的情况下提取总 rna。本手稿中介绍的方法经过了优化, 允许将全血作为样本类型使用, 并为大小的 mrna 和 ncrna 建立测序库。该方法经过优化, 可以分析所有可检测到的非编码 rna, 并保留病毒 rna 供以后调查13。总之, 我们优化的库制备协议允许从一个完整的血液样本中调查多个 rna 分子。
使用这种方法的总体目标是开发一个过程, 允许从一个全血样本中收集非编码 rna 和 mrna。这使我们能够对我们研究中的每一种动物拥有来自单个样本9的 mrna、ncrna 和病毒 rna。这最终可以在不增加动物成本的情况下进行更多的科学发现, 并更完整地反映每个样本的表达。所述方法允许检查基因表达的调节剂, 并允许完成使用单个全血样本比较 mrna 和非编码 rna 表达的相关研究。我们的研究使用这个协议来研究基因表达的变化和可能的表观遗传调节剂在病毒感染的9周大的雄性商业猪。
研究方案
动物规程得到了国家动物疾病中心 (usda-ars-nadc) 动物护理和使用委员会的批准。
1. 猪血样本收集
- 将血液样本收集到 rna 管中。收集约2.5 毫升或更多, 如果有较大的收集管可用。
2. 猪血样的加工
- 在室温 (15-25) 下, 以 5, 020 x克离心血液管10分钟。如果在离心前在室温下处理冷冻样品孵育管至少2小时。
- 去除上清液, 在颗粒中加入8毫升的无 rnase 水。关闭并旋涡颗粒, 直到它明显溶解。在室温下以 5, 020 x克离心样品管 10分钟, 以回收颗粒。丢弃所有上清液并保存颗粒。
3. 总 rna 和小 rna (mirna 分离试剂盒) 的有机提取
- 从步骤2.2 中将300μl 的裂解结合缓冲液移入颗粒, 开始总 rna 提取。
- 涡流并将混合物转移到新的标记 1.5 ml 离心管中。从试剂盒中加入30μl 的均质添加剂。将管子旋涡, 放在冰上10分钟。
- 取出试管, 从试剂盒中加入300μl 的酸性苯酚: 氯仿试剂。涡流管混合。在室温下以 10, 000 x g 离心5分钟。
- 小心地将水相 (300-350μl) 取出到新鲜的管中。请注意下一步的卷。
4. rna 总分离程序
- 根据水回收量 (300-350μl), 将 1.25 x 体积的 100% (~ 375μl) 乙醇添加到水相。使用移液器混合样品。
- 为每个样品设置包含滤芯的新收集管。将液相 ~ 675μl 的液滴/乙醇混合物放在滤芯上。
请注意:不要一次添加 & gt; 700μl 的滤芯。为更大的容量连续申请。 - 以 10, 000 x g的速度短暂离心 (~ 15-20秒), 通过过滤器中的液体。不要旋转比这更硬。
- 丢弃流经, 如有必要, 用剩余的裂解乙醇混合物重复离心, 直到全部应用。在下一步中保留相同的滤芯和收集管。
- 将套件中的700μl 洗涤溶液1短暂地添加到滤芯和离心机 (~ 10秒) 中, 以拉穿滤清器。丢弃流经并保留相同的滤芯和收集管。
- 加入500μl 的洗涤液 2/3。离心通过滤芯提取液体。丢弃流经。重复清洗步骤。
- 在同一根管子中, 将滤芯旋转 60秒, 以去除滤芯中的任何残留液体。将滤芯转移到新鲜的收集管。
- 在滤芯中心加入100μl 的预热 (95°c) 无核酸水。在桌面离心机的最大速度下旋转约20-30。
请注意:rna 包含在洗脱液中, 现在可以在-20°c 或以下进行进一步处理或存储。未对小型 rna 进行浓缩。
5. 减少全球 (根据针对猪全血样本优化的协议)14,15
注: 减少全局是为了使图书馆不会过多填充读取映射到球蛋白基因, 这将减少可映射到其他更感兴趣的基因14,15的读取次数。
- 球蛋白还原寡核苷酸杂交
- 通过将提取的样品移入 0.2 ml 薄壁无核反应管, 并在70°c 下放置在热循环器中 2分钟, 使 rna (最大7微米体积为6微米 rna 样品) 变性。它是冰管的关键, 立即在第一归变性步骤为最佳 rna 质量。不需要 dnase 处理。
- 当管子冷却制备400μl 的10倍球蛋白还原寡核苷酸混合物时: 两个 hba 寡核苷酸中的每一个 100μl (5 '-gatcggccccctggggt-3 ', 和 5 '-tacacggggggggcaa-3 ') 在 30μm, 两个六溴代二苯寡核苷酸 (5 '-aggggactgggggggggc-3 ', 和5 '-gtgggggggctcctcct-3 '), 每次反应为 120μm, 最终浓度为 7.5μm hba寡核苷酸和 30μm hbb寡核苷酸。制备10倍寡核苷酸杂交缓冲: 100 mm Tris-HCL, ph 7.6;200毫升 kcl。
- 准备杂交混合: 6 微米 rna 样品 (最大7μl 体积), 400μl 的10x 球蛋白还原寡核苷酸混合物 (最终浓度 2x), 10x 寡核苷酸混合液 (最终浓度 1x) 为1μl。将无核酸水添加到10μl 的最终体积中。
- 将热环素设置在 70°c 5分钟, 立即冷却至 4°c, 然后进行 rnase h 消化。
- rnase h 消化
- 用 1x rnase h 缓冲液稀释 10倍 rnase h (10 u/μl) 至 1x rnase h。
注: rnase 缓冲液为 10倍, 使用前需要用 1x rnase h 缓冲液稀释至1x。 - 将 10倍 rnase 缓冲液的2μl、1μl 的 rnase 抑制剂在2μl 的 1x rnase h 中制备 rnase h 反应混合物, 将5μl 的无核水与总体积为10μl。
- 将球蛋白还原杂交样品与 rnase h 反应混合物10μl 彻底混合, 在37°c 下消化 10分钟, 冷却至4°c。
- 通过在每个样品中添加 0.μl 0.5 m edta 来停止消化, 并立即进入清理步骤。
- 用 1x rnase h 缓冲液稀释 10倍 rnase h (10 u/μl) 至 1x rnase h。
- rnase h 治疗的总 rna 清除。
- 根据制造商的说明, 使用基于硅膜的洗脱净化清理试剂盒纯化 rnase h 处理的 rna。预混缓冲液: 对于温和的洗涤缓冲液, 添加44毫升的100% 乙醇。
- 不要将样品转移到新的试管中。加入80μl 的无 rnase 水和350μl 的裂解缓冲液。在稀释后的 rna 中加入 250μl 100% 乙醇, 并通过移液良好地混合。
注: 请勿离心。立即继续执行步骤5.3.3。 - 现在, 将700μl 样品转移到放置在2毫升收集管中的洗脱滤芯中, 以收集流经。离心速度为 15秒, 规格≥8, 000x g。放弃流通。
- 重复此操作, 将洗脱滤芯放入新的 2 ml 收集管中。在滤芯和离心机中加入500μl 的温和洗涤缓冲液, 以≥8, 000 x克的速度在15个月内, 以便清洗滤芯膜。放弃流通。5.3.5 的步骤中重复使用收集管。
- 现在使用相同的样品管, 在滤芯中再加入500μl 的80% 乙醇。这一次离心管 2分钟, ≥8, 000 x克。收集洗脱自旋柱的下一步, 并丢弃流经和收集管。
- 将最后一步的洗脱滤芯放入新的 2 ml 收集管中。将盖子打开滤芯, 并以全速离心 5分钟, 使旋转柱膜干燥, 防止乙醇携带。丢弃流经和收集管。
请注意:为了避免损坏角度, 盖子指向与转子相反的方向。 - 取干燥的滤芯, 放入新的 1.5 ml 收集管中。在滤芯膜中加入14μl 的无 rnase 水, 确保将无 rnase 的水直接添加到中心。全速离心 60秒, 以清除 rna。
- 评估减少球蛋白的 rna 样本的质量 (第6步)。进行 mrna 样品制备 (步骤 7) 和小 rna 文库制备 (步骤 8)16,17。
注: 减少球的 rna 样本现在可以储存在-20°c, 但建议在-80°c 储存, 以便长期保存。
6. rna 评估
- 使用分光光度计对 rna 浓度进行定量。检查260和280纳米的波长比。~ 2 或更高的比率被认为是纯 rna 的, 较低的值表示有一定的污染。该仪器使用此比率来确定 rna 浓度为 ng/μl。
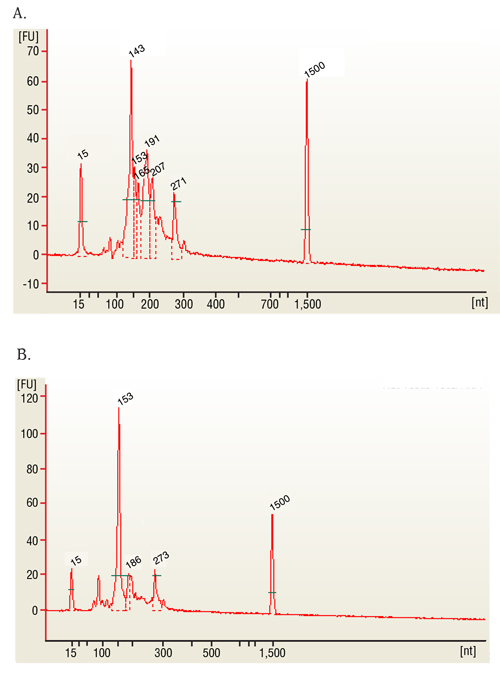
- 在适当的芯片上使用 1μl (100 纳克) 的样品来评估 rna 质量。最终产物应为 ~ 2 或更高的 rin, mrna 单读文库的峰值为 ~ 280。小 rna 文库峰值为143对应于 mirna。
7. 为 mrna 和长 ncrna 文库提供链式总 rna 样品制备。16
- 请注意:将洗脱缓冲液和 rrna 去除珠带到室温。预先标记0.2 毫升薄壁 pcr 管 (也可使用板材)。基于制造商说明的协议步骤 16。
- 从4μl 的球蛋白还原 rna 开始。加入6μl 的无核酸水, 5μl 的 rrna 结合缓冲液, 每管5μl 的 rrna 去除混合物 (1 管)。将移液器混合和重新盖上。在100°c 热盖的热囊中放置 5分钟, 在68°c 下取出并在室温下离开60秒。
- 用涡旋重新利用 rrna 去除珠;将35μl 的珠子加入新的 (第2)pcr 管和从第1个管中的移液器样品添加到第2管中的珠子上。在室温下培养 2和pcr 管3分钟。放置在磁支架上7分钟。
请注意:通过移液彻底混合每个样品, 以获得最佳的 rrna 消耗。在支架上的降管可能有助于加快分离过程。 - 将上清液从第二 pcr 管转移到匹配的第三个pcr 管, 并放置在磁支架上至少 60秒, 只有在珠子未移动到管侧的情况下, 才重复转移到新的 pcr 管中.
- 用涡流混合样品净化珠;在每一个第三pcr 管中加入99μl。允许在室温下坐 15分钟. 放置在磁架上 5分钟. 确保珠子移动到两侧。移液器丢弃上清液。
- 保持在磁性支架上设置第三管。加入200μl 的70% 乙醇, 同时注意不要挤珠。允许坐至少30秒和移液器丢弃上清液。重复步骤。
- 允许样品在室温下在磁支架上干燥15分钟。以 600 x g 的速度离心试剂盒洗脱缓冲液 5秒。取出管, 加入11μl 的洗脱缓冲液彻底混合。在长凳上生 2分钟, 然后在室温下在磁性支架上至少孵化5分钟。
- 将8.5μl 上清液从第3管转移到新的 (第4)pcr 管。从试剂盒中加入8.5μl 的 elute-primeme/frangese 高混合料。混合好。在94°c 时, 在4°c 时保持在预热盖8分钟的热环机中盖上并放置。短暂拆卸和离心机。
- 合成第一链 cdna
- 允许从试剂盒中进行第一股合成混合物, 以600xg 的温度进入室温, 在600xg 的温度下进行离心机. 将50μl 的逆转录酶转移到第一股合成混合物中。逆转录酶可以添加到第一链合成在1:9 的比例。
- 移液器的8μl 的联合逆转录酶/第一链合成混合到第四管与样品。盖和离心机 600 x g为 5 s. 放入热囊机, 预热盖设置在100°c。运行时间: 25°c 时 10分钟, 42°c 15分钟, 70°c 15分钟, 4°c 休息。最终体积约为每口井25μl。立即移动到下一步。
- 合成二阶 cdna
- 将末端修复试剂 (err) 和第二链混合 (ssm) 带到室温, 并在 600 x g 处的离心机 5 s. 在1:50 稀释时将 err 与再悬浮液混合。打开 4管的顶盖, 并在每个管中加入5μl 的稀释 err 和20μl 的 ssm;将移液器混合均匀。最终体积 ~ 50μl ds cdna。
- 在16°c 条件下, 在热环素中盖和孵育1小时。当循环完成时, 取下瓶盖, 并允许在台面上达到室温。
- ds cdna 清理步骤
- 首先将固相可逆固定化 (spri) 顺磁珠通过涡旋混合。将珠子的90μl 转移到第4管(ds cdna) 中的样品中, 然后混合。最终体积为 140μl. 允许管在室温下在磁支架上孵育 15分钟 ~ 5分钟。从每口井中去除 ~ 135μl 上清液。每口井应只剩下5μl。
- 将管子放在磁支架上, 加入200μl 的80% 乙醇进行清洗。允许坐 30秒, 然后取出并丢弃上清液。重复清洗步骤。将管子留在磁架上 15分钟, 以便干燥。
- 在室温下, 以 600 x g的速度离心再悬浮液, 为5秒。从支架上取下管子。将17.5μl 的再悬浮液转移到试管和混合中。让管子在台面上孵育 2分钟, 然后移动到磁架5分钟。
- 将含有 ds cdna 样本的上清液的移液器15μl 插入新的 (5)0.2 ml 薄壁管。迅速移动到下一步, 或密封和储存在-15°c 至-25°c 不超过7天。
- 腺苷酸 3 ' 结束。
- 将液相2.5μl 的室温再悬浮液放入样品管中, 然后加入12.5μl 的解冻 a-t着手混合料。通过移液完全混合。
注: 不使用 a 尾管控制。 - 在预热盖100°c 的热环机中盖上和孵育。在37°c 下运行 30分钟, 然后在70°c 下运行 5分钟, 盖上加热盖子。让热环器在4°c 下休息。
- 将液相2.5μl 的室温再悬浮液放入样品管中, 然后加入12.5μl 的解冻 a-t着手混合料。通过移液完全混合。
- 配体指数适配器
- 将 rna 适配器管和停止结扎缓冲液混合到室温。对于每个, 离心机在 600 x g为 5 s. 离开结扎混合在冰柜, 直到准备使用。索引的示例池排列应知道。
- 在每个样品管中加入2.5μl 的再悬浮液和2.5μl 的结扎混合物。现在移液器在2.5μl 的适当 rna 适配器到每个样品管。适配器的选择应根据制造商对所选套件的说明执行。
- 在 280 x g 的情况下, 通过离心进行重新计算和混合1分钟。在30°c 的热循环中孵育10分钟。加入5μl 的停止结扎缓冲液进行采样, 以停止反应并充分混合。
- 开始清理, 将 spri 顺磁珠子按涡旋混合 60秒. 将42μl 的珠子转移到每个管中。将长凳上的顶部彻底混合, 孵育15分钟。
- 将管子移至磁性支架, 然后离开, 直到液体变清 (~ 5分钟)。一旦清除并丢弃99.5μl 的上清液。将管子留在磁性支架上, 加入200μl 的80% 乙醇进行清洗。允许坐至少 30秒, 然后删除并丢弃上清液。重复清洗步骤。留在磁性支架上, 再留约 15分钟, 以便干燥。清洗步骤时不要打乱珠子。
- 在样品管中加入52.5μl 的再悬浮液, 搅拌至珠子完全重新悬浮。在台面上孵化2分钟。将样品管移回磁性支架, 直到液体变清 (~ 5分钟)。
- 放在支架上, 轻轻将50μl 的上清液输送到新的 (第6)管中。加入50μl 的涡旋 spri 珠。允许在台面上孵育15分钟。
- 移动到磁性支架, 让板保持直到液体变清 (~ 5分钟)。丢弃95μl 的上清液, 在每个管中留下5μl。
- 将管子留在磁性支架上, 加入200μl 的80% 乙醇进行清洗。允许管坐 30秒, 然后取出并丢弃上清液。重复清洗。让管子在磁支架上干燥15分钟。
- 加入22.5μl 的再悬浮液, 搅拌至珠子悬浮。在长凳上孵化 2分钟, 然后移动到磁性支架, 直到液体变清 (~ 5分钟)。将样品20μl 放入新的 pcr 管中的液器 20μl (第7个)。
- 将所需的套件组件带到室温下。将 5μl pcr 引物混合物和 25μl pcr 主混合物从试剂盒转移到含有指数样本的样品 (第7个pcr 管)。混合样品和盖管。
- 将管道放入带预热盖的热循环器中。运行程序: 98°c 为 30秒, 15个周期为98°c 为 10秒, 60°c 为 30秒, 72°c 为 30秒, 72°c 为 5分钟, 保持在4°c。
- 在样品管和移液器中加入50μl 的足够混合的 spri 珠子进行清洁。让反应在板凳顶部孵育15分钟。
- 将管子移动到磁性支架上, 孵育至液体变清 (~ 5分钟)。丢弃95μl 的上清液, 在每个管中留下5μl。
- 将管子留在磁性支架上, 加入200μl 的80% 乙醇进行清洗。允许坐至少 30秒, 然后取出并丢弃上清液。重复此清洗步骤。在磁性支架上停留 15分钟, 以便干燥。
- 加入32.5μl 的再悬浮液, 搅拌至珠子完全重新悬浮。在长凳上孵化 2分钟, 然后移动到磁性支架, 直到液体变清 (~ 5分钟)。将每个管的样品从一个新的 pcr 管中进入30μl。
- 使用适当的芯片重复步骤6.1 和 6.2, 评估 dna 的数量和质量。转到共用步骤 (步骤 9)。
8. 小 rna 文库制备的 snrna。17
注: 基于制造商说明的协议步骤 17。
- 开始小 rna 库准备与 ~ 220 ng-~ 1.1μg/μl 的球蛋白还原 rna 样本 (4μl)。
- 适配器结扎
- 通过将1μl 的适配器、2μl 的无核酸水和4μl 的减少球蛋白的 rna 样本彻底混合, 将 3 '-适配器连接在一起。在70°c 下孵化 2分钟, 然后立即放在冰上。
- 加入10μl 的2倍结扎缓冲液, 和3μl 的结扎酶混合, 混合和孵育 16°c 18小时。
注: 建议将样品孵育时间增加到16°c 较低温度下的 18小时, 用于对甲基化 rna 物种 (如 piwi rna) 感兴趣的研究。较长的孵育时间允许更高的结扎效率, 由于在 3 ' 适应结扎阶段的类修改。 - 在4.5 微米的无核酸酶水中加入1μl 的逆转录酶底漆, 以防止适配器二聚物形成超过 3 ' 的适应剂。
- 在75°c 时编程5分钟的预热热循环器中进行培养, 在37°c 时进行15分钟的孵化, 在25°c 时进行15分钟的孵化, 并在4°C 时保持。
- 在70°c 的热循环器中, 每个样品的预稀释 5 '-适配器的值 1μl, 时间为 2分钟, 然后立即放在冰上。
- 加入1μl 的变性 5 '-适配器, 1μl 的 10x 5 ' 结扎缓冲液和2.5μl 的 5 ' 结扎酶。混合孵育 25°c 1小时。
- cdna 合成和扩增
- 通过 5′/3′-adapter-ligated rna 30μl、8μl 第一链缓冲液、1μl rnase 抑制剂和1μl 逆转录酶进行彻底混合。将混合物在50°c 下培养 60分钟, 立即进行 pcr 扩增或加热, 在70°c 下灭活反应 15分钟, 并储存在-20°c。
- pcr 通过添加50μl 的 pcr 主混合物、2.5μl 的 rna pcr 引物、添加2.5 微米的指定 rna pcr 引物指数来扩增40μl 的反向转移酶酶反应, 并将无核酸酶引物总体积为 100μl. 通过移液混合。使用以下循环条件在热循环器中运行 pcr: 在94°c 下的初始变性 30秒;11次94°c 循环 15秒, 在62°c 退火 30秒, 在70°c 下延长 15秒;然后在70°c 下最后延长 5分钟;样品可在4°c 的时间内保持在自行车器中。
- 注: 添加索引是为了进行采样池。
- 样品清理
- 从 dna 清理试剂盒中添加500μl 的结合缓冲液 (5m gu-hcl, 30% 异丙醇) 到100μl 的 pcr 扩增样品中, 以便能够有效地结合到自旋柱膜。
注: 如果结合缓冲液的 ph 指示包含, 请检查混合物的颜色是否为黄色。 - 将样品和结合缓冲液混合到2毫升收集管内的滤芯中, 在桌面离心机中旋转 30-60, 以 17, 900 x克的速度旋转。丢弃流经。
- 用提供的 0.75 ml 套件洗涤缓冲液 (10 mm Tris-HCl ph 7.5, 80% 乙醇) 将滤芯清洗到同一收集管中, 方法是在桌面离心机中旋转30-60 秒 , 并丢弃流经。将滤芯旋转干燥, 再干60秒。
- 将滤芯放入干净的 1.5 ml 收集管中。加入30μl 洗脱缓冲液 (10 mm Tris-HCl ph 8.5), 让色谱代表 60秒, 然后在桌面离心机中旋转 60秒, 为 17, 900 x 克。
- 从 dna 清理试剂盒中添加500μl 的结合缓冲液 (5m gu-hcl, 30% 异丙醇) 到100μl 的 pcr 扩增样品中, 以便能够有效地结合到自旋柱膜。
- 使用适当的 dna 芯片重复步骤6.1 和6.2 评估样品质量。移动到池步骤 (步骤 9)。
9. 用于排序的样本池
- 池条形码和 qc 的样本, 通过设置和标记新的管 (或96孔 ncRNA 板), 以包含 mrna 或 ncRNA 样本。根据制造商的说明16、17, 将每个条形码10nm 库的13μl 传输到相应的管道 (或新板材中的井)。提交集合样本进行测序, 如果要在同一芯片上运行, 请务必选择相似的读取长度。
结果
我们研究中的代表性样本是球蛋白和核子耗尽的全血样本。该协议的代表性结果包括一个全球枯竭的库样本, 其 rna 完整性数 (rin) 高于 7 (图 1a) 和26-280 纳米浓度比率在2或以上 (图 1b 和 1c)。样品结果的验证是使用分光光度计进行的, 以给出每个文库和基于芯片的电泳的最终浓度, 给出 rin 数以及一个峰值?...
讨论
该协议中优化的第一个关键步骤包括增加的球蛋白耗尽步骤, 这使得从全血样本中获得高质量的读数成为可能。在测序研究中使用全血的最大限制之一是样本中的大量读数, 这些读数将映射到全球分子, 并减少可映射到其他感兴趣的分子的读数.因此, 在优化样本类型的协议时, 我们需要加入一个全局损耗步骤, 以确保通过测序实现尽可能高的 mrna 和非编码 rna 捕获。根据 choi等
披露声明
作者没有什么可透露的。
在本文中提及商品名称或商业产品仅用于提供特定信息, 并不意味着美国农业部的推荐或认可。美国农业部是一个机会平等的提供者和雇主。
致谢
这项工作主要得到了美国农业部 nifa afri 2013-67015-21236 的支持, 部分得到了美国农业部 nifa afri 2015-66015-23216 的支持。这项研究得到了美国能源部 (美国能源部) 之间机构间协议任命的由橡树岭科学和教育研究所 (orise) 管理的农业研究服务研究参与方案的部分支持。美国能源部) 和美国农业部。orise 由橡树 ridge 相关大学根据指定经营实体合同号管理。de-ac05-6or2310。
我们要感谢 kay faaberg 博士的 hp-prrs 感染克隆, susan Brockmeier 博士为参与实验的动物提供了帮助, sue ohlendorf 博士为编写手稿提供了秘书协助。
材料
| Name | Company | Catalog Number | Comments |
| PAXgene Tubes | PreAnalytix | 762165 | |
| Molecular Biology Grade Water | ThermoFisher | 10977-015 | |
| mirVana miRNA Isolation Kit | ThermoFisher | AM1560 | |
| Rneasy MinElute Clean Up Kit | QIAGEN | 74204 | |
| 100% Ethanol | Decon Labs, Inc. | 2716 | |
| 0.2 mL thin-walled tubes | ThermoFisher | 98010540 | |
| 1.5 mL RNase/DNase - free tubes | Any supplier | ||
| Veriti 96-well Thermocycler | ThermoFisher | 4375786R | |
| Globin Reduction Oligo (α 1) | Any supplier | Sequence GAT CTC CGA GGC TCC AGC TTA ACG GT | |
| Globin Reduction Oligo (α 2) | Any supplier | Sequence TCA ACG ATC AGG AGG TCA GGG TGC AA | |
| Globin Reduction Oligo (β 1) | Any supplier | Sequence AGG GGA ACT TAG TGG TAC TTG TGG GT | |
| Globin Reduction Oligo (β 2) | Any supplier | Sequence GGT TCA GAG GAA AAA GGG CTC CTC CT | |
| 10X Oligo Hybridization Buffer | |||
| -Tris-HCl, pH 7.6 | Fisher Scientific | BP1757-100 | |
| -KCl | Millipore Sigma | 60142-100ML-F | |
| 10X RNase H Buffer | |||
| -Tris-HCl, pH 7.6 | Fisher Scientific | BP1757-100 | |
| -DTT | ThermoFisher | Y00147 | |
| -MgCl2 | Promega | A351B | |
| -Molecular Biology Grade Water | ThermoFisher | 10977-015 | |
| RNase H | ThermoFisher | AM2292 | |
| SUPERase-IN | ThermoFisher | AM2694 | Rnase inhibitor |
| EDTA | Millipore Sigma | E7889 | |
| Microcentrifuge | Any supplier | ||
| 2100 Electrophoresis BioAnalyzer Instrument | Agilent Technologies | G2938C | |
| Agilent RNA 6000 Nano Kit | Agilent Technologies | 5067-1511 | |
| Agilent High Sensitivity DNA Kit | Agilent Technologies | 5067-4626 | |
| TruSeq Stranded Total RNA Library Prep Kit with Ribo-Zero | Illumina | RS-122-2201 | mRNA kit; Human/Mouse/Rat Set A (48 samples, 12 indexes) |
| TruSeq Stranded Total RNA Sample Preparation Guide | Illumina | Available on-line | |
| RNAClean XP Beads | BeckmanCoulter | A63987 | |
| AMPure XP Beads | BeckmanCoulter | A63880 | |
| MicroAmp Optical 8-tube Strip | ThermoFisher | N8010580 | 0.2 ml thin-walled tubes |
| MicroAmp Optical 8-tube Strip Cap | ThermoFisher | N801-0535 | |
| RNase/DNase - free reagent reservoirs | Any supplier | ||
| SuperScript II Reverse Transcriptase | ThermoFisher | 18064-014 | |
| MicroAmp Optical 96 well plates | ThermoFisher | N8010560 | These were used in place of .3mL plates as needed |
| MicroAmp Optical adhesive film | ThermoFisher | 4311971 | |
| NEBNext Multiplex Small RNA Library Prep Set for Illumina® (Set 1) | New England Biolabs | E73005 | small RNA kit |
| NEBNext Multiplex Small RNA Library Prep Set for Illumina® (Set 2) | New England Biolabs | E75805 | small RNA kit |
| QIAQuick PCR Purification Kit | QIAGEN | 28104 | |
| 96S Super Magnet Plate | ALPAQUA | A001322 |
参考文献
- Finotello, F., Di Camillo, B. Measuring differential gene expression with RNA-seq: challenges and strategies for data analysis. Briefings in Functional Genomics. 14 (2), 130-142 (2015).
- Coble, D. J., et al. RNA-seq analysis of broiler liver transcriptome reveals novel responses to high ambient temperature. BMC Genomics. 15, 1084 (2014).
- Koltes, J. E., et al. Identification of a putative quantitative trait nucleotide in guanylate binding protein 5 for host response to PRRS virus infection. BMC Genomics. 16, 412 (2015).
- Miller, L. C., et al. Comparative analysis of signature genes in PRRSV-infected porcine monocyte-derived cells to different stimuli. PLoS One. 12 (7), 0181256 (2017).
- Head, S. R., et al. Library construction for next-generation sequencing: overviews and challenges. Biotechniques. 56 (2), 61-64 (2014).
- Wang, Z., Gerstein, M., Snyder, M. RNA-Seq: a revolutionary tool for transcriptomics. Nature reviews. Genetics. 10 (1), 57-63 (2009).
- Dominic, O. N., Heike, G., Martin, S. Ribosomal RNA Depletion for Efficient Use of RNA-Seq Capacity. Current Protocols in Molecular Biology. 103 (1), 11-14 (2013).
- Fleming, D. S., Miller, L. C. Identification of small non-coding RNA classes expressed in swine whole blood during HP-PRRSV infection. Virology. 517, 56-61 (2018).
- Fleming, D. S., Miller, L. C. Small non-coding RNA expression status in animals faced with highly pathogenic porcine reproductive and respiratory syndrome virus (HP-PRRSV). Proceedings of the World Congress on Genetics Applied to Livestock Production. (11), (2018).
- Bivens Nathan, J., Zhou, M. RNA-Seq Library Construction Methods for Transcriptome Analysis. Current Protocols in Plant Biology. 1 (1), 197-215 (2016).
- Cui, P., et al. A comparison between ribo-minus RNA-sequencing and polyA-selected RNA-sequencing. Genomics. 96 (5), 259-265 (2010).
- Guo, Y., et al. RNAseq by Total RNA Library Identifies Additional RNAs Compared to Poly(A) RNA Library. BioMed Research International. 2015, 9 (2015).
- Choi, I., et al. Increasing gene discovery and coverage using RNA-seq of globin RNA reduced porcine blood samples. BMC Genomics. 15, 954 (2014).
- . TruSeq® Stranded Total RNA Sample Preparation Guide. Illumina. , (2018).
- . New England Biolabs Inc. Protocol for use with NEBNext Multiplex Small RNA Library Prep Kit for Illumina (Index Primers 1-48) (E7560) Available from: https://www.neb.com/~/media/Catalog/All-Protocols/758F75A29CDE4D03954E1EF75E78EA3D/Content/manualE7560_Figure_1.jpg (2018)
- Shin, H., et al. Variation in RNA-Seq transcriptome profiles of peripheral whole blood from healthy individuals with and without globin depletion. PLoS One. 9 (3), 91041 (2014).
- Costa, V., Angelini, C., De Feis, I., Ciccodicola, A. Uncovering the complexity of transcriptomes with RNA-Seq. Journal of Biomedicine and Biotechnology. 2010, 853916 (2010).
- Martens-Uzunova, E. S., Olvedy, M., Jenster, G. Beyond microRNA--novel RNAs derived from small non-coding RNA and their implication in cancer. Cancer Letters. 340 (2), 201-211 (2013).
{kind=link}
转载和许可
请求许可使用此 JoVE 文章的文本或图形
请求许可This article has been published
Video Coming Soon
版权所属 © 2025 MyJoVE 公司版权所有,本公司不涉及任何医疗业务和医疗服务。