
Method Article
Ein thermoplasmonischer Ansatz zur Untersuchung der Reparatur von Plasmamembranen in lebenden Zellen und Modellmembranen
In diesem Artikel
Zusammenfassung
Die thermoplasmonische Punktionsmethode integriert konfokale Mikroskopie, optische Pinzetten und Goldnanopartikel, um Proteinantworten während der Reparatur von Plasmamembranen in Zellen und riesigen unilamellären Vesikeln zu untersuchen. Die Technik ermöglicht eine schnelle und lokalisierte Membranpunktion und ermöglicht die Identifizierung von Schlüsselproteinen und ihrer funktionellen Rolle in der komplizierten Plasmamembran-Reparaturmaschinerie.
Zusammenfassung
Die Zellmembran ist entscheidend für das Überleben der Zelle, und die Gewährleistung ihrer Integrität ist von entscheidender Bedeutung, da die Zelle während ihres gesamten Lebenszyklus Verletzungen erleidet. Um eine Schädigung der Membran zu verhindern, haben Zellen effiziente Reparaturmechanismen der Plasmamembran entwickelt. Diese Reparaturmechanismen können durch die Kombination von konfokaler Mikroskopie und nanoskaliger Thermoplasmonik untersucht werden, um die Rolle von Schlüsselproteinen wie Annexinen zu identifizieren und zu untersuchen, die an der Oberflächenreparatur in lebenden Zellen und Membranmodellsystemen beteiligt sind.
Bei der Punktionsmethode wird ein Laser verwendet, um bei der Bestrahlung mit Nanopartikeln eine stark lokalisierte Erwärmung zu induzieren. Die Verwendung von Nahinfrarotlicht minimiert die Phototoxizität in der biologischen Probe, während der Großteil der Absorption im Nahinfrarot-resonanten plasmonischen Nanopartikel stattfindet. Diese thermoplasmonische Methode wurde für potenzielle photothermische und biophysikalische Forschung genutzt, um das Verständnis intrazellulärer Mechanismen und zellulärer Reaktionen durch Vesikel- und Zellfusionsstudien zu verbessern. Der Ansatz hat sich als komplementär zu bestehenden Methoden zur Membranzerstörung erwiesen, wie z. B. mechanisch, chemisch oder optisch induzierte Verletzungen, und bietet ein hohes Maß an Kontrolle durch die Zufügung extrem lokalisierter Verletzungen. Das Ausmaß der Verletzung ist auf die Nähe des sphärischen Nanopartikels beschränkt, und im Gegensatz zu gepulsten Lasern mit unterschiedlichen Wellenlängen treten entlang des Strahlengangs keine schädlichen Schäden auf. Trotz gewisser Einschränkungen, wie z.B. der Bildung von Nanobläschen, bietet die thermoplasmonische Methode ein einzigartiges Werkzeug, um zelluläre Reaktionen bei der Reparatur von Plasmamembranen in einer nahezu nativen Umgebung zu untersuchen, ohne die Lebensfähigkeit der Zellen zu beeinträchtigen.
In Kombination mit der konfokalen Mikroskopie kann die Punktionsmethode ein mechanistisches Verständnis der Membrandynamik in Modellmembransystemen sowie quantitative Informationen über Proteinreaktionen auf Membranschäden, einschließlich der Proteinrekrutierung und ihrer biophysikalischen Funktion, liefern. Insgesamt kann die Anwendung dieser Methode auf reduzierte Modellsysteme unser Verständnis der komplizierten Plasmamembran-Reparaturmaschinerie in lebenden Zellen verbessern.
Einleitung
Die Zellmembran, die sowohl als physikalische Barriere als auch als Signalplattform dient, ist für das Überleben der Zellen von entscheidender Bedeutung1. Während ihres gesamten Zellzyklus ist die Plasmamembran (PM) Schäden ausgesetzt, wie z. B. mechanischen 2,3,4,5- und chemischen 6-Stress-induzierten Verletzungen. Um die Integrität der Membran aufrechtzuerhalten und das Überleben der Zellen zu sichern, hat die Zelle robuste Mechanismen zur Reparatur der Plasmamembran (PMR) entwickelt. Diese Mechanismen hängen von verschiedenen Strategien ab, wie z. B. der Reorganisation des Zytoskeletts, der Membranfusion und den Membranersatzstrategien 7,8,9,10,11, die alle auf der Rekrutierung spezifischer Proteine beruhen. Insbesondere wurden Mitglieder der Annexin-Proteinfamilie als Schlüsselproteine identifiziert, die mit den Prozessen von PMR 1,9,12,13,14,15,16 assoziiert sind. Nach einer PM-Verletzung kommt es in der Zelle zu einem Einstrom von Kalziumionen (Ca2+), der eine unmittelbare Bedrohung für das Überleben der Zelle darstellt17. Als Reaktion auf denCa2+-Einstrom binden Annexinproteine, die sich überwiegend im Zytosol befinden, im Rahmen der PMR-Strategien an das innere Segel der geschädigten Plasmamembran18. Annexin A2 (ANXA2) war eines der ersten Mitglieder der Annexin-Familie, das mit PMR bei Dysferlin-defizienter Muskeldystrophie assoziiert wurde, und es wurde vorgeschlagen, die Reparatur durch die Fusion intrazellulärer Vesikel mit dem PM in der Nähe der Verletzungsstelle zu vermitteln 5,19,20,21. In der Folge wurden den Anhängen22 mehrere Funktionen zugeschrieben, und ihre Rolle in der PMR hat in den letzten 20 Jahren immer mehr Aufmerksamkeit erregt. Die genaue Rolle von Annexinen in der PMR ist jedoch noch nicht vollständig geklärt 15,18,21,22.
In diesem Artikel wird eine Methode zur Untersuchung der Protein-Membran-Interaktion und der Membrandynamik in einer kontrollierten und hochgradig lokalisierten Weise vorgeschlagen, wobei eine Kombination aus konfokaler Mikroskopie, optischer Pinzette und Goldnanopartikeln (AuNPs) verwendet wird. Diese Methode ermöglicht die quantitative Untersuchung von Protein-, Lipid- und niedermolekularen Wechselwirkungen als Reaktion auf Membranschäden und Ca2+ Einstrom. Trotz der Komplexität und Vielfalt der Komponenten, die am Prozess der Membranreparatur beteiligt sind, wurden vereinfachte Membransysteme, die die Plasmamembran nachahmen, eingesetzt, um ein tieferes mechanistisches Verständnis der Membrandynamik und der Reaktion von Annexinproteinen auf Membranstörungen zu erlangen16. Als Modellmembransystem wurden riesige unilamellare Lipidvesikel (GUVs) mit einer spezifizierten Lipidzusammensetzung gewählt. Die GUVs wurden mit der gelunterstützten Hydratationsmethode, insbesondere der Polyvinylalkohol-Gelhydratation, wie von Weinberger et al.23 beschrieben, erzeugt, die eine effiziente Verkapselung von Annexinen in GUVs ermöglichte.
Die Verwendung von Nahinfrarot-Laserbestrahlung (NIR) auf metallischen Nanopartikeln (NPs) induziert eine signifikante Erwärmung des NP, was sie zu einer effektiven Methode zur Etablierung einer lokalen Wärmequelle macht, die in biomedizinischen Anwendungen genutzt wird24. Die Methode wurde ursprünglich verwendet, um die Temperatur, die einen einzelnen AuNP umgibt, sowohl in 2D- als auch in 3D-biomimetischen Assays direkt zu messen. In diesen Assays 25,26 wurden die plasmonischen Nanopartikel auf einer geträgerten Lipiddoppelschicht bestrahlt oder optisch in der Nähe von GUVs gefangen, die bei lokaler Erwärmung einen lokalen thermischen Phasenübergang durchliefen, was eine Quantifizierung und Kontrolle des genauen Temperaturprofils um das Partikel herum ermöglichte. Dieses Referenztemperaturprofil wurde bei der Untersuchung oder Manipulation biologischer Proben verwendet. Weitere Fortschritte in der Methode haben die Induktion nanoskopischer Poren in Membranen27 erleichtert, was eine Vesikel- und Zellfusion ermöglicht28,29. Andere Studien haben das Verhalten von peripheren Membranproteinen in GUVs29 und Transmembranproteinen30 untersucht, indem sie neuartige Hybridvesikel geschaffen haben, während die zellspezifische Wirkstoffabgabe ebenfalls untersucht wurde, um zelluläre Reaktionen oder Genexpression zu kontrollieren und zu untersuchen 28,29,31,32,33. Kürzlich wurde die Methode verwendet, um Proteinantworten auf Membranschäden zu untersuchen 32,34,35.
Es gibt mehrere Methoden, um die Plasmamembran aufzubrechen, um zelluläre Reaktionen und die Membranreparatur zu untersuchen. Dazu gehören Mikronadelpunktionen, Mikroperlenschütteln und Zellschaben, die alle die Zellmembran mechanisch stören können 14,36,37. Chemisch induzierte Schäden können durch Zugabe von Detergenzien 5,38 oder bakteriellen Toxinen39,40 erreicht werden, die die Lipiddoppelschicht destabilisieren und Membranporen über der Plasmamembran erzeugen. Darüber hinaus wurden optisch induzierte Verletzungen durch kontinuierliche und gepulste Laser verwendet, um PMR-Komponenten, wie z. B. die Annexinproteine 5,14,21,41, in Kombination mit plasmonischen Nanopartikeln 42,43,44,45 zu untersuchen. Trotz der Effizienz von gepulsten Hochleistungslasern können sie entlang des Strahlengangs erhebliche Verletzungen und Schäden im Zellinneren verursachen. Darüber hinaus müssen die detaillierten Veränderungen, die bei gepulster Laserbestrahlung in der biologischen Materie auftreten, und ob dadurch eine gut definierte Pore entsteht, noch weiter untersucht werden. In diesem Artikel wird eine alternative Methode vorgestellt, bei der die Thermoplasmonik eingesetzt wird, um nanoskopische Löcher im Feinstaub auf kontrollierte Weise zu induzieren34,35, ohne die inneren Strukturen signifikant zu beschädigen. Dies wird erreicht, indem plasmonische NPs einem hochfokussierten NIR-Laser ausgesetzt werden, was zu einem extrem lokalisierten Temperaturanstieg führt, der leicht Temperaturen von über 200 °C erreichen kann, was zu kleinen nanoskopischen Explosionen führen kann 25,46,47. Dieser Prozess kann durch Einstellen der Laserintensität sowie der Größe, Form und Zusammensetzung der NPs48 gesteuert werden. Durch den Einsatz dieser Technik können Forscher die Rolle von Proteinen bei der PM-Reparatur in lebenden Zellen untersuchen, was dazu beitragen könnte, einige der unbeantworteten Fragen bezüglich der Beteiligung von Annexin-Proteinen an der Membranreparatur zu beantworten, ohne die Lebensfähigkeit der Zellen zu beeinträchtigen.
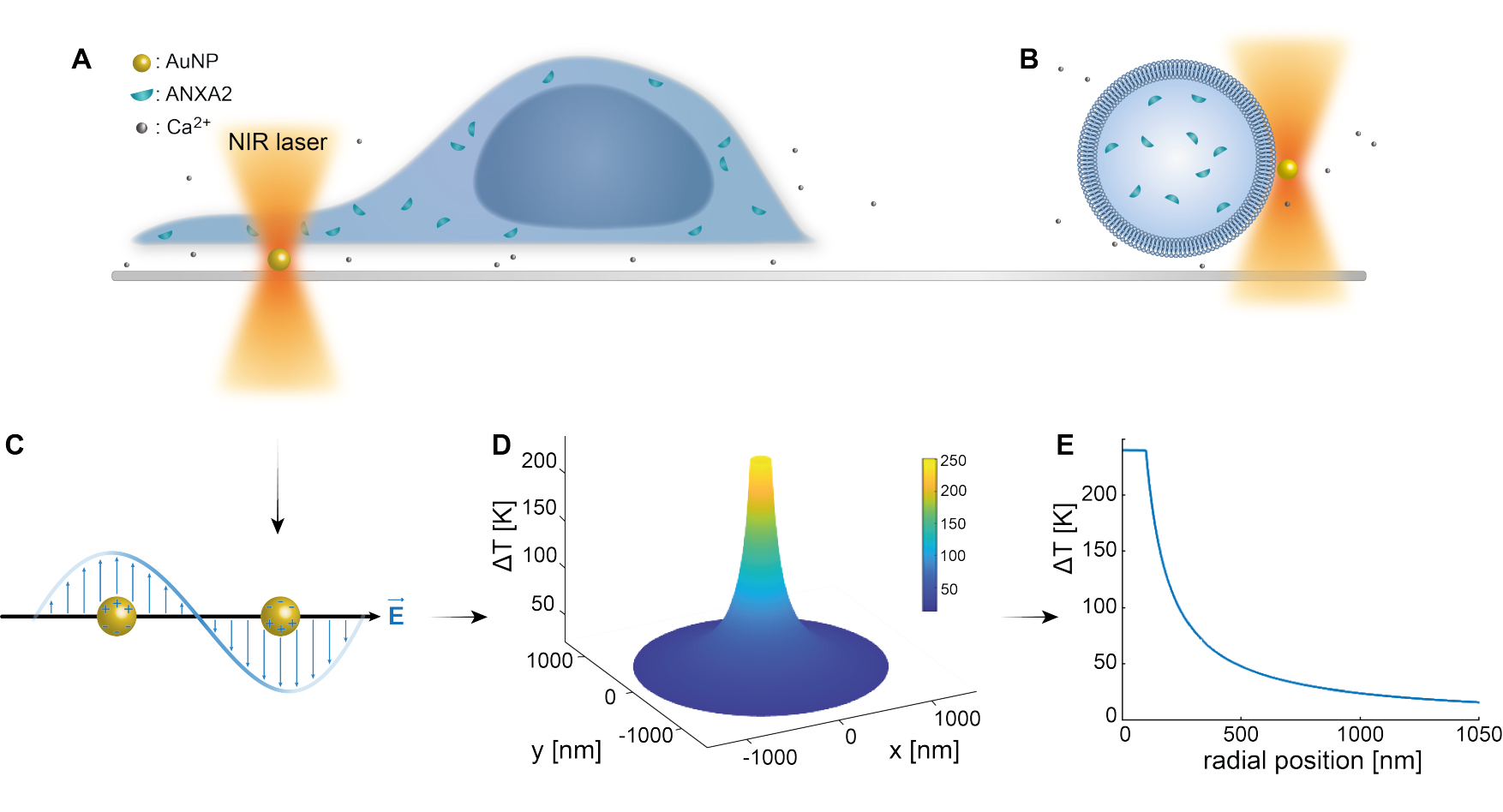
Das optische Einfangen von plasmonischen Nanopartikeln ist in früheren Studien gut etabliert 25,49,50,51,52; zusätzliche Erkenntnisse über die thermoplasmonischen Eigenschaften der Nanopartikel 53,54,55 können jedoch in den ergänzenden Materialien (Ergänzungsdatei 1) gewonnen werden. Mit der thermoplasmonischen Methode können nanoskopische Löcher im PM erzeugt werden, um die zelluläre Reaktion und Reparaturmechanismen zu untersuchen. Genauer gesagt kann die Punktion durch die optische Erwärmung von AuNPs in unmittelbarer Nähe der Membran erreicht werden, wie in den Abbildungen 1A und B gezeigt. Diese lokalisierte Punktion ermöglicht einen Ca2+ Einstrom, der durch einen Kalziumsensor verifiziert wurde, wodurch die PMR-Maschinerie aktiviert wurde. Für Lebendzellexperimente wurden AuNPs mit einem Durchmesser von 200 nm auf der Oberfläche unter der Zelle immobilisiert, um die Rolle von ANXA2 in der PMR mittels konfokaler Mikroskopie zu überwachen. Der NIR-Laser (Abbildung 1A, B) mit einer Wellenlänge von 1064 nm bestrahlt das AuNP unter Ausnutzung seiner plasmonischen Eigenschaften (Abbildung 1C), was zu einer erheblichen lokalen Erwärmung (Abbildung 1D) im biologischen Transparenzfenster49 führt, während die Zelle selbst nur minimal beschädigt wird. Der Hochtemperaturbereich, der den AuNP umgibt, nimmt in einem Abstand, der dem Radius des NP entspricht, schnell um 30-40% ab, wie in Abbildung 1E dargestellt, was eine extrem begrenzte Verletzung in allen drei Dimensionen ermöglicht.
Ergänzendes Dossier 1. Bitte klicken Sie hier, um diese Datei herunterzuladen.

Abbildung 1: Schematische Darstellung der experimentellen Methode. (A) ANXA-transfizierte Zellen befinden sich auf immobilisierten Goldnanopartikeln (AuNPs) auf der Oberfläche, oder (B) riesige unilamellare Vesikel (GUVs) mit eingekapseltem ANXA sind in einem Medium suspendiert, das AuNPs enthält. (C) Ein einzelnes AuNP wird von der optischen NIR-Falle bestrahlt, wobei die Wechselwirkung zwischen dem einfallenden elektromagnetischen Feld und den Leitungselektronen zu einer kollektiven Schwingung der Elektronen innerhalb des NP führt. (D) Dieser Prozess führt zu einem sehr begrenzten, aber signifikanten Temperaturanstieg. Um die Temperatur an der NP-Oberfläche abzuschätzen, wird die Mie-Theorie verwendet und ein (E)-Temperaturprofil für einen AuNP mit einem Durchmesser von 200 nm und einer Laserintensität I = 6,36 x 108 W/cm2 berechnet. Bitte klicken Sie hier, um eine größere Version dieser Abbildung anzuzeigen.
{kind=link}
Um den thermischen Effekt auf die Zellmembran zu minimieren, werden die AuNPs nur für ~1 Sekunde bestrahlt. Dies führt zu einem vorübergehenden und lokalen Erhitzungsschub, der die Schädigung von Proteinen reduziert, die normalerweise mehr Zeit benötigen, um sich zu entfalten. Bei der Membranpunktion werden Annexinproteine im Bruchteil einer Sekunde rekrutiert, und innerhalb weniger Sekunden bildet sich ein Annexin-Ring-ähnliches Gerüst um die Verletzungsstelle (Abbildung 2). Dieser Ansatz wurde auch angewendet, um die Beteiligung von ANXA5 sowohl in lebenden Zellen als auch in Modellmembranen16 zu untersuchen, um das vollständige Schema der Reparaturprozesse zu beleuchten. Während das Hauptaugenmerk auf der korrelierenden Rekrutierung verschiedener Annexin-Proteine lag, sind die biophysikalischen Aspekte des Reparaturmechanismus noch nicht geklärt.
Um die vorgeschlagene Methode vollständig umzusetzen, sind drei Schlüsselkomponenten erforderlich: konfokale Mikroskopie, optische Pinzette und Metallnanopartikel. Optische Pinzetten werden verwendet, um AuNPs einzufangen, und ihre Konstruktion kann nach dem von Neuman et al.49 beschriebenen Verfahren erreicht werden. Wenn sich der Bau einer optischen Pinzette jedoch als zu schwierig erweist, kann ein eng fokussierter NIR-Laser verwendet werden, um AuNPs zu bestrahlen, die unter den Zellen immobilisiert sind. Während für dieses Protokoll sphärische AuNPs ausgewählt wurden, konnte auch eine Vielzahl von plasmonischen Partikeln mit einstellbaren Absorptionsspektren verwendet werden, um einen stark lokalisierten Temperaturgradienten innerhalb des NIR-Bereichszu erreichen 48.
Die Fluoreszenzbildgebung ist notwendig, um die Rolle der fluoreszenzmarkierten Proteine zu beobachten, und daher könnte die Totalreflexionsmikroskopie (TIRF)56 als Alternative zur konfokalen Bildgebung in Betracht gezogen werden. Diese Technik erlaubt jedoch nur die Oberflächenbildgebung und wäre nicht kompatibel mit den Modell-Membranvesikel-Experimenten. Folglich sind sowohl die optische Pinzette als auch das konfokale Mikroskop für die präzise Lokalisierung des Nanopartikels und die detaillierte Untersuchung des lokalen Bereichs um die Zellverletzung unerlässlich. Um das Nanopartikel effektiv mit einem beugungsbegrenzten Laserfokus zu bestrahlen, ist es notwendig, das Nanopartikel sichtbar zu machen. Dies kann durch die Reflexionsmikroskopie, die ein Standard-Bildgebungsmerkmal der konfokalen Mikroskope von Leica ist, optimal erreicht werden. Wenn jedoch keine Reflexions- oder Streubildgebung verfügbar ist, können alternative Methoden, wie z. B. die weniger effiziente fluoreszierende AuNP-Markierung, in Betracht gezogen werden.
Zusammenfassend lässt sich sagen, dass die in dieser Studie vorgestellte hochgradig kontrollierbare und lokalisierte thermoplasmonische Methode das Potenzial hat, als hervorragende Plattform für die Untersuchung der molekularen Komponenten zu dienen, die an zellulären Reaktionen und PM-Reparaturmechanismen in lebenden Zellen beteiligt sind. Neben der Untersuchung der Proteinantwort auf PM-Schäden kann dieser Ansatz auch für die lokale Punktion von Vesikeln verwendet werden, wodurch eine Untersuchung der Proteinantwort sowohl in der Protein-Protein- als auch in der Protein-Membran-Dynamik ermöglicht wird. Darüber hinaus ermöglicht diese Methode eine quantitative Analyse der Wechselwirkungen zwischen Proteinen, Lipiden und kleinen Molekülen, wenn Membranen aufgebrochen werden. Zusammengenommen haben diese Fortschritte das Potenzial, einige der ungelösten Fragen in Bezug auf die komplizierte und komplexe Reparaturmaschinerie von Plasmamembranen zu beleuchten.
Protokoll
1. Vorbereitung der Zellmembranpunktion
- Zellaussaat (Tag 1)
- Kultivieren Sie menschliche embryonale Nierenzellen (HEK293T) in einem Kulturmedium bei 37 °C in einem 5%igen CO2 - Befeuchter-Inkubator, bis sie eine Konfluenz von 70 % erreichen.
- Lösen Sie die Zellen mit 500 μl Trypsin von der Oberfläche, zählen Sie 200.000 HEK293T Zellen und säen Sie sie in einer Kulturschale mit einem Gesamtvolumen von 3 ml Kulturmedium. Inkubieren Sie die Zellen bei 37 °C in einem 5%igen CO2 - Befeuchter-Inkubator für 24 Stunden.
HINWEIS: Um eine Zellanhäufung in der Mitte der Schale zu verhindern, verteilen Sie die Zellen gleichmäßig und vermeiden Sie es, die Schale zu verwirbeln, da dies die Transfektionseffizienz beeinträchtigen kann.
- Zelltransfektion (Tag 2)
HINWEIS: Die transfizierten Zellen können bis zu 48 Stunden nach der Transfektion verwendet werden.- Das interessierende Plasmid und das Transfektionsreagenz werden vor Gebrauch 5 s lang pipetiert.
- Mischen Sie in einem sterilen 2-ml-Röhrchen Folgendes in der angegebenen Reihenfolge: 500 μl reduziertes Serummedium, 5 μl Transfektionsreagenz (4-mal mehr als das Plasmid) und 1,25 μl Plasmid (1 μg/μl).
HINWEIS: Um den Kalziumeinstrom nach einem Membranriss zu untersuchen, gehen Sie auf die gleiche Weise vor, verwenden Sie jedoch die membrangebundene Kalziumsensorsonde GCaMP6-CAAX (1 μg/μl). - Die Transfektionsmischung wird vorsichtig, aber gründlich pipeppetiert und 30 Minuten lang bei Raumtemperatur (RT) inkubiert, bevor sie tropfenweise in die Zellen gegeben wird.
- Bevor Sie die Transfektionsmischung hinzufügen, nehmen Sie das Medium aus der Kulturschale, waschen Sie die Zellen vorsichtig mit 2 ml phosphatgepufferter Kochsalzlösung und geben Sie 2000 μl reduziertes Serummedium in die Schale.
- Die Zellen werden zusammen mit dem Transfektionsgemisch bei 37 °C in einem 5%igen CO2-Befeuchter-Inkubator für 2 h und 45 min inkubiert, bevor das Medium auf 3 ml Nährmedium umgestellt wird.
- Herstellung einer Gold-Nanopartikel-Lösung (AuNP) (Tag 2)
- Die 200 nm AuNP-Stammlösung wird 10 s lang auf Stufe 10 vorgetakt (siehe Materialtabelle für weitere Spezifizierungen der Apparatur), 5 min lang beschallt (maximale Amplitude) und erneut 10 s lang vortexed.
- Mischen Sie 150 μl AuNPs mit 850 μl destilliertem Wasser zu einem Gesamtvolumen von 1 ml.
HINWEIS: Die verdünnte AuNP-Lösung kann im Kühlschrank aufbewahrt und bis zu 1 Monat wiederverwendet werden.
- Zubereitung der Versuchsschale (Tag 2)
- Die Mikrotiterschale wird mit 150 μl 0,01%-0,1%iger Zellanhaftungslösung bestreicht und 15 min bei RT inkubiert.
- Waschen Sie die Glasoberfläche zweimal mit 500 μl destilliertem Wasser und lassen Sie sie ~10 Minuten an der Luft trocknen.
- 80 μl AuNP-Lösung tropfenweise auf die trockene Oberfläche geben.
HINWEIS: Vortex (10 s), Beschallung (5 min) und Vortex (10 s) der verdünnten AuNP-Lösung, bevor sie auf die beschichtete Glasoberfläche gegeben wird, um AuNP-Aggregate zu minimieren. - Warten Sie ~10 Minuten, bevor Sie 1,5 ml Nährmedium einführen. Das Gericht über Nacht bei 37 °C inkubieren lassen.
- Vorbereitung der Experimentierkammer (Tag 3)
HINWEIS: Die Versuchskammer kann entweder am 3. oder 4. Tag vorbereitet werden. Achten Sie jedoch darauf, dass die folgenden Vorbereitungen am selben Tag wie das Experiment durchgeführt werden.- Entfernen Sie das Medium aus den Zellen in der Kulturschale und waschen Sie die Zellen mit 2 ml phosphatgepufferter Kochsalzlösung.
HINWEIS: Dieser Schritt ist wichtig, um alle Restmedien und Ablagerungen zu entfernen, die die nachfolgenden Schritte beeinträchtigen könnten. - Geben Sie 500 μl enzymbasierte Zellablösungslösung in die Vertiefung und inkubieren Sie 1-3 Minuten lang, bis sich die Zellen von der Kulturschale gelöst haben.
- Fügen Sie 1,5 ml frisches Kulturmedium hinzu und pipettieren Sie die Zelllösung, um eine homogene Zelllösung zu erhalten, die die Wahrscheinlichkeit von Zellclustern minimiert.
- Entfernen Sie das Medium vorsichtig aus den AuNPs im experimentellen Mikrotopf.
- Geben Sie die Zelllösung (2 ml) in die Mikrovertiefung und lassen Sie sie mindestens 5 Stunden inkubieren, bevor Sie das Experiment durchführen.
HINWEIS: Um optimale Versuchsbedingungen zu gewährleisten, vermeiden Sie es, die Kammer zu verwirbeln, da dies dazu führen kann, dass sich Zellen in der Mitte der Kammer ansammeln.
- Entfernen Sie das Medium aus den Zellen in der Kulturschale und waschen Sie die Zellen mit 2 ml phosphatgepufferter Kochsalzlösung.
2. Experiment zur Punktion von Zellmembranen
- Optische Einstellungen des Experiments
- Führen Sie die Experimente mit einem konfokalen Rastermikroskop in Kombination mit einem 1064-nm-Fallenlaser57 durch.
- Führen Sie die optische Einfangung in der Fokusebene mit einem 63-fachen Wasserimmersionsobjektiv mit einer numerischen Apertur (NA) von 1,2 durch.
- Nehmen wir an, dass der Fokus die Größe einer Airy-Scheibe hat und dass die fokale Strahlbreite des Bestrahlungslasers einen Radius von ~540 nm beträgt.
- Rechnen Sie die Laserleistung (P) in die entsprechende Laserintensität (I) um, indem Sie die Laserleistung pro Fläche (W/cm2) berechnen.
- Verwenden Sie einen akusto-optischen Strahlteiler (AOBS), um mehrere Fluoreszenzsignale zu visualisieren, die mit Photomultiplier-Röhren detektiert werden, und gleichzeitig metallische NPs über ihr Streusignal zu detektieren.
HINWEIS: Nicht alle konfokalen Systeme sind mit AOBS ausgestattet, das eine Reflexionsbildgebung von metallischen NPs ermöglicht. Hier müssen andere Detektionsformen implementiert werden, oder es kann versucht werden, den NIR-Laser im Bereich unterhalb der Zelle sequentiell abzutasten. - Montieren Sie während der experimentellen Sitzungen eine offene Glasbodenkammer mit Zellen, molekularen Fluoreszenzsonden und Goldnanopartikeln auf dem Mikroskop.
- Verschieben Sie die Falle relativ zu den Zellen, indem Sie die auf einem piezoelektrischen Tisch montierte Probe verschieben, um nanometergenaue laterale Bewegungen zu ermöglichen16.
HINWEIS: Die optische Falle wird stationär gehalten.
- Experimentelle Einstellungen für die Zellmembranpunktion
- Platzieren Sie HEK293T Zellen, transfiziert mit Annexin-Plasmiden, die mit einem fluoreszierenden Protein gekoppelt sind, auf isolierten AuNPs, die auf der Glasoberfläche immobilisiert sind (Abbildung 1A).
- Verwenden Sie einen Argon 488-nm-Laser zur Visualisierung des GFP-Fluoreszenzsignals und einen HeNe-633-nm-Laser zur Beobachtung der AuNP-Reflexion im konfokalen Rastermikroskop.
- Verwenden Sie die optische Pinzette, die mit einem 1064-nm-NIR-Laser arbeitet, um ein einzelnes AuNP ~1 s lang zu bestrahlen, wodurch ein signifikanter lokaler Temperaturanstieg ausgelöst wird, der die Plasmamembran stört.
- Wenden Sie eine Bestrahlung zwischen 200 und 295 mW auf das fokussierte Partikel an, was zu einem erheblichen Temperaturanstieg führt.
HINWEIS: Es gibt einen erheblichen Leistungsverlust innerhalb der Optik, wobei die Laserleistung im Brennpunkt je nach Aufbau etwa 20 % des angegebenen Milliwatts erreicht. Die spezifische Intensität (gemessen in W/cm2) hängt von der exakten Ausrichtung des Systems ab, insbesondere von der Brennweite. Darüber hinaus ist die Wärmeleitung von Glas höher als die von Zelle und Wasser, und folglich wird die Verringerung der an die Plasmamembran abgegebenen Wärmemenge geringfügig reduziert48,58. - Erzielen Sie eine effektive und lokalisierte PM-Verletzung und eine anschließende Proteinreparaturreaktion, indem Sie die Lokalisierung des NIR-Laser-Fokuspunkts richtig kalibrieren. Dies wird erreicht, indem ein einzelner AuNP erfasst wird, der im selben Bildgebungsmedium suspendiert ist, und sichergestellt wird, dass der ausgewählte AuNP vor der Bestrahlung scharf gestellt wird.
ANMERKUNG: Das Nanopartikel gilt als scharf, wenn sein Streusignal am schärfsten erscheint, d. h. klare Kanten und das Fehlen eines Halo um das Partikel herum aufweist, wenn es in der konfokalen Mikroskopie beobachtet wird (Abbildung 2C (ii)).
- Zell- und AuNP-Dichtebedingungen
- Wählen Sie einzelne Zellen anstelle von Zellclustern, um eine Überlappung der Plasmamembranen zu vermeiden.
ANMERKUNG: Die Zellen sollten an der Oberfläche haften und eine abgeflachte Morphologie aufweisen (Ergänzende Abbildung 1), die eine Punktion der Zellperipherie ermöglicht und gleichzeitig eine Schädigung der Kernmembran verhindert (Ergänzende Abbildung 2). - Inkubieren Sie die Zellen gemäß dem Protokoll oder bis sie sich beruhigt und abgeflacht haben, um die Aufnahme von AuNP-Zellen zu verhindern. Vermeiden Sie eine übermäßige Inkubationszeit und verringern Sie die Wahrscheinlichkeit einer AuNP-Endozytose durch PEGylierte AuNPs.
- Stellen Sie sicher, dass die immobilisierten AuNPs als einzelne Partikel in ausreichendem Abstand zueinander vorhanden sind, um Aggregate zu vermeiden. Aggregate können zu einer deutlichen Erhöhung des thermischen Gradienten führen, was zu erhöhten Temperaturen führt, die einen erheblichen Teil der Zelle stören können.
- Tauschen Sie die Probe alle 1-2 Stunden aus, um die Zellgesundheit zu erhalten.
HINWEIS: Eine längere Exposition kann zu einer Verschlechterung der Zellgesundheit führen und ihre Fähigkeit beeinträchtigen, in Bezug auf die Membranreparatur genau auf Verletzungen zu reagieren. Diese Verschlechterung der Zellgesundheit wird typischerweise durch eine Veränderung der Zellform beobachtet, da die Zellen kugelförmiger und starrer erscheinen, was schließlich zum Zelltod führt.
- Wählen Sie einzelne Zellen anstelle von Zellclustern, um eine Überlappung der Plasmamembranen zu vermeiden.
Ergänzende Informationen. Bitte klicken Sie hier, um diese Datei herunterzuladen.
3. Vorbereitung des Riesen-Unilamellären Vesikels (GUV)
- Herstellung der Lipidmischung
- Stellen Sie die GUV-Lipidzusammensetzung her, indem Sie 1,2-Dioleoyl-sn-glycero-3-phosphocholin (DOPC) und 1,2-dioleoyl-sn-glycero-3-phospho-L-serin (DOPS)-Lipide in einem molaren Verhältnis von 4:1 kombinieren (siehe Tabelle 1). Aliquotieren Sie den Lipidstock je nach Versuchsbedarf in 1,5-ml-Glasfläschchen und lagern Sie sie bei -20 °C.
HINWEIS: Für eine längere Lipidkonservierung und um die Oxidation ungesättigter Lipide zu verhindern, ersetzen Sie die Luft in den aliquotierten Durchstechflaschen durch Argon. - Bevor Sie die Lipide mischen, reinigen Sie eine 50-μl- und eine 500-μl-Glas- oder Metallspritze fünfmal gründlich mit Chloroform, um sicherzustellen, dass sie frei von Verunreinigungen für die in Chloroform gelösten Lipide sind.
VORSICHT: Chloroform aufgrund seiner Toxizität in einem Abzug handhaben.- Das berechnete Chloroformvolumen wird in eine saubere Durchstechflasche aus Braunglas überführt, gefolgt von der angegebenen Menge jedes Lipids (siehe Tabelle 1).
HINWEIS: Um eine Kreuzkontamination zwischen den Lipidvorräten zu vermeiden, stellen Sie sicher, dass die Spritzen mit Chloroform gereinigt werden. - Zum Schluss fügen Sie den Membranfarbstoff hinzu und mischen die Lipide gründlich durch Pipettieren. Lagern Sie die vorbereitete Lipidmischung zur weiteren Verwendung bei -20 °C; Das Gemisch bleibt 2-3 Wochen lebensfähig, ohne nennenswerte Lipidschäden zu verursachen.
HINWEIS: Das Mischen sollte mit einer Metall- oder Glasspritze erfolgen. Bewahren Sie die Lipide immer auf Eis auf, wenn sie aus dem Gefrierschrank kommen.
- Das berechnete Chloroformvolumen wird in eine saubere Durchstechflasche aus Braunglas überführt, gefolgt von der angegebenen Menge jedes Lipids (siehe Tabelle 1).
- Stellen Sie die GUV-Lipidzusammensetzung her, indem Sie 1,2-Dioleoyl-sn-glycero-3-phosphocholin (DOPC) und 1,2-dioleoyl-sn-glycero-3-phospho-L-serin (DOPS)-Lipide in einem molaren Verhältnis von 4:1 kombinieren (siehe Tabelle 1). Aliquotieren Sie den Lipidstock je nach Versuchsbedarf in 1,5-ml-Glasfläschchen und lagern Sie sie bei -20 °C.
- Herstellung von Polyvinylalkohol (PVA)-Gel
- Bereiten Sie die GUVs mit der von Weinberger et al.23beschriebenen gelunterstützten Hydratationsmethode mit geringfügigen Modifikationen vor.
- Bereiten Sie das PVA-Gel vor, indem Sie 5 g PVA in 100 ml eines Puffers auflösen, der 50 mM Saccharose, 25 mM NaCl und 25 mM Tris (pH 7,4) enthält.
- Erhitzen Sie die PVA-Lösung auf 85 °C und rühren Sie, bis sie durchsichtig wird. Lassen Sie es auf RT abkühlen und bewahren Sie es zur weiteren Verwendung im Kühlschrank auf.
HINWEIS: PVA ist im Puffer nicht richtig gelöst, daher ist eine Erwärmung auf bis zu 85 °C erforderlich.
- Bereiten Sie die GUVs mit der von Weinberger et al.23beschriebenen gelunterstützten Hydratationsmethode mit geringfügigen Modifikationen vor.
- Präparation von Objektträgern
- Objektträger mit Ethanol reinigen und an der Luft trocknen lassen. Behandeln Sie die Objektträger anschließend mit einem Luft-Plasma-Reiniger, um Restverunreinigungen von der Glasoberfläche zu entfernen.
- Präparation von PVA-beschichteten Objektträgern
- Erwärmen Sie das PVA-Gel (5%) 30 Minuten lang auf 60 °C, um seine Fließfähigkeit zu erhöhen. 90 μl des warmen PVA auf den Objektträger geben, gleichmäßig verteilen und in einem Wärmeschrank bei 50 °C 50 min trocknen lassen.
- Sobald der PVA-Objektträger fertig ist, fügen Sie 30 μl der vorbereiteten Lipidmischung mit einer Glas- oder Metallspritze hinzu und verteilen Sie sie mit dem Rand der Nadel zu einem dünnen Film.
- Trocknen Sie das Lipidgemisch, indem Sie den Chloroformgehalt unter leichtem Stickstoffdruck verdampfen. Trocknen Sie die Objektträger 1,5-2 h lang unter Vakuum.
- Züchtung von GUVs in einer Kammer
- Montieren Sie die hauseigene Kammer, die in ihrem Design einem zuvor veröffentlichten Bericht59 ähnelt, mit dem vorbereiteten Glasobjektträger.
- In einem separaten 2-ml-Röhrchen wird das rekombinante Protein von Interesse (in diesem Fall ANXA5 oder ANXA4) auf eine Endkonzentration von 500 nM mit einem Wachstumspuffer (GB) verdünnt, der aus 80 mM Saccharose, 70 mM NaCl und 25 mM Tris-HCl (pH 7,4) besteht.
- Geben Sie 400 μl der verdünnten rekombinanten Proteinlösung in die Kammer. Inkubieren Sie die Kammer bei RT für 1 h, damit GUVs aus der abgeschiedenen Lipidschicht wachsen können. Verwenden Sie den gleichen Puffer, ohne das Protein, als Negativkontrolle.
HINWEIS: Wickeln Sie die Kammer in Polyolefinfolie ein, um eine Pufferverdunstung zu verhindern. - Sammeln Sie die GUVs, indem Sie 400 μl des Kammerinhalts in ein 2-ml-Röhrchen umfüllen.
- Entfernen Sie nicht verkapselte Proteine außerhalb der GUVs, indem Sie 1 ml Beobachtungspuffer (OB) mit 55 mM Glukose, 70 mM NaCl und 50 mM Tris-HCl (pH 7,4) zur gesammelten Lösung hinzufügen. Anschließend wird die Lösung bei 600 x g 10 min bei 13 °C zentrifugiert.
- Nach der Zentrifugation wird 1 ml des Überstandes durch ein gleiches Volumen Beobachtungspuffer ersetzt. Dispergieren Sie die GUVs durch sanftes Pipettieren und lagern Sie sie im Kühlschrank, bis sie in GUV-Experimenten verwendet werden.
4. GUV-Punktions-Experiment
- Vorbereitung der Kammer
- Verwenden Sie für das Experiment eine 35-mm-Glasbodenschale. Um zu verhindern, dass GUVs an der Oberfläche haften bleiben und platzen, beschichten Sie die Oberfläche 15-30 Minuten lang mit β-Casein (5 mg/ml).
- Für die β-Casein-Lösung werden 0,1 g des Proteins in einem 20-ml-Puffer aus 20 mM Tris (PH 7,5) und 100 mM NaCl gelöst. Filtern Sie die Proteinlösung, aliquotieren Sie sie in kleine Fläschchen und frieren Sie sie für die weitere Verwendung ein.
- Waschen Sie die Kammer zweimal mit Beobachtungspuffer, um überschüssige freie β-Kaseine von der Oberfläche zu entfernen, und lassen Sie sie bei RT trocknen.
- In einem separaten 2-ml-Röhrchen werden die gesammelten GUVs mit OB gemischt. Geben Sie CaCl2 in die Mischung, um die gewünschte Endkonzentration (in diesem Fall 200 μM) zu erreichen.
- 150-nm-Gold-Nanoschalen (AuNSs) in einem Verhältnis von 1:100 in die Mischung geben. Die endgültige Mischung besteht aus 250 μl GUVs, 225 μl OB, 20 μl CaCl2 (5 mM) und 5 μl der spezifizierten AuNSs.
- Verwenden Sie für das Experiment eine 35-mm-Glasbodenschale. Um zu verhindern, dass GUVs an der Oberfläche haften bleiben und platzen, beschichten Sie die Oberfläche 15-30 Minuten lang mit β-Casein (5 mg/ml).
- Versuchsaufbau
- Geben Sie die Mischung in die Kammer und montieren Sie sie auf dem Mikroskoptisch. Je nach Größe der Kammer entweder die gesamte Mischung oder einen Teil der Mischung hinzufügen.
HINWEIS: Das Timing ist entscheidend, da Kalziumionen die Membran passieren und die Bindung der Annexine an das innere Segel der Membran vermitteln können. - Verwenden Sie den gleichen optischen Aufbau, der für die Zellpunktionsexperimente verwendet wurde.
- Verwenden Sie die optische Pinzette, um einen einzelnen AuNS in unmittelbarer Nähe oder auf der Oberfläche eines GUV einzufangen, indem Sie eine Laserleistung von ~ 125 mW anwenden.
- Erhöhen Sie anschließend die Laserleistung auf ~ 300 mW. Dies führt zu einem stark lokalisierten Temperaturanstieg, der die Membran an der Zielstelle stört und durchsticht.
HINWEIS: AuNSs werden für die GUV-Experimente bevorzugt, da sie in der Lage sind, einen höheren transienten Temperaturanstieg zu erzeugen und gleichzeitig eine geringere Größe im Vergleich zu festen AuNPs beizubehalten.
- Geben Sie die Mischung in die Kammer und montieren Sie sie auf dem Mikroskoptisch. Je nach Größe der Kammer entweder die gesamte Mischung oder einen Teil der Mischung hinzufügen.

Tabelle 1: Tabelle zur Bestimmung der GUV-Zusammensetzung. Bitte klicken Sie hier, um diese Tabelle herunterzuladen.
5. Messungen und Datenanalyse der ANXA-Antwort in Zellpunktionsexperimenten
- Verwenden Sie MATLAB, um die Bilder zu analysieren und das AuNP-Temperaturprofil (Abbildung 1D, E) basierend auf den Gleichungen60,61 von Mie zu berechnen und darzustellen.
- Verarbeiten Sie außerdem alle konfokalen Bilder mit der FIJI ImageJ-Verteilung62,63.
- Berechnung des ANXA-Ringradius
- Schneiden Sie den interessierenden Bereich mit dem punktierten Bereich aus den Rohdaten vor der Analyse der Membranverletzung zu.
- Nutzen Sie den hauseigenen MATLAB-Workflow für die Verarbeitung jedes Bildes, indem Sie die Mitte des ANXA-Rings und seinen äußeren Umfang manuell markieren.
- Verwenden Sie diese Markierungen, um die Verarbeitungsgrenzwerte für den Interessenbereich festzulegen.
HINWEIS: Die Fläche wird anschließend radial innerhalb der eingestellten Grenzen bearbeitet, und die Fluoreszenzintensität in einem bestimmten Abstand zum Mittelpunkt ist der Mittelwert über den Vollkreis mit gleichem Achsabstand. Diese Nummer wird für jeden Radiusschritt gespeichert. - Verwenden Sie den Arbeitsablauf, um die gemittelten Intensitäten über den gesamten Scanbereich an eine eindimensionale Gaußsche Kurve anzupassen, um die Maxima (Rp) und die volle Breite bei halbem Maximum (FWHM) zu identifizieren, die Rext bzw. Rint entsprechen, wie in Abbildung 3A dargestellt.
- Verwenden Sie diese Markierungen, um die Verarbeitungsgrenzwerte für den Interessenbereich festzulegen.
- Bestimmen Sie schließlich den Radius des ANXA-Rings durch den Abstand vom Mittelpunkt des Rings zu Rext , der durch das vom MATLAB-Workflow generierte Diagramm gegeben ist.
- Dreidimensionale ANXA-Ringradiusberechnung
- Verfolgen Sie die Änderung des ANXA-Ringradius in z-Richtung mit demselben MATLAB-Analyse-Workflow wie in Schritt 5.1. Wenden Sie den Arbeitsablauf auf mehrere konfokale Z-Schnitte entlang der Membranwunde an, wie in Abbildung 3C, E dargestellt.
- Berechnen Sie die Steigung für jede Wunde basierend auf der Änderung der Radien über den Z-Abschnitten, wie in Abbildung 3F dargestellt.
- Zeitliche Entwicklung des ANXA-Ringradius
- In ähnlicher Weise können Sie die Entwicklung des ANXA-Rings des ANXA-Rings nach einer thermoplasmonischen Membranpunktion mithilfe des MATLAB-Analyse-Workflows aus Schritt 5.1 verfolgen. Analysieren Sie aufeinanderfolgende Zeitpunkte, um Änderungen im Zeitverlauf zu überwachen, wie in Abbildung 3D, G, H dargestellt.
Ergebnisse
In der vorliegenden Arbeit wird die thermoplasmonische Methode verwendet, um die Antwort des Annexin-Proteins auf den Aufschluss der Plasmamembran zu untersuchen; Jedes Protein, das bei einer Membranverletzung rekrutiert werden kann, kann jedoch mit diesem Assay untersucht werden, vorausgesetzt, das jeweilige Protein ist fluoreszenzmarkiert. Die Rekrutierung und Funktion von Proteinen wird durch konfokale Bildgebung sowohl in humanen embryonalen Nierenzellen (HEK293T) als auch in riesigen unilamellären Vesikeln (GUVs) überwacht. Abbildung 2 veranschaulicht die experimentellen Bedingungen, unter denen ein fokussierter NIR-Laserstrahl bei 1064 nm verwendet wird, um ein einzelnes AuNP zu bestrahlen (Abbildung 2A), was zu einer Membranverletzung und einem Einstrom von Ca2+ in die Zelle führt, wodurch die PMR-Maschinerie aktiviert wird. Anschließend werden Annexine schnell an die Verletzungsstelle rekrutiert, wo sie an die negativ geladenen Phospholipide am Wundrand binden und innerhalb von Sekunden eine ringförmige Struktur bilden (Abbildung 2B, i-iv). Die Modellmembranexperimente mit GUVs zeigten, dass Membranpunktionen in Abwesenheit von ANXA schnell wieder verschlossen wurden, wie in Abbildung 2C dargestellt. In Gegenwart von ANXA wurde jedoch nach der Membranpunktion eine schnelle Akkumulation von ANXA an der Verletzungsstelle beobachtet (Abbildung 2D). Bemerkenswert ist, dass ANXA die freiliegenden Kanten weiter rollte, was letztendlich zum Platzen des GUV führte. Es wird angenommen, dass dieser Rollmechanismus auf die Fähigkeit von ANXA zurückzuführen ist, Krümmungen zu induzieren und Lipidmembranen zu biegen20.

Abbildung 2: Reaktion von Annexinen (ANXAs) auf thermoplasmonisch induzierten Membranaufschluss. Zunächst (A) fungiert die Plasmamembran als Barriere zwischen der extrazellulären Umgebung, die einen hohen Gehalt an Ca2+-Ionen enthält, und der intrazellulären Umgebung mit eingekapselten ANXAs. Bei der Bestrahlung mit einem Nahinfrarotlaser (NIR) erzeugt der AuNP erhebliche Wärme, wodurch die Membran reißt und Ca2+ Ionen einströmen. Folglich wird die Plasmamembranreparatur-Maschinerie (PMR) aktiviert, die die Rekrutierung von ANXAs an die Verletzungsstelle beinhaltet, wo sie an negativ geladene Phospholipide binden. (B-D) Konfokalmikroskopische Aufnahmen einer ANXA2-haltigen Zelle und eines ANXA4-haltigen GUV demonstrieren diesen Prozess. (i) Vor der Bestrahlung zeigen die Bilder eine intakte Zelle oder GUV, wobei die Bestrahlungsstellen durch die weißen Pfeile gekennzeichnet sind. (ii) Bei Bestrahlung mit Nanopartikeln (B) werden ANXAs schnell an die Verletzungsstelle rekrutiert und bilden eine ringförmige Struktur um die Membranwunde (B [ii-iv]). Tafel (C) zeigt ein membrangefärbtes GUV ohne ANXA, das nach Punktion ohne beobachtbares Membranremodeling schnell wieder abdichtet. Auf der anderen Seite zeigt Bild (D) ein GUV, das rekombinantes ANXA4 enthält, das bereits vor der (i) Bestrahlung aufgrund vonCa2+-Leckagen über die Membran an die Membran gebunden ist. (ii) Bei der Punktion bindet ANXA4 an die freien Kanten, wodurch das GUV kollabiert, wenn die Membran von der Kante wegrollt. Die Maßstabsleisten messen 10 μm für Abbildung (B), 2 μm für B (i), 10 μm für (C) und 15 μm für (D). Diese Abbildung ist reproduziert von Moreno-Pescador et. al16 mit Genehmigung der Royal Society of Chemistry. Bitte klicken Sie hier, um eine größere Version dieser Abbildung anzuzeigen.
{kind=link}
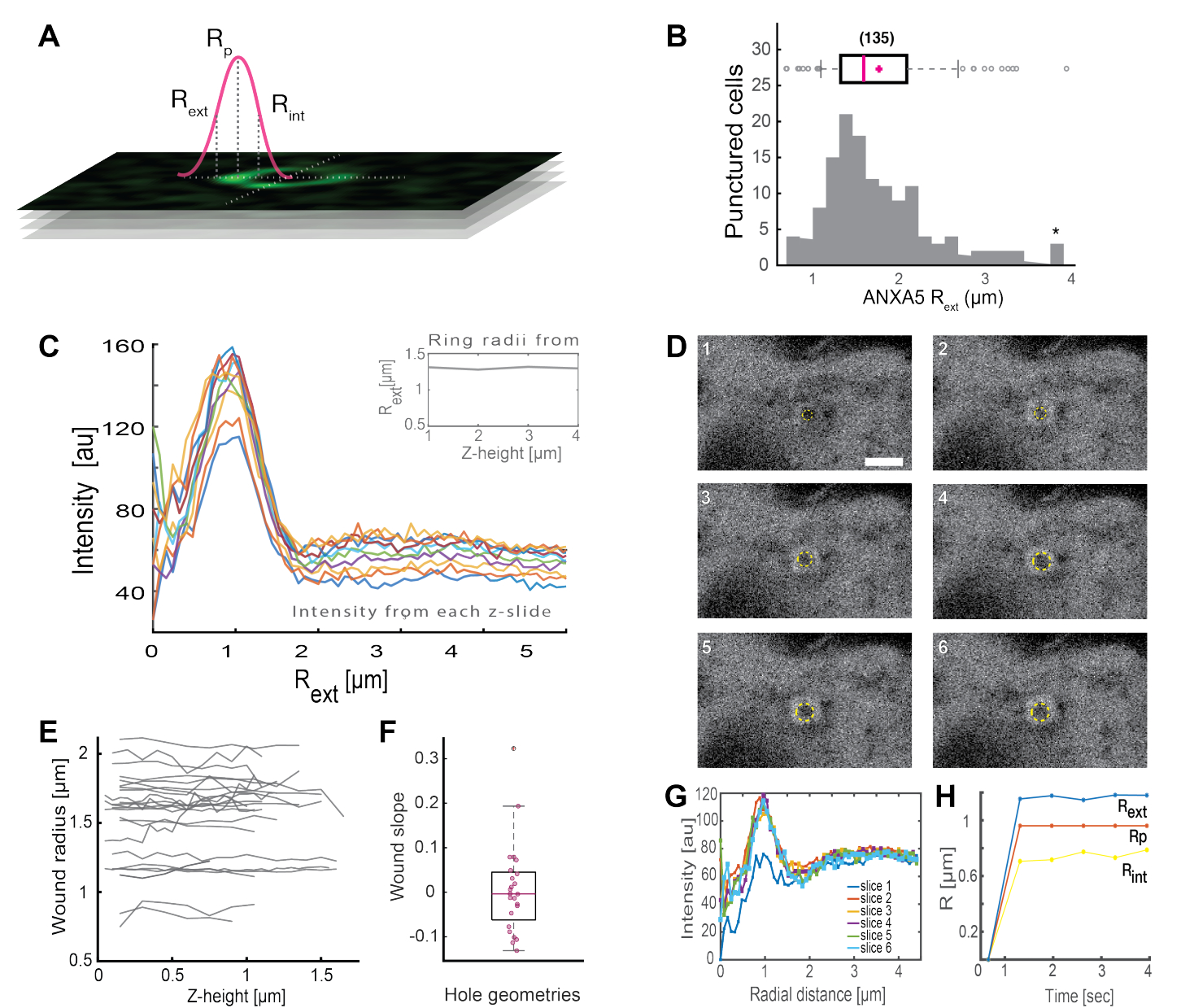
Die Analyse vollständiger ringförmiger ANXA-Strukturen (Abbildung 2B und ergänzende Abbildung 1) liefert nützliche Einblicke in die Wundgröße und -morphologie. Der Radius der ANXA-Ringstruktur kann über Zeit und Raum bestimmt werden, wie in Abschnitt 5 beschrieben. Moreno et al.16 analysierten über 135 Plasmamembranverletzungen in lebenden Zellen, wobei die Analyse der ANXA5-Ringstrukturen in Abbildung 3 dargestellt ist. Der Radius wurde bestimmt, indem der Abstand von der Mitte des Rings zum äußeren Radius der Kurve gemessen wurde, basierend auf dem Halbwertmaximum der vollen Breite des kollabierten Intensitätsprofils (Abbildung 3A). Die Ergebnisse zeigten eine heterogene Verteilung der ANXA5-Ringgrößen (Abbildung 3B), die über die Zeit (Abbildung 3D,G,H) und den Raum (Abbildung 3C,E,F) konstant blieb. Diese Ergebnisse deuten auf eine Akkumulation von ANXA5 um die Verletzungsstellen hin, was auf eine alternative ANXA5-vermittelte PMR-Strategie in lebenden Zellen zur hypothetischen trichterartigen Einknospenbildung der geschädigten Membran hindeutet5.

Abbildung 3: Analyse von ANXA5-Ringstrukturen, die eine Verletzungsstelle in lebenden Zellen umgeben. (A) Die schematische Darstellung veranschaulicht den analytischen Ansatz, der auf dem Maximum der vollen Breite und des halben Maximums des ANXA-Intensitätslinienprofils basiert. (B) Das Histogramm zeigt 135 gemessene ANXA5-Ringradien. (C) Fluoreszenzintensitätslinienprofile über einen ANXA5-GFP-Ring für jeden Z-Abschnitt des Z-Stapels. (D) Die konfokalen Bilder veranschaulichen die zeitliche Entwicklung einer Wunde, bei der sich ANXA5-GFP unmittelbar nach der Verletzung am Wundrand ansammelt, gefolgt von einer Wundstabilisierung. Die mit 1-6 gekennzeichneten Zeitrahmen wurden in Intervallen von 0,66 s pro Frame erfasst. Der Maßstabsbalken beträgt 2 μm. (E) Die Radien der Wunden wurden in Abhängigkeit von der Wundtiefe auf der Grundlage der in Tafel A vorgestellten Methode bestimmt. (F) Die Steigung der Wunde wurde als ANXA5-Ringradien als Funktion der z-Höhe basierend auf den aus Tafel E extrahierten Daten analysiert. (G) Die Fluoreszenzintensitätslinienprofile wurden über den ANXA5-GFP-Ring aus Tafel D gemessen. wobei jedes Zeitintervall 1-6 als Scheibe 1-6 bezeichnet wird. Schließlich wird (H) die zeitliche Entwicklung der ANXA5-GFP-Ringradien als Funktion der Zeit dargestellt, die aus den Daten in Panel G berechnet wird. Diese Abbildung ist reproduziert von Moreno-Pescador et. al16 mit Genehmigung der Royal Society of Chemistry. Bitte klicken Sie hier, um eine größere Version dieser Abbildung anzuzeigen.
{kind=link}
Im Vergleich zu den ringförmigen Strukturen, die allein durch die Störung des PM gebildet werden (Ergänzende Abbildung 1), können Verletzungen in unmittelbarer Nähe des Zellkerns (Ergänzende Abbildung 2A) die Entwicklung und Geometrie der Wunde beeinflussen. Gelegentlich war nur ein Bruchteil der ANXA-Ringe erkennbar (Ergänzende Abbildung 2B,C), die auch mit dem hauseigenen MATLAB-Analyse-Workflow analysiert werden können (siehe Abschnitt 5), obwohl zusätzliche Daten verloren gehen können. Typischerweise befinden sich ANXA-Ringbildungen, die in nicht-adhärenten Zellen beobachtet werden (Ergänzende Abbildung 2C), sowohl in der Nähe des Zellkerns als auch der Zellperipherie. In der Folge ist eine länglichere Ringstruktur zu beobachten, die für die vorgestellte Datenanalyse suboptimal ist. Darüber hinaus schienen nicht-adhärente Zellen anfälliger für den Zelltod nach einer PM-Verletzung zu sein. Darüber hinaus ist es bei der Betrachtung von Verletzungen, die durch die Bestrahlung von AuNP-Aggregaten verursacht werden, wichtig zu beachten, dass diese Verletzungen schwerwiegender und weniger kontrollierbar sein können. Dies ist auf die deutlich erhöhte plasmonische Erwärmung zurückzuführen, die einen großen Teil der Zelle schädigen kann. Daher wurden solche Verletzungen nicht in die ANXA5-Ringanalyse aufgenommen.
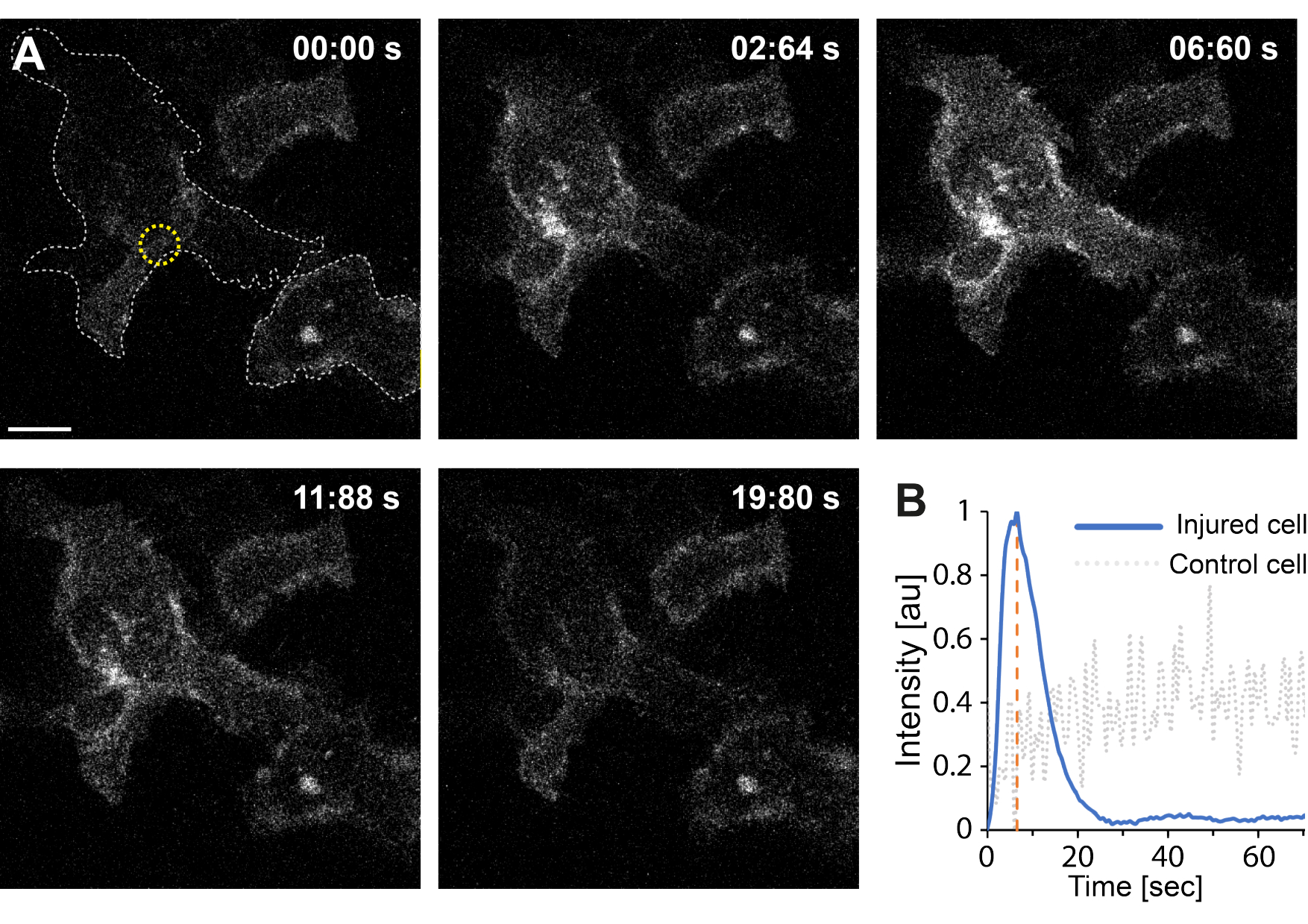
Darüber hinaus deuten die vorläufigen Ergebnisse darauf hin, dass die Störung der Plasmamembran mittels Thermoplasmonik zu erhöhten intrazellulärenCa2+ -Spiegeln führt. Dies wurde sogar bei Bestrahlung einzelner AuNPs mit geringer Intensität beobachtet, was auf eine PM-Permeabilisierung35 hindeutet, wie in Abbildung 4 dargestellt. Der Ca2+ Einstrom wurde in Zellen beobachtet, die eine membrangebundene Kalziumsonde, GCaMP6s-CAAX, exprimierten, die bei Ca2+ Einstrom eine Konformationsänderung erfährt, so dass eine Zunahme ihrer Intensität beobachtet werdenkann 64. Die Kalziumintensität wurde für den gesamten Fußabdruck der Zelle über die Zeit quantifiziert. Um Hintergrundgeräusche zu eliminieren, wurde das Hintergrundniveau von Ca2+ vor der Membranauffüllung und der Reparatur nach der Membran subtrahiert. Die maximale Intensität wurde durch Normalisierung der mittleren Ca2+ Intensität innerhalb der Zelle bestimmt, was zu einer Intensitätskurve führte, die einen schnellen anfänglichen Anstieg der Ca2+ Intensität zeigte, gefolgt von einer langsameren Abnahme, wie in Abbildung 4B dargestellt.
Die Zelle erreichte eine maximale Kalziumintensität bei ~ 6,6 s, was mit den Ergebnissen von Klenow et al.64 übereinstimmt, die darauf hinwiesen, dass die Zeit des Kalziumintensitätspeaks (t = tc) der Zeit entspricht, die für den Wundverschluss benötigt wird. Obwohl weitere Untersuchungen erforderlich sind, um den zugrundeliegenden Mechanismus der Membranreparatur und Wundheilung zu ermitteln, zeigten die vorläufigen Ergebnisse, dass dieser Ca2+ Prozess ausschließlich in der verletzten Zelle beobachtet wurde und nicht in der unverletzten Zelle, die als Positivkontrolle verwendet wurde. Dies bestätigt, dass die Zelle einen Ca2+ Einstrom bei der Zerlegung der thermoplasmonischen Membran erfuhr, bei dem das überschüssige intrazelluläre Kalzium nach erfolgreicher PMR aktiv abgepumpt wird, da der intrazelluläre Kalziumspiegel nicht mehr in Konkurrenz zum Ca2+ Einstrom steht, wodurch schließlich die Zellhomöostase erreichtwird 64.

Abbildung 4: Der Kalziumeinstrom tritt auf, wenn die Plasmamembran einer HEK293T Zelle durch Thermoplasmonik aufgerissen wird. Eine Reihe von konfokalen Bildern zeigt zwei Zellen (die Zelle von Interesse und eine unverletzte Zelle, die als Positivkontrolle verwendet wurde), die die membrangebundene Kalziumsonde GCaMP6s-CAAX exprimieren. Der Maßstabsbalken misst 10 μm. (A) Vor der Bestrahlung, um 00:00 Uhr, wird der Fußabdruck der beiden Zellen durch die grau gestrichelte Linie visualisiert, und der Bestrahlungsort wird durch den gelben Kreis bezeichnet. Bei Laserbestrahlung wird ein schneller Einstrom von Ca2+ beobachtet, der seine maximale Intensität bei ~ 6,6 s erreicht, gekennzeichnet durch die orange gestrichelte Linie, ein Zeitpunkt, von dem angenommen wird, dass er dem Zeitpunkt des Wundverschlusses64 entspricht. (B) Das Kalziumintensitätsprofil, das von der GCaMP6s-CAAX-Sonde in der verletzten Zelle (blaue Linie) erhalten wurde, wurde mit der Ca2+ -Intensität in einer benachbarten unverletzten Zelle (grau gestrichelte Linie) verglichen und zeigte einen deutlichen Ca2+ -Einstrom ausschließlich bei PM-Störung. Bitte klicken Sie hier, um eine größere Version dieser Abbildung anzuzeigen.
{kind=link}
Diskussion
Die Studie hebt den thermoplasmonischen Ansatz als vielversprechende Technik hervor, um Proteinantworten in lebenden Zellen und Modellmembranen nach Membranaufschluss zu untersuchen. Diese Methode liefert nicht nur umfangreiche Informationen über die Proteinrekrutierung, sondern auch über die biophysikalische Funktion von Proteinen, die an der Proteinmembrandynamik beteiligt sind. Folglich erleichtert es die Identifizierung molekularer Komponenten, die für die Oberflächenreparatur verantwortlich sind, und fördert das Verständnis der komplexen, aber lebenswichtigen Maschinerie der Plasmamembranreparatur. Obwohl es verschiedene Methoden gibt, um Membranaufschluss zu induzieren, wie z. B. mechanische, chemische und optische Techniken, leiden diese Methoden unter Einschränkungen, wie z. B. dass sie nicht spezifisch für Zellen sind, mehrere Verletzungen der Zellmembran verursachen oder eine erhebliche Schädigung der Membran verursachen und internes Zellmaterial entlang des Laserpfads abtragen, wenn gepulste Hochleistungslaser verwendet werden. Während die Integration von konfokaler Mikroskopie und optischer Pinzette die umfassendsten Informationen bietet, könnten auch alternative Bildgebungsmodalitäten verwendet werden. Da beispielsweise die Abbildung des plasmonischen Nanopartikels mit Hilfe der Reflexionsmikroskopie, einem eingebauten Bildgebungsmodus in konfokalen Leica-Mikroskopen, erreicht wird, könnten zusätzliche bildgebende Verfahren wie die Dunkelfeldmikroskopie65,66, andere Streumethoden wie iSCAT67,68 oder die Fluoreszenzmarkierung des Nanopartikels für die AuNP-Visualisierung eingesetzt werden, obwohl dies die Anwendbarkeit der Methode einschränken könnte.
Die vorgestellte Methode ist zusätzlich in der Lage, nanoskopische Löcher in Modellmembranen zu induzieren, was die Untersuchung von Synergieeffekten zwischen verschiedenen Anhängsen ermöglicht. Dies wird durch die Verkapselung unterschiedlich markierter rekombinanter Annexine, z.B. RFP bzw. GFP, erreicht, gefolgt von einer thermoplasmonischen Punktion. Dieses Modellsystem gibt Aufschluss darüber, wie Annexine mit Membranen in der Nähe freier Kanten interagieren, wie in Abbildung 2D gezeigt. Anders als in Zellen dehnen sich die Löcher bei GUVs jedoch weiter aus, gefolgt von einer Destabilisierung des Vesikels. Die Abbildung der Lochentwicklung mit konfokaler Mikroskopie kann aufgrund der schnellen Ausdehnung des Lochdurchmessers eine Herausforderung darstellen, kann aber durch die Erfassung mehrerer Z-Stapel im Laufe der Zeit erreicht werden. Eine alternative Methode wäre die Verwendung einer konfokalen rotierenden Scheibe für eine schnellere Bildgebung. Darüber hinaus liefert der thermoplasmonische Ansatz in der Regel eine begrenzte Anzahl optimaler Ergebnisse pro Stunde, wenn er auf Einzelzellen oder GUV-Experimente angewendet wird, in der Regel zwei bis drei, bei Probentemperaturen zwischen 20 °C und 30 °C. Um eine möglichst genaue Beobachtung der Protein-Membran-Dynamik zu erhalten, wird empfohlen, die Zellen in einem HEPES-haltigen Puffer zu halten und die Probe stündlich auszutauschen. Alternativ könnte das experimentelle Fenster erweitert werden, indem die Versuche in einer Zellinkubationskammer, d.h. bei einer konstanten Temperatur von 37 °C mit 5 %CO2, durchgeführt werden. Darüber hinaus könnte die Kombination dieses Ansatzes mit anderen bildgebenden Verfahren, wie der stochastischen optischen Rekonstruktionsmikroskopie (STORM), ein tieferes Verständnis der biophysikalischen Funktion und Interaktion von Schlüsselproteinen liefern, die an der Membranreparatur auf Einzelmolekülebene beteiligt sind. Dies könnte detaillierte Informationen über den Ort der Verletzung liefern, einschließlich der Wundgeometrie und der Lage der Annexin-Proteine, sowie andere Schlüsselakteure identifizieren, die an der Reparatur der Membranoberfläche beteiligt sind.
Um maximale Effektivität und Präzision bei der Induktion von Membranverletzungen zu erreichen, ist es zwingend erforderlich, die Position des Laserfokus vor jedem Experiment zu überprüfen und sicherzustellen, dass die axiale Position des Laserfokus mit dem konfokalen Fokus übereinstimmt. Diese Ausrichtung optimiert die Intensität während der AuNP-Bildgebung, was zu einem maximalen lokalen Temperaturanstieg und einer daraus resultierenden Membranverletzung bei geringerer Laserleistung führt. Dieser Prozess wird manuell ausgeführt und ist daher anfällig für Schwankungen in der Membranbrucheffizienz, da der Fokus manuell in eine Position verschoben wird, die mit der Position des Partikels übereinstimmt. Bei Mikroskopen, die keinen Reflexionsmodus haben, wie z. B. in einigen kommerziellen Systemen, kann die Kolokalisierung von Laserfokus und Partikel eine Herausforderung darstellen. In solchen Fällen können alternative Bildgebungsmodi (z. B. Hellfeld) verwendet werden, und ein langsamer Rasterscan kann um die erwartete Partikelposition herum durchgeführt werden. Es sollte beachtet werden, dass eine niedrige Laserleistung wahrscheinlich nur eine Membranpermeabilisierung induziert, während eine hohe Laserleistung Temperaturen um den NP herum erzeugen kann, die den Siedepunkt von Wasser überschreiten, selbst wenn die Glasoberfläche einen kühlenden Effekt hat. Es wird geschätzt, dass die Bildung von Nanoblasen, die die NPs umgeben, zwischen 200 °C und 300 °Cstattfindet 25,48, wobei die explosive Hitze entweder zu einer Partikelverschiebung aus dem Laserfokus oder zu einer Partikelfragmentierung führen kann. Darüber hinaus stellt die Bildung von Nano- oder Mikrobläschen beim Erhitzen eine Herausforderung für diese Methode dar. Da Luftgrenzflächen die Membranen entfeuchten und eine Proteindestabilisierung verursachen können, was unerwünscht ist, ist es zwingend erforderlich, die Erwärmung bei der Untersuchung der Membranreparatur zu begrenzen. Bemerkenswert ist, dass Goldnanoschalen keine hohen Temperaturen tolerieren und sich unter diesen Bedingungen abbauen, wie die hochauflösende Mikroskopie58 zeigt.
Dieser Artikel enthält ein detailliertes Protokoll für die Verwendung von Thermoplasmonik zur Durchführung stark lokalisierter Punktionen in Membranen, das sowohl auf Zellen als auch auf Modellmembranen anwendbar ist. Um das Ausmaß der Erwärmung weiter zu reduzieren, können kleinere Nanopartikel verwendet werden, die mit NIR-Licht in Resonanz stehen, was intrazelluläre Punktionen in Endosomen, im endoplasmatischen Retikulum und in der Kernhülle ermöglicht. Solche Nanopartikel, einschließlich Stäbchen und Nanomatroschkas48, können verwendet werden, um die Reparatur der Kernhülle zu untersuchen, indem sie auf endozytosierte Goldnanopartikel abzielen, die leicht an der Zelloberfläche aufgenommen und in Richtung des Zellkerns69 transportiert werden. Insgesamt ermöglicht diese Technik die Identifizierung und Untersuchung von molekularen Schlüsselkomponenten, die an der PMR beteiligt sind, und klärt ihre biophysikalische Funktion und Rolle auf, während die Lebensfähigkeit der Zellen erhalten bleibt.
Offenlegungen
Die Autoren haben nichts zu verraten.
Danksagungen
Wir danken Jesper Nylandsted für die Bereitstellung von rekombinanten Annexin-Proteinen und Plasmiden, die für Annexine kodieren. Diese Arbeit wurde finanziell unterstützt vom Dänischen Rat für unabhängige Forschung, Naturwissenschaften (DFF-4181-00196), vom Interdisziplinären Synergieprogramm 2018 der Novo Nordisk Foundation (NNF18OC0034936), dem Wissenschaftlichen Komitee der Dänischen Krebsgesellschaft (R90-A5847-14-S2), der Lundbeck Foundation (R218-2016-534) und vom Lundbeck Foundation Center of Excellence (Biomembranes in Nanomedicine).
Materialien
| Name | Company | Catalog Number | Comments |
| 1064 nm trapping laser | Spectra Physics | N/A | Spectra Physics J201-BL-106C, Nd: YVO4 NIR laser |
| 160 nm Gold Nanoshells | NanoComposix | NCXGSIR150 | |
| 200 nm Gold Nanoparticles | BBI Solutions | EM.GC200/7 | |
| 35 mm glass surface MatTex microwell | MATTEK | P35G-1.5-14-C | |
| Amber-glass vials | Supelco Sigma Aldrich | 243438 | |
| Annexin A2 plasmids | N/A | N/A | Received from our collaborator at the Danish Cancer Research Center |
| Annexin A4 recombinant-protein | N/A | N/A | N-terminal GFP tagged ANXA4 received from our collaborator at the Danish Cancer Research Center |
| Annexin A5 recombinant-protein | N/A | N/A | N-terminal GFP tagged ANXA5 received from our collaborator at the Danish Cancer Research Center |
| beta-casein | Sigma Life Science | C6905-1G | |
| CaCl2 | Suprlco (sigma Aldrich) | 10035-04-8 | |
| Centrifuge 5702 | Eppendorf | 5702 | |
| Chloroform | VWR Chemicals | 67-66-3 | |
| Culture dish (Nunclon Delta Surface) | Thermo scientific | 150460 | |
| DID cell-labelling Solution | Invitrogen | 7757 | |
| Distilled water | Gibco | 15230-089 | |
| DOPC | Avanti Polar Lipids | 850375C | Dissolved in chloroform |
| DOPS | Avanti Polar Lipids | 840035C | Dissolved in chloroform |
| Dulbecco's Modified Eage's Medium | Thermo Fisher Scientific | 11995065 | |
| FIJI ImageJ distribution | ImageJ2 | N/A | |
| GCaMP6s-CAAX | N/A | Received from our collaborator at the Danish Cancer Research Center | |
| Gibco Fetal Bovine Serum | Fisher Scientific | 11573397 | 10% of the culture medium |
| Glucose | PROLABO | 24 374.297 | |
| Hamilton syringes | Hamilton Company | N/A | 50 and 500 microliters |
| Harrick Plasma Cleaner PDG-002 | Harrick Plasma | N/A | |
| HEK293T cells | N/A | Received from our collaborator at the Danish Cancer Research Center | |
| Leica Acousto-Optical Beam Splitter (AOBS) | Leica | N/A | |
| Leica PL APO 63x water immersion objective, NA = 1.2 | Leica | N/A | |
| Leica SP5 confocal scanning microscope | Leica | N/A | |
| Lipofectamine | Fisher Scientific | 15338030 | |
| MatLab | The Mathworks, Inc., Natick, Massachusetts, United States | N/A | |
| NaCl | VWR Chemicals | 7647-14-5 | |
| Opti-MEM Reduced-Serum Medium | Thermo Fisher Scientific | 11058021 | |
| Parafilm | Bemis | PM-992 | |
| Penicillin-Streptomycin | Thermo Fisher Scientific | 15140122 | 1% of the culture medium |
| Phosphate Buffered Saline (PBS) | Thermo Fisher Scientific | 10010023 | |
| Piezoelectric stage (PI 731.20) | Physik Instrumente (Germany) | N/A | |
| Poly-L-Lysine | Sigma-Aldrich | P8920-100ML | 0.01-0.1% for coating |
| Polyvinyl alcohol | Sigma-Aldrich | 363065-25G | |
| round glass slide 25 mm Ø | VWR | 631-1584 | |
| Sonicator Brandson 2800 | Brandson | N/A | |
| sucrose | Sigma Life Science | 57-50-1 | |
| T25 tissue culture flask | Falcon | 353108 | Blue Vented cap |
| Tris-HCl | Invitrogen | 15567-027 | |
| TrypLE | Thermo Fisher Scientific | A1285901 | |
| Trypsin-EDTA | Fisher Scientific | 11590626 | |
| VWR Mixer mini vortex 230V EU | VWR | 12620-84 | ECN: 444-2790, SN: 150713022 |
Referenzen
- Bendix, P. M., et al. Interdisciplinary synergy to reveal mechanisms of annexin-mediated plasma membrane shaping and repair. Cells. 9 (4), 1029 (2020).
- Gajic, O., Lee, J., Doerr, C. H., Berrios, J. C., Myers, J. L., Hubmayr, R. D. Ventilator-induced Cell Wounding and Repair in the Intact Lung. American Journal of Respiratory and Critical Care Medicine. 167, 1057-1063 (2003).
- McNeil, P. L., Khakee, R. Disruptions of muscle fiber plasma membranes. Role in exercise-induced damage. The American Journal of Pathology. 140 (5), 1097-1109 (1992).
- Yu, Q. C., McNeil, P. L. Transient disruptions of aortic endothelial cell plasma membranes. The American Journal of Pathology. 141 (6), 1349-1360 (1992).
- Boye, T. L., et al. Annexin A4 and A6 induce membrane curvature and constriction during cell membrane repair. Nature Communications. 8, 1623 (2017).
- Bischofberger, M., Gonzalez, M. R., van der Goot, F. G. Membrane injury by pore-forming proteins. Current Opinion in Cell Biology. 21, 589-595 (2009).
- Tang, S. K. Y., Marshall, W. F. Self-repairing cells. Science (New York, N.Y.). 356, 1022-1025 (2017).
- Abreu-Blanco, M. T., Verboon, J. M., Parkhurst, S. M. Single cell wound repair: Dealing with life's little traumas. Bioarchitecture. 1, 114-121 (2011).
- Sønder, S. L., et al. Annexin A7 is required for ESCRT III-mediated plasma membrane repair. Scientific Reports. 9, 6726 (2019).
- Andrews, N. W., Almeida, P. E., Corrotte, M. Damage control: cellular mechanisms of plasma membrane repair. Trends in Cell Biology. 24 (12), 734-742 (2014).
- Idone, V., Tam, C., Goss, J. W., Toomre, D., Pypaert, M., Andrews, N. W. Repair of injured plasma membrane by rapid Ca2+-dependent endocytosis. The Journal of Cell Biology. 180 (5), 905-914 (2008).
- Lauritzen, S. P., Boye, T. L., Nylandsted, J. Annexins are instrumental for efficient plasma membrane repair in cancer cells. Seminars in Cell & Developmental Biology. 45, 32-38 (2015).
- Häger, S. C., Nylandsted, J. Annexins: players of single cell wound healing and regeneration. Communicative & Integrative Biology. 12 (1), 162-165 (2019).
- Jaiswal, J. K., et al. S100A11 is required for efficient plasma membrane repair and survival of invasive cancer cells. Nature Communications. 5, 3795 (2014).
- Draeger, A., Monastyrskaya, K., Babiychuk, E. B. Plasma membrane repair and cellular damage control: The annexin survival kit. Biochemical Pharmacology. 81 (6), 703-712 (2011).
- Moreno-Pescador, G. S., et al. Thermoplasmonic nano-rupture of cells reveals annexin V function in plasma membrane repair. Nanoscale. 14 (21), 7778-7787 (2022).
- Zhivotovsky, B., Orrenius, S. Calcium and cell death mechanisms: A perspective from the cell death community. Cell Calcium. 50 (3), 211-221 (2011).
- Gerke, V., Moss, S. E. Annexins: From structure to function. Physiological Reviews. 82 (2), 331-371 (2002).
- Idone, V., Tam, C., Andrews, N. W. Two-way traffic on the road to plasma membrane repair. Trends in Cell Biology. 18 (11), 552-559 (2008).
- Boye, T. L., et al. Annexins induce curvature on free-edge membranes displaying distinct morphologies. Scientific Reports. 8, 10309 (2018).
- Bouter, A., et al. Annexin-A5 assembled into two-dimensional arrays promotes cell membrane repair. Nature Communications. 2, 270 (2011).
- Boye, T. L., Nylandsted, J. Annexins in plasma membrane repair. Biological Chemistry. 397 (10), 961-969 (2016).
- Weinberger, A., et al. Gel-assisted formation of giant unilamellar vesicles. Biophysical Journal. 105 (1), 154-164 (2013).
- Numata, T., Tatsuta, H., Morita, Y., Otani, Y., Umeda, N. Localized thermal processing with a laser-trapped and heated metal nanoparticle. IEEJ Transactions on Electrical and Electronic Engineering. 2, 398-401 (2007).
- Bendix, P. M., Reihani, S. N. S., Oddershede, L. B. Direct measurements of heating by electromagnetically trapped gold nanoparticles on supported lipid bilayers. ACS Nano. 4 (4), 2256-2262 (2010).
- Kyrsting, A., Bendix, P. M., Stamou, D. G., Oddershede, L. B. Heat profiling of three-dimensionally optically trapped gold nanoparticles using vesicle cargo release. Nano Letters. 11 (2), 888-892 (2011).
- Andersen, T., Kyrsting, A., Bendix, P. M. Local and transient permeation events are associated with local melting of giant liposomes. Soft Matter. 10 (24), 4268-4274 (2014).
- Bahadori, A., Oddershede, L. B., Bendix, P. M. Hot-nanoparticle-mediated fusion of selected cells. Nano Research. 10, 2034-2045 (2017).
- Rørvig-Lund, A., Bahadori, A., Semsey, S., Bendix, P. M., Oddershede, L. B. Vesicle fusion triggered by optically heated gold nanoparticles. Nano Letters. 15 (6), 4183-4188 (2015).
- Moreno-Pescador, G., Arastoo, M. R., Ruhoff, V. T., Chiantia, S., Daniels, R., Bendix, P. M. Thermoplasmonic vesicle fusion reveals membrane phase segregation of influenza spike proteins. Nano Letters. 23 (8), 3377-3384 (2023).
- Bahadori, A., Lund, A. R., Semsey, S., Oddershede, L. B., Bendix, P. M. Controlled cellular fusion using optically trapped plasmonic nano-heaters. SPIE Proceedings. SPIE 9922, Optical Trapping and Optical Micromanipulation XIII. 992211, (2016).
- Bahadori, A., Moreno-Pescador, G., Oddershede, L. B., Bendix, P. M. Remotely controlled fusion of selected vesicles and living cells: a key issue review. Reports on Progress in Physics. 81 (3), 32602 (2018).
- Moreno-Pescador, G., Arastoo, M. R., Chiantia, S., Daniels, R., Bendix, P. M. Thermoplasmonic induced vesicle fusion for investigating membrane protein phase affinity. bioRxiv. , (2022).
- Pescador, G. S. M., et al. Investigating plasma-membrane repair employing thermoplasmonics. Biophysical Journal. 120 (3), 45A (2021).
- Moreno-Pescador, G. S., Qoqaj, I., Thusgaard Ruhoff, V., Iversen, J., Nylandsted, J., Bendix, P. M. Effect of local thermoplasmonic heating on biological membranes. SPIE 11083, Optical Trapping and Optical Micromanipulation XVI. 110830M, (2019).
- Bement, W. M., Mandato, C. A., Kirsch, M. N. Wound-induced assembly and closure of an actomyosin purse string in Xenopus oocytes. Current Biology. 9 (11), 579-587 (1999).
- Weisleder, N., et al. Recombinant MG53 protein modulates therapeutic cell membrane repair in treatment of muscular dystrophy. Science Translational Medicine. 4 (139), 139ra85 (2012).
- Sudji, I. R., Subburaj, Y., Frenkel, N., García-Sáez, A. J., Wink, M. Membrane disintegration caused by the steroid saponin digitonin is related to the presence of cholesterol. Molecules. 20 (11), 20146-20160 (2015).
- Babiychuk, E. B., Monastyrskaya, K., Potez, S., Draeger, A. Intracellular Ca2+ operates a switch between repair and lysis of streptolysin O-perforated cells. Cell Death & Differentiation. 16, 1126-1134 (2009).
- Nygård Skalman, L., Holst, M. R., Larsson, E., Lundmark, R. Plasma membrane damage caused by listeriolysin O is not repaired through endocytosis of the membrane pore. Biology Open. 7 (10), bio035287 (2018).
- Swaggart, K. A., et al. Annexin A6 modifies muscular dystrophy by mediating sarcolemmal repair. Proceedings of the National Academy of Sciences of the United States of America. 111, 6004-6009 (2014).
- Yeheskely-Hayon, D., Minai, L., Golan, L., Dann, E. J., Yelin, D. Optically induced cell fusion using bispecific nanoparticles. Small. 9 (22), 3771-3777 (2013).
- Minai, L., Yeheskely-Hayon, D., Golan, L., Bisker, G., Dann, E. J., Yelin, D. Optical nanomanipulations of malignant cells: Controlled cell damage and fusion. Small. 8 (11), 1732-1739 (2012).
- Lukianova-Hleb, E., et al. Plasmonic nanobubbles as transient vapor nanobubbles generated around plasmonic nanoparticles. ACS Nano. 4 (4), 2109-2123 (2010).
- Vogel, A., Noack, J., Hüttman, G., Paltauf, G. Mechanisms of femtosecond laser nanosurgery of cells and tissues. Applied Physics B. 81, 1015-1047 (2005).
- Baffou, G., Polleux, J., Rigneault, H., Monneret, S. Super-heating and micro-bubble generation around plasmonic nanoparticles under cw illumination. Journal of Physical Chemistry C. 118 (9), 4890-4898 (2014).
- Sasikumar, K., Liang, Z., Cahill, D. G., Keblinski, P. Curvature induced phase stability of an intensely heated liquid. Journal of Chemical Physics. 140 (23), 234506 (2014).
- Jauffred, L., Samadi, A., Klingberg, H., Bendix, P. M., Oddershede, L. B. Plasmonic heating of nanostructures. Chemical Reviews. 119 (13), 8087-8130 (2019).
- Neuman, K. C., Nagy, A. Single-molecule force spectroscopy: optical tweezers, magnetic tweezers and atomic force microscopy. Nature Methods. 5 (6), 491-505 (2008).
- Bendix, P. M., Jauffred, L., Norregaard, K., Oddershede, L. B. Optical trapping of nanoparticles and quantum dots. IEEE Journal of Selected Topics in Quantum Electronics. 20, 15-26 (2014).
- Samadi, A., Bendix, P. M., Oddershede, L. B. Optical manipulation of individual strongly absorbing platinum nanoparticles. Nanoscale. 46, 18449-18455 (2017).
- Jørgensen, J. T., Norregaard, K., Tian, P., Bendix, P. M., Kjaer, A., Oddershede, L. B. Single particle and PET-based platform for identifying optimal plasmonic nano-heaters for photothermal cancer therapy. Scientific Reports. 6, 30076 (2016).
- Goldenberg, H., Tranter, C. J. Heat flow in an infinite medium heated by a sphere. British Journal of Applied Physics. 3 (9), 296-298 (1952).
- Eustis, S., El-Sayed, M. A. Why gold nanoparticles are more precious than pretty gold: Noble metal surface plasmon resonance and its enhancement of the radiative and nonradiative properties of nanocrystals of different shapes. Chemical Society Reviews. 35, 209-217 (2006).
- Landau, L. D., Lifshitz, E. M. . Fluid Mechanics: Landau and Lifshitz: Course of Theoretical Physics. 6, (2013).
- Niederauer, C., Seynen, M., Zomerdijk, J., Kamp, M., Ganzinger, K. A. The K2: Open-source simultaneous triple-color TIRF microscope for live-cell and single-molecule imaging. HardwareX. 13, e00404 (2023).
- Richardson, A. C., Reihani, N., Oddershede, L. B. Combining confocal microscopy with precise force-scope optical tweezers. SPIE Proceedings:SPIE 6326, Optical Trapping and Optical Micromanipulation III. 632628, (2006).
- Samadi, A., Klingberg, H., Jauffred, L., Kjær, A., Bendix, P. M., Oddershede, L. B. Platinum nanoparticles: a non-toxic, effective and thermally stable alternative plasmonic material for cancer therapy and bioengineering. Nanoscale. 10 (19), 9097-9107 (2018).
- . Available from: https://www.thermofisher.com/order/catalog/product/A7816 (2023)
- Kreibig, U., Vollmer, M. Theoretical considerations. In: Optical Properties of Metal Clusters. 25, (1995).
- Mie, G. Beiträge zur Optik trüber Medien, speziell kolloidaler Metallösungen. Annalen der Physik. 330 (3), 377-445 (1908).
- Rueden, C. T., et al. ImageJ2: ImageJ for the next generation of scientific image data. BMC Bioinformatics. 18 (1), 529 (2017).
- Schindelin, J., et al. Fiji: An open-source platform for biological-image analysis. Nature Methods. 9, 676-682 (2012).
- Klenow, M. B., Heitmann, A. S. B., Nylandsted, J., Simonsen, A. C. Timescale of hole closure during plasma membrane repair estimated by calcium imaging and numerical modeling. Scientific Reports. 11, 4226 (2021).
- Li, T., Wu, X., Liu, F., Li, N. Analytical methods based on the light-scattering of plasmonic nanoparticles at the single particle level with dark-field microscopy imaging. Analyst. 142 (2), 248-256 (2017).
- Gibbs-Flournoy, E. A., Bromberg, P. A., Hofer, T. P. J., Samet, J. M., Zucker, R. M. Darkfield-Confocal Microscopy detection of nanoscale particle internalization by human lung cells. Particle and Fibre Toxicology. 8 (1), 2 (2011).
- Taylor, R. W., Sandoghdar, V. Interferometric scattering microscopy: Seeing single nanoparticles and molecules via Rayleigh scattering. Nano Letters. 19 (8), 4827-4835 (2019).
- Wu, Y., Ali, M. R. K., Chen, K., Fang, N., El-Sayed, M. A. Gold nanoparticles in biological optical imaging. Nano Today. 24, 120-140 (2019).
- Klingberg, H., Oddershede, L. B., Loeschner, K., Larsen, E. H., Loft, S., Møller, P. Uptake of gold nanoparticles in primary human endothelial cells. Toxicology Research. 4 (3), 566-666 (2015).
Nachdrucke und Genehmigungen
Genehmigung beantragen, um den Text oder die Abbildungen dieses JoVE-Artikels zu verwenden
Genehmigung beantragenThis article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. Alle Rechte vorbehalten