
Method Article
Uma Abordagem Termoplasmônica para Investigação do Reparo da Membrana Plasmática em Células Vivas e Membranas Modelo
Neste Artigo
Resumo
O método de punção termoplasmônica integra microscopia confocal, pinças ópticas e nanopartículas de ouro para estudar as respostas proteicas durante o reparo da membrana plasmática em células e vesículas unilamelares gigantes. A técnica permite a punção rápida e localizada da membrana, permitindo a identificação de proteínas-chave e seus papéis funcionais na intrincada maquinaria de reparo da membrana plasmática.
Resumo
A membrana celular é crucial para a sobrevivência celular, e garantir sua integridade é essencial, pois a célula sofre lesões ao longo de todo o seu ciclo de vida. Para evitar danos à membrana, as células desenvolveram mecanismos eficientes de reparo da membrana plasmática. Esses mecanismos de reparo podem ser estudados combinando microscopia confocal e termoplasmônica em nanoescala para identificar e investigar o papel de proteínas-chave, como as anexinas, envolvidas no reparo de superfície em células vivas e sistemas modelo de membrana.
O método de punção emprega um laser para induzir aquecimento altamente localizado sobre a irradiação de nanopartículas. O uso de luz infravermelha próxima minimiza a fototoxicidade na amostra biológica, enquanto a maior parte da absorção ocorre na nanopartícula plasmônica ressonante do infravermelho próximo. Este método termoplasmônico tem sido explorado para potenciais pesquisas fototérmicas e biofísicas para melhorar a compreensão dos mecanismos intracelulares e respostas celulares através de estudos de fusão de vesículas e células. A abordagem tem se mostrado complementar aos métodos existentes para ruptura de membranas, como lesões induzidas mecanicamente, quimicamente ou opticamente, e fornece um alto nível de controle ao infligir lesões extremamente localizadas. A extensão da lesão é limitada à vizinhança da nanopartícula esférica, e nenhum dano prejudicial ocorre ao longo do caminho do feixe, em oposição aos lasers pulsados usando diferentes comprimentos de onda. Apesar de certas limitações, como a formação de nanobolhas, o método termoplasmônico oferece uma ferramenta única para investigar respostas celulares no reparo da membrana plasmática em um ambiente quase nativo sem comprometer a viabilidade celular.
Quando integrado à microscopia confocal, o método de punção pode fornecer uma compreensão mecanística da dinâmica da membrana em sistemas de membrana modelo, bem como informações quantitativas sobre as respostas proteicas a danos na membrana, incluindo o recrutamento de proteínas e sua função biofísica. Em geral, a aplicação deste método a sistemas modelo reduzidos pode melhorar nossa compreensão da intrincada maquinaria de reparo da membrana plasmática em células vivas.
Introdução
A membrana celular, servindo tanto como barreira física quanto como plataforma de sinalização, é vital para a sobrevivência celular1. Durante todo o seu ciclo celular, a membrana plasmática (PM) está sujeita a danos, tais como lesões mecânicas 2,3,4,5 e químicas6 induzidas por estresse. Para manter a integridade da membrana e garantir a sobrevivência celular, a célula desenvolveu mecanismos robustos de reparo da membrana plasmática (PMR). Esses mecanismos dependem de várias estratégias, como reorganização do citoesqueleto, fusão de membranas e estratégias de substituição de membranas7,8,9,10,11, as quais dependem do recrutamento de proteínas específicas. Notavelmente, membros da família de proteínas anexinas têm sido identificados como proteínas-chave associadas aos processos de PMR 1,9,12,13,14,15,16. Após a lesão de PM, a célula experimenta um influxo de íons cálcio (Ca2+), o que representa uma ameaça imediata à sobrevivência da célula17. Em resposta ao influxo de Ca2+, as proteínas anexinas, predominantemente localizadas no citosol, ligam-se ao folheto interno da membrana plasmática lesada como parte das estratégias de RPM18. A anexina A2 (ANXA2) foi um dos primeiros membros da família das anexinas a ser associada à RPM na distrofia muscular deficiente em disferlina e foi sugerida para mediar o reparo pela fusão de vesículas intracelulares ao PM próximo ao local da lesão 5,19,20,21. Posteriormente, várias funções foram atribuídas às anexinas22, e seu papel na PMR ganhou maior atenção nos últimos 20 anos. No entanto, o papel exato das anexinas na RPM ainda não é totalmente compreendido 15,18,21,22.
Este artigo propõe um método para investigar a interação proteína-membrana e a dinâmica da membrana de forma controlada e altamente localizada, utilizando uma combinação de microscopia confocal, pinças ópticas e nanopartículas de ouro (AuNPs). Este método permite o estudo quantitativo de interações proteicas, lipídicas e de pequenas moléculas em resposta a danos na membrana e influxo de Ca2+ . Apesar da complexidade e multiplicidade de componentes envolvidos no processo de reparo da membrana, sistemas de membrana simplificados que mimetizam a membrana plasmática têm sido empregados para obter uma compreensão mecanicista mais profunda da dinâmica da membrana e da resposta das proteínas da anexina à ruptura da membrana16. Vesículas lipídicas unilamelares gigantes (GUVs) foram escolhidas como o sistema de membrana modelo com uma composição lipídica especificada. As GUVs foram geradas utilizando o método de hidratação assistida por gel, especificamente a hidratação em gel de álcool polivinílico, como descrito por Weinberger et al.23, o que permitiu um encapsulamento eficiente de anexinas em GUVs.
A utilização da irradiação com laser no infravermelho próximo (NIR) em nanopartículas metálicas (NPs) induz aquecimento significativo do NP, tornando-se um método eficaz para estabelecer uma fonte de calor local explorada em aplicações biomédicas24. O método foi inicialmente usado para medir diretamente a temperatura ao redor de uma única AuNP em ensaios biomiméticos 2D e 3D. Nestes ensaios25,26, as nanopartículas plasmônicas foram irradiadas em uma bicamada lipídica suportada ou aprisionadas opticamente próximas a GUVs submetidas a uma transição de fase térmica local após aquecimento local, permitindo a quantificação e o controle do perfil exato de temperatura ao redor da partícula. Este perfil de temperatura de referência tem sido utilizado na investigação ou manipulação de espécimes biológicos. Novos avanços no método facilitaram a indução de poros nanoscópicos nas membranas27, permitindo a fusão de vesículas e células28,29. Outros estudos investigaram o comportamento de proteínas de membrana periférica em GUVs29 e proteínas transmembrana30 por meio da criação de novas vesículas híbridas, enquanto a liberação de fármacos específicos para células também tem sido explorada para controlar e estudar respostas celulares ou expressão gênica 28,29,31,32,33. Recentemente, o método tem sido utilizado para investigar as respostas proteicas ao dano de membrana 32,34,35.
Existem vários métodos para romper a membrana plasmática para explorar as respostas celulares e o reparo da membrana. Estes incluem punções com microagulhas, agitação de microesferas e raspagem celular, que podem romper mecanicamente a membrana celular 14,36,37. Danos quimicamente induzidos podem ser obtidos pela adição de detergentes 5,38 ou toxinas bacterianas39,40 que desestabilizam a bicamada lipídica e geram poros de membrana através da membrana plasmática. Além disso, lesões opticamente induzidas por ondas contínuas e lasers pulsados têm sido utilizados para estudar componentes do PMR, como as proteínas da anexina 5,14,21,41, em combinação com nanopartículas plasmônicas 42,43,44,45. Apesar da eficiência dos lasers pulsados de alta potência, eles podem causar lesões significativas e danos ao interior da célula ao longo do caminho do feixe. Além disso, as mudanças detalhadas que ocorrem na matéria biológica após a irradiação com laser pulsado e se ela cria um poro bem definido ainda precisam ser mais investigadas. Um método alternativo é apresentado neste artigo, empregando termoplasmônicos para induzir orifícios nanoscópicos no MP de forma controlada34,35 sem causar danos significativos às estruturas internas. Isso é conseguido expondo NPs plasmônicos a um laser NIR altamente focado, resultando em um aumento de temperatura extremamente localizado que pode facilmente atingir temperaturas superiores a 200 °C, o que pode levar a pequenas explosões nanoscópicas 25,46,47. Esse processo pode ser controlado ajustando-se a intensidade do laser, bem como o tamanho, a forma e a composição dos NPs48. Ao empregar esta técnica, os pesquisadores podem explorar o papel das proteínas no reparo da PM em pilhas vivas, o que poderia ajudar a resolver algumas das perguntas não respondidas sobre o envolvimento das proteínas da anexina no reparo da membrana sem comprometer a viabilidade celular.
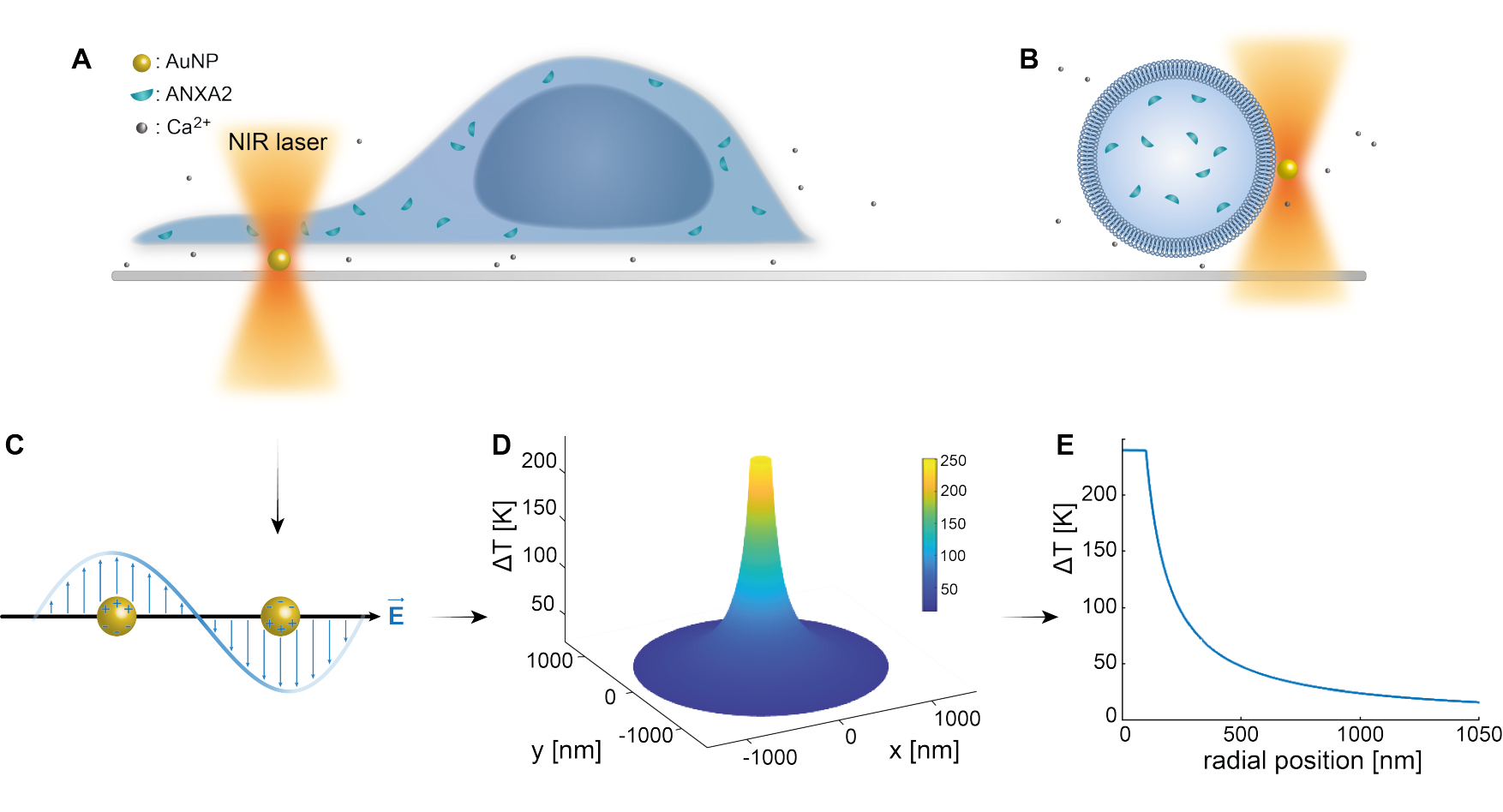
O aprisionamento óptico de nanopartículas plasmônicas foi bem estabelecido por estudos prévios 25,49,50,51,52; entretanto, informações adicionais sobre as propriedades termoplasmônicas das nanopartículas 53,54,55 podem ser obtidas nos materiais complementares (Arquivo Suplementar 1). O método termoplasmônico pode ser utilizado para criar orifícios nanoscópicos no MP com a finalidade de estudar a resposta celular e mecanismos de reparo. Mais precisamente, a punção pode ser obtida através do aquecimento óptico das AuNPs em estreita proximidade com a membrana, como mostram as Figuras 1A e B. Essa punção localizada permite o influxo de Ca2+, que foi verificado por um sensor de cálcio, ativando assim o maquinário PMR. Para experimentos com células vivas, AuNPs com diâmetro de 200 nm foram imobilizadas na superfície abaixo da célula para monitorar o papel de ANXA2 na RPM via microscopia confocal. O laser NIR (Figura 1A,B), com comprimento de onda de 1064 nm, irradia a AuNP, explorando suas propriedades plasmônicas (Figura 1C), resultando em substancial aquecimento local (Figura 1D) na janela de transparência biológica49 e causando danos mínimos à própria célula. A região de alta temperatura ao redor da AuNP diminui rapidamente em 30-40% a uma distância correspondente ao raio da NP, como ilustrado na Figura 1E, permitindo uma lesão extremamente confinada nas três dimensões.
Arquivo Suplementar 1. Clique aqui para baixar este arquivo.

Figura 1: Esquema do método experimental. (A) células transfectadas por ANXA estão situadas sobre nanopartículas de ouro imobilizadas (AuNPs) na superfície, ou (B) vesículas unilamelares gigantes (GUVs) com ANXA encapsulado são suspensas em um meio contendo AuNPs. (C) Uma única AuNP é irradiada pela armadilha óptica NIR, onde a interação entre o campo eletromagnético de entrada e os elétrons de condução leva à oscilação coletiva dos elétrons dentro do NP. (D) Este processo resulta em um aumento altamente confinado, mas significativo da temperatura. Para estimar a temperatura na superfície NP, a teoria de Mie é empregada, e um perfil de temperatura (E) é calculado para uma AuNP com diâmetro de 200 nm e intensidade de laser I = 6,36 x 108 W/cm2. Clique aqui para ver uma versão maior desta figura.
{kind=link}
Para minimizar o efeito térmico na membrana celular, as AuNPs são irradiadas apenas por ~1 segundo. Isso causa uma explosão transitória e local de aquecimento, o que reduz os danos às proteínas que normalmente exigem mais tempo para se desdobrar. Após a punção da membrana, as proteínas da anexina são recrutadas em uma fração de segundo e, em poucos segundos, um arcabouço semelhante a um anel de anexina é formado ao redor do local da lesão (Figura 2). Esta abordagem também tem sido aplicada para explorar o envolvimento de ANXA5 em células vivas e membranas modelo16 em um esforço para esclarecer o esquema completo dos processos de reparo. Embora o foco principal tenha sido o recrutamento correlacionado de várias proteínas de anexinas, os aspectos biofísicos do mecanismo de reparo ainda precisam ser elucidados.
Para implementar completamente o método proposto, três componentes principais são necessários: microscopia confocal, pinças ópticas e nanopartículas metálicas. Pinças ópticas são usadas para aprisionar AuNPs, e sua construção pode ser obtida seguindo o procedimento descrito por Neuman et al.49. No entanto, se a construção de uma pinça óptica se mostrar muito desafiadora, um laser NIR fortemente focado pode ser usado para irradiar AuNPs imobilizadas sob as células. Enquanto AuNPs esféricas foram escolhidas para este protocolo, uma variedade de partículas plasmônicas com espectros de absorção ajustáveis também pode ser utilizada para alcançar um gradiente de temperatura altamente localizado dentro da região NIR48.
A imagem por fluorescência é necessária para observar o papel das proteínas marcadas fluorescentemente e, portanto, a microscopia de reflexão interna total (TIRF)56 pode ser considerada uma alternativa à imagem confocal. No entanto, essa técnica permite apenas imagens de superfície e não seria compatível com os experimentos de vesículas de membrana modelo. Consequentemente, tanto a pinça óptica quanto o microscópio confocal são essenciais para a localização precisa da nanopartícula e investigação detalhada da área local ao redor da lesão celular. Para irradiar efetivamente a nanopartícula com um foco de laser limitado por difração, é necessário visualizar a nanopartícula. Isso pode ser obtido de forma otimizada pela microscopia de reflexão, que é um recurso de imagem padrão dos microscópios confocais Leica. No entanto, se a imagem de reflexão ou espalhamento não estiver disponível, métodos alternativos, como a marcação fluorescente de AuNP menos eficiente, podem ser considerados.
Em resumo, o método termoplasmônico altamente controlável e localizado apresentado neste estudo tem o potencial de servir como uma excelente plataforma para investigar os componentes moleculares envolvidos nas respostas celulares e mecanismos de reparo de MP em células vivas. Além de estudar a resposta proteica após danos de PM, essa abordagem também pode ser utilizada para puncionar vesículas localmente, permitindo assim uma investigação da resposta proteica tanto na dinâmica proteína-proteína quanto na proteína-membrana. Além disso, esse método permite uma análise quantitativa das interações entre proteínas, lipídios e pequenas moléculas quando as membranas são rompidas. Coletivamente, esses avanços têm o potencial de lançar luz sobre algumas das questões não resolvidas sobre a intrincada e complexa maquinaria de reparo da membrana plasmática.
Protocolo
1. Preparação da punção da membrana celular
- Semeadura celular (Dia 1)
- Cultivar células de rim embrionário humano (HEK293T) em meio de cultura a 37 °C em estufa de umidificador de CO2 a 5% até atingirem 70% de confluência.
- Separar as células da superfície usando 500 μL de tripsina, contar 200.000 células HEK293T e semeá-las em uma placa de cultura com um volume total de 3 mL de meio de cultura. Incubar as células a 37 °C numa estufa de umidificador de CO2 a 5% durante 24 horas.
NOTA: Para evitar o agrupamento de células no centro do prato, espalhe as células uniformemente e evite girar o prato, pois isso pode diminuir a eficiência da transfecção.
- Transfecção celular (Dia 2)
OBS: As células transfectadas podem ser utilizadas até 48 h após a transfecção.- Pipetar o plasmídeo de interesse e o reagente de transfecção por 5 s antes do uso.
- Em um tubo estéril de 2 mL, misturar o seguinte na ordem específica: 500 μL de meio de soro reduzido, 5 μL de reagente de transfecção (4 vezes mais que o plasmídeo) e 1,25 μL de plasmídeo (1 μg/μL).
NOTA: Para investigar o influxo de cálcio na ruptura da membrana, siga o mesmo procedimento, mas use a sonda do sensor de cálcio ligada à membrana GCaMP6-CAAX (1 μg/μL). - Pipetar suavemente, mas completamente, a mistura de transfecção e incubá-la à temperatura ambiente (TR) por 30 minutos antes de adicioná-la gota às células.
- Antes de adicionar a mistura de transfecção, retire o meio da placa de cultura, lave suavemente as células com 2 mL de solução salina tamponada com fosfato e adicione 2000 μL de meio de soro reduzido à placa.
- Incubar as células juntamente com a mistura de transfecção a 37 °C em uma estufa de umidificador de CO2 a 5% por 2 h e 45 min antes de trocar o meio para 3 mL de meio de cultura.
- Preparação de solução de nanopartículas de ouro (AuNP) (Dia 2)
- Vórtice a solução estoque de AuNP de 200 nm por 10 s no nível 10 (consulte Tabela de materiais para especificação adicional do aparelho), sonicate por 5 min (amplitude máxima) e vórtice novamente por 10 s.
- Misturar 150 μL de AuNPs com 850 μL de água destilada para um volume total de 1 mL.
NOTA: A solução de AuNP diluída pode ser armazenada na geladeira e reutilizada por até 1 mês.
- Preparo do prato experimental (Dia 2)
- Cubra a placa de micropoço com 150 μL de solução de fixação celular a 0,01%-0,1% e incube por 15 min no TR.
- Lave a superfície do vidro duas vezes com 500 μL de água destilada e deixe secar ao ar por ~10 min.
- Adicionar 80 μL de solução de AuNP gota a gota à superfície seca.
NOTA: Vórtice (10 s), sonicato (5 min) e vórtice (10 s) a solução de AuNP diluída antes de adicioná-la à superfície de vidro revestida para minimizar os agregados de AuNP. - Aguarde ~10 min antes de introduzir 1,5 mL de meio de cultura. Deixe o prato incubar a 37 °C durante a noite.
- Preparo da câmara experimental (Dia 3)
OBS: A câmara experimental pode ser preparada no dia 3 ou 4; No entanto, certifique-se de que as seguintes preparações sejam feitas no mesmo dia que o experimento.- Retire o meio das células na placa de cultura e lave as células com 2 mL de solução salina tamponada com fosfato.
NOTA: Esta etapa é essencial para remover qualquer meio residual e detritos que possam interferir com as etapas subsequentes. - Adicionar 500 μL de solução de descolamento celular à base de enzima ao poço e incubar por 1-3 min até que as células se desprendam da placa de cultura.
- Adicionar 1,5 mL de meio de cultura fresco e pipetar a solução celular para obter uma solução celular homogênea para minimizar a probabilidade de aglomerados celulares.
- Remova cuidadosamente o meio das AuNPs no micropoço experimental.
- Adicionar a solução celular (2 mL) ao micropoço e deixar incubar por pelo menos 5 h antes de realizar o experimento.
NOTA: Para condições experimentais ideais, evite girar a câmara, pois isso pode fazer com que as células se agrupem no meio da câmara.
- Retire o meio das células na placa de cultura e lave as células com 2 mL de solução salina tamponada com fosfato.
2. Experimento de punção de membrana celular
- Configurações ópticas do experimento
- Realizar os experimentos utilizando um microscópio de varredura confocal combinado com um laser de aprisionamento de 1064 nm57.
- Realizar o aprisionamento óptico no plano focal utilizando uma objetiva de imersão em água de 63x com uma abertura numérica (NA) de 1,2.
- Suponha que o foco é do tamanho de um disco Airy e que a largura do feixe focal do laser de irradiação é de ~540 nm de raio.
- Converter a potência do laser (P) para a intensidade do laser correspondente (I) calculando a potência do laser por área (W/cm2).
- Utilize um divisor de feixe óptico aousto (AOBS) para visualizar múltiplos sinais fluorescentes detectados usando tubos fotomultiplicadores e detecção simultânea de NPs metálicos através de seu sinal de espalhamento.
NOTA: Nem todos os sistemas confocais são equipados com AOBS, que permite imagens de reflexão de NPs metálicos. Aqui, outras formas de detecção devem ser implementadas, ou uma varredura sequencial do laser NIR na área abaixo da célula pode ser tentada. - Durante as sessões experimentais, monte uma câmara aberta com fundo de vidro contendo células, sondas fluorescentes moleculares e nanopartículas de ouro no microscópio.
- Mover a armadilha em relação às células traduzindo a amostra montada em um estágio piezelétrico, permitindo movimentos laterais com precisão nanométrica16.
NOTA: A armadilha óptica é mantida estacionária.
- Cenários experimentais para a punção da membrana celular
- Colocar HEK293T células, transfectadas com plasmídeos de anexina acoplados a uma proteína fluorescente, sobre AuNPs isoladas que são imobilizadas na superfície de vidro (Figura 1A).
- Utilize um laser de argônio 488 nm para visualizar o sinal de fluorescência GFP e um laser HeNe 633 nm para observar a reflexão AuNP no microscópio confocal de varredura.
- Empregar o tweezer óptico, que opera com um laser NIR de 1064 nm, para irradiar uma única AuNP por ~1 s, induzindo um aumento local significativo da temperatura que rompe a membrana plasmática.
- Aplicar irradiação entre 200-295 mW na partícula focalizada, resultando em um aumento substancial da temperatura.
NOTA: Há uma perda substancial de potência dentro da óptica, com a potência do laser no ponto focal atingindo cerca de 20% do miliwatt declarado, dependendo da configuração específica. A intensidade específica (medida em W/cm2) depende do alinhamento exato do sistema, especificamente do tamanho focal. Além disso, a condução térmica do vidro é maior do que a da célula e da água e, consequentemente, a redução da quantidade de calor liberado para a membrana plasmática é ligeiramente reduzida48,58. - Alcance uma lesão de PM efetiva e localizada e uma resposta de reparo proteica subsequente calibrando adequadamente a localização do ponto de foco do laser NIR. Isso é conseguido capturando uma única AuNP suspensa no mesmo meio de imagem e garantindo que a AuNP selecionada esteja em foco antes da irradiação.
OBS: A nanopartícula é considerada em foco quando seu sinal de espalhamento aparece mais nítido, ou seja, exibindo bordas claras e ausência de halo ao redor da partícula, ao observá-la em microscopia confocal (Figura 2C(ii)).
- Condições de densidade celular e AuNP
- Escolha células únicas em vez de aglomerados de células para evitar a sobreposição de membranas plasmáticas.
OBS: As células devem estar aderidas à superfície, exibindo morfologia achatada (Figura 1 Suplementar), o que permite a punção da periferia celular e evita danos à membrana nuclear (Figura 2 Suplementar). - Incubar as células de acordo com o protocolo ou até que elas se estabeleçam e se achatem para evitar a captação celular de AuNP. Evitar o tempo excessivo de incubação e reduzir a probabilidade de endocitose de AuNP usando AuNPs PEGylated.
- Certifique-se de que as AuNPs imobilizadas estejam presentes como partículas únicas, adequadamente espaçadas entre si para evitar agregados. Os agregados podem levar a um aumento significativo no gradiente térmico, resultando em temperaturas elevadas que podem interromper uma fração substancial da célula.
- Substitua a amostra a cada 1-2 h para manter a saúde celular.
NOTA: A exposição prolongada pode levar a uma deterioração na saúde celular, comprometendo sua capacidade de responder com precisão a lesões em termos de reparo de membranas. Este declínio na saúde celular é tipicamente observado por uma mudança na forma celular, à medida que as células parecem mais esféricas e mais rígidas, culminando eventualmente na morte celular.
- Escolha células únicas em vez de aglomerados de células para evitar a sobreposição de membranas plasmáticas.
Informações Complementares. Clique aqui para baixar este arquivo.
3. Preparação da vesícula unilamelar gigante (GUV)
- Preparo da mistura lipídica
- Completar a composição lipídica do GUV combinando os lipídios 1,2-dioleoil-sn-glicero-3-fosfocolina (DOPC) e 1,2-dioleoil-sn-glicero-3-fosfo-L-serina (DOPS) em uma proporção molar de 4:1 (ver Tabela 1). Aliquotar o estoque lipídico em frascos de vidro de 1,5 mL de acordo com as necessidades experimentais e armazená-los a -20 °C.
NOTA: Para preservação lipídica prolongada e para evitar a oxidação de lipídios insaturados, substitua o ar por argônio nos frascos para injetáveis com alíquotas. - Antes de misturar os lipídios, limpe cuidadosamente uma seringa de vidro ou metal de 50 μL e 500 μL com clorofórmio cinco vezes para garantir que estejam livres de contaminantes para os lipídios dissolvidos de clorofórmio.
CUIDADO: Manuseie o clorofórmio em uma capela devido à sua toxicidade.- Transfira o volume calculado de clorofórmio para um frasco para injetáveis limpo de vidro âmbar, seguido da quantidade especificada de cada lípido (ver Tabela 1).
NOTA: Para evitar a contaminação cruzada entre os estoques lipídicos, certifique-se de que as seringas sejam limpas com clorofórmio. - Finalmente, adicione o corante de membrana e misture bem os lipídios por pipetagem. Conservar a mistura lipídica preparada a -20 °C para utilização posterior; A mistura permanece viável por 2-3 semanas sem danos lipídicos consideráveis.
OBS: A mistura deve ser feita com seringa metálica ou de vidro. Mantenha sempre os lipídios no gelo quando estiverem fora do freezer.
- Transfira o volume calculado de clorofórmio para um frasco para injetáveis limpo de vidro âmbar, seguido da quantidade especificada de cada lípido (ver Tabela 1).
- Completar a composição lipídica do GUV combinando os lipídios 1,2-dioleoil-sn-glicero-3-fosfocolina (DOPC) e 1,2-dioleoil-sn-glicero-3-fosfo-L-serina (DOPS) em uma proporção molar de 4:1 (ver Tabela 1). Aliquotar o estoque lipídico em frascos de vidro de 1,5 mL de acordo com as necessidades experimentais e armazená-los a -20 °C.
- Preparação de gel de álcool polivinílico (PVA)
- Preparar as GUVs usando o método de hidratação assistida por gel descrito por Weinberger et al.23com pequenas modificações.
- Preparar o gel de PVA dissolvendo 5 g de PVA em 100 mL de um tampão contendo 50 mM de sacarose, 25 mM de NaCl e 25 mM de Tris (pH 7,4).
- Aqueça a solução de PVA a 85 °C e mexa até ficar transparente. Deixe esfriar até RT e guarde na geladeira para uso posterior.
NOTA: O PVA não está devidamente dissolvido no tampão e, por conseguinte, é necessário aquecer até 85 °C.
- Preparar as GUVs usando o método de hidratação assistida por gel descrito por Weinberger et al.23com pequenas modificações.
- Preparação de lâminas de vidro
- Limpe as lâminas de vidro com etanol e deixe-as secar ao ar. Em seguida, trate as lâminas com um limpador de plasma de ar para remover qualquer contaminação residual da superfície de vidro.
- Preparação de lâminas de vidro revestidas com PVA
- Aquecer o gel de PVA (5%) até 60 °C durante 30 min para aumentar a sua fluidez. Aplicar 90 μL do PVA quente sobre a lâmina de vidro, espalhar uniformemente e deixar secar em um armário de aquecimento a 50 °C por 50 min.
- Quando a lâmina de vidro PVA estiver pronta, adicione 30 μL da mistura lipídica preparada usando uma seringa de vidro ou metal e espalhe-a em um filme fino usando a borda da agulha.
- Secar a mistura lipídica evaporando seu teor de clorofórmio sob suave pressão de nitrogênio. Além disso, seque as lâminas de vidro sob vácuo por 1,5-2 h.
- Cultivando GUVs em uma câmara
- Montar a câmara interna, semelhante em design a um relatório59 publicado anteriormente, usando a lâmina de vidro preparada.
- Em um tubo separado de 2 mL, diluir a proteína recombinante de interesse (neste caso, ANXA5 ou ANXA4) para uma concentração final de 500 nM com um tampão de crescimento (GB) composto por 80 mM de sacarose, 70 mM de NaCl e 25 mM de Tris-HCl (pH 7,4).
- Adicionar 400 μL da solução proteica recombinante diluída à câmara. Incubar a câmara em TR por 1 h para permitir que os GUVs cresçam a partir da camada lipídica depositada. Use o mesmo tampão, excluindo a proteína, como controle negativo.
NOTA: Embrulhe a câmara em filme de poliolefina para evitar a evaporação do tampão. - Recolher os GUVs transferindo 400 μL do conteúdo da câmara para um tubo de 2 ml.
- Remover proteínas não encapsuladas fora das GUVs adicionando 1 mL de tampão de observação (OB) contendo 55 mM de glicose, 70 mM de NaCl e 50 mM de Tris-HCl (pH 7,4) à solução coletada. Em seguida, centrifugar a solução a 600 x g durante 10 min a 13 °C.
- Após centrifugação, substituir 1 mL do sobrenadante por um volume igual de tampão de observação. Disperse os GUVs através de pipetagem suave e guarde-os na geladeira até que sejam usados em experimentos de GUV.
4. Experimento de punção GUV
- Preparação da câmara
- Use um prato com fundo de vidro de 35 mm para o experimento. Para evitar que os GUVs grudem na superfície e estourem, cubra a superfície com β-caseína (5 mg/mL) por 15-30 min.
- Para a solução de β-caseína, dissolver 0,1 g da proteína em um tampão de 20 mL de Tris 20 mM (PH 7,5) e NaCl 100 mM. Filtre a solução proteica, aliquote-a em pequenos frascos para injetáveis e congele-a para utilização posterior.
- Lavar a câmara duas vezes com tampão de observação para remover qualquer excesso de β-caseínas livres da superfície e deixar secar em RT.
- Em um tubo separado de 2 mL, misture os GUVs coletados com OB. Adicione CaCl2 à mistura para obter a concentração final desejada (neste caso, 200 μM).
- Introduzir nanocascas de ouro de 150 nm (AuNSs) na mistura na proporção de 1:100. A mistura final é composta por 250 μL de GUVs, 225 μL de OB, 20 μL de CaCl2 (5 mM) e 5 μL de AuNSs especificados.
- Use um prato com fundo de vidro de 35 mm para o experimento. Para evitar que os GUVs grudem na superfície e estourem, cubra a superfície com β-caseína (5 mg/mL) por 15-30 min.
- Arranjo experimental
- Transfira a mistura para a câmara e monte-a no palco do microscópio. Dependendo do tamanho da câmara, adicione a mistura inteira ou uma porção da mistura.
NOTA: O tempo é crítico, uma vez que os iões de cálcio podem atravessar a membrana e mediar a ligação das anexinas ao folheto interno da membrana. - Empregar a mesma configuração óptica usada para os experimentos de punção celular.
- Use a pinça óptica para prender um AuNS individual nas proximidades ou na superfície de um GUV aplicando uma potência de laser de ~ 125 mW.
- Posteriormente, aumente a potência do laser para ~ 300 mW. Isso produz um aumento de temperatura altamente localizado, que interrompe e perfura a membrana no local alvo.
NOTA: AuNSs são preferidos para os experimentos GUV devido à sua capacidade de gerar um aumento de temperatura transiente mais alto, mantendo um tamanho menor em comparação com AuNPs sólidos.
- Transfira a mistura para a câmara e monte-a no palco do microscópio. Dependendo do tamanho da câmara, adicione a mistura inteira ou uma porção da mistura.

Tabela 1: Tabela para determinação da composição do GUV. Clique aqui para baixar esta tabela.
5. Medidas e análise de dados da resposta do ANXA em experimentos de punção celular
- Utilizar o MATLAB para analisar as imagens e calcular e plotar o perfil de temperatura AuNP (Figura 1D, E) com base nas equações de Mie60,61.
- Além disso, processe todas as imagens confocais usando a distribuição FIJI ImageJ62,63.
- Cálculo do raio do anel ANXA
- Cortar a região de interesse que contém a área puncionada a partir dos dados brutos antes da análise da lesão por membrana.
- Utilize o fluxo de trabalho interno do MATLAB para processar cada imagem, marcando manualmente o centro do anel ANXA e seu perímetro externo.
- Use essas marcações para definir os limites de processamento para a região de interesse.
NOTA: A área é subsequentemente processada radialmente dentro dos limites definidos, e a intensidade da fluorescência a uma distância específica do centro é a média sobre o círculo completo com a mesma distância central. Esse número é armazenado para cada etapa de raio. - Use o fluxo de trabalho para ajustar as intensidades médias em todo o intervalo de varredura a uma curva Gaussiana unidimensional para identificar os máximos (Rp) e a largura total na metade máxima (FWHM) correspondentes a Rext e Rint, respectivamente, conforme ilustrado na Figura 3A.
- Use essas marcações para definir os limites de processamento para a região de interesse.
- Finalmente, determine o raio do anel ANXA pela distância do centro do anel aoramal R dada pelo gráfico gerado pelo fluxo de trabalho do MATLAB.
- Cálculo do raio do anel ANXA tridimensional
- Acompanhe a alteração no raio do anel ANXA na direção z usando o mesmo fluxo de trabalho de análise MATLAB da etapa 5.1. Aplique o fluxo de trabalho a várias z-seções confocais em toda a ferida da membrana, conforme ilustrado na Figura 3C, E.
- Calcular a inclinação de cada ferida com base na mudança dos raios sobre as seções z, conforme ilustrado na Figura 3F.
- Evolução temporal do raio do anel ANXA
- Da mesma forma, acompanhe a evolução do anel ANXA do anel ANXA após a punção da membrana termoplasmônica usando o fluxo de trabalho de análise MATLAB da etapa 5.1. Analise pontos de tempo consecutivos para monitorar as mudanças ao longo do tempo, conforme demonstrado na Figura 3D, G, H.
Resultados
No presente estudo, o método termoplasmônico é utilizado para investigar a resposta da proteína anexina à ruptura da membrana plasmática; no entanto, qualquer proteína que possa ser recrutada após lesão de membrana pode ser investigada usando este ensaio, uma vez que a respectiva proteína é marcada fluorescentemente. O recrutamento e a função de proteínas são monitorados por imagens confocais em células de rim embrionário humano (HEK293T) e vesículas unilamelares gigantes (GUVs). Para elaborar, a Figura 2 ilustra as condições experimentais em que um feixe de laser NIR focalizado a 1064 nm é usado para irradiar uma única AuNP (Figura 2A), resultando em lesão da membrana e um influxo de Ca2+ para dentro da célula, ativando a maquinaria PMR. Posteriormente, as anexinas são rapidamente recrutadas para o local da lesão, onde se ligam aos fosfolipídios carregados negativamente no perímetro da ferida, formando uma estrutura em forma de anel em segundos (Figura 2B, i-iv). Os experimentos de membrana modelo, usando GUVs, demonstraram que as punções de membrana foram rapidamente seladas na ausência de ANXA, como mostrado na Figura 2C. Entretanto, na presença de ANXA, observou-se rápido acúmulo de ANXA no local da lesão após a punção da membrana (Figura 2D). Notavelmente, ANXA continuou a rolar as bordas expostas, levando ao estouro do GUV. Acredita-se que esse mecanismo de rolamento decorra da capacidade da ANXA de induzir curvatura e dobrar as membranas lipídicas20.

Figura 2: Resposta das anexinas (ANXAs) à ruptura da membrana induzida por termoplasmônicos. Inicialmente, (A) a membrana plasmática atua como uma barreira entre o meio extracelular contendo altos níveis de íons Ca2+ e o meio intracelular com ANXAs encapsulados. Após a irradiação com um laser de infravermelho próximo (NIR), o AuNP gera calor substancial, causando a ruptura da membrana e resultando em um influxo de íons Ca2+. Consequentemente, a maquinaria de reparo da membrana plasmática (PMR) é ativada, o que envolve o recrutamento de ANXAs para o local da lesão, onde eles se ligam a fosfolipídios carregados negativamente. (B-D) Imagens de microscopia confocal de uma célula contendo ANXA2 e um GUV contendo ANXA4 demonstram esse processo. (i) Antes da irradiação, as imagens mostram uma célula intacta, ou GUV, com os locais de irradiação denotados pelas setas brancas. (ii) Após a irradiação com nanopartículas, (B) os ANXAs são rapidamente recrutados para o local da lesão, formando uma estrutura em forma de anel ao redor da ferida da membrana (B [ii-iv]). O painel (C) mostra um GUV corado pela membrana sem ANXA, que, após a punção, resela rapidamente sem remodelamento observável da membrana. Por outro lado, o painel (D) mostra um GUV contendo ANXA4 recombinante, que já está ligado à membrana antes da (i) irradiação devido ao vazamento de Ca2+ através da membrana. (ii) Após a punção, ANXA4 liga-se às bordas livres, fazendo com que o GUV entre em colapso à medida que a membrana se afasta da borda. As barras de escala medem 10 μm para a figura (B), 2 μm para B (i), 10 μm para (C) e 15 μm para (D). Esta figura é reproduzida de Moreno-Pescador et. al16 com permissão da Royal Society of Chemistry. Clique aqui para ver uma versão maior desta figura.
{kind=link}
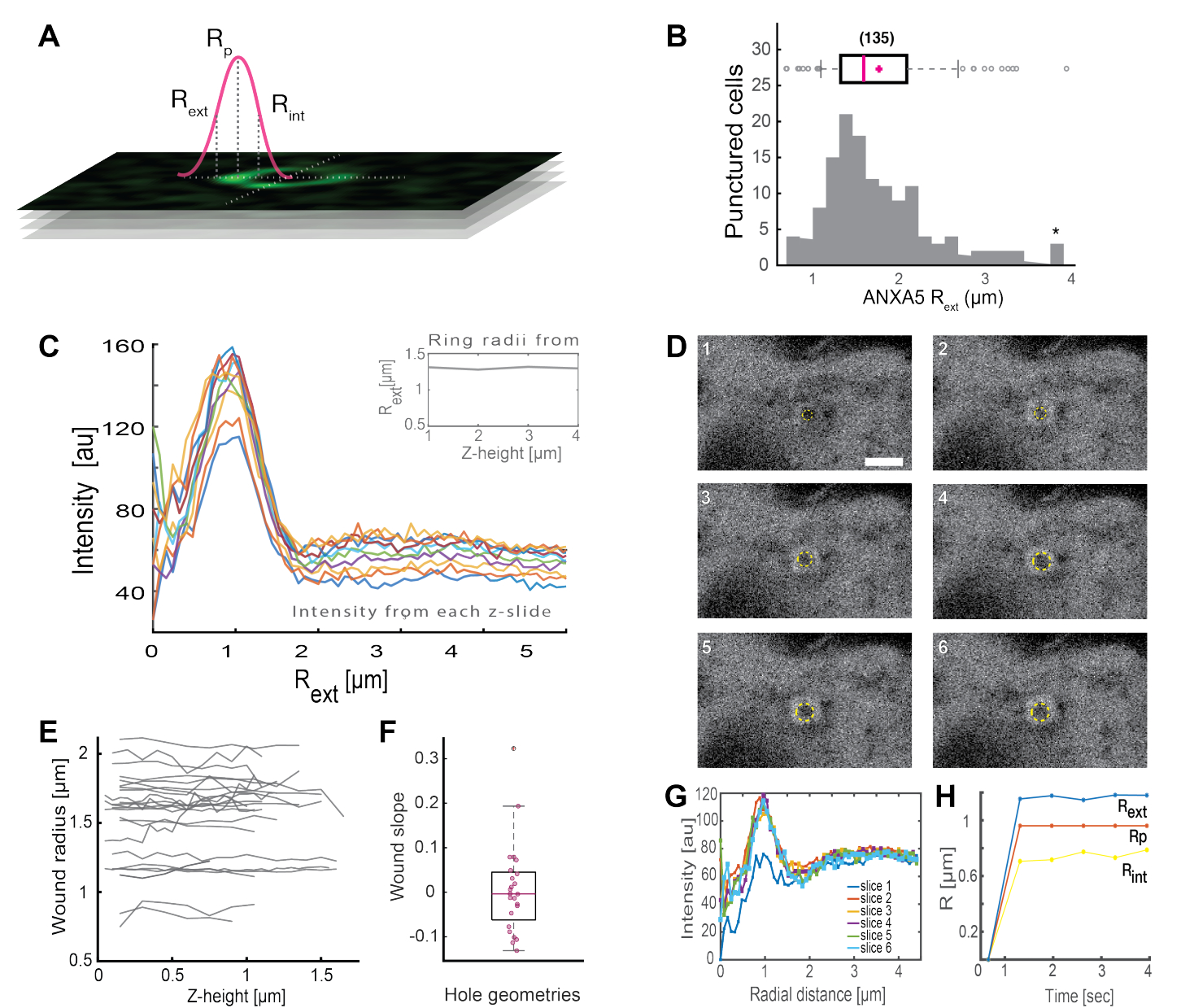
A análise de estruturas anpilares ANXA completas (Figura 2B e Figura 1 Suplementar) fornece informações úteis sobre o tamanho e a morfologia da ferida. O raio da estrutura do anel ANXA pode ser determinado ao longo do tempo e do espaço, conforme descrito na Seção 5. Moreno et al.16 analisaram mais de 135 lesões da membrana plasmática em células vivas, onde a análise das estruturas do anel ANXA5 está representada na Figura 3. O raio foi determinado medindo-se a distância do centro do anel ao raio externo da curva com base na largura total-metade-máxima do perfil de intensidade colapsada (Figura 3A). Os achados ilustraram uma distribuição heterogênea dos tamanhos dos anéis ANXA5 (Figura 3B), que se manteve constante ao longo do tempo (Figura 3D,G,H) e do espaço (Figura 3C,E,F). Esses resultados sugerem um acúmulo de ANXA5 ao redor dos locais de lesão, indicativo de uma estratégia alternativa de RPM mediada por ANXA5 em células vivas ao hipotético brotamento interno semelhante a um funil da membrana lesada5.

Figura 3: Análise das estruturas dos anéis ANXA5 ao redor de um local de lesão em células vivas. (A) A representação esquemática ilustra a abordagem analítica, que se baseia no máximo de largura total e metade do perfil da linha de intensidade ANXA. (B) O histograma ilustra 135 raios do anel ANXA5 medidos. (C) Perfis de linha de intensidade de fluorescência através de um anel ANXA5-GFP para cada seção z da pilha z. (D) As imagens confocais ilustram a evolução temporal de uma ferida, onde ANXA5-GFP se acumula no perímetro da ferida imediatamente após a lesão, seguido de estabilização da ferida. Os períodos de tempo rotulados de 1 a 6 foram capturados em intervalos de 0,66 s por quadro. A barra de escala é de 2 μm. (E) Os raios das feridas foram determinados em função da profundidade da ferida com base no método apresentado no painel A. (F) A inclinação da ferida foi analisada como raios do anel ANXA5 em função da altura z com base nos dados extraídos do painel E. (G) Os perfis de linha de intensidade de fluorescência foram medidos através do anel ANXA5-GFP do painel D, onde cada intervalo de tempo 1-6 é denotado como fatia 1-6. Finalmente, (H) a evolução temporal dos raios do anel ANXA5-GFP é apresentada em função do tempo calculado a partir dos dados do painel G. Esta figura é reproduzida de Moreno-Pescador et. al16 com permissão da Royal Society of Chemistry. Clique aqui para ver uma versão maior desta figura.
{kind=link}
Em comparação com as estruturas anelares formadas pela ruptura isolada do PM (Figura 1 Suplementar), lesões próximas ao núcleo (Figura 2A Suplementar) podem influenciar a evolução e a geometria da ferida. Ocasionalmente, apenas uma fração dos anéis ANXA foram evidentes (Figura 2B,C Suplementar), que também pode ser analisada usando o fluxo de trabalho interno de análise MATLAB (ver Seção 5), embora dados adicionais possam ser perdidos. Tipicamente, as formações de anéis ANXA observadas em células não aderentes (Figura 2C Suplementar) estão localizadas próximas ao núcleo e à periferia da célula. Como consequência, pode-se observar uma estrutura de anel mais alongada, o que é subótimo para a análise dos dados apresentados. Além disso, as células não aderentes parecem ser mais suscetíveis à morte celular após a lesão de PM. Além disso, ao considerar as lesões resultantes da irradiação de agregados de AuNP, é importante ressaltar que essas lesões podem ser mais graves e menos controláveis. Isso se deve ao aumento significativo do aquecimento plasmônico, que pode danificar uma grande fração da célula. Como resultado, tais lesões não foram incorporadas à análise do anel ANXA5.
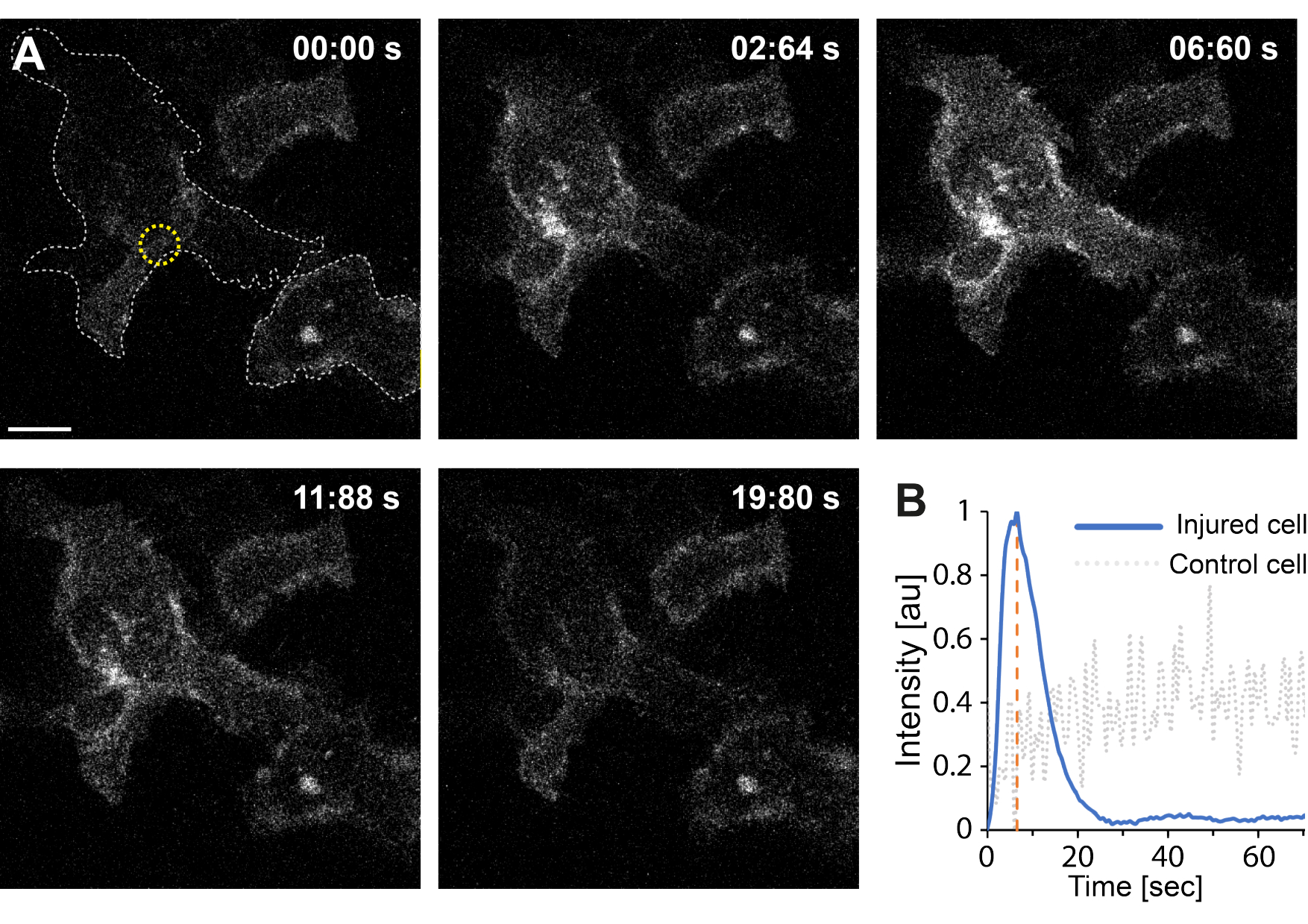
Além disso, os resultados preliminares indicam que a ruptura da membrana plasmática usando termoplasmônica resulta em níveis elevados de Ca2+ intracelular. Isso foi observado mesmo com irradiação de baixa intensidade de AuNPs isoladas, sugerindo permeabilização de MP35, como mostra a Figura 4. O influxo de Ca2+ foi observado em células que expressam uma sonda de cálcio ligada à membrana, GCaMP6s-CAAX, que sofre mudança conformacional com o influxo de Ca2+ , podendo-se observar, assim, um aumento na sua intensidade64. A intensidade de cálcio foi quantificada para toda a impressão plantar da célula ao longo do tempo. Para eliminar o ruído de fundo, o nível de Ca2+ de fundo foi subtraído antes da ruptura da membrana e do reparo pós-membrana. A intensidade máxima foi determinada normalizando a intensidade média de Ca2+ dentro da célula, resultando em uma curva de intensidade exibindo um rápido aumento inicial da intensidade de Ca2+ seguido por uma diminuição mais lenta, como mostrado na Figura 4B.
A célula atingiu uma intensidade máxima de cálcio em ~ 6,6 s, o que é consistente com os achados de Klenow et al.64, que sugeriram que o tempo do pico de intensidade de cálcio (t = tc) corresponde ao tempo necessário para o fechamento da ferida. No entanto, embora mais investigações sejam necessárias para estabelecer o mecanismo subjacente de reparo da membrana e cicatrização de feridas, os resultados preliminares mostraram que esse processo de Ca2+ foi observado exclusivamente na célula lesada e não na célula não lesada usada como controle positivo. Isso confirma que a célula experimentou influxo de Ca2+ após a ruptura da membrana termoplasmônica, onde o excesso de cálcio intracelular é ativamente bombeado para fora após o sucesso da RPM, pois o nível de cálcio intracelular não está mais em competição com o influxo de Ca2+ , eventualmente atingindo a homeostática celular64.

Figura 4: O influxo de cálcio ocorre quando a membrana plasmática de uma célula HEK293T é rompida pela termoplasmônica. Uma série de imagens confocais mostra duas células (a célula de interesse e uma célula não lesada usada como controle positivo) expressando a sonda de cálcio ligada à membrana, GCaMP6s-CAAX. A barra de escala mede 10 μm. (A) Antes da irradiação, às 00:00 s, a impressão das duas células é visualizada pela linha pontilhada cinza, e o local da irradiação é denotado pelo círculo amarelo. À irradiação com laser, observa-se um rápido influxo de Ca2+ , atingindo intensidade máxima de ~ 6,6 s, denotada pela linha tracejada laranja, momento que se presume corresponder ao tempo de fechamento da ferida64. (B) O perfil de intensidade de cálcio obtido da sonda GCaMP6s-CAAX na célula lesada (linha azul) foi comparado com a intensidade de Ca2+ em uma célula vizinha não lesada (linha pontilhada cinza), mostrando um influxo claro de Ca2+ exclusivamente após a ruptura do PM. Clique aqui para ver uma versão maior desta figura.
{kind=link}
Discussão
O estudo destaca a abordagem termoplasmônica como uma técnica promissora para explorar respostas de proteínas em células vivas e membranas modelo após ruptura de membrana. Este método não só fornece extensa informação sobre o recrutamento de proteínas, mas também sobre a função biofísica de proteínas envolvidas na dinâmica proteína-membrana. Consequentemente, facilita a identificação de componentes moleculares responsáveis pelo reparo de superfície e avança na compreensão da complexa, mas vital maquinaria de reparo da membrana plasmática. Embora existam vários métodos para induzir ruptura de membrana, como mecânica, química e óptica, esses métodos sofrem limitações, como não serem específicos das células, gerar múltiplas lesões na membrana celular ou causar danos significativos à membrana e ablar material celular interno ao longo do trajeto do laser quando se utilizam lasers pulsados de alta potência. Embora a integração de microscopia confocal e pinça óptica ofereça as informações mais abrangentes, modalidades alternativas de imagem também podem ser usadas. Por exemplo, como a imagem da nanopartícula plasmônica é obtida usando microscopia de reflexão, um modo de imagem incorporado em microscópios confocais Leica, técnicas de imagem adicionais, como microscopia de campo escuro65,66, outros métodos de espalhamento como iSCAT 67,68, ou marcação fluorescente da nanopartícula, poderiam ser empregadas para visualização de AuNP, embora isso possa limitar a aplicabilidade do método.
O método apresentado é adicionalmente capaz de induzir orifícios nanoscópicos em membranas modelo, permitindo a investigação dos efeitos sinérgicos entre diferentes anexinas. Isso é conseguido encapsulando anexinas recombinantes marcadas de forma diferente, por exemplo, RFP e GFP, respectivamente, seguidas de punção termoplasmônica. Este sistema modelo fornece informações sobre como as anexinas interagem com membranas nas proximidades das bordas livres, como demonstrado na Figura 2D. No entanto, ao contrário das células, os orifícios infligidos aos GUVs continuam a se expandir, seguidos de desestabilização da vesícula. A obtenção de imagens da evolução do orifício usando microscopia confocal pode ser desafiadora devido à rápida expansão do diâmetro do furo, mas pode ser obtida capturando várias pilhas z ao longo do tempo. Um método alternativo seria usar um disco giratório confocal para imagens mais rápidas. Além disso, a abordagem termoplasmônica normalmente produz um número limitado de resultados ótimos por hora quando aplicada a células únicas ou experimentos de GUV, geralmente dois a três, em temperaturas de amostra entre 20 °C e 30 °C. Para obter a observação mais precisa da dinâmica proteína-membrana, recomenda-se manter as células em um tampão contendo HEPES e substituir a amostra a cada hora. Alternativamente, a janela experimental poderia ser estendida realizando-se os experimentos em uma câmara de incubação celular, ou seja, a uma temperatura constante de 37 °C com 5% de CO2. Além disso, a combinação dessa abordagem com outras técnicas de imagem, como a microscopia de reconstrução óptica estocástica (STORM), poderia fornecer uma compreensão mais profunda da função biofísica e da interação de proteínas-chave envolvidas no reparo de membrana em nível de molécula única. Isso poderia fornecer informações detalhadas sobre o local da lesão, incluindo a geometria da ferida e a localização das proteínas da anexina, bem como identificar outros atores-chave envolvidos no reparo da superfície da membrana.
Para alcançar a máxima eficácia e precisão na indução de lesão de membrana, é imperativo verificar a localização do foco do laser antes de cada experimento e garantir que a posição axial do foco do laser coincida com o foco confocal. Esse alinhamento otimiza a intensidade durante a aquisição de AuNP, o que leva a um aumento máximo da temperatura local e consequente lesão da membrana com menor potência do laser. Este processo é executado manualmente e, portanto, suscetível à variabilidade na eficiência de ruptura da membrana, pois o foco é traduzido manualmente para uma posição que coincide com a localização da partícula. Em microscópios que não possuem um modo de reflexão, como em alguns sistemas comerciais, a co-localização do foco do laser e da partícula pode ser um desafio. Nesses casos, modos de imagem alternativos (por exemplo, campo brilhante) podem ser empregados, e uma varredura raster lenta pode ser realizada em torno da posição esperada da partícula. Deve-se notar que a baixa potência do laser provavelmente induzirá apenas a permeabilização da membrana, enquanto a alta potência do laser pode gerar temperaturas ao redor do NP que excedem o ponto de ebulição da água, mesmo que a superfície do vidro tenha um efeito de resfriamento. Estima-se que a formação de nanobolhas ao redor dos NPs ocorra entre 200 °C e 300 °C25,48, onde o calor explosivo pode resultar tanto no deslocamento das partículas do foco do laser quanto na fragmentação das partículas. Além disso, a formação de nano ou microbolhas durante o aquecimento representa um desafio para este método. Como as interfaces de ar desumedecem as membranas e podem causar desestabilização proteica, o que é indesejável, é imperativo limitar o aquecimento ao investigar o reparo da membrana. Notavelmente, nanocascas de ouro não toleram altas temperaturas e se degradam nessas condições, como demonstrado por microscopia de alta resolução58.
Este artigo fornece um protocolo detalhado para o uso de termoplasmônicos para realizar punções altamente localizadas em membranas, que é aplicável tanto a células quanto a membranas modelo. Para reduzir ainda mais a extensão do aquecimento, nanopartículas menores ressonantes com luz NIR podem ser utilizadas, permitindo punções intracelulares em endossomos, no retículo endoplasmático e no envelope nuclear. Tais nanopartículas, incluindo bastões e nanomatryoshkas48, podem ser usadas para investigar o reparo do envelope nuclear visando nanopartículas de ouro endocitosadas que são prontamente absorvidas na superfície celular e trafegadas em direção ao núcleo69. De modo geral, essa técnica permite a identificação e o exame dos principais componentes moleculares envolvidos na RPM, elucidando sua função e papel biofísicos, preservando a viabilidade das células.
Divulgações
Os autores não têm nada a revelar.
Agradecimentos
Gostaríamos de agradecer a Jesper Nylandsted por nos fornecer proteínas recombinantes de anexina e plasmídeos que codificam para anexinas. Este trabalho foi financiado pelo Conselho Dinamarquês de Pesquisa Independente, Ciências Naturais (DFF-4181-00196), pelo Programa de Sinergia Interdisciplinar da Fundação Novo Nordisk 2018 (NNF18OC0034936), pelo Comitê Científico da Sociedade Dinamarquesa de Câncer (R90-A5847-14-S2), pela Fundação Lundbeck (R218-2016-534) e pelo Centro de Excelência da Fundação Lundbeck (Biomembranas em Nanomedicina).
Materiais
| Name | Company | Catalog Number | Comments |
| 1064 nm trapping laser | Spectra Physics | N/A | Spectra Physics J201-BL-106C, Nd: YVO4 NIR laser |
| 160 nm Gold Nanoshells | NanoComposix | NCXGSIR150 | |
| 200 nm Gold Nanoparticles | BBI Solutions | EM.GC200/7 | |
| 35 mm glass surface MatTex microwell | MATTEK | P35G-1.5-14-C | |
| Amber-glass vials | Supelco Sigma Aldrich | 243438 | |
| Annexin A2 plasmids | N/A | N/A | Received from our collaborator at the Danish Cancer Research Center |
| Annexin A4 recombinant-protein | N/A | N/A | N-terminal GFP tagged ANXA4 received from our collaborator at the Danish Cancer Research Center |
| Annexin A5 recombinant-protein | N/A | N/A | N-terminal GFP tagged ANXA5 received from our collaborator at the Danish Cancer Research Center |
| beta-casein | Sigma Life Science | C6905-1G | |
| CaCl2 | Suprlco (sigma Aldrich) | 10035-04-8 | |
| Centrifuge 5702 | Eppendorf | 5702 | |
| Chloroform | VWR Chemicals | 67-66-3 | |
| Culture dish (Nunclon Delta Surface) | Thermo scientific | 150460 | |
| DID cell-labelling Solution | Invitrogen | 7757 | |
| Distilled water | Gibco | 15230-089 | |
| DOPC | Avanti Polar Lipids | 850375C | Dissolved in chloroform |
| DOPS | Avanti Polar Lipids | 840035C | Dissolved in chloroform |
| Dulbecco's Modified Eage's Medium | Thermo Fisher Scientific | 11995065 | |
| FIJI ImageJ distribution | ImageJ2 | N/A | |
| GCaMP6s-CAAX | N/A | Received from our collaborator at the Danish Cancer Research Center | |
| Gibco Fetal Bovine Serum | Fisher Scientific | 11573397 | 10% of the culture medium |
| Glucose | PROLABO | 24 374.297 | |
| Hamilton syringes | Hamilton Company | N/A | 50 and 500 microliters |
| Harrick Plasma Cleaner PDG-002 | Harrick Plasma | N/A | |
| HEK293T cells | N/A | Received from our collaborator at the Danish Cancer Research Center | |
| Leica Acousto-Optical Beam Splitter (AOBS) | Leica | N/A | |
| Leica PL APO 63x water immersion objective, NA = 1.2 | Leica | N/A | |
| Leica SP5 confocal scanning microscope | Leica | N/A | |
| Lipofectamine | Fisher Scientific | 15338030 | |
| MatLab | The Mathworks, Inc., Natick, Massachusetts, United States | N/A | |
| NaCl | VWR Chemicals | 7647-14-5 | |
| Opti-MEM Reduced-Serum Medium | Thermo Fisher Scientific | 11058021 | |
| Parafilm | Bemis | PM-992 | |
| Penicillin-Streptomycin | Thermo Fisher Scientific | 15140122 | 1% of the culture medium |
| Phosphate Buffered Saline (PBS) | Thermo Fisher Scientific | 10010023 | |
| Piezoelectric stage (PI 731.20) | Physik Instrumente (Germany) | N/A | |
| Poly-L-Lysine | Sigma-Aldrich | P8920-100ML | 0.01-0.1% for coating |
| Polyvinyl alcohol | Sigma-Aldrich | 363065-25G | |
| round glass slide 25 mm Ø | VWR | 631-1584 | |
| Sonicator Brandson 2800 | Brandson | N/A | |
| sucrose | Sigma Life Science | 57-50-1 | |
| T25 tissue culture flask | Falcon | 353108 | Blue Vented cap |
| Tris-HCl | Invitrogen | 15567-027 | |
| TrypLE | Thermo Fisher Scientific | A1285901 | |
| Trypsin-EDTA | Fisher Scientific | 11590626 | |
| VWR Mixer mini vortex 230V EU | VWR | 12620-84 | ECN: 444-2790, SN: 150713022 |
Referências
- Bendix, P. M., et al. Interdisciplinary synergy to reveal mechanisms of annexin-mediated plasma membrane shaping and repair. Cells. 9 (4), 1029 (2020).
- Gajic, O., Lee, J., Doerr, C. H., Berrios, J. C., Myers, J. L., Hubmayr, R. D. Ventilator-induced Cell Wounding and Repair in the Intact Lung. American Journal of Respiratory and Critical Care Medicine. 167, 1057-1063 (2003).
- McNeil, P. L., Khakee, R. Disruptions of muscle fiber plasma membranes. Role in exercise-induced damage. The American Journal of Pathology. 140 (5), 1097-1109 (1992).
- Yu, Q. C., McNeil, P. L. Transient disruptions of aortic endothelial cell plasma membranes. The American Journal of Pathology. 141 (6), 1349-1360 (1992).
- Boye, T. L., et al. Annexin A4 and A6 induce membrane curvature and constriction during cell membrane repair. Nature Communications. 8, 1623 (2017).
- Bischofberger, M., Gonzalez, M. R., van der Goot, F. G. Membrane injury by pore-forming proteins. Current Opinion in Cell Biology. 21, 589-595 (2009).
- Tang, S. K. Y., Marshall, W. F. Self-repairing cells. Science (New York, N.Y.). 356, 1022-1025 (2017).
- Abreu-Blanco, M. T., Verboon, J. M., Parkhurst, S. M. Single cell wound repair: Dealing with life's little traumas. Bioarchitecture. 1, 114-121 (2011).
- Sønder, S. L., et al. Annexin A7 is required for ESCRT III-mediated plasma membrane repair. Scientific Reports. 9, 6726 (2019).
- Andrews, N. W., Almeida, P. E., Corrotte, M. Damage control: cellular mechanisms of plasma membrane repair. Trends in Cell Biology. 24 (12), 734-742 (2014).
- Idone, V., Tam, C., Goss, J. W., Toomre, D., Pypaert, M., Andrews, N. W. Repair of injured plasma membrane by rapid Ca2+-dependent endocytosis. The Journal of Cell Biology. 180 (5), 905-914 (2008).
- Lauritzen, S. P., Boye, T. L., Nylandsted, J. Annexins are instrumental for efficient plasma membrane repair in cancer cells. Seminars in Cell & Developmental Biology. 45, 32-38 (2015).
- Häger, S. C., Nylandsted, J. Annexins: players of single cell wound healing and regeneration. Communicative & Integrative Biology. 12 (1), 162-165 (2019).
- Jaiswal, J. K., et al. S100A11 is required for efficient plasma membrane repair and survival of invasive cancer cells. Nature Communications. 5, 3795 (2014).
- Draeger, A., Monastyrskaya, K., Babiychuk, E. B. Plasma membrane repair and cellular damage control: The annexin survival kit. Biochemical Pharmacology. 81 (6), 703-712 (2011).
- Moreno-Pescador, G. S., et al. Thermoplasmonic nano-rupture of cells reveals annexin V function in plasma membrane repair. Nanoscale. 14 (21), 7778-7787 (2022).
- Zhivotovsky, B., Orrenius, S. Calcium and cell death mechanisms: A perspective from the cell death community. Cell Calcium. 50 (3), 211-221 (2011).
- Gerke, V., Moss, S. E. Annexins: From structure to function. Physiological Reviews. 82 (2), 331-371 (2002).
- Idone, V., Tam, C., Andrews, N. W. Two-way traffic on the road to plasma membrane repair. Trends in Cell Biology. 18 (11), 552-559 (2008).
- Boye, T. L., et al. Annexins induce curvature on free-edge membranes displaying distinct morphologies. Scientific Reports. 8, 10309 (2018).
- Bouter, A., et al. Annexin-A5 assembled into two-dimensional arrays promotes cell membrane repair. Nature Communications. 2, 270 (2011).
- Boye, T. L., Nylandsted, J. Annexins in plasma membrane repair. Biological Chemistry. 397 (10), 961-969 (2016).
- Weinberger, A., et al. Gel-assisted formation of giant unilamellar vesicles. Biophysical Journal. 105 (1), 154-164 (2013).
- Numata, T., Tatsuta, H., Morita, Y., Otani, Y., Umeda, N. Localized thermal processing with a laser-trapped and heated metal nanoparticle. IEEJ Transactions on Electrical and Electronic Engineering. 2, 398-401 (2007).
- Bendix, P. M., Reihani, S. N. S., Oddershede, L. B. Direct measurements of heating by electromagnetically trapped gold nanoparticles on supported lipid bilayers. ACS Nano. 4 (4), 2256-2262 (2010).
- Kyrsting, A., Bendix, P. M., Stamou, D. G., Oddershede, L. B. Heat profiling of three-dimensionally optically trapped gold nanoparticles using vesicle cargo release. Nano Letters. 11 (2), 888-892 (2011).
- Andersen, T., Kyrsting, A., Bendix, P. M. Local and transient permeation events are associated with local melting of giant liposomes. Soft Matter. 10 (24), 4268-4274 (2014).
- Bahadori, A., Oddershede, L. B., Bendix, P. M. Hot-nanoparticle-mediated fusion of selected cells. Nano Research. 10, 2034-2045 (2017).
- Rørvig-Lund, A., Bahadori, A., Semsey, S., Bendix, P. M., Oddershede, L. B. Vesicle fusion triggered by optically heated gold nanoparticles. Nano Letters. 15 (6), 4183-4188 (2015).
- Moreno-Pescador, G., Arastoo, M. R., Ruhoff, V. T., Chiantia, S., Daniels, R., Bendix, P. M. Thermoplasmonic vesicle fusion reveals membrane phase segregation of influenza spike proteins. Nano Letters. 23 (8), 3377-3384 (2023).
- Bahadori, A., Lund, A. R., Semsey, S., Oddershede, L. B., Bendix, P. M. Controlled cellular fusion using optically trapped plasmonic nano-heaters. SPIE Proceedings. SPIE 9922, Optical Trapping and Optical Micromanipulation XIII. 992211, (2016).
- Bahadori, A., Moreno-Pescador, G., Oddershede, L. B., Bendix, P. M. Remotely controlled fusion of selected vesicles and living cells: a key issue review. Reports on Progress in Physics. 81 (3), 32602 (2018).
- Moreno-Pescador, G., Arastoo, M. R., Chiantia, S., Daniels, R., Bendix, P. M. Thermoplasmonic induced vesicle fusion for investigating membrane protein phase affinity. bioRxiv. , (2022).
- Pescador, G. S. M., et al. Investigating plasma-membrane repair employing thermoplasmonics. Biophysical Journal. 120 (3), 45A (2021).
- Moreno-Pescador, G. S., Qoqaj, I., Thusgaard Ruhoff, V., Iversen, J., Nylandsted, J., Bendix, P. M. Effect of local thermoplasmonic heating on biological membranes. SPIE 11083, Optical Trapping and Optical Micromanipulation XVI. 110830M, (2019).
- Bement, W. M., Mandato, C. A., Kirsch, M. N. Wound-induced assembly and closure of an actomyosin purse string in Xenopus oocytes. Current Biology. 9 (11), 579-587 (1999).
- Weisleder, N., et al. Recombinant MG53 protein modulates therapeutic cell membrane repair in treatment of muscular dystrophy. Science Translational Medicine. 4 (139), 139ra85 (2012).
- Sudji, I. R., Subburaj, Y., Frenkel, N., García-Sáez, A. J., Wink, M. Membrane disintegration caused by the steroid saponin digitonin is related to the presence of cholesterol. Molecules. 20 (11), 20146-20160 (2015).
- Babiychuk, E. B., Monastyrskaya, K., Potez, S., Draeger, A. Intracellular Ca2+ operates a switch between repair and lysis of streptolysin O-perforated cells. Cell Death & Differentiation. 16, 1126-1134 (2009).
- Nygård Skalman, L., Holst, M. R., Larsson, E., Lundmark, R. Plasma membrane damage caused by listeriolysin O is not repaired through endocytosis of the membrane pore. Biology Open. 7 (10), bio035287 (2018).
- Swaggart, K. A., et al. Annexin A6 modifies muscular dystrophy by mediating sarcolemmal repair. Proceedings of the National Academy of Sciences of the United States of America. 111, 6004-6009 (2014).
- Yeheskely-Hayon, D., Minai, L., Golan, L., Dann, E. J., Yelin, D. Optically induced cell fusion using bispecific nanoparticles. Small. 9 (22), 3771-3777 (2013).
- Minai, L., Yeheskely-Hayon, D., Golan, L., Bisker, G., Dann, E. J., Yelin, D. Optical nanomanipulations of malignant cells: Controlled cell damage and fusion. Small. 8 (11), 1732-1739 (2012).
- Lukianova-Hleb, E., et al. Plasmonic nanobubbles as transient vapor nanobubbles generated around plasmonic nanoparticles. ACS Nano. 4 (4), 2109-2123 (2010).
- Vogel, A., Noack, J., Hüttman, G., Paltauf, G. Mechanisms of femtosecond laser nanosurgery of cells and tissues. Applied Physics B. 81, 1015-1047 (2005).
- Baffou, G., Polleux, J., Rigneault, H., Monneret, S. Super-heating and micro-bubble generation around plasmonic nanoparticles under cw illumination. Journal of Physical Chemistry C. 118 (9), 4890-4898 (2014).
- Sasikumar, K., Liang, Z., Cahill, D. G., Keblinski, P. Curvature induced phase stability of an intensely heated liquid. Journal of Chemical Physics. 140 (23), 234506 (2014).
- Jauffred, L., Samadi, A., Klingberg, H., Bendix, P. M., Oddershede, L. B. Plasmonic heating of nanostructures. Chemical Reviews. 119 (13), 8087-8130 (2019).
- Neuman, K. C., Nagy, A. Single-molecule force spectroscopy: optical tweezers, magnetic tweezers and atomic force microscopy. Nature Methods. 5 (6), 491-505 (2008).
- Bendix, P. M., Jauffred, L., Norregaard, K., Oddershede, L. B. Optical trapping of nanoparticles and quantum dots. IEEE Journal of Selected Topics in Quantum Electronics. 20, 15-26 (2014).
- Samadi, A., Bendix, P. M., Oddershede, L. B. Optical manipulation of individual strongly absorbing platinum nanoparticles. Nanoscale. 46, 18449-18455 (2017).
- Jørgensen, J. T., Norregaard, K., Tian, P., Bendix, P. M., Kjaer, A., Oddershede, L. B. Single particle and PET-based platform for identifying optimal plasmonic nano-heaters for photothermal cancer therapy. Scientific Reports. 6, 30076 (2016).
- Goldenberg, H., Tranter, C. J. Heat flow in an infinite medium heated by a sphere. British Journal of Applied Physics. 3 (9), 296-298 (1952).
- Eustis, S., El-Sayed, M. A. Why gold nanoparticles are more precious than pretty gold: Noble metal surface plasmon resonance and its enhancement of the radiative and nonradiative properties of nanocrystals of different shapes. Chemical Society Reviews. 35, 209-217 (2006).
- Landau, L. D., Lifshitz, E. M. . Fluid Mechanics: Landau and Lifshitz: Course of Theoretical Physics. 6, (2013).
- Niederauer, C., Seynen, M., Zomerdijk, J., Kamp, M., Ganzinger, K. A. The K2: Open-source simultaneous triple-color TIRF microscope for live-cell and single-molecule imaging. HardwareX. 13, e00404 (2023).
- Richardson, A. C., Reihani, N., Oddershede, L. B. Combining confocal microscopy with precise force-scope optical tweezers. SPIE Proceedings:SPIE 6326, Optical Trapping and Optical Micromanipulation III. 632628, (2006).
- Samadi, A., Klingberg, H., Jauffred, L., Kjær, A., Bendix, P. M., Oddershede, L. B. Platinum nanoparticles: a non-toxic, effective and thermally stable alternative plasmonic material for cancer therapy and bioengineering. Nanoscale. 10 (19), 9097-9107 (2018).
- . Available from: https://www.thermofisher.com/order/catalog/product/A7816 (2023)
- Kreibig, U., Vollmer, M. Theoretical considerations. In: Optical Properties of Metal Clusters. 25, (1995).
- Mie, G. Beiträge zur Optik trüber Medien, speziell kolloidaler Metallösungen. Annalen der Physik. 330 (3), 377-445 (1908).
- Rueden, C. T., et al. ImageJ2: ImageJ for the next generation of scientific image data. BMC Bioinformatics. 18 (1), 529 (2017).
- Schindelin, J., et al. Fiji: An open-source platform for biological-image analysis. Nature Methods. 9, 676-682 (2012).
- Klenow, M. B., Heitmann, A. S. B., Nylandsted, J., Simonsen, A. C. Timescale of hole closure during plasma membrane repair estimated by calcium imaging and numerical modeling. Scientific Reports. 11, 4226 (2021).
- Li, T., Wu, X., Liu, F., Li, N. Analytical methods based on the light-scattering of plasmonic nanoparticles at the single particle level with dark-field microscopy imaging. Analyst. 142 (2), 248-256 (2017).
- Gibbs-Flournoy, E. A., Bromberg, P. A., Hofer, T. P. J., Samet, J. M., Zucker, R. M. Darkfield-Confocal Microscopy detection of nanoscale particle internalization by human lung cells. Particle and Fibre Toxicology. 8 (1), 2 (2011).
- Taylor, R. W., Sandoghdar, V. Interferometric scattering microscopy: Seeing single nanoparticles and molecules via Rayleigh scattering. Nano Letters. 19 (8), 4827-4835 (2019).
- Wu, Y., Ali, M. R. K., Chen, K., Fang, N., El-Sayed, M. A. Gold nanoparticles in biological optical imaging. Nano Today. 24, 120-140 (2019).
- Klingberg, H., Oddershede, L. B., Loeschner, K., Larsen, E. H., Loft, S., Møller, P. Uptake of gold nanoparticles in primary human endothelial cells. Toxicology Research. 4 (3), 566-666 (2015).
Reimpressões e Permissões
Solicitar permissão para reutilizar o texto ou figuras deste artigo JoVE
Solicitar PermissãoThis article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. Todos os direitos reservados