Method Article
Une approche thermoplasmonique pour étudier la réparation de la membrane plasmique dans les cellules vivantes et les membranes modèles
Dans cet article
Résumé
La méthode de ponction thermoplasmonique intègre la microscopie confocale, la pince optique et les nanoparticules d’or pour étudier les réponses des protéines lors de la réparation de la membrane plasmique dans les cellules et les vésicules unilamellaires géantes. Cette technique permet une ponction membranaire rapide et localisée, permettant l’identification de protéines clés et de leurs rôles fonctionnels dans la machinerie complexe de réparation de la membrane plasmique.
Résumé
La membrane cellulaire est cruciale pour la survie des cellules, et il est essentiel d’assurer son intégrité car la cellule subit des blessures tout au long de son cycle de vie. Pour éviter d’endommager la membrane, les cellules ont développé des mécanismes efficaces de réparation de la membrane plasmique. Ces mécanismes de réparation peuvent être étudiés en combinant la microscopie confocale et la thermoplasmonique à l’échelle nanométrique pour identifier et étudier le rôle de protéines clés, telles que les annexines, impliquées dans la réparation de surface dans les cellules vivantes et les systèmes modèles membranaires.
La méthode de ponction utilise un laser pour induire un échauffement très localisé lors de l’irradiation de nanoparticules. L’utilisation de la lumière proche infrarouge minimise la phototoxicité dans l’échantillon biologique, tandis que la majorité de l’absorption a lieu dans la nanoparticule plasmonique résonante proche infrarouge. Cette méthode thermoplasmonique a été exploitée pour des recherches photothermiques et biophysiques potentielles afin d’améliorer la compréhension des mécanismes intracellulaires et des réponses cellulaires grâce à des études de fusion de vésicules et de cellules. L’approche s’est avérée complémentaire aux méthodes existantes de rupture membranaire, telles que les lésions induites mécaniquement, chimiquement ou optiquement, et offre un haut niveau de contrôle en infligeant des blessures extrêmement localisées. L’étendue de la lésion est limitée au voisinage de la nanoparticule sphérique, et aucun dommage préjudiciable ne se produit le long du trajet du faisceau, contrairement aux lasers pulsés utilisant des longueurs d’onde différentes. Malgré certaines limitations, telles que la formation de nanobulles, la méthode thermoplasmonique offre un outil unique pour étudier les réponses cellulaires dans la réparation de la membrane plasmique dans un environnement presque natif sans compromettre la viabilité cellulaire.
Lorsqu’elle est intégrée à la microscopie confocale, la méthode de ponction peut fournir une compréhension mécaniste de la dynamique membranaire dans les systèmes membranaires modèles ainsi que des informations quantitatives sur les réponses des protéines aux dommages membranaires, y compris le recrutement des protéines et leur fonction biophysique. Dans l’ensemble, l’application de cette méthode à des systèmes modèles réduits peut améliorer notre compréhension de la machinerie complexe de réparation de la membrane plasmique dans les cellules vivantes.
Introduction
La membrane cellulaire, qui sert à la fois de barrière physique et de plate-forme de signalisation, est vitale pour la survie des cellules1. Tout au long de son cycle cellulaire, la membrane plasmique (PM) est soumise à des dommages, tels que des lésions mécaniques 2,3,4,5 et chimiques6 induites par le stress. Pour maintenir l’intégrité de la membrane et assurer la survie de la cellule, celle-ci a développé des mécanismes robustes de réparation de la membrane plasmique (PMR). Ces mécanismes dépendent de diverses stratégies, telles que la réorganisation du cytosquelette, la fusion membranaire et les stratégies de remplacement membranaire 7,8,9,10,11, qui reposent toutes sur le recrutement de protéines spécifiques. Notamment, les membres de la famille des protéines annexines ont été identifiés comme des protéines clés associées aux processus de PMR 1,9,12,13,14,15,16. À la suite d’une lésion des particules, la cellule subit un afflux d’ions calcium (Ca2+), ce qui constitue une menace immédiate pour la survie de la cellule17. En réponse à l’afflux de Ca2+, les protéines de l’annexine, qui sont principalement situées dans le cytosol, se lient au feuillet interne de la membrane plasmique endommagée dans le cadre des stratégies PMR18. L’annexine A2 (ANXA2) a été l’un des premiers membres de la famille de l’annexine à être associée à la RPM dans la dystrophie musculaire déficiente en dysferline et il a été suggéré qu’elle intermédie la réparation en fusionnant des vésicules intracellulaires à la MP près du site de la lésion 5,19,20,21. Par la suite, plusieurs fonctions ont été attribuées aux annexines22, et leur rôle dans la RPM a suscité une attention accrue au cours des 20 dernières années. Cependant, le rôle exact des annexines dans la RPM n’est pas encore entièrement compris 15,18,21,22.
Cet article propose une méthode pour étudier l’interaction protéine-membrane et la dynamique membranaire de manière contrôlée et hautement localisée, en utilisant une combinaison de microscopie confocale, de pinces optiques et de nanoparticules d’or (AuNPs). Cette méthode permet l’étude quantitative des interactions entre les protéines, les lipides et les petites molécules en réponse aux dommages membranaires et à l’afflux de Ca2+ . Malgré la complexité et la multiplicité des composants impliqués dans le processus de réparation membranaire, des systèmes membranaires simplifiés qui imitent la membrane plasmique ont été utilisés pour acquérir une compréhension mécanistique plus approfondie de la dynamique membranaire et de la réponse des protéines annexines à la perturbation membranaire16. Des vésicules lipidiques unilamellaires géantes (GUV) ont été choisies comme système membranaire modèle avec une composition lipidique spécifiée. Les GUV ont été générés à l’aide de la méthode d’hydratation assistée par gel, en particulier l’hydratation par gel d’alcool polyvinylique, telle que décrite par Weinberger et al.23, qui a permis une encapsulation efficace des annexines dans les GUV.
L’utilisation de l’irradiation laser dans le proche infrarouge (NIR) sur les nanoparticules métalliques (NP) induit un échauffement important du NP, ce qui en fait une méthode efficace pour établir une source de chaleur locale exploitée dans les applications biomédicales24. La méthode a d’abord été utilisée pour mesurer directement la température autour d’un seul AuNP dans des tests biomimétiques 2D et 3D. Dans ces essais25,26, les nanoparticules plasmoniques ont été irradiées sur une bicouche lipidique supportée ou piégées optiquement à proximité des GUV subissant une transition de phase thermique locale lors d’un chauffage local, ce qui a permis de quantifier et de contrôler le profil de température exact autour de la particule. Ce profil de température de référence a été utilisé lors de l’étude ou de la manipulation d’échantillons biologiques. D’autres progrès dans la méthode ont facilité l’induction de pores nanoscopiques dans les membranes27, permettant la fusion des vésicules et des cellules28,29. D’autres études ont étudié le comportement des protéines membranaires périphériques dans les GUVs29 et les protéines transmembranaires30 en créant de nouvelles vésicules hybrides, tandis que l’administration de médicaments spécifiques aux cellules a également été explorée pour contrôler et étudier les réponses cellulaires ou l’expression des gènes 28,29,31,32,33. Récemment, la méthode a été utilisée pour étudier les réponses des protéines aux lésions membranaires 32,34,35.
Il existe plusieurs méthodes pour perturber la membrane plasmique afin d’explorer les réponses cellulaires et la réparation des membranes. Il s’agit notamment des ponctions de micro-aiguilles, de l’agitation des microbilles et du grattage cellulaire, qui peuvent tous perturber mécaniquement la membrane cellulaire 14,36,37. Les dommages induits chimiquement peuvent être obtenus en ajoutant des détergents 5,38 ou des toxines bactériennes39,40 qui déstabilisent la bicouche lipidique et génèrent des pores membranaires à travers la membrane plasmique. De plus, des lésions induites optiquement par des lasers à ondes continues et pulsées ont été utilisées pour étudier des composants PMR, tels que les protéines annexine 5,14,21,41, en combinaison avec des nanoparticules plasmoniques 42,43,44,45. Malgré l’efficacité des lasers pulsés de haute puissance, ils peuvent causer des blessures importantes et des dommages à l’intérieur de la cellule le long du trajet du faisceau. De plus, les changements détaillés qui se produisent dans la matière biologique lors de l’irradiation laser pulsée et si elle crée un pore bien défini restent à étudier. Une méthode alternative est présentée dans cet article, utilisant la thermoplasmonique pour induire des trous nanoscopiques dans les particules de manière contrôlée34,35 sans causer de dommages significatifs aux structures internes. Ceci est accompli en exposant les NP plasmoniques à un laser NIR hautement focalisé, ce qui entraîne une augmentation de température extrêmement localisée qui peut facilement atteindre des températures supérieures à 200 °C, ce qui peut conduire à de petites explosions nanoscopiques 25,46,47. Ce processus peut être contrôlé en ajustant l’intensité du laser ainsi que la taille, la forme et la composition des NPs48. En utilisant cette technique, les chercheurs peuvent explorer le rôle des protéines dans la réparation des particules dans les cellules vivantes, ce qui pourrait aider à répondre à certaines des questions sans réponse concernant l’implication des protéines d’annexine dans la réparation membranaire sans compromettre la viabilité cellulaire.
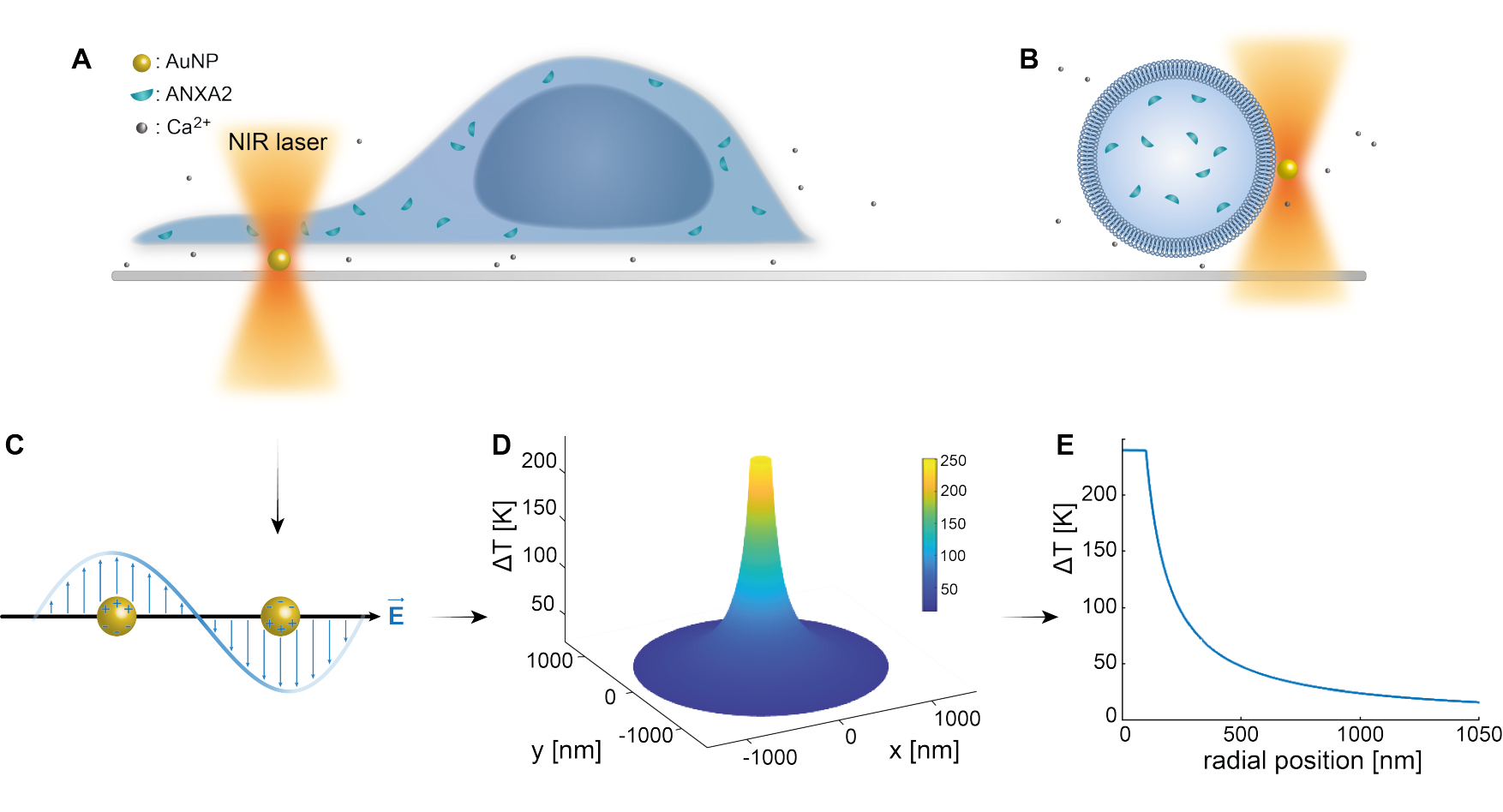
Le piégeage optique des nanoparticules plasmoniques a été bien établi par des études antérieures 25,49,50,51,52 ; cependant, des informations supplémentaires concernant les propriétés thermoplasmoniques des nanoparticules 53,54,55 peuvent être obtenues dans les matériaux supplémentaires (Fichier supplémentaire 1). La méthode thermoplasmonique peut être utilisée pour créer des trous nanoscopiques dans les particules dans le but d’étudier la réponse cellulaire et les mécanismes de réparation. Plus précisément, la ponction peut être réalisée par le chauffage optique des AuNPs à proximité immédiate de la membrane, comme le montrent les figures 1A et B. Cette ponction localisée permet un afflux de Ca2+, qui a été vérifié par un capteur de calcium, activant ainsi la machinerie PMR. Pour les expériences sur cellules vivantes, des AuNPs d’un diamètre de 200 nm ont été immobilisés à la surface sous la cellule pour surveiller le rôle d’ANXA2 dans la PMR par microscopie confocale. Le laser NIR (Figure 1A,B), d’une longueur d’onde de 1064 nm, irradie l’AuNP, exploitant ses propriétés plasmoniques (Figure 1C), ce qui entraîne un échauffement local important (Figure 1D) dans la fenêtre de transparence biologique49 tout en causant des dommages minimes à la cellule elle-même. La région à haute température entourant l’AuNP diminue rapidement de 30 à 40 % à une distance correspondant au rayon du NP, comme le montre la figure 1E, ce qui permet une lésion extrêmement confinée dans les trois dimensions.
Dossier supplémentaire 1. Veuillez cliquer ici pour télécharger ce fichier.

Figure 1 : Schéma de la méthode expérimentale. (A) les cellules transfectées ANXA sont situées au-dessus de nanoparticules d’or immobilisées (AuNPs) à la surface, ou (B) les vésicules monolamellaires géantes (GUVs) avec ANXA encapsulé sont en suspension dans un milieu contenant des AuNPs. (C) Un seul AuNP est irradié par le piège optique NIR, où l’interaction entre le champ électromagnétique entrant et les électrons de conduction conduit à l’oscillation collective des électrons dans le NP. (D) Ce processus entraîne une augmentation très confinée mais significative de la température. Pour estimer la température à la surface du NP, la théorie de Mie est utilisée, et un profil de température (E) est calculé pour un AuNP d’un diamètre de 200 nm et d’une intensité laser I = 6,36 x 108 W/cm2. Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}
Pour minimiser l’effet thermique sur la membrane cellulaire, les AuNPs ne sont irradiés que pendant ~1 seconde. Cela provoque une explosion transitoire et locale d’échauffement, ce qui réduit les dommages aux protéines qui nécessitent généralement plus de temps pour se déployer. Lors de la ponction membranaire, les protéines d’annexine sont recrutées en une fraction de seconde et, en quelques secondes, un échafaudage en forme d’anneau d’annexine se forme autour du site de la lésion (Figure 2). Cette approche a également été appliquée pour explorer l’implication d’ANXA5 à la fois dans les cellules vivantes et dans les membranes modèles16 dans le but de faire la lumière sur le schéma complet des processus de réparation. Bien que l’accent ait été mis sur le recrutement corrélé de diverses protéines annexines, les aspects biophysiques du mécanisme de réparation n’ont pas encore été élucidés.
Pour mettre pleinement en œuvre la méthode proposée, trois composants clés sont nécessaires : la microscopie confocale, la pince optique et les nanoparticules métalliques. Des pinces optiques sont utilisées pour piéger les AuNPs, et leur construction peut être réalisée en suivant la procédure décrite par Neuman et al.49. Cependant, si la construction d’une pince optique s’avère trop difficile, un laser NIR à focalisation serrée peut être utilisé pour irradier les AuNPs immobilisés sous les cellules. Bien que des AuNP sphériques aient été choisis pour ce protocole, une variété de particules plasmoniques avec des spectres d’absorption accordables pourrait également être utilisée pour obtenir un gradient de température hautement localisé dans la région NIR48.
L’imagerie par fluorescence est nécessaire pour observer le rôle des protéines marquées par fluorescence et, par conséquent, la microscopie à réflexion interne totale (TIRF)56 pourrait être considérée comme une alternative à l’imagerie confocale. Cependant, cette technique ne permet que l’imagerie de surface et ne serait pas compatible avec les expériences modèles de vésicules membranaires. Par conséquent, la pince optique et le microscope confocal sont essentiels pour la localisation précise de la nanoparticule et l’étude détaillée de la zone locale entourant la lésion cellulaire. Pour irradier efficacement la nanoparticule avec un foyer laser limité par diffraction, il est nécessaire de visualiser la nanoparticule. Cela peut être réalisé de manière optimale par microscopie à réflexion, qui est une caractéristique d’imagerie standard des microscopes confocaux Leica. Cependant, si l’imagerie par réflexion ou diffusion n’est pas disponible, d’autres méthodes, telles que le marquage fluorescent AuNP moins efficace, peuvent être envisagées.
En résumé, la méthode thermoplasmonique hautement contrôlable et localisée présentée dans cette étude a le potentiel de servir d’excellente plate-forme pour étudier les composants moléculaires impliqués dans les réponses cellulaires et les mécanismes de réparation des particules dans les cellules vivantes. En plus d’étudier la réponse protéique lors de dommages aux particules, cette approche peut également être utilisée pour perforer localement les vésicules, permettant ainsi d’étudier la réponse protéique dans la dynamique protéine-protéine et protéine-membrane. De plus, cette méthode permet une analyse quantitative des interactions entre les protéines, les lipides et les petites molécules lorsque les membranes sont perturbées. Collectivement, ces avancées ont le potentiel de faire la lumière sur certaines des questions non résolues concernant la machinerie complexe et complexe de réparation des membranes plasmiques.
Protocole
1. Préparation de la ponction de la membrane cellulaire
- Ensemencement cellulaire (Jour 1)
- Cultiver des cellules rénales embryonnaires humaines (HEK293T) dans un milieu de culture à 37 °C dans un incubateur humidificateur à 5 % de CO2 jusqu’à ce qu’elles atteignent 70 % de confluence.
- Détacher les cellules de la surface à l’aide de 500 μL de trypsine, compter 200 000 cellules HEK293T et les ensemencer dans une boîte de culture d’un volume total de 3 mL de milieu de culture. Incuber les cellules à 37 °C dans un humidificateur à 5 % de CO2 pendant 24 h.
REMARQUE : Pour éviter que les cellules ne se regroupent au centre de la boîte, répartissez les cellules uniformément et évitez de faire tourbillonner la boîte, car cela peut diminuer l’efficacité de la transfection.
- Transfection cellulaire (Jour 2)
REMARQUE : Les cellules transfectées peuvent être utilisées jusqu’à 48 h après la transfection.- Pipeter le plasmide d’intérêt et le réactif de transfection pendant 5 s avant utilisation.
- Dans un tube stérile de 2 mL, mélanger les éléments suivants dans l’ordre spécifique : 500 μL de milieu sérique réduit, 5 μL de réactif de transfection (4 fois plus que le plasmide) et 1,25 μL de plasmide (1 μg/μL).
REMARQUE : Pour étudier l’afflux de calcium lors de la rupture de la membrane, suivez la même procédure, mais utilisez la sonde de capteur de calcium liée à la membrane GCaMP6-CAAX (1 μg/μL). - Pipeter doucement mais soigneusement le mélange de transfection et l’incuber à température ambiante (RT) pendant 30 minutes avant de l’ajouter goutte à goutte aux cellules.
- Avant d’ajouter le mélange de transfection, retirez le milieu de la boîte de culture, lavez délicatement les cellules avec 2 mL de solution saline tamponnée au phosphate et ajoutez 2000 μL de milieu sérique réduit dans la boîte.
- Incuber les cellules avec le mélange de transfection à 37 °C dans un humidificateur à 5 % de CO2 pendant 2 h et 45 min avant de changer le milieu pour 3 mL de milieu de culture.
- Préparation d’une solution de nanoparticules d’or (AuNP) (Jour 2)
- Vortex la solution mère AuNP de 200 nm pendant 10 s au niveau 10 (voir le tableau des matériaux pour plus de détails sur l’appareil), soniser pendant 5 min (amplitude maximale) et vortex à nouveau pendant 10 s.
- Mélanger 150 μL d’AuNPs avec 850 μL d’eau distillée jusqu’à un volume total de 1 mL.
REMARQUE : La solution d’AuNP diluée peut être conservée au réfrigérateur et réutilisée jusqu’à 1 mois.
- Préparation du plat expérimental (Jour 2)
- Enduire la boîte de micropuits de 150 μL de solution de fixation cellulaire à 0,01 % à 0,1 % et incuber pendant 15 min à RT.
- Lavez deux fois la surface du verre avec 500 μL d’eau distillée et laissez-la sécher à l’air libre pendant ~10 min.
- Ajouter 80 μL de solution d’AuNP goutte à goutte sur la surface sèche.
REMARQUE : Vortex (10 s), sonicate (5 min) et vortex (10 s) la solution d’AuNP diluée avant de l’ajouter à la surface du verre enduit pour minimiser les agrégats d’AuNP. - Attendre ~10 min avant d’introduire 1,5 mL de milieu de culture. Laissez incuber le plat à 37 °C pendant la nuit.
- Préparation de la chambre expérimentale (Jour 3)
REMARQUE : La chambre expérimentale peut être préparée le jour 3 ou 4 ; Cependant, assurez-vous que les préparations suivantes sont effectuées le même jour que l’expérience.- Retirez le milieu des cellules dans la boîte de culture et lavez les cellules avec 2 mL de solution saline tamponnée au phosphate.
REMARQUE : Cette étape est essentielle pour éliminer tout milieu résiduel et débris qui pourraient interférer avec les étapes suivantes. - Ajouter 500 μL de solution de détachement cellulaire à base d’enzymes dans le puits et incuber pendant 1 à 3 minutes jusqu’à ce que les cellules se soient détachées de la boîte de culture.
- Ajouter 1,5 mL de milieu de culture frais et pipeter la solution cellulaire pour obtenir une solution cellulaire homogène afin de minimiser la probabilité d’amas cellulaires.
- Retirez délicatement le milieu des AuNPs dans le micropuits expérimental.
- Ajouter la solution cellulaire (2 mL) dans le micropuits et laisser incuber pendant au moins 5 h avant d’effectuer l’expérience.
REMARQUE : Pour des conditions expérimentales optimales, évitez de faire tourbillonner la chambre, car cela pourrait provoquer un regroupement de cellules au milieu de la chambre.
- Retirez le milieu des cellules dans la boîte de culture et lavez les cellules avec 2 mL de solution saline tamponnée au phosphate.
2. Expérience de ponction de membrane cellulaire
- Paramètres optiques de l’expérience
- Mener les expériences à l’aide d’un microscope confocal à balayage combiné à un laser de piégeage de 1064 nm57.
- Effectuez le piégeage optique au niveau du plan focal à l’aide d’un objectif à immersion dans l’eau 63x avec une ouverture numérique (NA) de 1,2.
- Supposons que la mise au point soit de la taille d’un disque d’Airy et que la largeur du faisceau focal du laser d’irradiation soit de ~540 nm de rayon.
- Convertissez la puissance laser (P) en intensité laser correspondante (I) en calculant la puissance laser par zone (W/cm2).
- Utilisez un séparateur de faisceau optique acoustique (AOBS) pour visualiser plusieurs signaux fluorescents détectés à l’aide de tubes photomultiplicateurs et la détection simultanée de NP métalliques via leur signal de diffusion.
REMARQUE : Tous les systèmes confocaux ne sont pas équipés d’AOBS, qui permet l’imagerie par réflexion des NP métalliques. Dans ce cas, d’autres formes de détection doivent être mises en œuvre, ou un balayage séquentiel du laser NIR dans la zone située sous la cellule peut être tenté. - Pendant les sessions expérimentales, montez une chambre ouverte à fond de verre contenant des cellules, des sondes fluorescentes moléculaires et des nanoparticules d’or sur le microscope.
- Déplacer le piège par rapport aux cellules en déplaçant l’échantillon monté sur une platine piézoélectrique, ce qui permet des mouvements latéraux de l’ordre du nanomètre16.
REMARQUE : Le piège optique est maintenu immobile.
- Paramètres expérimentaux pour la ponction membranaire cellulaire
- Placer HEK293T cellules, transfectées avec des plasmides d’annexine couplés à une protéine fluorescente, au-dessus des AuNPs isolés qui sont immobilisés sur la surface du verre (Figure 1A).
- Utilisez un laser Argon 488 nm pour visualiser le signal de fluorescence GFP et un laser HeNe 633 nm pour observer la réflexion de l’AuNP dans le microscope confocal à balayage.
- Utilisez la pince optique, qui fonctionne avec un laser NIR de 1064 nm, pour irradier un seul AuNP pendant ~1 s, induisant une augmentation locale significative de la température qui perturbe la membrane plasmique.
- Appliquer une irradiation comprise entre 200 et 295 mW à la particule focalisée, ce qui entraîne une augmentation substantielle de la température.
REMARQUE : Il y a une perte de puissance substantielle dans l’optique, la puissance laser au point focal atteignant environ 20% du milliwatt indiqué, selon la configuration spécifique. L’intensité spécifique (mesurée en W/cm2) dépend de l’alignement exact du système, en particulier de la taille focale. De plus, la conduction thermique du verre est supérieure à celle de la cellule et de l’eau et, par conséquent, la réduction de la quantité de chaleur libérée vers la membrane plasmique est légèrement réduite48,58. - Obtenir une intervention efficace et localisée en cas de lésions particulaires et une réponse de réparation des protéines en calibrant correctement la localisation du point de focalisation du laser NIR. Pour ce faire, il suffit de capturer un seul AuNP en suspension dans le même milieu d’imagerie et de s’assurer que l’AuNP sélectionné est mis au point avant l’irradiation.
NOTA : La nanoparticule est considérée comme nette lorsque son signal de diffusion apparaît le plus net, c’est-à-dire qu’elle présente des bords nets et l’absence de halo autour de la particule, lorsqu’on l’observe en microscopie confocale (Figure 2C (ii)).
- Conditions de densité de cellules et d’AuNP
- Choisissez des cellules individuelles plutôt que des amas de cellules pour éviter le chevauchement des membranes plasmiques.
NOTA : Les cellules doivent être collées à la surface, présentant une morphologie aplatie (figure supplémentaire 1), ce qui permet de perforer la périphérie de la cellule tout en évitant d’endommager la membrane nucléaire (figure supplémentaire 2). - Incuber les cellules selon le protocole ou jusqu’à ce qu’elles se soient stabilisées et aplaties pour empêcher l’absorption cellulaire de l’AuNP. Évitez un temps d’incubation excessif et réduisez la probabilité d’endocytose de l’AuNP à l’aide d’AuNPs PEGylés.
- S’assurer que les AuNPs immobilisés sont présents sous forme de particules individuelles, suffisamment espacées les unes des autres pour éviter les agrégats. Les agrégats peuvent entraîner une augmentation significative du gradient thermique, ce qui entraîne des températures élevées qui peuvent perturber une fraction substantielle de la cellule.
- Remplacez l’échantillon toutes les 1 à 2 heures pour maintenir la santé des cellules.
REMARQUE : Une exposition prolongée peut entraîner une détérioration de la santé des cellules, compromettant leur capacité à réagir avec précision aux blessures en termes de réparation de la membrane. Ce déclin de la santé cellulaire est généralement observé par un changement de forme cellulaire, car les cellules apparaissent plus sphériques et plus rigides, aboutissant finalement à la mort cellulaire.
- Choisissez des cellules individuelles plutôt que des amas de cellules pour éviter le chevauchement des membranes plasmiques.
Informations complémentaires. Veuillez cliquer ici pour télécharger ce fichier.
3. Préparation de la vésicule unilamellaire géante (GUV)
- Préparation du mélange lipidique
- Obtenir la composition lipidique GUV en combinant les lipides 1,2-dioleoyl-sn-glycéro-3-phosphocholine (DOPC) et 1,2-dioleoyl-sn-glycéro-3-phospho-L-sérine (DOPS) dans un rapport molaire de 4 :1 (voir tableau 1). Aliquoter le bouillon lipidique dans des flacons en verre de 1,5 mL selon les besoins expérimentaux et les conserver à -20 °C.
REMARQUE : Pour une conservation prolongée des lipides et pour éviter l’oxydation des lipides insaturés, remplacez l’air par de l’argon dans les flacons alicités. - Avant de mélanger les lipides, nettoyez soigneusement une seringue en verre ou en métal de 50 μL et de 500 μL avec du chloroforme cinq fois pour vous assurer qu’elles sont exemptes de contaminants pour les lipides dissous par le chloroforme.
ATTENTION : Manipuler le chloroforme dans une hotte en raison de sa toxicité.- Transférez le volume calculé de chloroforme dans un flacon en verre ambré propre, suivi de la quantité spécifiée de chaque lipide (voir le tableau 1).
REMARQUE : Pour éviter la contamination croisée entre les stocks de lipides, assurez-vous que les seringues sont nettoyées avec du chloroforme. - Enfin, ajoutez le colorant membranaire et mélangez bien les lipides par pipetage. Conservez le mélange lipidique préparé à -20 °C pour une utilisation ultérieure ; Le mélange reste viable pendant 2 à 3 semaines sans dommages lipidiques considérables.
REMARQUE : Le mélange doit être fait avec une seringue métallique ou en verre. Conservez toujours les lipides sur de la glace lorsqu’ils sortent du congélateur.
- Transférez le volume calculé de chloroforme dans un flacon en verre ambré propre, suivi de la quantité spécifiée de chaque lipide (voir le tableau 1).
- Obtenir la composition lipidique GUV en combinant les lipides 1,2-dioleoyl-sn-glycéro-3-phosphocholine (DOPC) et 1,2-dioleoyl-sn-glycéro-3-phospho-L-sérine (DOPS) dans un rapport molaire de 4 :1 (voir tableau 1). Aliquoter le bouillon lipidique dans des flacons en verre de 1,5 mL selon les besoins expérimentaux et les conserver à -20 °C.
- Préparation d’un gel d’alcool polyvinylique (PVA)
- Préparez les GUV en utilisant la méthode d’hydratation assistée par gel décrite par Weinberger et al.23avec des modifications mineures.
- Préparez le gel PVA en dissolvant 5 g de PVA dans 100 mL d’un tampon contenant 50 mM de saccharose, 25 mM de NaCl et 25 mM de Tris (pH 7,4).
- Chauffez la solution PVA à 85 °C et remuez jusqu’à ce qu’elle devienne transparente. Laissez-le refroidir jusqu’à ce qu’il soit RT et conservez-le au réfrigérateur pour une utilisation ultérieure.
REMARQUE : Le PVA n’est pas correctement dissous dans le tampon et il est donc nécessaire de le chauffer jusqu’à 85 °C.
- Préparez les GUV en utilisant la méthode d’hydratation assistée par gel décrite par Weinberger et al.23avec des modifications mineures.
- Préparation des lames de verre
- Nettoyez les lames de verre avec de l’éthanol et laissez-les sécher à l’air libre. Ensuite, traitez les lames avec un nettoyeur plasma d’air pour éliminer toute contamination résiduelle de la surface du verre.
- Préparation de lames de verre revêtues de PVA
- Réchauffez le gel PVA (5%) jusqu’à 60 °C pendant 30 min pour augmenter sa fluidité. Appliquez 90 μL de PVA chaud sur la lame de verre, étalez-la uniformément et laissez-la sécher dans une armoire chauffante à 50 °C pendant 50 min.
- Une fois que la lame de verre PVA est prête, ajoutez 30 μL du mélange lipidique préparé à l’aide d’une seringue en verre ou en métal et étalez-la en un film mince à l’aide du bord de l’aiguille.
- Séchez le mélange lipidique en évaporant sa teneur en chloroforme sous une légère pression d’azote. De plus, séchez les lames de verre sous vide pendant 1,5 à 2 h.
- Cultiver des GUV dans une chambre
- Assemblez la chambre interne, dont la conception est similaire à celle d’un rapport59 publié précédemment, à l’aide de la lame de verre préparée.
- Dans un tube séparé de 2 mL, diluer la protéine recombinante d’intérêt (dans ce cas, ANXA5 ou ANXA4) à une concentration finale de 500 nM avec un tampon de croissance (GB) composé de 80 mM de saccharose, 70 mM de NaCl et 25 mM de Tris-HCl (pH 7,4).
- Ajouter 400 μL de la solution de protéines recombinantes diluée dans la chambre. Incuber la chambre à RT pendant 1 h pour permettre aux GUV de se développer à partir de la couche lipidique déposée. Utilisez le même tampon, à l’exclusion de la protéine, comme témoin négatif.
REMARQUE : Enveloppez la chambre dans un film de polyoléfine pour éviter l’évaporation du tampon. - Recueillir les GUV en transférant 400 μL du contenu de la chambre dans un tube de 2 mL.
- Éliminer les protéines non encapsulées à l’extérieur des GUV en ajoutant 1 mL de tampon d’observation (OB) contenant 55 mM de glucose, 70 mM de NaCl et 50 mM de Tris-HCl (pH 7,4) à la solution recueillie. Ensuite, centrifugez la solution à 600 x g pendant 10 min à 13 °C.
- Après centrifugation, remplacer 1 mL de surnageant par un volume égal de tampon d’observation. Dispersez les GUV par pipetage doux et conservez-les au réfrigérateur jusqu’à ce qu’ils soient utilisés dans des expériences GUV.
4. Expérience de ponction GUV
- Préparation de la chambre
- Utilisez un plat à fond en verre de 35 mm pour l’expérience. Pour éviter que les GUV ne collent à la surface et n’éclatent, enduisez la surface de β-caséine (5 mg/mL) pendant 15 à 30 minutes.
- Pour la solution de β-caséine, dissoudre 0,1 g de la protéine dans un tampon de 20 mL de 20 mM de Tris (PH 7,5) et de 100 mM de NaCl. Filtrez la solution protéique, aliquotez-la dans de petits flacons et congelez-la pour une utilisation ultérieure.
- Lavez la chambre deux fois avec un tampon d’observation pour éliminer tout excès de β-caséines libres de la surface et laissez sécher à RT.
- Dans un tube séparé de 2 mL, mélanger les GUV collectés avec l’OB. Ajouter le CaCl2 au mélange pour obtenir la concentration finale souhaitée (dans ce cas, 200 μM).
- Introduire des nanocoquilles d’or (AuNS) de 150 nm dans le mélange dans un rapport de 1 :100. Le mélange final comprend 250 μL de GUV, 225 μL d’OB, 20 μL de CaCl2 (5 mM) et 5 μL d’AuNS spécifiés.
- Utilisez un plat à fond en verre de 35 mm pour l’expérience. Pour éviter que les GUV ne collent à la surface et n’éclatent, enduisez la surface de β-caséine (5 mg/mL) pendant 15 à 30 minutes.
- Dispositif expérimental
- Transférez le mélange dans la chambre et montez-le sur la platine du microscope. Selon la taille de la chambre, ajoutez soit le mélange entier, soit une partie du mélange.
REMARQUE : Le moment est critique car les ions calcium peuvent traverser la membrane et médier la liaison des annexines au feuillet interne de la membrane. - Utiliser la même configuration optique que celle utilisée pour les expériences de ponction cellulaire.
- Utilisez la pince optique pour piéger un AuNS individuel à proximité ou sur la surface d’un GUV en appliquant une puissance laser de ~ 125 mW.
- Par la suite, augmentez la puissance du laser à ~ 300 mW. Cela produit une augmentation de température très localisée, qui perturbe et perce la membrane au site cible.
REMARQUE : Les AuNS sont préférés pour les expériences GUV en raison de leur capacité à générer une augmentation transitoire de température plus élevée tout en maintenant une taille plus petite par rapport aux AuNP solides.
- Transférez le mélange dans la chambre et montez-le sur la platine du microscope. Selon la taille de la chambre, ajoutez soit le mélange entier, soit une partie du mélange.

Tableau 1 : Tableau de détermination de la composition de la GUV. Veuillez cliquer ici pour télécharger ce tableau.
5. Mesures et analyse des données de la réponse ANXA dans les expériences de ponction cellulaire
- Utilisez MATLAB pour analyser les images et pour calculer et tracer le profil de température AuNP (Figure 1D, E) basé sur les équations de Mie60,61.
- De plus, traitez toutes les images confocaux à l’aide de la distribution FIJI ImageJ62,63.
- Calcul du rayon de l’anneau ANXA
- Recadrez la région d’intérêt contenant la zone perforée à partir des données brutes avant l’analyse des lésions membranaires.
- Utilisez le workflow interne de MATLAB pour traiter chaque image en marquant manuellement le centre de l’anneau ANXA et son périmètre extérieur.
- Utilisez ces marquages pour définir les limites de traitement pour la région d’intérêt.
REMARQUE : La zone est ensuite traitée radialement dans les limites définies, et l’intensité de fluorescence à une distance spécifique du centre est la moyenne sur le cercle complet avec la même distance centrale. Ce nombre est stocké pour chaque pas de rayon. - Utilisez le flux de travail pour ajuster les intensités moyennes sur toute la plage de balayage à une courbe gaussienne unidimensionnelle afin d’identifier les maxima (Rp) et la largeur totale à la moitié du maximum (FWHM) correspondant respectivement à Rext etR int, comme illustré à la figure 3A.
- Utilisez ces marquages pour définir les limites de traitement pour la région d’intérêt.
- Enfin, déterminez le rayon de l’anneau ANXA en fonction de la distance entre le centre de l’anneau etl’extrémité R donnée par le tracé généré par le workflow MATLAB.
- Calcul tridimensionnel du rayon de l’anneau ANXA
- Suivez l’évolution du rayon de l’anneau ANXA dans la direction z à l’aide du même workflow d’analyse MATLAB qu’à l’étape 5.1. Appliquez le flux de travail à plusieurs sections confocaux en Z à travers la plaie de la membrane, comme illustré à la figure 3C, E.
- Calculez la pente de chaque plaie en fonction de la variation des rayons sur les sections z, comme illustré à la figure 3F.
- Évolution temporelle du rayon de l’anneau ANXA
- De même, suivez l’évolution de l’anneau ANXA de l’anneau ANXA après la ponction de la membrane thermoplasmonique à l’aide du workflow d’analyse MATLAB de l’étape 5.1. Analysez des points temporels consécutifs pour surveiller les changements au fil du temps, comme illustré dans les figures 3D, G, H.
Résultats
Dans la présente étude, la méthode thermoplasmonique est utilisée pour étudier la réponse de la protéine de l’annexine à la perturbation de la membrane plasmique ; Cependant, toute protéine susceptible d’être recrutée lors d’une lésion membranaire peut être étudiée à l’aide de ce test, étant donné que la protéine concernée est marquée par fluorescence. Le recrutement et la fonction des protéines sont surveillés par imagerie confocale dans les cellules rénales embryonnaires humaines (HEK293T) et les vésicules unilamellaires géantes (GUV). Pour élaborer, la figure 2 illustre les conditions expérimentales dans lesquelles un faisceau laser NIR focalisé à 1064 nm est utilisé pour irradier un seul AuNP (Figure 2A), ce qui entraîne une lésion de la membrane et un afflux de Ca2+ dans la cellule, activant la machinerie PMR. Par la suite, les annexines sont rapidement recrutées sur le site de la lésion, où elles se lient aux phospholipides chargés négativement sur le périmètre de la plaie, formant une structure en forme d’anneau en quelques secondes (Figure 2B, i-iv). Les expériences sur les membranes modèles, à l’aide de GUV, ont démontré que les perforations membranaires étaient rapidement refermées en l’absence d’ANXA, comme le montre la figure 2C. Cependant, en présence d’ANXA, une accumulation rapide d’ANXA a été observée au site de la lésion après la ponction de la membrane (Figure 2D). Notamment, ANXA a continué à rouler les bords exposés, ce qui a finalement conduit à l’éclatement du GUV. On pense que ce mécanisme de roulement provient de la capacité d’ANXA à induire une courbure et à plier les membranes lipidiques20.

Figure 2 : Réponse des annexines (ANXA) à la perturbation membranaire induite par les thermoplasmes. Initialement, (A) la membrane plasmique agit comme une barrière entre l’environnement extracellulaire contenant des niveaux élevés d’ions Ca2+ et l’environnement intracellulaire avec des ANXA encapsulés. Lors de l’irradiation avec un laser proche infrarouge (NIR), l’AuNP génère une chaleur substantielle, provoquant la rupture de la membrane et entraînant un afflux d’ions Ca2+. Par conséquent, la machinerie de réparation de la membrane plasmique (PMR) est activée, ce qui implique le recrutement d’ANXA sur le site de la lésion, où ils se lient aux phospholipides chargés négativement. (B-D) Les images de microscopie confocale d’une cellule contenant ANXA2 et d’un GUV contenant ANXA4 démontrent ce processus. (i) Avant l’irradiation, les images montrent une cellule intacte, ou GUV, avec les sites d’irradiation indiqués par les flèches blanches. (ii) Lors de l’irradiation par nanoparticules, (B) les ANXA sont rapidement recrutés sur le site de la lésion, formant une structure en forme d’anneau autour de la plaie membranaire (B [ii-iv]). Le panneau (C) montre un GUV coloré par la membrane sans ANXA, qui, en cas de perforation, se referme rapidement sans remodelage de la membrane observable. D’autre part, le panneau (D) montre un GUV contenant de l’ANXA4 recombinant, qui est déjà lié à la membrane avant (i) l’irradiation due à une fuite de Ca2+ à travers la membrane. (ii) Lors de la perforation, ANXA4 se lie aux bords libres, provoquant l’effondrement du GUV lorsque la membrane s’éloigne du bord. Les barres d’échelle mesurent 10 μm pour la figure (B), 2 μm pour B (i), 10 μm pour (C) et 15 μm pour (D). Cette figure est reproduite d’après Moreno-Pescador et. al16 avec l’autorisation de la Royal Society of Chemistry. Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}
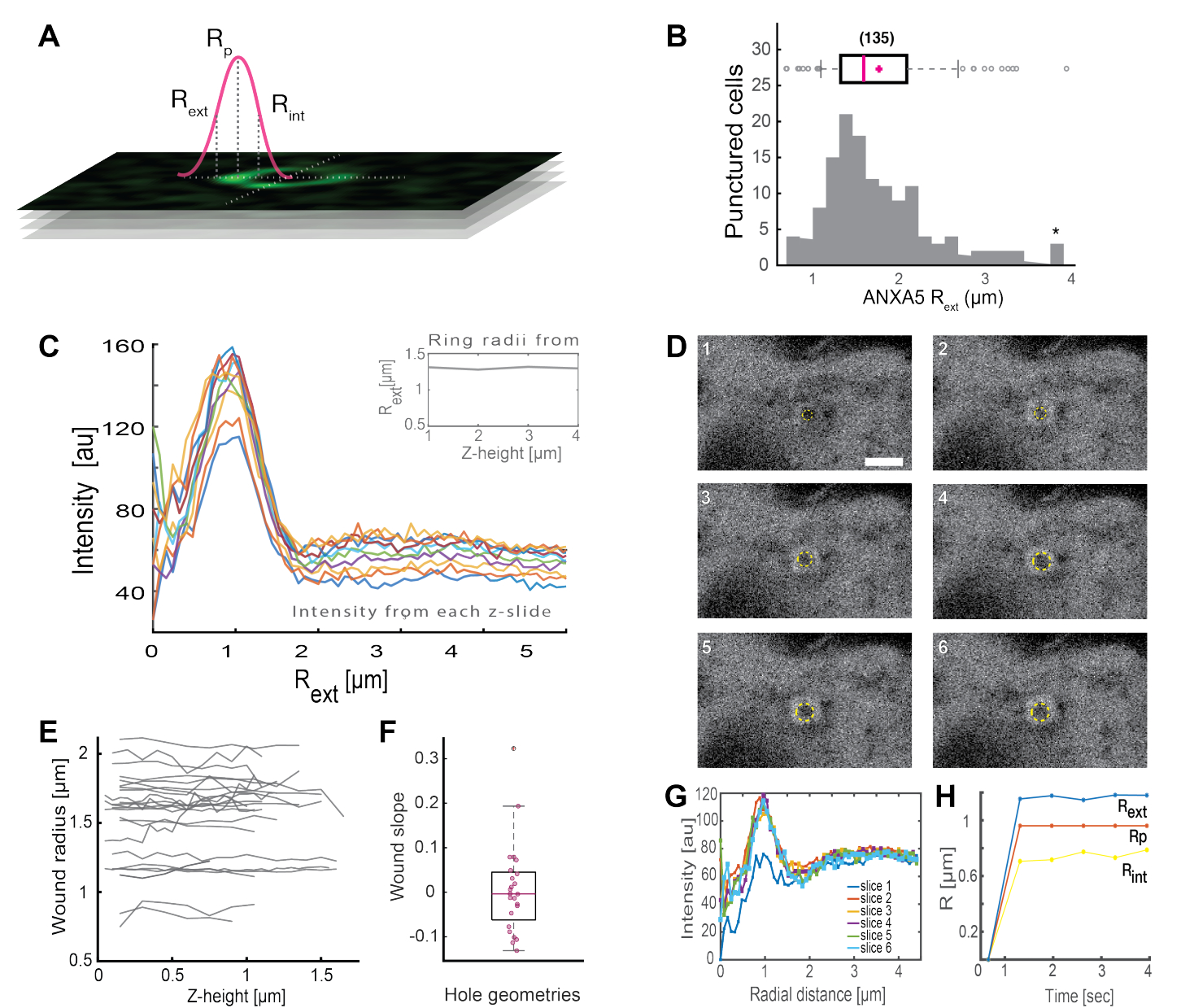
L’analyse des structures complètes en forme d’anneau ANXA (Figure 2B et Figure supplémentaire 1) fournit des informations utiles sur la taille et la morphologie de la plaie. Le rayon de la structure de l’anneau ANXA peut être déterminé dans le temps et dans l’espace, comme décrit dans la section 5. Moreno et al.16 ont analysé plus de 135 lésions de la membrane plasmique dans des cellules vivantes, où l’analyse des structures de l’anneau ANXA5 est représentée à la figure 3. Le rayon a été déterminé en mesurant la distance entre le centre de l’anneau et le rayon externe de la courbe en fonction de la demi-largeur maximale du profil d’intensité replié (figure 3A). Les résultats ont illustré une distribution hétérogène des tailles d’anneaux ANXA5 (Figure 3B), qui sont restées constantes dans le temps (Figure 3D, G, H) et dans l’espace (Figure 3C, E, F). Ces résultats suggèrent une accumulation d’ANXA5 autour des sites de lésion, ce qui indique une stratégie PMR alternative médiée par ANXA5 dans les cellules vivantes à l’hypothétique bourgeonnement vers l’intérieur de la membrane endommagée en forme d’entonnoir5.

Figure 3 : Analyse des structures annulaires d’ANXA5 entourant un site de lésion dans des cellules vivantes. (A) La représentation schématique illustre l’approche analytique, qui est basée sur la largeur maximale du profil de la raie d’intensité ANXA. (B) L’histogramme illustre 135 rayons d’anneaux ANXA5 mesurés. (C) Profils de raies d’intensité de fluorescence à travers un anneau ANXA5-GFP pour chaque section z de la pile z. (D) Les images confocales illustrent l’évolution temporelle d’une plaie, où ANXA5-GFP s’accumule au périmètre de la plaie immédiatement après la blessure, suivie de la stabilisation de la plaie. Les intervalles de temps étiquetés de 1 à 6 ont été capturés à des intervalles de 0,66 s par image. La barre d’échelle est de 2 μm. (E) Les rayons des plaies ont été déterminés en fonction de la profondeur de la plaie sur la base de la méthode présentée dans le panneau A. (F) La pente de la plaie a été analysée en tant que rayons de l’anneau ANXA5 en fonction de la hauteur z sur la base des données extraites du panneau E. (G) Les profils de raies d’intensité de fluorescence ont été mesurés à travers l’anneau ANXA5-GFP du panneau D, où chaque intervalle de temps 1 à 6 est noté tranche 1 à 6. Enfin, (H) l’évolution temporelle des rayons de l’anneau ANXA5-GFP est présentée en fonction du temps calculé à partir des données du panneau G. Cette figure est reproduite d’après Moreno-Pescador et. al16 avec l’autorisation de la Royal Society of Chemistry. Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}
Par rapport aux structures en forme d’anneau formées par la perturbation des particules seules (figure supplémentaire 1), les lésions à proximité immédiate du noyau (figure supplémentaire 2A) peuvent influencer l’évolution et la géométrie de la plaie. Parfois, seule une fraction des anneaux ANXA était évidente (Figure supplémentaire 2B,C), qui peut également être analysée à l’aide du workflow d’analyse interne de MATLAB (voir Section 5), bien que des données supplémentaires puissent être perdues. En règle générale, les formations d’anneaux ANXA observées dans les cellules non adhérentes (figure supplémentaire 2C) sont situées à proximité du noyau et de la périphérie de la cellule. En conséquence, une structure annulaire plus allongée peut être observée, ce qui n’est pas optimal pour l’analyse des données présentées. De plus, les cellules non adhérentes semblaient être plus sensibles à la mort cellulaire après une lésion des particules. De plus, lorsque l’on considère les blessures résultant de l’irradiation des agrégats d’AuNP, il est important de noter que ces blessures peuvent être plus graves et moins contrôlables. Cela est dû à l’augmentation significative de l’échauffement plasmonique, qui peut endommager une grande partie de la cellule. Par conséquent, ces blessures n’ont pas été intégrées dans l’analyse de l’anneau ANXA5.
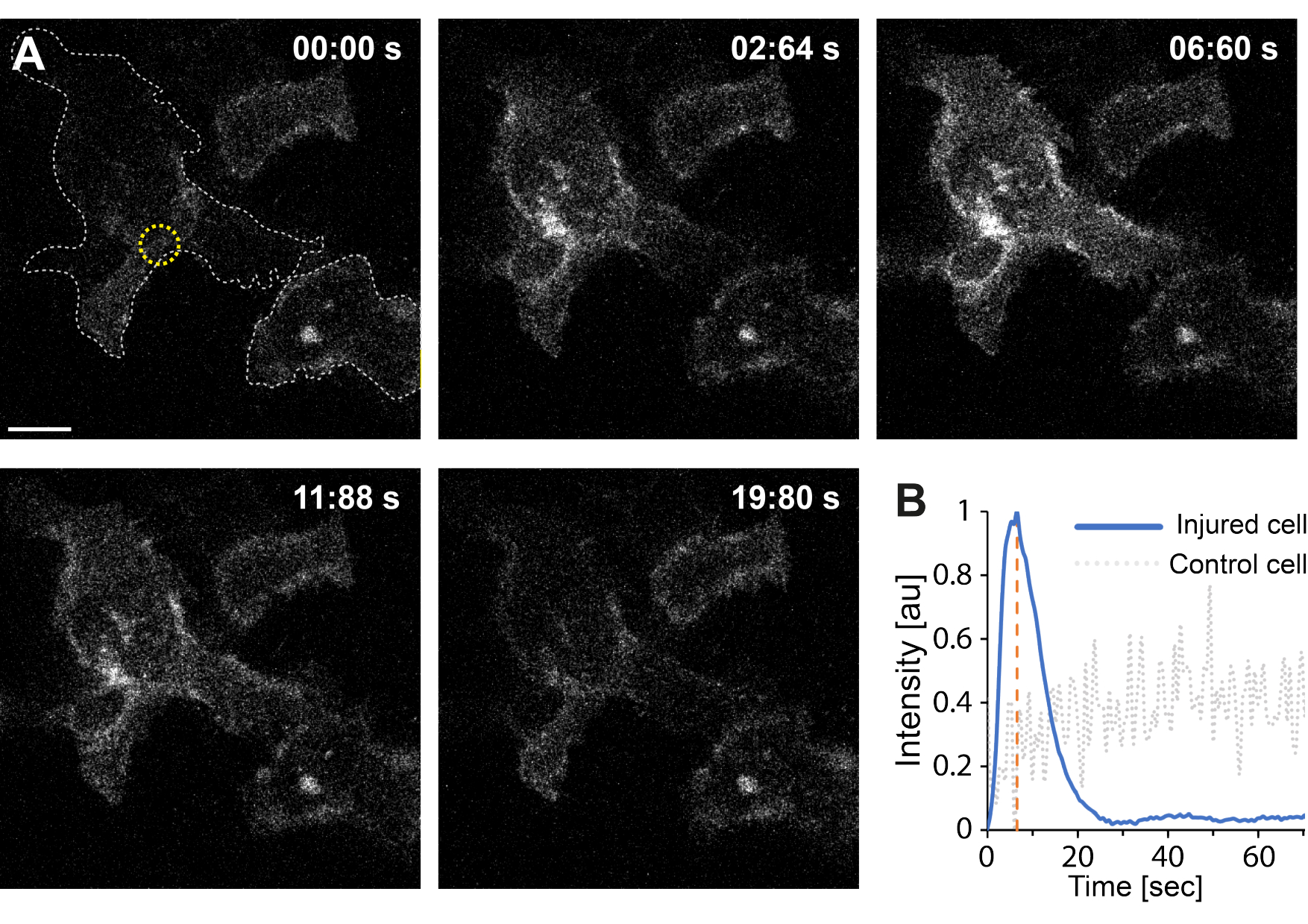
De plus, les résultats préliminaires indiquent que la perturbation de la membrane plasmique à l’aide de la thermoplasmonique entraîne des niveaux intracellulaires élevés de Ca2+ . Cela a été observé même à une irradiation de faible intensité d’AuNPs uniques, suggérant une perméabilisation des particules35, comme le montre la figure 4. L’afflux de Ca2+ a été observé dans des cellules exprimant une sonde de calcium liée à la membrane, GCaMP6s-CAAX, qui subit un changement conformationnel lors de l’afflux de Ca2+ , et donc, une augmentation de son intensité peut être observée64. L’intensité calcique a été quantifiée pour l’ensemble de l’empreinte de la cellule au fil du temps. Pour éliminer le bruit de fond, le niveau de fond de Ca2+ a été soustrait avant la perturbation de la membrane et après la réparation de la membrane. L’intensité maximale a été déterminée en normalisant l’intensité moyenne du Ca2+ à l’intérieur de la cellule, ce qui a donné lieu à une courbe d’intensité montrant une augmentation initiale rapide de l’intensité du Ca2+ suivie d’une diminution plus lente, comme le montre la figure 4B.
La cellule a atteint une intensité calcique maximale à ~ 6,6 s, ce qui est cohérent avec les résultats de Klenow et al.64, qui ont suggéré que le temps du pic d’intensité calcique (t = tc) correspond au temps nécessaire à la fermeture de la plaie. Néanmoins, bien que d’autres investigations soient nécessaires pour établir le mécanisme sous-jacent de la réparation membranaire et de la cicatrisation des plaies, les résultats préliminaires ont montré que ce processus de Ca2+ a été observé exclusivement dans la cellule lésée et non dans la cellule non blessée utilisée comme témoin positif. Cela confirme que la cellule a subi un afflux de Ca2+ lors d’une perturbation de la membrane thermoplasmonique, où l’excès de calcium intracellulaire est activement pompé après une PMR réussie, car le niveau de calcium intracellulaire n’est plus en compétition avec l’afflux de Ca2+ , atteignant finalement l’homéostase cellulaire64.

Figure 4 : L’afflux de calcium se produit lorsque la membrane plasmique d’une cellule HEK293T est rompue par la thermoplasmonique. Une série d’images confocales montre deux cellules (la cellule d’intérêt et une cellule non blessée utilisée comme témoin positif) exprimant la sonde calcique membranaire, GCaMP6s-CAAX. La barre d’échelle mesure 10 μm. (A) Avant l’irradiation, à 00 :00 s, l’empreinte des deux cellules est visualisée par la ligne pointillée grise, et le site d’irradiation est indiqué par le cercle jaune. Lors de l’irradiation laser, un afflux rapide de Ca2+ est observé, atteignant une intensité maximale à ~ 6,6 s, indiquée par la ligne pointillée orange, un point de temps qui est présumé correspondre au moment de la fermeture de la plaie64. (B) Le profil d’intensité calcique obtenu à partir de la sonde GCaMP6s-CAAX dans la cellule lésée (ligne bleue) a été comparé à l’intensité de Ca2+ dans une cellule voisine non blessée (ligne pointillée grise), montrant un afflux clair de Ca2+ exclusivement lors de la perturbation des particules. Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}
Discussion
L’étude met en évidence l’approche thermoplasmonique comme une technique prometteuse pour explorer les réponses protéiques dans les cellules vivantes et les membranes modèles après une rupture membranaire. Cette méthode fournit non seulement des informations détaillées sur le recrutement des protéines, mais aussi sur la fonction biophysique des protéines impliquées dans la dynamique protéine-membrane. Par conséquent, il facilite l’identification des composants moléculaires responsables de la réparation de surface et fait progresser la compréhension de la machinerie complexe mais vitale de la réparation des membranes plasmiques. Bien qu’il existe diverses méthodes pour induire une perturbation membranaire, telles que les techniques mécaniques, chimiques et optiques, ces méthodes souffrent de limitations, telles que le fait d’être non spécifiques aux cellules, de générer de multiples lésions de la membrane cellulaire ou de causer des dommages importants à la membrane et d’abler le matériel cellulaire interne le long du trajet du laser lors de l’utilisation de lasers pulsés de haute puissance. Bien que l’intégration de la microscopie confocale et de la pince optique offre les informations les plus complètes, d’autres modalités d’imagerie pourraient également être utilisées. Par exemple, comme l’imagerie de la nanoparticule plasmonique est réalisée à l’aide de la microscopie par réflexion, un mode d’imagerie intégré dans les microscopes confocaux Leica, des techniques d’imagerie supplémentaires, telles que la microscopie à fond noir 65,66, d’autres méthodes de diffusion comme iSCAT67,68 ou le marquage fluorescent de la nanoparticule, pourraient être utilisées pour la visualisation de l’AuNP, bien que cela puisse limiter l’applicabilité de la méthode.
La méthode présentée est en outre capable d’induire des trous nanoscopiques dans des membranes modèles, ce qui permet d’étudier les effets de synergie entre différentes annexines. Ceci est réalisé en encapsulant des annexines recombinantes marquées différemment, par exemple, RFP et GFP, respectivement, suivies d’une ponction thermoplasmonique. Ce système modèle donne un aperçu de la façon dont les annexines interagissent avec les membranes au voisinage des bords libres, comme le montre la figure 2D. Cependant, contrairement aux cellules, les trous infligés aux GUV continuent de s’étendre, suivis d’une déstabilisation de la vésicule. L’imagerie de l’évolution du trou à l’aide de la microscopie confocale peut être difficile en raison de l’expansion rapide du diamètre du trou, mais peut être accomplie en capturant plusieurs piles z au fil du temps. Une autre méthode consisterait à utiliser un disque confocal rotatif pour une imagerie plus rapide. De plus, l’approche thermoplasmonique donne généralement un nombre limité de résultats optimaux par heure lorsqu’elle est appliquée à des cellules uniques ou à des expériences GUV, généralement deux à trois, à des températures d’échantillon comprises entre 20 °C et 30 °C. Pour obtenir l’observation la plus précise possible de la dynamique protéine-membrane, il est recommandé de conserver les cellules dans un tampon contenant HEPES et de remplacer l’échantillon toutes les heures. Alternativement, la fenêtre expérimentale pourrait être prolongée en effectuant les expériences dans une chambre d’incubation cellulaire, c’est-à-dire à une température constante de 37 °C avec 5% de CO2. De plus, la combinaison de cette approche avec d’autres techniques d’imagerie, telles que la microscopie de reconstruction optique stochastique (STORM), pourrait permettre de mieux comprendre la fonction biophysique et l’interaction des protéines clés impliquées dans la réparation membranaire au niveau d’une seule molécule. Cela pourrait fournir des informations détaillées sur le site de la blessure, y compris la géométrie de la plaie et l’emplacement des protéines d’annexine, ainsi que l’identification d’autres acteurs clés impliqués dans la réparation de la surface de la membrane.
Afin d’obtenir une efficacité et une précision maximales dans l’induction d’une lésion membranaire, il est impératif de vérifier l’emplacement de la mise au point laser avant chaque expérience et de s’assurer que la position axiale de la mise au point laser coïncide avec la mise au point confocale. Cet alignement optimise l’intensité pendant l’imagerie AuNP, ce qui conduit à une augmentation locale maximale de la température et à une lésion de la membrane conséquente à une puissance laser plus faible. Ce processus est exécuté manuellement et est donc susceptible de varier l’efficacité de la rupture de la membrane, car la mise au point est traduite manuellement vers une position qui coïncide avec l’emplacement de la particule. Dans les microscopes qui n’ont pas de mode de réflexion, comme dans certains systèmes commerciaux, la colocalisation du foyer laser et de la particule peut être difficile. Dans de tels cas, d’autres modes d’imagerie (par exemple, fond clair) peuvent être utilisés, et un balayage raster lent peut être effectué autour de la position attendue de la particule. Il convient de noter qu’une faible puissance laser est susceptible d’induire uniquement une perméabilisation de la membrane, tandis qu’une puissance laser élevée peut générer des températures autour du NP qui dépassent le point d’ébullition de l’eau, même si la surface du verre a un effet de refroidissement. On estime que la formation de nanobulles entourant les NP se produit entre 200 °C et 300 °C25,48, où la chaleur explosive peut entraîner soit un déplacement des particules par rapport au foyer laser, soit une fragmentation des particules. De plus, la formation de nano ou de microbulles pendant le chauffage pose un défi à cette méthode. Comme l’air s’interface avec les membranes et peut provoquer une déstabilisation des protéines, ce qui n’est pas souhaitable, il est impératif de limiter l’échauffement lors de l’étude de la réparation des membranes. Notamment, les nanocoquilles d’or ne tolèrent pas les températures élevées et se dégradent dans ces conditions, comme l’a démontré la microscopie à haute résolution58.
Cet article fournit un protocole détaillé pour l’utilisation de la thermoplasmonique pour effectuer des ponctions hautement localisées dans les membranes, qui est applicable à la fois aux cellules et aux membranes modèles. Pour réduire davantage l’étendue de l’échauffement, des nanoparticules plus petites résonnant avec la lumière NIR peuvent être utilisées, ce qui permet des ponctions intracellulaires dans les endosomes, le réticulum endoplasmique et l’enveloppe nucléaire. De telles nanoparticules, y compris les bâtonnets et les nanomatriochkas48, peuvent être utilisées pour étudier la réparation de l’enveloppe nucléaire en ciblant les nanoparticules d’or endocytosées qui sont facilement absorbées à la surface de la cellule et trafiquées vers le noyau69. Dans l’ensemble, cette technique permet d’identifier et d’examiner les composants moléculaires clés impliqués dans la PMR, d’élucider leur fonction et leur rôle biophysiques tout en préservant la viabilité des cellules.
Déclarations de divulgation
Les auteurs n’ont rien à divulguer.
Remerciements
Nous tenons à remercier Jesper Nylandsted de nous avoir fourni des protéines d’annexine recombinantes et des plasmides codant pour les annexines. Ce travail a été soutenu financièrement par le Conseil danois pour la recherche indépendante en sciences naturelles (DFF-4181-00196), par le programme de synergie interdisciplinaire 2018 de la Fondation Novo Nordisk (NNF18OC0034936), le comité scientifique de la Société danoise du cancer (R90-A5847-14-S2), la Fondation Lundbeck (R218-2016-534) et par le Centre d’excellence de la Fondation Lundbeck (biomembranes en nanomédecine).
matériels
| Name | Company | Catalog Number | Comments |
| 1064 nm trapping laser | Spectra Physics | N/A | Spectra Physics J201-BL-106C, Nd: YVO4 NIR laser |
| 160 nm Gold Nanoshells | NanoComposix | NCXGSIR150 | |
| 200 nm Gold Nanoparticles | BBI Solutions | EM.GC200/7 | |
| 35 mm glass surface MatTex microwell | MATTEK | P35G-1.5-14-C | |
| Amber-glass vials | Supelco Sigma Aldrich | 243438 | |
| Annexin A2 plasmids | N/A | N/A | Received from our collaborator at the Danish Cancer Research Center |
| Annexin A4 recombinant-protein | N/A | N/A | N-terminal GFP tagged ANXA4 received from our collaborator at the Danish Cancer Research Center |
| Annexin A5 recombinant-protein | N/A | N/A | N-terminal GFP tagged ANXA5 received from our collaborator at the Danish Cancer Research Center |
| beta-casein | Sigma Life Science | C6905-1G | |
| CaCl2 | Suprlco (sigma Aldrich) | 10035-04-8 | |
| Centrifuge 5702 | Eppendorf | 5702 | |
| Chloroform | VWR Chemicals | 67-66-3 | |
| Culture dish (Nunclon Delta Surface) | Thermo scientific | 150460 | |
| DID cell-labelling Solution | Invitrogen | 7757 | |
| Distilled water | Gibco | 15230-089 | |
| DOPC | Avanti Polar Lipids | 850375C | Dissolved in chloroform |
| DOPS | Avanti Polar Lipids | 840035C | Dissolved in chloroform |
| Dulbecco's Modified Eage's Medium | Thermo Fisher Scientific | 11995065 | |
| FIJI ImageJ distribution | ImageJ2 | N/A | |
| GCaMP6s-CAAX | N/A | Received from our collaborator at the Danish Cancer Research Center | |
| Gibco Fetal Bovine Serum | Fisher Scientific | 11573397 | 10% of the culture medium |
| Glucose | PROLABO | 24 374.297 | |
| Hamilton syringes | Hamilton Company | N/A | 50 and 500 microliters |
| Harrick Plasma Cleaner PDG-002 | Harrick Plasma | N/A | |
| HEK293T cells | N/A | Received from our collaborator at the Danish Cancer Research Center | |
| Leica Acousto-Optical Beam Splitter (AOBS) | Leica | N/A | |
| Leica PL APO 63x water immersion objective, NA = 1.2 | Leica | N/A | |
| Leica SP5 confocal scanning microscope | Leica | N/A | |
| Lipofectamine | Fisher Scientific | 15338030 | |
| MatLab | The Mathworks, Inc., Natick, Massachusetts, United States | N/A | |
| NaCl | VWR Chemicals | 7647-14-5 | |
| Opti-MEM Reduced-Serum Medium | Thermo Fisher Scientific | 11058021 | |
| Parafilm | Bemis | PM-992 | |
| Penicillin-Streptomycin | Thermo Fisher Scientific | 15140122 | 1% of the culture medium |
| Phosphate Buffered Saline (PBS) | Thermo Fisher Scientific | 10010023 | |
| Piezoelectric stage (PI 731.20) | Physik Instrumente (Germany) | N/A | |
| Poly-L-Lysine | Sigma-Aldrich | P8920-100ML | 0.01-0.1% for coating |
| Polyvinyl alcohol | Sigma-Aldrich | 363065-25G | |
| round glass slide 25 mm Ø | VWR | 631-1584 | |
| Sonicator Brandson 2800 | Brandson | N/A | |
| sucrose | Sigma Life Science | 57-50-1 | |
| T25 tissue culture flask | Falcon | 353108 | Blue Vented cap |
| Tris-HCl | Invitrogen | 15567-027 | |
| TrypLE | Thermo Fisher Scientific | A1285901 | |
| Trypsin-EDTA | Fisher Scientific | 11590626 | |
| VWR Mixer mini vortex 230V EU | VWR | 12620-84 | ECN: 444-2790, SN: 150713022 |
Références
- Bendix, P. M., et al. Interdisciplinary synergy to reveal mechanisms of annexin-mediated plasma membrane shaping and repair. Cells. 9 (4), 1029 (2020).
- Gajic, O., Lee, J., Doerr, C. H., Berrios, J. C., Myers, J. L., Hubmayr, R. D. Ventilator-induced Cell Wounding and Repair in the Intact Lung. American Journal of Respiratory and Critical Care Medicine. 167, 1057-1063 (2003).
- McNeil, P. L., Khakee, R. Disruptions of muscle fiber plasma membranes. Role in exercise-induced damage. The American Journal of Pathology. 140 (5), 1097-1109 (1992).
- Yu, Q. C., McNeil, P. L. Transient disruptions of aortic endothelial cell plasma membranes. The American Journal of Pathology. 141 (6), 1349-1360 (1992).
- Boye, T. L., et al. Annexin A4 and A6 induce membrane curvature and constriction during cell membrane repair. Nature Communications. 8, 1623 (2017).
- Bischofberger, M., Gonzalez, M. R., van der Goot, F. G. Membrane injury by pore-forming proteins. Current Opinion in Cell Biology. 21, 589-595 (2009).
- Tang, S. K. Y., Marshall, W. F. Self-repairing cells. Science (New York, N.Y.). 356, 1022-1025 (2017).
- Abreu-Blanco, M. T., Verboon, J. M., Parkhurst, S. M. Single cell wound repair: Dealing with life's little traumas. Bioarchitecture. 1, 114-121 (2011).
- Sønder, S. L., et al. Annexin A7 is required for ESCRT III-mediated plasma membrane repair. Scientific Reports. 9, 6726 (2019).
- Andrews, N. W., Almeida, P. E., Corrotte, M. Damage control: cellular mechanisms of plasma membrane repair. Trends in Cell Biology. 24 (12), 734-742 (2014).
- Idone, V., Tam, C., Goss, J. W., Toomre, D., Pypaert, M., Andrews, N. W. Repair of injured plasma membrane by rapid Ca2+-dependent endocytosis. The Journal of Cell Biology. 180 (5), 905-914 (2008).
- Lauritzen, S. P., Boye, T. L., Nylandsted, J. Annexins are instrumental for efficient plasma membrane repair in cancer cells. Seminars in Cell & Developmental Biology. 45, 32-38 (2015).
- Häger, S. C., Nylandsted, J. Annexins: players of single cell wound healing and regeneration. Communicative & Integrative Biology. 12 (1), 162-165 (2019).
- Jaiswal, J. K., et al. S100A11 is required for efficient plasma membrane repair and survival of invasive cancer cells. Nature Communications. 5, 3795 (2014).
- Draeger, A., Monastyrskaya, K., Babiychuk, E. B. Plasma membrane repair and cellular damage control: The annexin survival kit. Biochemical Pharmacology. 81 (6), 703-712 (2011).
- Moreno-Pescador, G. S., et al. Thermoplasmonic nano-rupture of cells reveals annexin V function in plasma membrane repair. Nanoscale. 14 (21), 7778-7787 (2022).
- Zhivotovsky, B., Orrenius, S. Calcium and cell death mechanisms: A perspective from the cell death community. Cell Calcium. 50 (3), 211-221 (2011).
- Gerke, V., Moss, S. E. Annexins: From structure to function. Physiological Reviews. 82 (2), 331-371 (2002).
- Idone, V., Tam, C., Andrews, N. W. Two-way traffic on the road to plasma membrane repair. Trends in Cell Biology. 18 (11), 552-559 (2008).
- Boye, T. L., et al. Annexins induce curvature on free-edge membranes displaying distinct morphologies. Scientific Reports. 8, 10309 (2018).
- Bouter, A., et al. Annexin-A5 assembled into two-dimensional arrays promotes cell membrane repair. Nature Communications. 2, 270 (2011).
- Boye, T. L., Nylandsted, J. Annexins in plasma membrane repair. Biological Chemistry. 397 (10), 961-969 (2016).
- Weinberger, A., et al. Gel-assisted formation of giant unilamellar vesicles. Biophysical Journal. 105 (1), 154-164 (2013).
- Numata, T., Tatsuta, H., Morita, Y., Otani, Y., Umeda, N. Localized thermal processing with a laser-trapped and heated metal nanoparticle. IEEJ Transactions on Electrical and Electronic Engineering. 2, 398-401 (2007).
- Bendix, P. M., Reihani, S. N. S., Oddershede, L. B. Direct measurements of heating by electromagnetically trapped gold nanoparticles on supported lipid bilayers. ACS Nano. 4 (4), 2256-2262 (2010).
- Kyrsting, A., Bendix, P. M., Stamou, D. G., Oddershede, L. B. Heat profiling of three-dimensionally optically trapped gold nanoparticles using vesicle cargo release. Nano Letters. 11 (2), 888-892 (2011).
- Andersen, T., Kyrsting, A., Bendix, P. M. Local and transient permeation events are associated with local melting of giant liposomes. Soft Matter. 10 (24), 4268-4274 (2014).
- Bahadori, A., Oddershede, L. B., Bendix, P. M. Hot-nanoparticle-mediated fusion of selected cells. Nano Research. 10, 2034-2045 (2017).
- Rørvig-Lund, A., Bahadori, A., Semsey, S., Bendix, P. M., Oddershede, L. B. Vesicle fusion triggered by optically heated gold nanoparticles. Nano Letters. 15 (6), 4183-4188 (2015).
- Moreno-Pescador, G., Arastoo, M. R., Ruhoff, V. T., Chiantia, S., Daniels, R., Bendix, P. M. Thermoplasmonic vesicle fusion reveals membrane phase segregation of influenza spike proteins. Nano Letters. 23 (8), 3377-3384 (2023).
- Bahadori, A., Lund, A. R., Semsey, S., Oddershede, L. B., Bendix, P. M. Controlled cellular fusion using optically trapped plasmonic nano-heaters. SPIE Proceedings. SPIE 9922, Optical Trapping and Optical Micromanipulation XIII. 992211, (2016).
- Bahadori, A., Moreno-Pescador, G., Oddershede, L. B., Bendix, P. M. Remotely controlled fusion of selected vesicles and living cells: a key issue review. Reports on Progress in Physics. 81 (3), 32602 (2018).
- Moreno-Pescador, G., Arastoo, M. R., Chiantia, S., Daniels, R., Bendix, P. M. Thermoplasmonic induced vesicle fusion for investigating membrane protein phase affinity. bioRxiv. , (2022).
- Pescador, G. S. M., et al. Investigating plasma-membrane repair employing thermoplasmonics. Biophysical Journal. 120 (3), 45A (2021).
- Moreno-Pescador, G. S., Qoqaj, I., Thusgaard Ruhoff, V., Iversen, J., Nylandsted, J., Bendix, P. M. Effect of local thermoplasmonic heating on biological membranes. SPIE 11083, Optical Trapping and Optical Micromanipulation XVI. 110830M, (2019).
- Bement, W. M., Mandato, C. A., Kirsch, M. N. Wound-induced assembly and closure of an actomyosin purse string in Xenopus oocytes. Current Biology. 9 (11), 579-587 (1999).
- Weisleder, N., et al. Recombinant MG53 protein modulates therapeutic cell membrane repair in treatment of muscular dystrophy. Science Translational Medicine. 4 (139), 139ra85 (2012).
- Sudji, I. R., Subburaj, Y., Frenkel, N., García-Sáez, A. J., Wink, M. Membrane disintegration caused by the steroid saponin digitonin is related to the presence of cholesterol. Molecules. 20 (11), 20146-20160 (2015).
- Babiychuk, E. B., Monastyrskaya, K., Potez, S., Draeger, A. Intracellular Ca2+ operates a switch between repair and lysis of streptolysin O-perforated cells. Cell Death & Differentiation. 16, 1126-1134 (2009).
- Nygård Skalman, L., Holst, M. R., Larsson, E., Lundmark, R. Plasma membrane damage caused by listeriolysin O is not repaired through endocytosis of the membrane pore. Biology Open. 7 (10), bio035287 (2018).
- Swaggart, K. A., et al. Annexin A6 modifies muscular dystrophy by mediating sarcolemmal repair. Proceedings of the National Academy of Sciences of the United States of America. 111, 6004-6009 (2014).
- Yeheskely-Hayon, D., Minai, L., Golan, L., Dann, E. J., Yelin, D. Optically induced cell fusion using bispecific nanoparticles. Small. 9 (22), 3771-3777 (2013).
- Minai, L., Yeheskely-Hayon, D., Golan, L., Bisker, G., Dann, E. J., Yelin, D. Optical nanomanipulations of malignant cells: Controlled cell damage and fusion. Small. 8 (11), 1732-1739 (2012).
- Lukianova-Hleb, E., et al. Plasmonic nanobubbles as transient vapor nanobubbles generated around plasmonic nanoparticles. ACS Nano. 4 (4), 2109-2123 (2010).
- Vogel, A., Noack, J., Hüttman, G., Paltauf, G. Mechanisms of femtosecond laser nanosurgery of cells and tissues. Applied Physics B. 81, 1015-1047 (2005).
- Baffou, G., Polleux, J., Rigneault, H., Monneret, S. Super-heating and micro-bubble generation around plasmonic nanoparticles under cw illumination. Journal of Physical Chemistry C. 118 (9), 4890-4898 (2014).
- Sasikumar, K., Liang, Z., Cahill, D. G., Keblinski, P. Curvature induced phase stability of an intensely heated liquid. Journal of Chemical Physics. 140 (23), 234506 (2014).
- Jauffred, L., Samadi, A., Klingberg, H., Bendix, P. M., Oddershede, L. B. Plasmonic heating of nanostructures. Chemical Reviews. 119 (13), 8087-8130 (2019).
- Neuman, K. C., Nagy, A. Single-molecule force spectroscopy: optical tweezers, magnetic tweezers and atomic force microscopy. Nature Methods. 5 (6), 491-505 (2008).
- Bendix, P. M., Jauffred, L., Norregaard, K., Oddershede, L. B. Optical trapping of nanoparticles and quantum dots. IEEE Journal of Selected Topics in Quantum Electronics. 20, 15-26 (2014).
- Samadi, A., Bendix, P. M., Oddershede, L. B. Optical manipulation of individual strongly absorbing platinum nanoparticles. Nanoscale. 46, 18449-18455 (2017).
- Jørgensen, J. T., Norregaard, K., Tian, P., Bendix, P. M., Kjaer, A., Oddershede, L. B. Single particle and PET-based platform for identifying optimal plasmonic nano-heaters for photothermal cancer therapy. Scientific Reports. 6, 30076 (2016).
- Goldenberg, H., Tranter, C. J. Heat flow in an infinite medium heated by a sphere. British Journal of Applied Physics. 3 (9), 296-298 (1952).
- Eustis, S., El-Sayed, M. A. Why gold nanoparticles are more precious than pretty gold: Noble metal surface plasmon resonance and its enhancement of the radiative and nonradiative properties of nanocrystals of different shapes. Chemical Society Reviews. 35, 209-217 (2006).
- Landau, L. D., Lifshitz, E. M. . Fluid Mechanics: Landau and Lifshitz: Course of Theoretical Physics. 6, (2013).
- Niederauer, C., Seynen, M., Zomerdijk, J., Kamp, M., Ganzinger, K. A. The K2: Open-source simultaneous triple-color TIRF microscope for live-cell and single-molecule imaging. HardwareX. 13, e00404 (2023).
- Richardson, A. C., Reihani, N., Oddershede, L. B. Combining confocal microscopy with precise force-scope optical tweezers. SPIE Proceedings:SPIE 6326, Optical Trapping and Optical Micromanipulation III. 632628, (2006).
- Samadi, A., Klingberg, H., Jauffred, L., Kjær, A., Bendix, P. M., Oddershede, L. B. Platinum nanoparticles: a non-toxic, effective and thermally stable alternative plasmonic material for cancer therapy and bioengineering. Nanoscale. 10 (19), 9097-9107 (2018).
- . Available from: https://www.thermofisher.com/order/catalog/product/A7816 (2023)
- Kreibig, U., Vollmer, M. Theoretical considerations. In: Optical Properties of Metal Clusters. 25, (1995).
- Mie, G. Beiträge zur Optik trüber Medien, speziell kolloidaler Metallösungen. Annalen der Physik. 330 (3), 377-445 (1908).
- Rueden, C. T., et al. ImageJ2: ImageJ for the next generation of scientific image data. BMC Bioinformatics. 18 (1), 529 (2017).
- Schindelin, J., et al. Fiji: An open-source platform for biological-image analysis. Nature Methods. 9, 676-682 (2012).
- Klenow, M. B., Heitmann, A. S. B., Nylandsted, J., Simonsen, A. C. Timescale of hole closure during plasma membrane repair estimated by calcium imaging and numerical modeling. Scientific Reports. 11, 4226 (2021).
- Li, T., Wu, X., Liu, F., Li, N. Analytical methods based on the light-scattering of plasmonic nanoparticles at the single particle level with dark-field microscopy imaging. Analyst. 142 (2), 248-256 (2017).
- Gibbs-Flournoy, E. A., Bromberg, P. A., Hofer, T. P. J., Samet, J. M., Zucker, R. M. Darkfield-Confocal Microscopy detection of nanoscale particle internalization by human lung cells. Particle and Fibre Toxicology. 8 (1), 2 (2011).
- Taylor, R. W., Sandoghdar, V. Interferometric scattering microscopy: Seeing single nanoparticles and molecules via Rayleigh scattering. Nano Letters. 19 (8), 4827-4835 (2019).
- Wu, Y., Ali, M. R. K., Chen, K., Fang, N., El-Sayed, M. A. Gold nanoparticles in biological optical imaging. Nano Today. 24, 120-140 (2019).
- Klingberg, H., Oddershede, L. B., Loeschner, K., Larsen, E. H., Loft, S., Møller, P. Uptake of gold nanoparticles in primary human endothelial cells. Toxicology Research. 4 (3), 566-666 (2015).
Réimpressions et Autorisations
Demande d’autorisation pour utiliser le texte ou les figures de cet article JoVE
Demande d’autorisationThis article has been published
Video Coming Soon
À PROPOS DE JoVE
Copyright © 2025 MyJoVE Corporation. Tous droits réservés.