
Se requiere una suscripción a JoVE para ver este contenido. Inicie sesión o comience su prueba gratuita.
Method Article
Aislamiento y la incorporación de recolección de luz las antenas de la diatomea Cyclotella Meneghiniana en liposomas con lípidos de tilacoides
En este artículo
Resumen
Aquí, presentamos un protocolo para aislar fucoxantina clorofila/c proteínas de unión (FCP) de diatomeas y los incorpore a liposomas con composiciones de lípidos naturales para el estudio de transferencia de energía de excitación a cambios en la composición de iones.
Resumen
El desempeño fotosintético de las plantas, algas y diatomeas depende en la regulación rápida y eficiente de la luz cosecha energía y procesos de transferencia en la membrana de los tilacoides de los cloroplastos. Transferencia eficiente a la reacción fotosintética centros así como en cuanto a la fotoprotección de la luz excesiva y la luz cosecha antena de diatomeas, el llamado fucoxantina clorofila/c proteínas de unión (FCP), son necesarios para la absorción de la luz. El interruptor entre estas dos funciones es una cuestión de muchos años de investigación. Muchos de estos estudios se llevan a con FCP en micelas de detergente. Estudios de interacción, se han eliminado los detergentes, que condujo a una agregación inespecífica de los complejos FCP. En este enfoque, es difícil discriminar entre artefactos y datos fisiológicamente relevantes. Por lo tanto, puede obtenerse información más valiosa acerca de la FCP y otra luz de membrana destino cosecha complejos mediante el estudio de las interacciones proteína-proteína, transferencia de energía y otras características espectroscópicas si están integrados en su entorno nativo del lípido. La principal ventaja es que los liposomas tienen un tamaño definido y una proporción de lípidos/proteínas definidas por el cual se controla el grado de clustering de FCP. Además, fácilmente se pueden simular cambios en la composición de pH y de iones que regulan la luz en vivo . En comparación con la membrana de los tilacoides, los liposomas son más homogénea y menos compleja, que resulta más fácil obtener y entender datos espectroscópicos. El protocolo describe el procedimiento de aislamiento de FCP, purificación, preparación de liposomas e incorporación de FCP en liposomas con composición lipídica natural. Resulta de una aplicación típica se da y se discuten.
Introducción
Organismos fotosintéticos tales como diatomeas deben lidiar con condiciones de luz cambiantes y responder con mecanismos sofisticados de aclimatación que mantener alta eficiencia fotosintética y protegen de daño foto-oxidativo causado por la luz excesiva. Un importante proceso de luz protectora en eucariotas fotosintéticos es la alta energía amortiguamiento (qE) de la luz absorbida que se presenta como la principal contribución a la Temple no fotoquímica (NPQ) bajo condiciones de estrés ligero1,2 ,3. Los complejos antena cosecha luz (LHC) están implicados en la regulación de vías de transferencia de energía de excitación. En respuesta a la luz alta inducida por el bajo pH en el lumen del cloroplasto, los interruptores del sistema de antena de la cosecha de estado al estado apagando la luz. Este estado disipativo de energía protege fotosistemas (PS) y otros complejos en la membrana del thylakoid de la foto-oxidación. En eucariotas fotosintéticos, qE es generalmente inducida por dos factores1,2,3. Un factor es la luz especializada recolección proteína que responde a bajo pH. La proteína subsidiarios induce la qE en las plantas superiores4. LhcSRs5, modulado por la actividad de subsidiarios, inducir la qE en algas verdes6. Las diatomeas poseen Lhcx-como proteínas relacionadas estructuralmente con LHCSRs7,8,9,10.
El segundo factor de qE es el ciclo de la xantofila donde los carotenoides de la antena son convertidos en forma de foto protección de-epoxidación y revertidos por epoxidación. En plantas y algas verdes, violaxantina se convierte en zeaxantina. En diatomeas, diadinoxanthin se convierte en diatoxanthin, que luego se correlaciona con el grado de NPQ11. La luz de la diatomea cosecha antena posee algunas peculiaridades aunque es evolutivo relacionado con plantas y algas LHCs. El interruptor de la luz cosecha a fotoprotección es enormemente rápido y la capacidad NPQ es superior comparada con las plantas12. Esta podría ser una razón por la cual las diatomeas muy exitosas en diferentes nichos ecológicos de manera que son responsables de hasta un 45% de la producción primaria neta oceánico13. Por lo tanto, luz cosecha sistemas de diatomeas son un interesante objeto de investigación de la fotosíntesis.
Diatomeas, como las centradas en especies de Cyclotella meneghiniana, poseen tilacoides luz intrínseca cosecha sistemas nombrados después de los pigmentos unen - fucoxantina, clorofila (chl) a y c, por lo tanto luz FCP. recolección de proteínas, como la FCPs, son incrustado en el sistema de membranas de tilacoides que comprende varias capas de membrana. Las diatomeas forman bandas de tres tilacoides. Este complejo situación dificulta estudiar en el nivel molecular en la membrana de los tilacoides. Además, muchos componentes contribuyen a la regulación de la luz cosecha (véase arriba). Por lo tanto, en muchos enfoques, los complejos fueron aislados de la membrana usando detergentes suaves, tales como n-dodecil-β-D-maltopyranoside (β-DDM), que solubilizan la membrana pero mantener intacto los complejos FCP. Se realizaron muchos estudios espectroscópicos uso FCP solubilizado para investigar energía intramolecular transferencia14,15,16,17. Sin embargo, este enfoque anterior era limitada ya que la regulación de la transferencia de energía necesita interacción excitonic con otros complejos de antena o fotosistemas. Por lo tanto, este tipo de estudios no se puede realizar con complejos solubles debido a la interacción entre complejos se pierde.
Una característica importante en la regulación de la antena es la "apretadura molecular" de la antena y fotosistemas en la membrana tilacoides del18. Anteriormente, se llevó a cabo un enfoque simple para simular este efecto en vitro. El detergente fue quitado, que conduce a la agregación aleatoria de complejos antena. Aunque se obtuvieron algunos datos razonables por este enfoque17,19, detergente no refleja la situación en vivo y tiene algunas limitaciones ya que los complejos no están interactuando en los terciarios regulares estructura.
El uso de liposomas supera varias de las limitaciones anteriores. La estructura terciaria sigue siendo totalmente intacta. La membrana del liposoma proporciona un entorno cuasi-nativo para los complejos de antena. La membrana separa el interior de la liposoma del ambiente exterior. Por estos medios, liposomas proporcionan dos compartimientos de la reacción para estudios de gradientes de iones y pH, así como por procesos de transporte. Además, pueden controlarse más fácilmente los parámetros del sistema experimental de estudios de la membrana del thylakoid. Liposomas ya fueron demostrados para ser una excelente herramienta para el estudio de complejos fotosintéticos. Un enfoque principal en el pasado fue en la planta donde se probó el efecto de la composición lipídica alterada en LHC II20LHC. En otros enfoques, interacción de la proteína-proteína entre diferentes LHC II fueron investigados21. También, algunos en algas verdes se realizaron estudios que describen la agrupación espontánea entre el LHC22. Considerando la importancia de las diatomeas para los ecosistemas acuáticos, relativamente pocos estudios se realizaron con los complejos antena de diatomeas. Dos estudios investigaron los complejos antena de la centrada en la Cyclotella meneghiniana, donde se muestra el agrupamiento de la FCP antena23 y capacidad de respuesta de FCP a gradientes electroquímicos24 . Así, los liposomas son una excelente herramienta para el estudio de diatomeas antenas y su interacción y regulación en condiciones casi nativas. Los liposomas son versátiles desde muchas condiciones tales como la composición de lípido, liposomas tamaño, densidad de la proteína y la fase acuosa circundante puede ser controlada. Además, el método requiere pequeñas cantidades de muestras. El sistema experimental es robusto y altamente reproducibles. Permite la compartimentalización de liposomas para el estudio de pH y los gradientes de iones, que son importantes factores en la regulación de los complejos antena.
Aquí, describimos el aislamiento de complejos de antena FCP de C. meneghiniana y su incorporación en los liposomas con composición de lípido natural tilacoides. También, proporcionamos datos ejemplares para la caracterización espectroscópica de FCP solubilizado y compararlos con FCP en liposomas. El método resume conocimientos y protocolos estandarizados de las mejoras de Gundermann y Büchel 201223, Natali et al 201622y Ahmad y Dietzel 201724.

Figura 1: representación esquemática del flujo de trabajo. (1) se refiere al párrafo 1 que describe el crecimiento celular, desorganización y aislamiento de tilacoides con la siguiente separación de FCP en gradientes de densidad de sacarosa; M. C. -Células deCyclotella meneghiniana . (2) preparación de la mezcla de lípidos naturales tilacoides (MGDG, DGDG y SQDG) había descrito en el párrafo 2 y creación de micelas de detergente de lípidos con octylglycoside (OG). Se consigue un tamaño de micela de lípidos definidos por extrusión utilizando membranas de diámetro de poro definido. FCP y lípido micelas se unifican en un lípido predefinido: relación y los detergentes OG y β-DDM son quitado vía controlado diálisis formando proteoliposomes FCP. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}
Protocolo
Nota: Complejos fotosintéticos como FCPs son altamente vulnerables a la luz y el calor. Trabaje siempre en el hielo y bajo una luz muy tenue.
1. aislamiento de FCP de células
-
Aislamiento de tilacoides de las células C. meneghiniana
- Crecer meneghiniana C. en cinco matraces de 500 mL que cada una llena con 300 mL de medio de ASP23,25 y 50 millones de células. Conecte los matraces con un tapón de algodón y permita que las células a crecer hasta la fase de crecimiento exponencial durante aproximadamente una semana en un agitador a 120 rpm con una 16 h luz y fase oscura 8 h 40 μmol photons/(m²s) blanca luz y una temperatura entre 15-18 ° C. Verifique que el numero de celular es entre 1,5 millones de células/mL con una cámara del contador de célula.
- Centrifugar las células a 4.000 x g en un rotor preenfriado con frascos de 500 mL centrífuga (4 ° C) durante 15 minutos en una centrifugadora de alta velocidad. Resuspender el pellet celular en 12 mL de buffer de homogenización (HB, tabla 1) mediante pipeteo.
- Transferir la suspensión a un tubo de plástico de 50 mL solo. Almacenar las muestras a-80 ° C o proceda al paso 1.1.3.
- Pre-enfriar el molino de grano y el equipo. Llenar el vaso de precipitados de 50 mL del molino de grano hasta un 75% con una mezcla de grano de cristal y añadir la suspensión de células. Para la interrupción de la célula, utiliza pulsos de 7 x 45 s a toda velocidad con 30 s de enfriamiento entre cada pulso. Tomar 20 μl de células interrumpidas por los controles de calidad en el paso 1.1.6.
- Filtrar las células interrumpidas sobre un embudo de filtro de cristal y lavado vertiendo la HB en los granos de cristal hasta que aparezcan claros. La fracción de lavado con el filtrado de la piscina. Mantener el volumen final menor de 150 mL.
- Centrifugar la muestra durante 15 min a 140 x g utilizando tres tubos de plástico de 50 mL para que sedimenten los restos celulares. Transferir el sobrenadante a viales de 20 mL policarbonato ultracentrifugación y descartar el precipitado.
- Llenar los frascos con HB, equilibrar el peso y centrífuga de un rotor adecuado para 1 h en 300.000 x g y 4 ° C para que sedimenten las membranas tilacoides.
- Utilizar el tiempo de centrifugación para comprobar la proporción de células interrumpidas por microscopia con 400 aumentos con la 20 μl de muestra en 1.1.3. Calcular la relación libre de cloroplasto cloroplasto que contiene frustules (conchas de sílice).
Nota: Las paredes celulares de diatomeas están hechas de sílice, que es visible como altamente diffracting la sustancia en el microscopio. Cloroplastos que ocurren puntos como verdes deben han sido liberados de las células si trabajó la interrupción de la célula. - Resuspenda el sedimento de membrana con buffer de lavado poco como sea posible (0.5-1 mL) con pincel de un pintor pequeño. Llene los frascos de la ultracentrifugación de policarbonato lavado tampón (tabla 1), equilibre su peso y centrifugar durante 20 min a 200.000 x g a 4 ° C.
- Vuelva a suspender las membranas lavadas con cepillo de pintor. Añadir tampón de lavado sólo si es necesario para mantener la concentración de tilacoides más alta posible. Los tilacoides en frasco (15 mL) de una muestra de la piscina.
- Pre-diluir las muestras con 10 μl de la muestra de 90 μl de acetona al 100%. Centrifugar a 12.000 x g durante 5 minutos para que sedimenten los precipitados de la proteína. Tomar 10 μl de la dilución y se mezcla con 990 μl de acetona al 90%.
- Medir la absorbancia (ABS) de clorofila a y c a 664 nm y 630 nm en acetona al 90%. Reste el ABS750nm de ambos valores. Determinar el contenido de clorofila total usando la siguiente fórmula:24
1)
2)
- Medir la absorbancia (ABS) de clorofila a y c a 664 nm y 630 nm en acetona al 90%. Reste el ABS750nm de ambos valores. Determinar el contenido de clorofila total usando la siguiente fórmula:24
- Alícuotas tilacoides en porciones de 0,5 mg de clorofila total en un tubos de reacción de 1,5 mL, congelar en nitrógeno líquido y almacenarlas a-80 ° C hasta su uso posterior.
-
Separación y concentración de complejos FCP
- Preparar una solución de gradiente de sacarosa y llenar los tubos de la ultracentrífuga hasta la parte superior menos el volumen de carga (300 – 500 μl). Se congelan los tubos a-20 ° C hasta que estén completamente congelados. Permita que los tubos descongelar a + 4 ° C, que toma 3-4 h de un tubo de 17 mL.
- Repetir el ciclo de congelación-descongelación dos veces para refinar el gradiente para la mejor resolución.
- Utilizar las muestras obtenidas en 1.1.9 correspondientes a 0,5 mg de clorofila y ajustar con el almacenador intermediario B1 (tabla 1) hasta un volumen final de 2 mL. Para la solubilización, añadir n-dodecil-β-D-maltopyranoside (b-DDM) a una concentración final de 20 mM.
- Invertir el tubo 3 veces y coloque en hielo por 20 min con agitación suave para evitar la espuma. Centrifugar durante 5 min, a 12.000 x g en una centrífuga de mesa previamente enfriado a + 4 ° C.
- El sobrenadante en el gradiente de carga. No cargue más de 125 μg de clorofila total por gradiente si se utilizan frascos de 17 mL. Centrífuga de 22 h a 100.000 x g y + 4 ° C.
- Recuperarse las fracciones deseadas de FCP marrón degradado utilizando una jeringa (figura 2A). Tomar la alícuota de 5 μl y diluirlo con 995 μl de B1a.
- Medir el espectro de absorbancia (ABS) entre 370-750 nm en un espectrofotómetro UV-VIS. Utilizar cubetas semi-micro vidrio óptico.
- Abra el software de administrador de espectros, modo de espectro y registrar una línea base de 370 a 750 nm seguido por medidas de muestra. Utilice la siguiente configuración: scan velocidad, 200 nm/min; campo de datos, 0,5 nm; respuesta, medio.
- Medir el volumen de la muestra recuperada con una micropipeta. Lave los complejos FCP añadiendo dos veces el volumen recuperado con tampón de B1 (tabla 1). Se concentran en un concentrador de membrana con un corte a 1.000 x g y + 4 ° C a un ABS672nm de al menos 20 kDa 30.
Nota: La concentración de b-DDM podría aumentar debido al enriquecimiento de la micela en el compartimiento de muestra. ¡Esto podría llevar a la mayor solubilización de los complejos FCP! Evitar exceso de solubilización mediante la adición de detergente búferes libres B1 si lava más pasos son necesarios para extraer la sacarosa residual. - Tomar la alícuota para los controles de 20 μl. Descarga congelar las muestras en nitrógeno líquido y almacenarlas a-80 ° C.
- Preparar una solución de gradiente de sacarosa y llenar los tubos de la ultracentrífuga hasta la parte superior menos el volumen de carga (300 – 500 μl). Se congelan los tubos a-20 ° C hasta que estén completamente congelados. Permita que los tubos descongelar a + 4 ° C, que toma 3-4 h de un tubo de 17 mL.

Figura 2: purificación de FCP, controles espectroscópicos y comprobar pureza. (A) aspecto típico de un gradiente de densidad de sacarosa después de la centrifugación durante la noche. Todas las bandas de color marrón contienen la piscina FCP consisten en pigmentos FCPa y FCPb.. - pigmentos, PS - espectros de absorción (B) de fotosistemas de FCP antes (línea azul) y después de concentración (línea naranja discontinua) con dispositivos de filtro centrífugo de 30 kDa de corte . Particularmente, los carotenoides son propensos a la pérdida de la FCP, que daría como resultado menor absorbancia en la región entre 500-550 nm. Los gráficos se normalizan para la clorofila Qy máximo a ~ 670 nm. Espectros de emisión de clorofila a (C) con excitación de chl c (465 nm) para las pruebas de la transferencia de energía de excitación funcional. Si la transferencia de energía de chl c a chl a es obstaculizado, una banda de fluorescencia adicional en ~ 640 nm (chl c) tendría lugar. Los gráficos se normalizan para la emisión máxima. (D) espectros de excitación grabado en 675 nm (chl una fluorescencia máxima) para probar la transferencia de energía a chl un de los pigmentos de absorción entre 370 nm y 600 nm. Si la transferencia de energía a chl a es menos eficiente, lo que disminuiría el rendimiento de fluorescencia especialmente entre 465 y 550 nm. Los gráficos se normalizan al máximo alrededor de 440 nm. Los espectros (B), (C) y (D) son casi idénticos si la concentración funcionó bien. (E) Compruebe la pureza de la FCP aislada usando un gel Tris-Tricina del28. FCPa y FCPb tienen subunidades entre 18-19 kDa. Todas visibles plata-manchados las proteínas más grandes que 20 kDa son contaminantes. Thyl. -Tilacoides haga clic aquí para ver una versión más grande de esta figura.
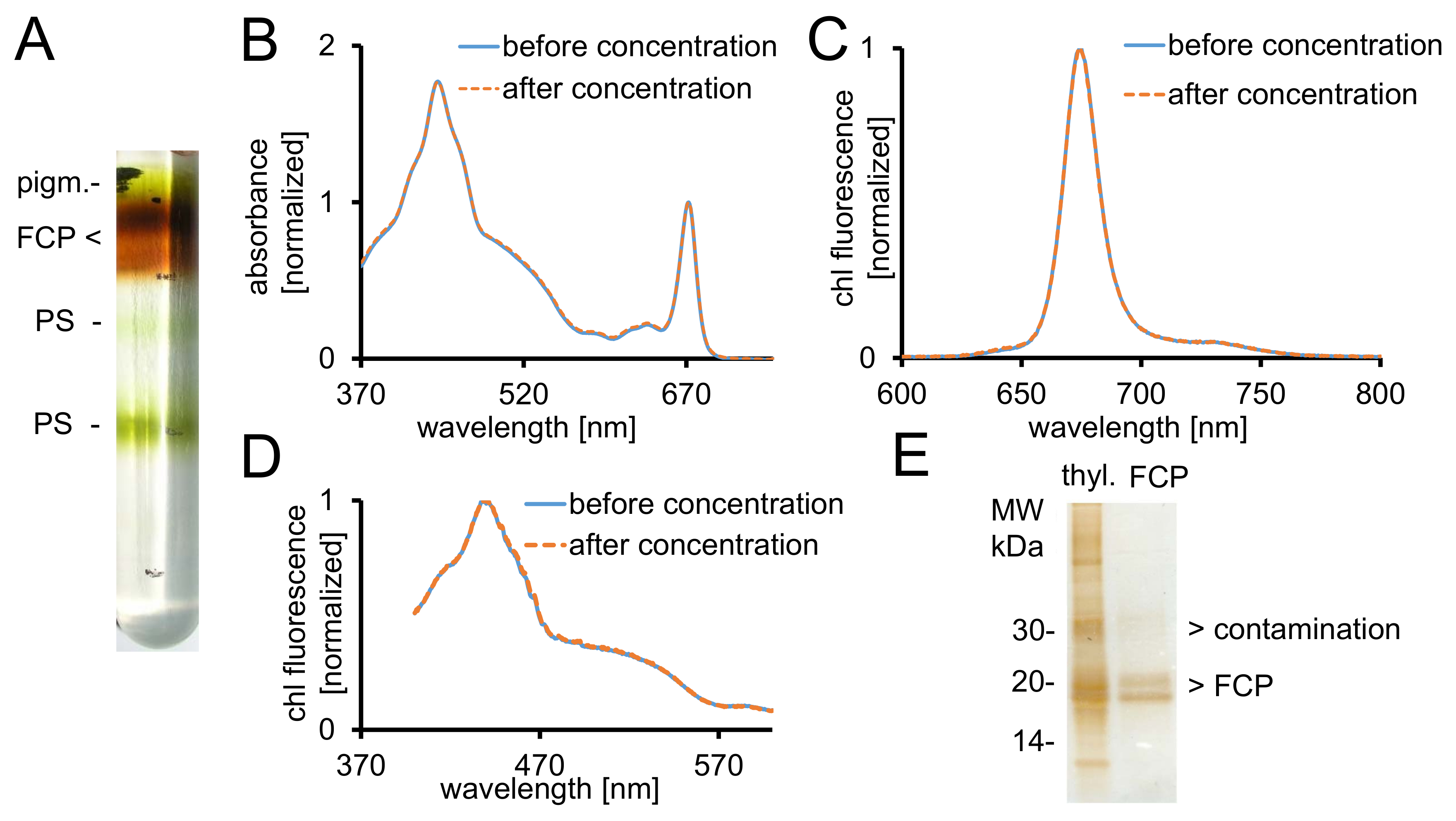{kind=link}
-
Controles basados en gel y espectroscópicos
- Registre la absorbancia entre 370-750 nm de 5 μl de FCP en 995 μl de B1a después paso 1.2.5. Compararlo con el espectro obtenido en el paso 1.2.4. Utiliza la misma instrumentación y configuración como se describe en el paso 1.2.4.
- Exportar datos a *.csv e importar los datos en una hoja de cálculo. Normalizar ambos espectros hasta el máximo de chl un en la banday Q unos 672 nm como se muestra en la figura 2B.
- Calcular el factor de dilución de las muestras del paso 1.2.4 y 1.3.1 dividiendo la medida ABS672nm de la deseada ABS672nm de 0.03. Diluir las muestras en consecuencia con B1a para cubetas de vidrio especial de la fluorescencia.
- Grabar chl un espectro de emisión de fluorescencia con un spectrofluorometer a revelar la transferencia de energía de la luz intacta de chl c a chl (figura 2).
- Utilizar el software de espectrómetro y vaya al modo de medición de espectro con los siguientes parámetros: modo, emisión; corte ancho excitación y emisión, 3 nm; sensibilidad, medio; velocidad de escaneo, 100 nm/min; campo de datos, 0,5 nm; longitud de onda de excitación, 465 nm; emisión, 600-800 nm. Realizar AUTOCERO y medida.
- Registrar espectros de excitación con la misma muestra y equipos para la transferencia de la energía intacta de todos los pigmentos a chl (Figura 2D). Cambiar configuración: modo, excitación; longitud de onda de emisión, 675 nm; excitación, 370-600 nm. Registrar los espectros.
- Corregir con un espectro de rodamina en el mismo rango de propiedades espectrales de la luz – cf. instrucciones en el manual del usuario.
- Las muestras de mezcla FCP corresponde a 1 μg de chl una con 10 μl de tampón de carga SDS. Incubar 10 min a 25 ° C. Centrifugar durante 5 minutos a 12000 x g en una centrífuga de mesa.
- Cargar el sobrenadante en un Tris-Tricina gel28. Separadas por 2 h a 150 V y plata mancha lo después de los40de la separación.
Nota: Las subunidades FCP separan en dos bandas prominentes de proteína entre 18-19 kDa, que son constituyentes de la FCPa y FCPb29 (Figura 2E).
- Cargar el sobrenadante en un Tris-Tricina gel28. Separadas por 2 h a 150 V y plata mancha lo después de los40de la separación.
- Registre la absorbancia entre 370-750 nm de 5 μl de FCP en 995 μl de B1a después paso 1.2.5. Compararlo con el espectro obtenido en el paso 1.2.4. Utiliza la misma instrumentación y configuración como se describe en el paso 1.2.4.
2. preparación de liposomas e incorporación de FCP
-
Preparación de la mezcla de lípidos y micelas de detergente de lípidos
Nota: lípidos son susceptibles a las cálidas temperaturas combinadas con condiciones oxidativas. Trate de mantener los lípidos refrigerados y bajo atmósfera de N2 .- Calcular las proporciones de lípidos tilacoides deseada para C. meneghiniana según Vieler et al. 200730. Consulte el ejemplo en suplementales tabla 1. Preparar el lípido soluciones recomendadas por el fabricante en un contenedor de prueba solvente.
- Pipetear la cantidad deseada de lípidos en un tubo de ensayo 2 mL y evaporar el cloroformo utilizando un flujo suave de nitrógeno y tratar de difundir los lípidos sobre el área entero de la base del tubo. Que el flujo de2 N hasta que se evapora el solvente todos.
- Solubilizar la mezcla de lípidos en 29 μl de solución de β-D-glucopyranoside de n-octilo (OG) a 4 ° C por 4 h. Incubar la mezcla de lípidos durante 10 minutos a 30 ° C. Incubar los lípidos en un baño sonicador para 3 x 3 min a 25 ° C interrumpido por 30 s en el hielo.
- Añadir 221 μl de tampón tricino y 250 μl de buffer de diálisis de 4 x.
- Utilizar una extrusora con membranas de policarbonato de 0,1 μm para el de 50-70 nm de diámetro definido liposoma. Montar la extrusora con el apoyo de membrana y filtro. Evitar las burbujas de aire y apriete el conjunto de fondo.
- Llene una jeringa con buffer de diálisis de 4 x y Humedezca previamente el estirador hasta que las burbujas no se pueden ver en la segunda jeringa.
- Aplicar las micelas de detergente de lípidos a la extrusora y presione la solución de una jeringa al otro adelante y atrás. Repita este paso 5 veces hasta que la solución aparece homogénea.
Nota: Esta solución puede conservarse a 4 ° C durante varios días. ¡No lo congele!
-
Incorporación de complejos FCP y eliminación de detergentes y agregados
Nota: En este ejemplo utilizamos un lípido/chl la proporción 12:1, que corresponde a una relación proteína/lípidos de aproximadamente 100: 1.- Añadir FCP igual a 20 μg de chl una en un volumen total de 500 μl de tampón de B1a a 250 μL de las micelas de lípidos extruidas y 250 μl de buffer de diálisis 4. Incubar las muestras por 3 x 3 min a 25 ° C en un Termomezcladores a 1.500-3.000 rpm interrumpido por una pausa 30 de s sobre el hielo.
- Cortar la tapa del tubo de reacción de 1,5 mL cuatro justo debajo de la parte superior dando un anillo que todavía cabe en la tapa. Preparar piezas de 1,5 cm x 1,5 cm de la membrana de diálisis y lávelos con 20 mL de buffer de diálisis x 1
- Llenar 250 μl de la muestra para cada tapa. Cuidadosamente, coloque la membrana sobre la tapa para que el compartimiento se llena completamente con la muestra y sin burbujas de aire se producen. Apriete el anillo del tubo de reacción en la Asamblea con el fin de tener un compartimento cerrado.
- Dializan las muestras en 50 mL de 1 x de buffer de diálisis durante la noche (12-16 h) en el hielo en una coctelera de agitación. El buffer de diálisis usado con uno nuevo y se añade 7 mg de adsorbente granos para quitar los restos de detergente durante al menos 6 h.
- Vuelva a colocar el buffer de diálisis y dializarse para otro 12 h. recuperar los liposomas por la perforación de la membrana de diálisis con una punta de micropipeta de 200 μl y aspirar los liposomas de la tapa del tubo de reacción.
- Paso opcional: si de alta pureza (> 95%) es necesario. Preparar un gradiente de densidad discontinuo en 17 mL frascos de ultracentrifugación con pasos que contiene copolímero epichlorhydrin de sacarosa 6%, 10%, 15% y 20% en buffer de diálisis. Los liposomas y ultracentrífuga a 100.000 x g durante 4 horas en un rotor que hace pivotar del cubo de la carga.
- Recuperar la banda superior de color marrón con una jeringa, diluir la muestra 1:5 con DP y proceder al siguiente paso.
- Centrifugue los liposomas FCP en al menos 2 mL de 1 x de buffer de diálisis durante 1,5 h a 100.000 x g a 4 ° C. Recuperar los liposomas girando el tubo de centrífuga en un ángulo de 45°. Permita que los liposomas para abajo durante 1 minuto (Figura 3A).
- Recuperar los liposomas FCP en un volumen final de 25-50 μl. evitar perturbar el precipitado.
-
Controles 1: absorbancia, espectroscopia de fluorescencia
- Añadir una alícuota de 3 μl de los liposomas de FCP a un volumen final de 1 mL de 1 x de buffer de diálisis y centrifugar durante 5 min a 12.000 x g.
- Registre la absorbancia entre 370 y 750 nm de FCP-liposomas con el mismo equipo como se describe en 1.2.4. Normalizar el espectro al máximo en la regióny de Q de chl un (670-680 nm) y se compara con el espectro normalizado de FCP solubilizado (figura 3).
- Preparar liposomas FCP en 1 mL de DP con una absorbancia (ABS) = 0.03 con respecto a la máxima entre 670-680 nm. Ajustar la FCP solubles en detergente de paso 1.2.6 el mismo ABS diluir con B1a.
- Registrar los espectros de absorbancia de ambas muestras como se describe en 1.2.4. Grabar chl un espectro de emisión de fluorescencia de ambas muestras como se describe en 1.3.3.
Nota: El rendimiento de fluorescencia está disminuido en la muestra de FCP-liposoma (figura 3D y CF. discusión.)
- Añadir una alícuota de 3 μl de los liposomas de FCP a un volumen final de 1 mL de 1 x de buffer de diálisis y centrifugar durante 5 min a 12.000 x g.

Figura 3: aislamiento de FCP proteoliposomes seguido por controles espectroscópicos y proyección de imagen confocal. (A) recuperación de liposomas FCP después de la centrifugación. Gire el tubo de centrifugación a 45° y espere aproximadamente 1 minuto, los liposomas se moverá hacia abajo mientras que el FCP agregados que no están incorporados en liposomas se adhieren a la pared del tubo. (B) comparación de espectros de absorbancia de FCP solubles en detergente (azul) y FCP en liposomas (naranja) (C) los mismos espectros como en (B) normalizaron a chl un máximo en la región roja (~ 670 nm - Qy pico); FCP solubles en detergente (azul) y FCP en liposomas (naranja). Potencialmente, podría haber una pérdida de pigmento principalmente de carotenoides visible en la región de 500-550 nm. El agrupamiento de FCP en los liposomas puede conducir a una ampliación de pico y un cambio leve de la chl un máximo (~ 670 nm) al rojo. (D) espectros de emisión de FCP solubles en detergente y FCP en liposomas. Agrupamiento de FCP en el liposoma mejora energéticas interacciones de complejos FCP que baja el rendimiento de fluorescencia (curva naranja) y cambia los máximos de emisión ligeramente al rojo. Haga clic aquí para ver una versión más grande de esta figura.
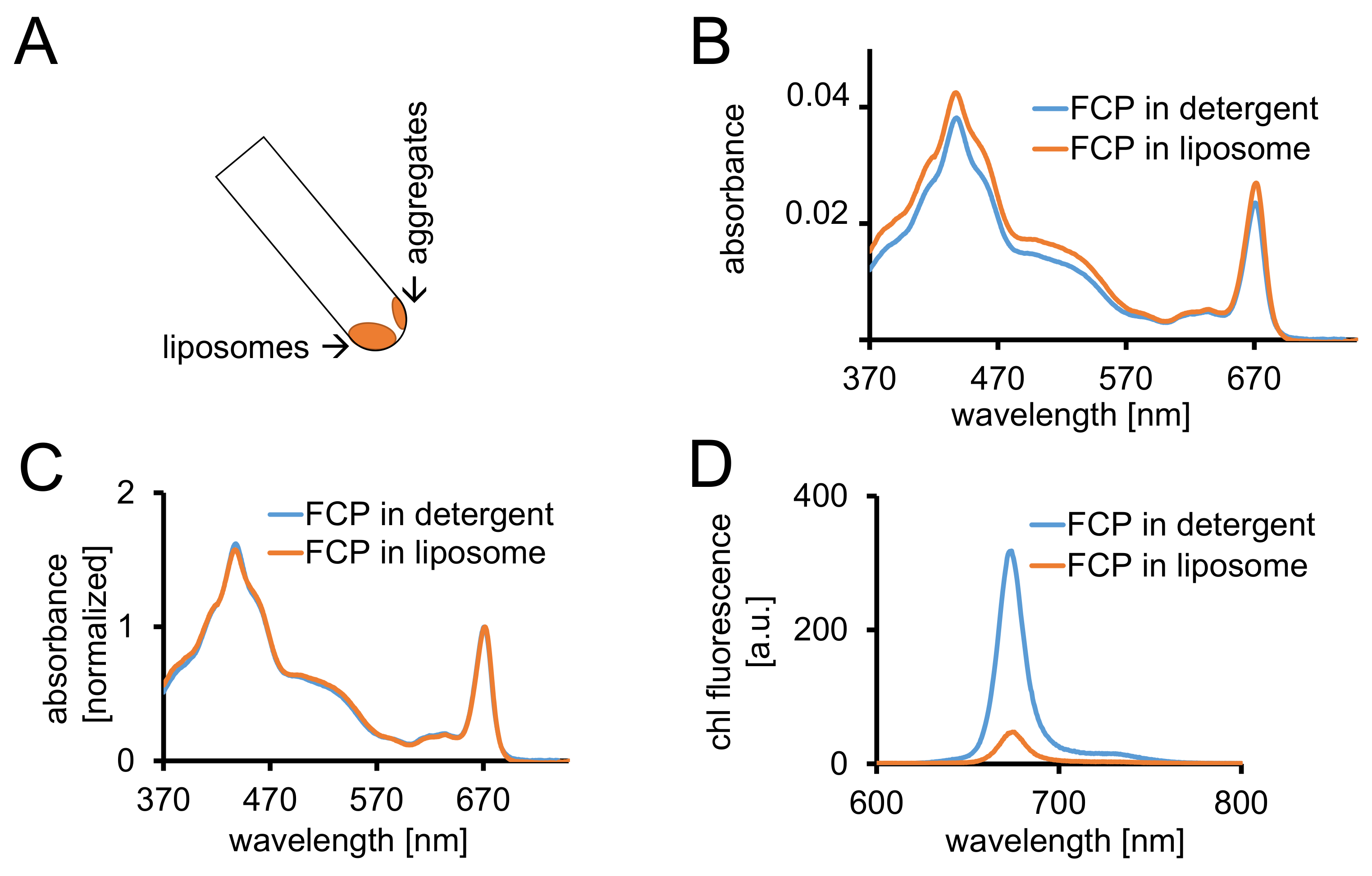{kind=link}
Resultados
El protocolo describe el aislamiento de la fracción total de FCP de Cyclotella meneghiniana e incorporación de liposomas con composición de lípido nativo. El aislamiento de thylakoid es altamente reproducible, pero puede cambiar el rendimiento de tilacoides. El resultado es aceptable si más del 50% de todos los pigmentos son recuperado en el paso 1.1.4. Más del 80% es óptima.
La solubilización de los tilacoides ...
Discusión
Liposomas FCP con la composición de lípido natural proporcionan una herramienta práctica, simple y reproducible para investigar propiedades espectroscópicas in vitro. El medio lipídico en liposomas FCP se asemeja a la situación dentro de la membrana del thylakoid, dando lugar a resultados experimentales que están más cercanos de las condiciones naturales.
Hay varias ventajas de utilizar meneghiniana C. como sistema modelo para antena FCP. Crece relativamente rápido y...
Divulgaciones
Los autores no tienen nada que revelar.
Agradecimientos
Agradecemos a Rana Adeel Ahmad para ayuda en la purificación de la FCP. Prof. Claudia Büchel se reconoce para las discusiones útiles y leyendo el manuscrito. Este trabajo fue financiado por la Fundación alemana de investigación de LD (DI1956-1/1) y la Fundación Humboldt para una Beca Feodor Lynen para ld
Materiales
| Name | Company | Catalog Number | Comments |
| 500 ml centrifuge vials | |||
| high speed centrifuge | Heraeus | ||
| Bead Mill VI 2 | Edmund-Bühler (edmund-buehler.de) | newer version: Vibrogen-Zellmühle Vl 6 | |
| Silibeads S 400 µm | Sigmund-Lindner.com | 5223-7 | |
| Silibeads S 1,-1,3 mm | Sigmund-Lindner.com | 4504 | |
| VitraPOR filter funnel - por1 | ROBU GmbH | 21121 | |
| polycarbonate ultracentrifuagtion vials (30 mL) for T-865 | Beranek Laborgeräte (Laborgeraete-beranek.de) | 314348 | |
| Ultracentrifuge Discovery 90SE | Sorvall | n.a. | |
| rotor T 865 | ThermoFisher Scientific (thermofisher.com) | 51411 | |
| Neubauer Cell Counter Chamber (improved) | Carl Roth Laborbedarf (Carlroth.com) | T729.1 | |
| Zeiss Mikroskop Primostar (7) | Optik-Pro (optik-pro.de) | 51428 | |
| optical glass cuvettes (6040-OG) | Hellma Analytics (hellma-analytics.com) | "6040-10-10" | |
| V-630 UV-VIS Spectrophotometer (incl. software) | Jasco (jasco.de) | V-630 | |
| n-Dodecyl-β-D-Maltopyranoside | ANATRACE (anatrace.com) | D310LA | |
| Ultra-Clear tubes 17 ml for AH629 | Beranek Laborgeräte (Laborgeraete-beranek.de) | 344061 | |
| rotor AH629-17-mL | ThermoFisher Scientific (thermofisher.com) | 54285 | |
| Membrane concentrator_Centriprep 30 kDa cutoff | Millipore (merckmillipore.com) | 4307 | |
| Biometra Minigel-Twin | Analytik Jena AG (analytik-jena.de) | 846-010-100 | |
| Silver Stain Plus Kit | Bio-Rad (bio-rad.com) | 1610449 | |
| libre office spread sheet | The document foundation | https://de.libreoffice.org/download/libreoffice-still/ | |
| special glass cuvettes for fluorescence (101-0S) | Hellma Analytics (hellma-analytics.com) | 101-10-20 | |
| Spectrofluorometer FP-6500 (incl. Software) | Jasco (jasco.de) | FP-6500 | |
| SDS-loading buffer Roti-Load | ROTH (carlroth.com) | K929.1 | |
| n-octyl β-D-glucopyranoside | ANATRACE (anatrace.com) | O311 | |
| Monogalactosyl Diaclyglycerol (MGDG) | Larodan AB (larodan.com) | 59-1300 | make stock solution in chloroform |
| Digalactosyl Diacylglycerol (DGDG) | Larodan AB (larodan.com) | 59-1310 | make stock solution in chloroform |
| Sulphoquinovosyl Diacylglycerol (SQDG) | Larodan AB (larodan.com) | 59-1230 | make stock solution in chloroform |
| L-alpha-Phosphatidylglycerol (PG) | Larodan AB (larodan.com) | 37-0150 | make stock solution in chloroform |
| L-α-Phosphatidylcholine | Sigma-Aldrich (sigmaaldrich.com) | P3782 SIGMA | make stock solution in chloroform |
| sonicator bath S-50TH | Sonicor (getmedonline.com | SONICOR-S-50TH | |
| mini-Extruder | Avanti Polar Lipids (Avanti.com) | 610000 | |
| Nuleopore polycarbonate membrane | Avanti Polar Lipids (Avanti.com) | 610005 | |
| dialysis membrane Visking 14 kDa cutoff | ROTH (carlroth.com) | 0653.1 | boil in destilled water before use |
| Biobeads SM2 Adsorbent | Biorad (Bio-rad.com) | 152-3920 | |
| sucrose epichlorhydrin copolymer - Ficoll 400 | Sigma-Aldrich (sigmaaldrich.com) | F4375 | |
| Polycarbonate ultracentrifuagtion vials (2.7 mL) for TFT 80.4 | Beranek Laborgeräte (Laborgeraete-beranek.de) | 252150 | |
| rotor TFT 80.4 | Millipore (merckmillipore.com) | 54356 | |
| material listed in order of appearance | For specific safety instructions please refer to material safety sheets and repective manuals. Standard lab material and substances are not listed. |
Referencias
- Eberhard, S., Finazzi, G., Wollman, F. A. The Dynamics of Photosynthesis. Annual Review of Genetics. 42, 463-515 (2008).
- Li, Z. R., Wakao, S., Fischer, B. B., Niyogi, K. K. Sensing and Responding to Excess Light. Annual Review of Plant Biology. 60, 239-260 (2009).
- Niyogi, K. K., Truong, T. B. Evolution of flexible non-photochemical quenching mechanisms that regulate light harvesting in oxygenic photosynthesis. Current Opinion in Plant Biology. 16 (3), 307-314 (2013).
- Li, X. -. P., et al. A pigment-binding protein essential for regulation of photosynthetic light harvesting. Nature. 403 (6768), 391-395 (2000).
- Peers, G., et al. An ancient light-harvesting protein is critical for the regulation of algal photosynthesis. Nature. 462 (7272), 518-521 (2009).
- Correa-Galvis, V., et al. Photosystem II Subunit PsbS Is Involved in the Induction of LHCSR Protein-dependent Energy Dissipation in Chlamydomonas reinhardtii. The Journal of biological chemistry. 291 (33), 17478-17487 (2016).
- Bailleul, B., et al. An atypical member of the light-harvesting complex stress-related protein family modulates diatom responses to light. Proceedings of the National Academy of Sciences of the United States of America. 107 (42), 18214-18219 (2010).
- Taddei, L., et al. Multisignal control of expression of the LHCX protein family in the marine diatom Phaeodactylum tricornutum. Journal of experimental botany. 67 (13), 3939-3951 (2016).
- Lepetit, B., et al. The diatom Phaeodactylum tricornutum adjusts nonphotochemical fluorescence quenching capacity in response to dynamic light via fine-tuned Lhcx and xanthophyll cycle pigment synthesis. New Phytologist. 214 (1), 205-218 (2017).
- Büchel, C. Evolution and function of light harvesting proteins. Journal of Plant Physiology. 172, 62-75 (2015).
- Lavaud, J., Rousseau, B., van Gorkom, H. J., Etienne, A. -. L. Influence of the Diadinoxanthin Pool Size on Photoprotection in the Marine Planktonic Diatom Phaeodactylum tricornutum. Plant Physiology. 129 (3), 1398-1406 (2002).
- Ruban, A., et al. The super-excess energy dissipation in diatom algae: comparative analysis with higher plants. Photosynthesis Research. 82 (2), 165-175 (2004).
- Mann, D. G. The species concept in diatoms. Phycologia. 38 (6), 437-495 (1999).
- Papagiannakis, E., van Stokkum, I. H. M., Fey, H., Büchel, C., van Grondelle, R. Spectroscopic Characterization of the Excitation Energy Transfer in the Fucoxanthin-Chlorophyll Protein of Diatoms. Photosynthesis Research. 86 (1-2), 241-250 (2005).
- Premvardhan, L., Robert, B., Beer, A., Büchel, C. Pigment organization in fucoxanthin chlorophyll a/c2 proteins (FCP) based on resonance Raman spectroscgopy and sequence analysis. Biochimica et Biophysica Acta-Bioenergetics. 1797 (9), 1647-1656 (2010).
- Gildenhoff, N., Herz, J., Gundermann, K., Büchel, C., Wachtveitl, J. The excitation energy transfer in the trimeric fucoxanthin-chlorophyll protein from Cyclotella meneghiniana analyzed by polarized transient absorption spectroscopy. Chemical Physics. 373 (1), 104-109 (2010).
- Ramanan, C., et al. Exploring the mechanism(s) of energy dissipation in the light harvesting complex of the photosynthetic algae Cyclotella meneghiniana. Biochimica et Biophysica Acta-Bioenergetics. 1837 (9), 1507-1513 (2014).
- Haferkamp, S., Kirchhoff, H. Significance of molecular crowding in grana membranes of higher plants for light harvesting by photosystem II. Photosynthesis Research. 95 (2-3), 129-134 (2008).
- Wahadoszamen, M., et al. Stark fluorescence spectroscopy reveals two emitting sites in the dissipative state of FCP antennas. Biochimica et Biophysica Acta-Bioenergetics. 1837 (1), 193-200 (2014).
- Zhou, F., et al. Effect of monogalactosyldiacylglycerol on the interaction between photosystem II core complex and its antenna complexes in liposomes of thylakoid lipids. Photosynthesis Research. 99 (3), 185-193 (2009).
- Moya, I., Silvestri, M., Vallon, O., Cinque, G., Bassi, R. Time-resolved fluorescence analysis of the photosystem II antenna proteins in detergent micelles and liposomes. Biochemistry. 40 (42), 12552-12561 (2001).
- Natali, A., et al. Light-harvesting Complexes (LHCs) Cluster Spontaneously in Membrane Environment Leading to Shortening of Their Excited State Lifetimes. Journal of Biological Chemistry. 291 (32), 16730-16739 (2016).
- Gundermann, K., Büchel, C. Factors determining the fluorescence yield of fucoxanthin-chlorophyll complexes (FCP) involved in non-photochemical quenching in diatoms. Biochimica et Biophysica Acta-Bioenergetics. 1817 (7), 1044-1052 (2012).
- Ahmad, R. A., Dietzel, L. Relaxation of cellular K+ gradients by valinomycin induces diatoxanthin accumulation in Cyclotella meneghiniana cells and alters FCPa fluorescence yield in vitro. Physiologia Plantarum. , 171-180 (2017).
- Provasoli, L., McLaughlin, J. J. A., Droop, M. R. The development of artificial media for marine algae. Archiv für Mikrobiologie. 25 (4), 392-428 (1957).
- Jeffrey, S., Humphrey, G. New spectrophotometry equations for determining chlorophyll a, chlorophyll b, chlorophyll c-1 and chlorophyll c-2 in higher plants, algae and natural phytoplankton. Biochemie und Physiologie der Pflanzen. 167, 191-194 (1975).
- Beer, A., Gundermann, K., Beckmann, J., Büchel, C. Subunit Composition and Pigmentation of Fucoxanthin−Chlorophyll Proteins in Diatoms: Evidence for a Subunit Involved in Diadinoxanthin and Diatoxanthin Binding. Biochemistry. 45 (43), 13046-13053 (2006).
- Schägger, H., von Jagow, G. Tricine-sodium dodecyl sulfate-polyacrylamide gel electrophoresis for the separation of proteins in the range from 1 to 100 kDa. Analytical Biochemistry. 166 (2), 368-379 (1987).
- Büchel, C. Fucoxanthin-Chlorophyll Proteins in Diatoms: 18 and 19 kDa Subunits Assemble into Different Oligomeric States. Biochemistry. 42 (44), 13027-13034 (2003).
- Vieler, A., Wilhelm, C., Goss, R., Süß, R., Schiller, J. The lipid composition of the unicellular green alga Chlamydomonas reinhardtii and the diatom Cyclotella meneghiniana investigated by MALDI-TOF MS and TLC. Chemistry and Physics of Lipids. 150 (2), 143-155 (2007).
- Gundermann, K., Büchel, C. The fluorescence yield of the trimeric fucoxanthin-chlorophyll-protein FCPa in the diatom Cyclotella meneghiniana is dependent on the amount of bound diatoxanthin. Photosynthesis Research. 95 (2-3), 229-235 (2008).
- Miloslavina, Y., et al. Ultrafast fluorescence study on the location and mechanism of non-photochemical quenching in diatoms. Biochimica et Biophysica Acta-Bioenergetics. 1787 (10), 1189-1197 (2009).
- Grouneva, I., Jakob, T., Wilhelm, C., Goss, R. The regulation of xanthophyll cycle activity and of non-photochemical fluorescence quenching by two alternative electron flows in the diatoms Phaeodactylum tricornutum and Cyclotella meneghiniana. Biochimica et Biophysica Acta-Bioenergetics. 1787 (7), 929-938 (2009).
- Chukhutsina, V. U., Büchel, C., van Amerongen, H. Disentangling two non-photochemical quenching processes in Cyclotella meneghiniana by spectrally-resolved picosecond fluorescence at 77 K. Biochimica et Biophysica Acta-Bioenergetics. 1837 (6), 899-907 (2014).
- Ghazaryan, A., Akhtar, P., Garab, G., Lambrev, P. H., Büchel, C. Involvement of the Lhcx protein Fcp6 of the diatom Cyclotella meneghiniana in the macro-organisation and structural flexibility of thylakoid membranes. Biochimica Et Biophysica Acta-Bioenergetics. 1857 (9), 1373-1379 (2016).
- Darley, W. M. Biochemical composition. The biology of diatoms. 13, 198-223 (1977).
- Milsman, M. H. W., Schwendner, R. A., Weder, H. G. Preparation of large single bilayer liposomes by a fast and controlled dialysis. Biochimica Et Biophysica Acta. 512 (1), 147-155 (1978).
- Zumbuehl, O., Weder, H. G. Liposomes of controllable size in the range of 40 to 180 nm by defined dialysis of lipid-detergent-mixed micelles. Biochimica Et Biophysica Acta. 640 (1), 252-262 (1981).
- Verchere, A., Broutin, I., Picard, M. Photo-induced proton gradients for the in vitro investigation of bacterial efflux pumps. Scientific Reports. 2 (306), (2012).
- Chevallet, M., Luche, S., Rabilloud, T. Silver staining of proteins in polyacrylamide gels. Nature Protocols. 1 (4), 1852-1858 (2006).
Reimpresiones y Permisos
Solicitar permiso para reutilizar el texto o las figuras de este JoVE artículos
Solicitar permisoThis article has been published
Video Coming Soon
ACERCA DE JoVE
Copyright © 2025 MyJoVE Corporation. Todos los derechos reservados