
È necessario avere un abbonamento a JoVE per visualizzare questo. Accedi o inizia la tua prova gratuita.
Method Article
Isolando e incorporando luce-raccolta di antenne da diatomea Cyclotella Meneghiniana in liposomi con i lipidi Thylakoid
In questo articolo
Riepilogo
Qui, presentiamo un protocollo per isolare le proteine che legano fucoxantina clorofilla a/c (FCP) da diatomee e incorporarli nei liposomi con composizioni lipidiche naturali per studiare il trasferimento di energia di eccitazione su cambiamenti della composizione dello ione.
Abstract
Le prestazioni fotosintetiche delle piante, alghe e diatomee dipendono fortemente la regolazione veloce ed efficiente di raccolta luce ed energia dei processi di trasferimento nella membrana tilacoide dei cloroplasti. Trasferimento efficiente alla reazione fotosintetica centri anche per quanto riguarda foto-protezione dalla luce eccessiva e la luce raccolta dell'antenna di diatomee, le cosiddette proteine di legame di fucoxantina clorofilla a/c (FCP), sono necessari per l'assorbimento della luce. Il passaggio tra queste due funzioni è una questione di vecchia data di ricerca. Molti di questi studi sono stati effettuati con FCP in micelle detergente. Per gli studi di interazione, i detergenti sono stati rimossi, che ha condotto ad un'aggregazione aspecifica dei complessi FCP. In questo approccio, è difficile distinguere tra manufatti e fisiologicamente rilevanti dati. Quindi, più preziose informazioni sulla FCP e altra luce di membrana associata complessi di raccolta possono essere ottenuti dallo studio di interazioni proteina-proteina, il trasferimento di energia e altre caratteristiche spettroscopiche se sono incorporati nel loro ambiente nativo del lipido. Il vantaggio principale è che i liposomi hanno una dimensione definita e un rapporto definito dei lipidi/proteine mediante il quale viene controllata nella misura di FCP clustering. Ulteriormente, i cambiamenti della composizione ionica e pH che regolano la luce raccolta in vivo facilmente possono essere simulati. In confronto la membrana tilacoide, i liposomi sono più omogeneo e meno complessa, che lo rende più facile ottenere e comprendere dati spettroscopici. Il protocollo descrive la procedura di FCP isolamento e purificazione, preparazione dei liposomi e incorporazione di FCP in liposomi con composizione lipidica naturale. Risultati da un'applicazione tipica sono data e discusse.
Introduzione
Gli organismi fotosintetici come le diatomee devono affrontare condizioni di luce mutevoli e rispondere con meccanismi sofisticati di acclimatazione che sostengono ad alta efficienza fotosintetica e proteggono da danno foto-ossidativo causato dalla luce eccessiva. Un importante processo di luce-protettivo negli eucarioti fotosintetici è l'alta energia tempra (qE) di luce assorbita che si presenta come il principale contributo per la tempra non fotochimica (NPQ) sotto condizioni di luce lo stress1,2 ,3. Il luce complessi antenna raccolta (LHC) sono coinvolti nella regolazione delle vie di trasferimento di energia di eccitazione. In risposta a luce alta indotta basso pH nel lume cloroplasto, gli interruttori del sistema antenna dalla luce raccolta allo stato dissetante. Questo stato che tende a dissipare energia protegge fotosistema (PS) e altri complessi nella membrana tilacoide da foto-ossidazione. Negli eucarioti fotosintetici, la qE solitamente è indotta da due fattori1,2,3. Un fattore è la luce specializzata raccolta della proteina che risponde al basso pH. La proteina PsbS induce il qE in più piante4. LhcSRs5, modulata da attività PsbS, indurre il qE alghe verdi6. Le diatomee possiedono proteine Lhcx-like che strutturalmente correlata alla LHCSRs7,8,9,10.
Il secondo fattore di qE è il ciclo della xantofilla dove i carotenoidi dell'antenna sono convertiti in un formato di foto-protettivo dal de-epossidazione e ripristinati da epossidazione. Nelle piante e alghe verdi, violaxantina viene convertito in zeaxantina. A diatomee, diadinoxanthin viene convertito in diatoxanthin, che quindi correla con il grado di NPQ11. La luce della diatomea raccolta antenna possiede alcune peculiarità, anche se è evolutiva legate alla pianta e LHCs d'alghe. L'interruttore da luce raccolta foto-protezione è estremamente veloce e la capacità NPQ è superiore rispetto a piante12. Questo potrebbe essere una ragione perché le diatomee sono molto successo in diverse nicchie ecologiche, in modo che essi sono responsabili fino al 45% della produzione primaria netta oceanico13. Di conseguenza, della diatomea luce sistemi di raccolta sono un interessante oggetto di ricerca di fotosintesi.
Diatomee, come le specie centric Cyclotella meneghiniana, possiedono luce intrinseca thylakoid prende i pigmenti hanno sistemi di raccolta associare - fucoxantina, clorofilla (chl) a e c, quindi FCP. luce raccolta proteine, quali FCPs, sono incorporato nel sistema della membrana tilacoide formati da diversi strati di membrana. Diatomee formano le fasce dei tre tilacoidi. Questo complesso situazione rende difficile per loro studiare a livello molecolare nella membrana tilacoide. Inoltre, molti componenti contribuiscono alla regolazione della luce raccolta (Vedi sopra). Di conseguenza, in molti approcci, i complessi sono stati isolati dalla membrana utilizzando un detergente delicato, come n-dodecil-β-D-maltopyranoside (β-DDM), che solubilizzano la membrana, ma mantenere intatti i complessi FCP. Molti studi spettroscopici sono stati eseguiti utilizzando solubilizzate FCP per indagare intramolecolare energia trasferimento14,15,16,17. Tuttavia, questo approccio ex era limitato poiché il regolamento del trasferimento di energia ha bisogno eccitoniche interazione con altri complessi antenna o fotosistema. Quindi, questi generi di studi non possono essere effettuati con solubilizzate complessi perché l'interazione fra i complessi è perso.
Una caratteristica importante nella regolazione dell'antenna è il "affollamento molecolare" del fotosistema nella membrana tilacoide18e dell'antenna. Precedentemente, è stato realizzato un approccio semplice per simulare questo effetto in vitro. Il detersivo è stato rimosso, che porta all'aggregazione casuale di complessi antenna. Anche se alcuni dati ragionevoli è stati ottenuti da questo approccio17,19, la rimozione del detersivo non riflette la situazione in vivo e presenta alcune limitazioni, poiché i complessi non interagiscono nel loro terziari regolari struttura.
L'uso dei liposomi supera molte delle limitazioni ex. La struttura terziaria è ancora completamente intatta. La membrana del liposoma fornisce un ambiente quasi nativo per i complessi dell'antenna. La membrana separa l'interno del liposoma dall'ambiente esterno. Con questi mezzi, liposomi forniscono due scomparti di reazione per studi di gradienti ionici e pH pure per quanto riguarda i processi di trasporto. Ulteriormente, i parametri del sistema sperimentale possono essere controllati più facilmente per gli studi nella membrana tilacoide. Liposomi già sono stati indicati per essere un ottimo strumento per lo studio di complessi fotosintetici. Degli obiettivi principali in passato era sulla pianta LHC dove l'effetto della composizione lipidica alterato è stato testato su LHC II20. In altri approcci, interazione proteina-proteina tra diversi LHC II sono stati studiati21. Inoltre, alcuni studi in alghe verdi sono stati effettuati che descrivono il clustering spontanea tra LHC22. Considerando l'importanza di diatomee per gli ecosistemi acquatici, relativamente pochi studi sono stati effettuati con complessi antenna di diatomee. Due studi hanno studiato i complessi antenna della centrica Cyclotella meneghiniana, dove il clustering della FCP antenna23 e reattività di FCP a gradienti elettrochimici24 sono stati indicati. Così, i liposomi sono un ottimo strumento per lo studio della diatomea antenne e loro interazione e regolazione in condizioni quasi native. I liposomi sono versatili da molte condizioni come composizione lipidica, liposoma dimensioni, densità di proteina e la fase acquosa circostante può essere controllata. Inoltre, il metodo richiede basse quantità di campioni. Il sistema sperimentale è robusto e altamente riproducibili. La compartimentazione dei liposomi permette lo studio di pH e gradienti ionici, che sono importanti fattori nella regolazione dei complessi antenna.
Qui, descriviamo l'isolamento di complessi antenna FCP da c. meneghiniana e la loro incorporazione in liposomi con composizione lipidica naturale thylakoid. Inoltre, forniamo dati esemplari per la caratterizzazione spettroscopica di FCP solubilizzate e confrontarli con FCP in liposomi. Il metodo riassume conoscenze e protocolli standardizzati ottenuti dai miglioramenti di Gundermann e Büchel 201223, Natali et al 201622e Ahmad e Dietzel 201724.

Figura 1: rappresentazione schematica del flusso di lavoro. (1) fa riferimento al paragrafo 1 che descrive la crescita delle cellule, rottura e tilacoidale isolamento con seguito di FCP separazione su gradienti di densità di saccarosio; M. c. -Cyclotella meneghiniana cellule. (2) preparazione della miscela di lipidi naturali thylakoid (MGDG, DGDG e SQDG) descritto nel paragrafo 2 e creazione di micelle lipidiche-detergente con octylglycoside (OG). Una dimensione definita del lipido-micella avviene mediante estrusione utilizzando membrane di un diametro dei pori definiti. FCP e lipido-micelle sono unificate presso un lipide predefinito: rapporto proteine e i detergenti OG e β-DDM vengono rimossi tramite controllata dialisi formando FCP proteoliposomi. Clicca qui per visualizzare una versione più grande di questa figura.
{kind=link}
Protocollo
Nota: Fotosintetici complessi quali FCPs sono altamente vulnerabili alla luce e al calore. Sempre lavorare sul ghiaccio e sotto una luce molto fioca.
1. isolamento di FCP dalle cellule
-
Thylakoid isolamento dalle cellule di c. meneghiniana
- C. meneghiniana in cinque palloni 500 mL che ciascuno riempito con 300 mL di ASP-medio23,25 e 50 milioni di cellule si sviluppano. Collegare le beute con un tappo di cotone e permettono alle cellule di crescere per la fase di crescita esponenziale per circa una settimana su un agitatore a 120 giri/min con un 16 h luce e fase scura di 8h alle 40 µmol photons/(m²s) bianco luce e una temperatura compresa tra 15-18 ° C. Verifica che il numero di cellulare è tra 1,5 milioni di cellule/mL con un alloggiamento del contatore di cella.
- Centrifugare le cellule a 4.000 x g in un rotore preraffreddato con fiale di 500 mL centrifuga (4 ° C) per 15 min in una centrifuga ad alta velocità. Pipettando, risospendere il pellet cellulare in 12 mL di tampone di omogeneizzazione (HB, tabella 1).
- Trasferire la sospensione in una provetta di plastica singolo 50 mL. Conservare i campioni a-80 ° C o passare al punto 1.1.3.
- Pre-raffreddare il laminatoio del branello e attrezzature. Riempire il becher 50ml del mulino perlina al 75% con una miscela di microsfere di vetro e aggiungere la sospensione cellulare. Per distruzione cellulare, usare 7x45 s impulsi a piena velocità con 30 s di raffreddamento tra ogni impulso. Prendere 20 µ l di celle interrotte per controlli di qualità nel passaggio 1.1.6.
- Filtrare le celle interrotte sopra un imbuto di vetro e lavare versando l'HB sopra le perline di vetro fino a quando non appaiono chiare. Piscina la frazione di lavare con il filtrato. Tenere il volume finale inferiore a 150 mL.
- Centrifugare il campione per 15 min a 140 x g usando tre provette di plastica da 50 mL a pellet i detriti cellulari. Attentamente trasferire il surnatante a 20 mL in policarbonato ultracentrifugazione fiale e scartare il pellet.
- Riempire i flaconi con HB, equilibrare il peso e centrifugare in un rotore adatto per 1 h a 300.000 x g e a 4 ° C per appallottolare le membrane tilacoidali.
- Utilizzare il tempo di centrifugazione per controllare la proporzione di cellule perturbate da microscopia a 400 ingrandimenti con i 20 µ l di campione preso a 1.1.3. Calcolare il rapporto tra cloroplasto gratis e cloroplasto contenente frustules (gusci di silice).
Nota: Pareti della cellula della diatomea sono costituite da silice, che sono visibile come altamente diffracting sostanza nel microscopio. Cloroplasti che si verificano punti verdi come avrebbe dovuto essere rilasciati dalle cellule se ha funzionato la distruzione cellulare. - Risospendere il pellet di membrana con come piccolo tampone di lavaggio come possibile (0,5-1 mL) utilizzando il pennello di un pittore piccolo. Riempire i flaconi di ultracentrifugazione in policarbonato con lavaggio tampone (tabella 1), equilibrare il loro peso e centrifugare per 20 min a 200.000 x g e a 4 ° C.
- Risospendere le membrane lavate con pennello di un pittore. Aggiungere tampone di lavaggio solo se necessario per mantenere la concentrazione di thylakoid più in alto possibile. Tutti i tilacoidi nel flacone di un campione (15 mL) della piscina.
- Pre-diluire i campioni utilizzando 10 µ l di campione con 90 µ l di acetone 100%. E centrifugare a 12.000 x g per 5 min a pellet proteina precipitata. Prendere 10 µ l della prediluizione e mescolare con 990 µ l di acetone 90%.
- Misurare l'assorbanza (ABS) di clorofilla a e c a 664 nm e 630 nm in 90% di acetone. Sottrarre l' ABS750nm da entrambi i valori. Determinare il contenuto di clorofilla totale utilizzando la seguente formula:24
1)
2)
- Misurare l'assorbanza (ABS) di clorofilla a e c a 664 nm e 630 nm in 90% di acetone. Sottrarre l' ABS750nm da entrambi i valori. Determinare il contenuto di clorofilla totale utilizzando la seguente formula:24
- Aliquotare tilacoidi in porzioni di 0,5 mg di clorofilla totale in un di provette da 1,5 mL, congelarli in azoto liquido e conservarli a-80 ° C fino all'utilizzo ulteriore.
-
Separazione e concentrazione di complessi FCP
- Preparare una soluzione di gradiente di saccarosio e riempire le provette ultracentrifuga fino alla parte superiore meno il volume di carico (300 – 500 µ l). Congelare le provette a-20 ° C fino a quando sono completamente congelati. Lasciare le provette per lo scongelamento a + 4 ° C, che dura 3-4 ore per un tubo da 17 mL.
- Ripetere il ciclo gelo-disgelo due volte per affinare la sfumatura per una migliore risoluzione.
- Utilizzare i campioni ottenuti in 1.1.9 pari a 0,5 mg di clorofilla e regolare con buffer B1 (tabella 1) ad un volume finale di 2 mL. Per solubilizzazione, aggiungere n-dodecil-β-D-maltopyranoside (b-DDM) ad una concentrazione finale di 20 mM.
- Capovolgere la provetta 3 volte e metterlo in ghiaccio per 20 minuti con agitazione delicata per evitare di schiuma. Centrifugare per 5 min, 12.000 x g in una centrifuga pre-raffreddata tabella superiore a + 4 ° C.
- Caricare il surnatante sulla sfumatura. Non caricare più di 125 µ g di clorofilla totale al gradiente se 17 mL fiale vengono utilizzati. Centrifuga per 22 h a 100.000 x g e + 4 ° C.
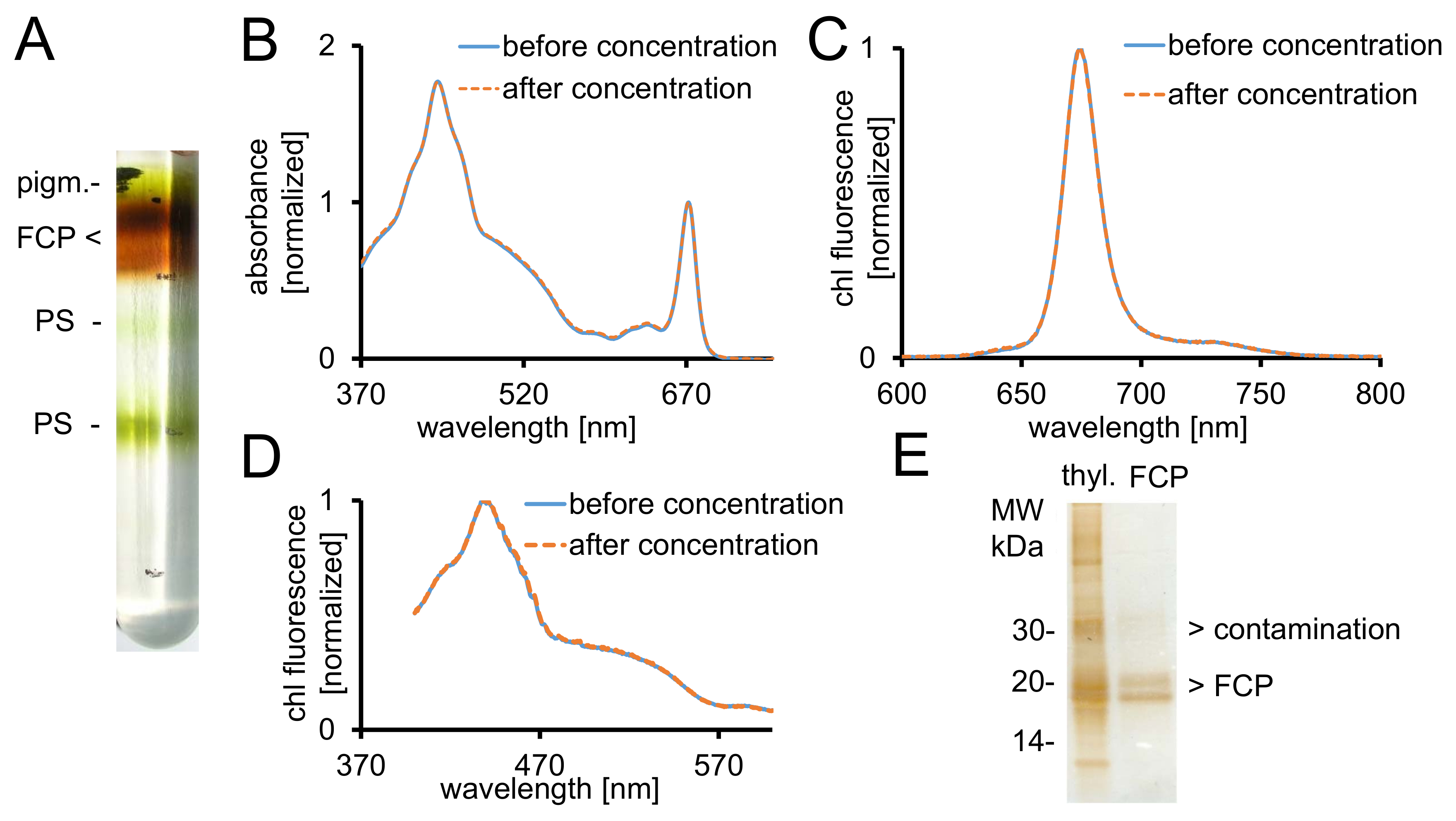
- Recuperare le frazioni FCP marrone desiderate dal gradiente usando una siringa (Figura 2A). Prendere un 5 µ l aliquota e diluirla con 995 µ l di B1a.
- Misurare lo spettro di assorbanza (ABS) tra 370-750 nm in uno spettrofotometro UV-VIS. Utilizzare cuvette semimicro vetro ottico.
- Aprire il software di gestione di spettri, andare in modalità spettro e registrare una previsione da 370 a 750 nm seguita da misure campione. Utilizzare le seguenti impostazioni: scansione velocità, 200 nm/min; dati passo, 0.5 nm; risposta, media.
- Misurare il volume del campione recuperato con una micropipetta. Lavare i complessi FCP aggiungendo due volte il volume recuperato con buffer B1 (tabella 1). Concentrare in un concentratore di membrana con un taglio a 1.000 x g e + 4 ° C per un ABS672nm di almeno 20 kDa 30.
Nota: La concentrazione di b-DDM potrebbe aumentare a causa di arricchimento micella nel vano del campione. Questo potrebbe portare ad ulteriore solubilizzazione dei complessi FCP! Evitare eccessiva solubilizzazione aggiungendo detergente buffer liberi B1 se ulteriori fasi di lavaggio sono tenuti a rimuovere residuo saccarosio. - Prendere un 20 µ l aliquota per i controlli. Shock congelare i campioni in azoto liquido e conservarli a-80 ° C.
- Preparare una soluzione di gradiente di saccarosio e riempire le provette ultracentrifuga fino alla parte superiore meno il volume di carico (300 – 500 µ l). Congelare le provette a-20 ° C fino a quando sono completamente congelati. Lasciare le provette per lo scongelamento a + 4 ° C, che dura 3-4 ore per un tubo da 17 mL.

Figura 2: purificazione di FCP, controlli spettroscopici e purezza check. (A) l'aspetto tipico di un gradiente di densità di saccarosio dopo centrifugazione durante la notte. Tutte le bande marroni contengono il pool FCP costituito FCPa e FCPb. pigm. - unbound pigmenti, PS - fotosistema spettri di assorbanza (B) di FCP prima (linea blu) e dopo concentrazione (linea tratteggiata arancione) utilizzando dispositivi centrifughi con 30 kDa cutoff . In particolare, i carotenoidi sono soggetti a perdita da FCP, che si tradurrebbe in assorbanza inferiore nella regione tra 500-550 nm. Grafici sono normalizzati alla clorofilla Qy massima a ~ 670 nm. (C) spettri di emissione di clorofilla a con eccitazione di chl c (465 nm) per il test di trasferimento di energia di eccitazione funzionale. Se l'energia Trasferisci c chl chl un è ostacolato, una band di fluorescenza ulteriori ~ 640 nm (chl c) si sarebbe verificato. Grafici sono normalizzati per l'emissione massima. (D) spettri di eccitazione registrata a 675 nm (chl una fluorescenza massima) per il test di trasferimento di energia alla chl un da tutti i pigmenti assorbenti tra 370 nm e 600 nm. Se l'energia si trasferisce a chl un è meno efficiente, la resa di fluorescenza farebbe diminuire soprattutto tra 550 e 465 nm. I grafici sono normalizzati al massimo circa 440 nm. Gli spettri in (B), (C) e (D) sono quasi identici se la concentrazione ha funzionato bene. (E) si verifica per la purezza della FCP isolata utilizzando un gel di Tris-tricine28. FCPa e FCPb hanno subunità tra 18-19 kDa. Tutte le proteine visibile argento-macchiate più grandi di 20 kDa sono contaminanti. Thyl. -Tilacoidi Clicca qui per visualizzare una versione più grande di questa figura.
{kind=link}
-
Spettroscopici e gel a base di controlli
- Annotare l'assorbanza tra 370-750 nm di 5 µ l di FCP in 995 µ l di B1a dopo passo 1.2.5. Confrontarlo con lo spettro ottenuto nel passaggio 1.2.4. Utilizzare la stessa strumentazione e impostazioni come descritto al punto 1.2.4.
- Esportare i dati come CSV e importare i dati in un foglio di calcolo. Due spettri al massimo di chl di normalizzare nella banday Q di circa 672 nm come raffigurato nella Figura 2B.
- Calcolare il fattore di diluizione dei campioni dal punto 1.2.4 e 1.3.1 dividendo l' ABS misurato672nm dalla desiderata ABS672nm di 0,03. Diluire i campioni di conseguenza con B1a e trasferirli cuvette di fluorescenza di vetro speciale.
- Registrare chl uno spettro di emissione di fluorescenza con una Spettrofluorimetro per rivelare il trasferimento di energia luce intatta da chl c alla chl un (Figura 2).
- Utilizzare il software di spettrometro e andare per la modalità di misurazione dello spettro con le seguenti impostazioni: modalità, emissione; a taglio larghezza eccitazione e emissione, 3 nm; sensibilità, medio; velocità di scansione, 100 nm/min; dati passo, 0.5 nm; lunghezza d'onda di eccitazione, 465 nm; emissione, 600-800 nm. Eseguire autozero e misura.
- Registrare spettri di eccitazione con lo stesso campione e attrezzature per rivelare il trasferimento di energia intatto da tutti i pigmenti alla chl (Figura 2D). Modificare le impostazioni per: modalità, eccitazione; lunghezza d'onda di emissione, 675 nm; eccitazione, 370-600 nm. Registrare gli spettri.
- Correggere con uno spettro di rodamina nello stesso intervallo per proprietà spettrali della lampada – cf. istruzioni nel manuale dell'utente.
- Campioni di mix FCP corrispondenti a 1 µ g di chl un con 10 µ l di tampone di caricamento SDS. Incubare per 10 minuti a 25 ° C. Centrifugare per 5 min a 12000 x g in una centrifuga di piano d'appoggio.
- Caricare il surnatante su un gel di Tris-tricine28. Separarlo per 2 ore a 150 V e argento-macchia dopo separazione40.
Nota: Le subunità FCP separano in due bande proteiche prominente tra 18-19 kDa, che sono costituenti di FCPa e FCPb29 (Figura 2E).
- Caricare il surnatante su un gel di Tris-tricine28. Separarlo per 2 ore a 150 V e argento-macchia dopo separazione40.
- Annotare l'assorbanza tra 370-750 nm di 5 µ l di FCP in 995 µ l di B1a dopo passo 1.2.5. Confrontarlo con lo spettro ottenuto nel passaggio 1.2.4. Utilizzare la stessa strumentazione e impostazioni come descritto al punto 1.2.4.
2. preparazione dei liposomi e incorporazione di FCP
-
Preparazione della miscela del lipido e detergente micelle lipidiche
Nota: i lipidi sono suscettibili di combinato con circostanze ossidative di temperature calde. Cercare di mantenere i lipidi refrigerati e sotto un'atmosfera di2 N.- Calcolare i rapporti del lipido thylakoid desiderata per c. meneghiniana secondo Vieler et al. 200730. Fare riferimento all'esempio fornito nella Supplemental tabella 1. Preparare il lipido stock soluzioni consigliate dal produttore in un contenitore di prova solvente.
- Pipettare la quantità desiderata di lipidi in una provetta di reazione di 2 mL ed evaporare il cloroformio utilizzando un flusso di azoto delicato e cercare di diffondere i lipidi su tutta la zona della base del tubo. Lasciate il flusso di N2 fino a quando tutti i solvente è evaporato.
- Solubilizzare la miscela di lipidi in 29 µ l di soluzione di n-ottile β-D-glucopyranoside (OG) a 4 ° C per 4 h. Incubare la miscela di lipidi per 10 min a 30 ° C. Incubare i lipidi in un bagno sonicatore per 3 x 3 min a 25 ° C, interrotto da 30 s sul ghiaccio.
- Aggiungere 221 µ l di Tampone tricina e 250 µ l di tampone di dialisi x 4.
- Utilizzare un estrusore con membrane di policarbonato 0,1 µm per un diametro definito liposoma di 50-70 nm. Montare l'estrusore con il supporto della membrana e filtro. Evitare le bolle d'aria e serrare accuratamente l'assembly.
- Riempire una siringa con 4 x dialisi tampone e pre-bagnato l'estrusore, fino a quando non ci sono bolle possono essere visto nella seconda siringa.
- Applicare il lipido-detergente-micelle all'estrusore e premere la soluzione da una siringa a altra avanti e indietro. Ripetere questo passaggio 5 volte fino a quando la soluzione appare omogenea.
Nota: Questa soluzione può essere conservata a 4 ° C per diversi giorni. Non congelare!
-
Incorporazione di FCP complessi e rimozione dei detergenti e aggregati
Nota: In questo esempio usiamo un lipido/chl un rapporto di 12:1, che corrisponde a un rapporto lipidi/proteine circa 100: 1.- Aggiungere FCP uguale a 20 µ g di chl un in un volume totale di 500 µ l di tampone di B1a a 250 μL dell'estruso del lipido-micelle e 250 µ l di tampone di dialisi: 4x. Incubare i campioni per 3 x 3 min a 25 ° C in un thermomixer a 1.500-3.000 giri/min interrotto da una pausa di 30 s sul ghiaccio.
- Tagliare il coperchio del tubo di reazione 1,5 mL quattro appena sotto la parte superiore dà un anello che si inserisce ancora sul coperchio. Preparare pezzi di 1,5 x 1,5 cm della membrana di dialisi e lavarli in 20 mL di tampone dialisi 1x
- Riempire 250 µ l di campione per ogni coperchio. Appoggiare delicatamente, la membrana sul coperchio in modo che il vano è completamente riempito con il campione e si verificano senza bolle d'aria. Stringere l'anello di tubo di reazione sull'Assemblea al fine di avere un vano chiuso.
- Dializzare i campioni in 50 mL di tampone di dialisi durante la notte: 1x (12-16 h) sul ghiaccio su un agitatore di barilatura. Sostituire il buffer di dialisi peritoneale con una fresca e aggiungere 7 mg di adsorbente perline per rimuovere i rimanenti detergenti per almeno 6 ore.
- Sostituire di nuovo il buffer di dialisi e Dializzare per un altro 12 h. recuperare i liposomi forando la membrana di dialisi con una punta di una micropipetta 200 µ l e aspirare tutti i liposomi dal coperchio del tubo di reazione.
- Passaggio facoltativo: se elevata purezza (> 95%) è necessario. Preparare un gradiente di densità discontinua in 17 mL fiale ultracentrifugazione con passaggi contenente 6%, 10%, 15% e 20% copolimero epicloridrina di saccarosio nel buffer di dialisi. Caricare i liposomi sulla parte superiore e ultracentrifuga a 100.000 x g per 4 h in un rotore oscillante.
- Recuperare la banda superiore marrone con una siringa, diluire il campione 1:5 con DP e procedere al passaggio successivo.
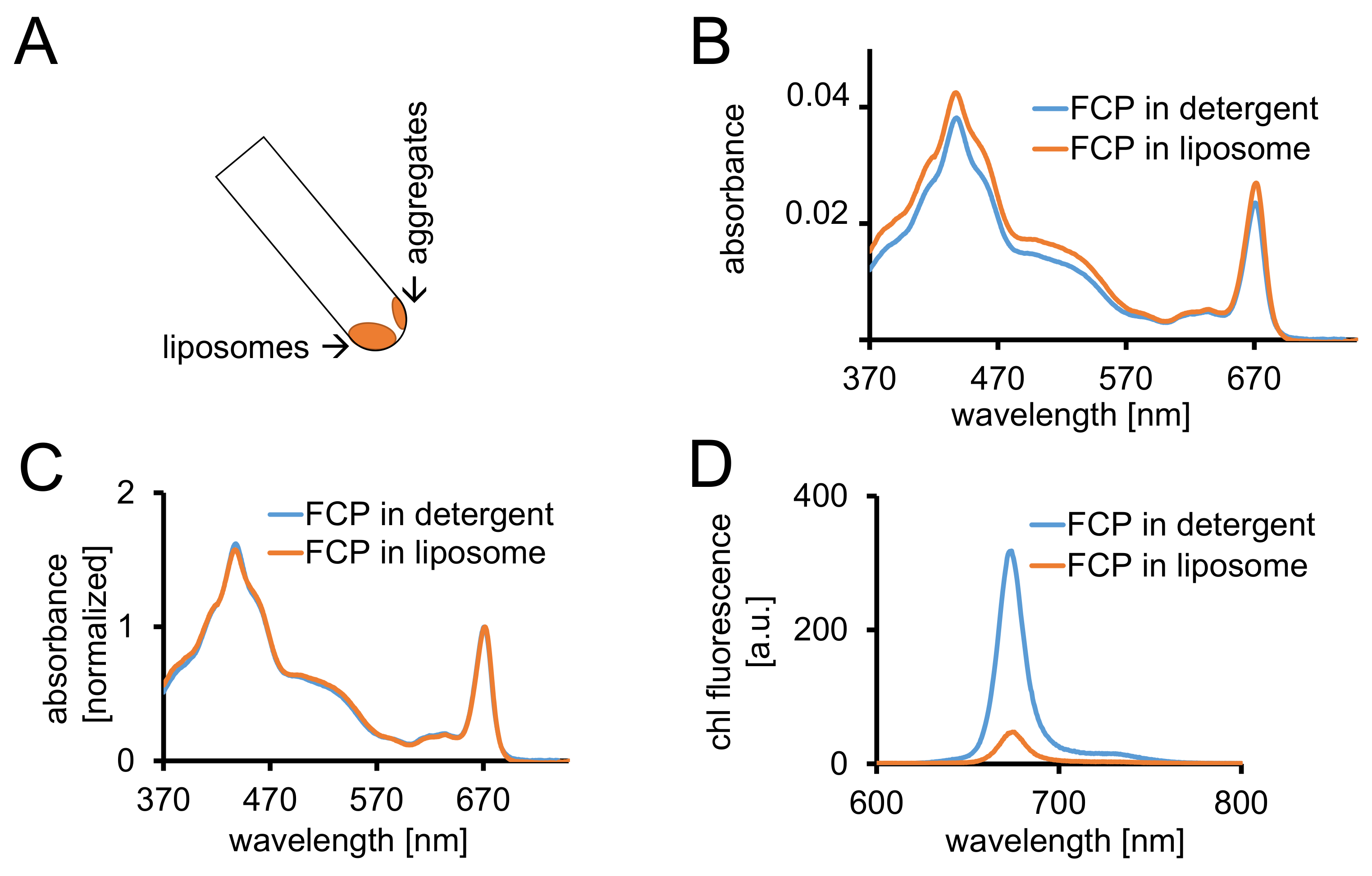
- Centrifugare i liposomi FCP in almeno 2 mL di 1x buffer di dialisi per 1,5 h a 100.000 x g e a 4 ° C. Recuperare i liposomi ruotando il tubo da centrifuga con un angolo di 45°. Consentire i liposomi spostare verso il basso per 1 min (Figura 3A).
- Recuperare i liposomi FCP in un volume finale di 25-50 µ l. evitare di disturbare il precipitato.
-
Controlli 1: assorbanza, spettroscopia di fluorescenza
- Aggiungere un'aliquota di 3 µ l di FCP-liposomi ad un volume finale di 1 mL di 1x buffer di dialisi e centrifugare per 5 min a 12.000 x g.
- Annotare l'assorbanza tra 370 e 750 nm di FCP-liposomi con la stessa attrezzatura come descritto in 1.2.4. Normalizzare lo spettro al massimo della regioney Q della chl un (670-680 nm) e confrontarlo con lo spettro normalizzato di solubilizzate FCP (Figura 3).
- Preparare FCP-liposomi in 1 mL di DP con un'assorbanza (ABS) = 0.03 per quanto riguarda il massimo tra 670-680 nm. Regolare la FCP solubilizzate in detergente dal punto 1.2.6 ABS stesso diluendo con B1a.
- Registrare spettri di assorbanza di entrambi i campioni come descritto in 1.2.4. Registrare chl uno spettro di emissione di fluorescenza di entrambi i campioni come descritto in 1.3.3.
Nota: Il rendimento di fluorescenza è diminuito nel campione FCP-liposoma (Figura 3D e cfr discussione.)
- Aggiungere un'aliquota di 3 µ l di FCP-liposomi ad un volume finale di 1 mL di 1x buffer di dialisi e centrifugare per 5 min a 12.000 x g.

Figura 3: isolamento di FCP proteoliposomi seguita da controlli spettroscopici e imaging confocale. (A) recupero di liposomi FCP dopo la centrifugazione. Girare il tubo di centrifugazione a 45° e attendere circa 1 min - liposomi si sposterà verso il basso mentre la FCP aggregati che non sono incorporati in liposomi stick per la parete del tubo. (B) confronto degli spettri di assorbanza di FCP solubilizzate in detersivo (blu) e FCP in liposomi (arancione) (C) gli stessi spettri come in (B) normalizzato alla chl un massimo nella regione rossa (~ 670 nm - Qy picco); FCP solubilizzate in detersivo (blu) e FCP in liposomi (arancione). Potenzialmente, ci potrebbe essere una perdita di pigmento principalmente dei carotenoidi visibile nella regione dei 500-550 nm. Il clustering di FCP nei liposomi può portare a un allargamento del picco e un leggero spostamento della chl un massimo (~ 670 nm) al rosso. (D) spettri di emissione di FCP solubilizzate in detersivo e FCP in liposomi. Clustering di FCP nel liposoma migliora energici interazioni dei complessi FCP che abbassa il rendimento di fluorescenza (curva arancia) e i massimi di emissione si sposta leggermente al rosso. Clicca qui per visualizzare una versione più grande di questa figura.
{kind=link}
Risultati
Il protocollo descrive l'isolamento della frazione totale di FCP da Cyclotella meneghiniana e incorporazione in liposomi con composizione lipidica nativa. L'isolamento di thylakoid è altamente riproducibile, ma la resa di thylakoid potrebbe cambiare. Il risultato è accettabile se più del 50% di tutti i pigmenti sono recuperati nel passaggio 1.1.4. Più dell'80% è ottimale.
La solubilizzazione dei tilacoidi è un pas...
Discussione
Liposomi FCP con composizione lipidica naturale forniscono uno strumento pratico, semplice e riproducibile per indagare le proprietà spettroscopiche in vitro. L'ambiente del lipido in liposomi FCP ricorda la situazione all'interno della membrana tilacoide, dando luogo a risultati sperimentali che sono più vicini alle condizioni naturali.
Ci sono diversi vantaggi di usando c. meneghiniana come sistema modello per antenna FCP. Si sviluppa relativamente veloce ed è più robus...
Divulgazioni
Gli autori non hanno nulla a rivelare.
Riconoscimenti
Ringraziamo la Rana Adeel Ahmad per assistenza nella purificazione di FCP. Prof. ssa Claudia Büchel è riconosciuto per le utili discussioni e leggere il manoscritto. Questo lavoro è stato supportato da German Research Foundation LD (DI1956-1/1) e la fondazione Humboldt per una borsa di Feodor-Lynen di LD
Materiali
| Name | Company | Catalog Number | Comments |
| 500 ml centrifuge vials | |||
| high speed centrifuge | Heraeus | ||
| Bead Mill VI 2 | Edmund-Bühler (edmund-buehler.de) | newer version: Vibrogen-Zellmühle Vl 6 | |
| Silibeads S 400 µm | Sigmund-Lindner.com | 5223-7 | |
| Silibeads S 1,-1,3 mm | Sigmund-Lindner.com | 4504 | |
| VitraPOR filter funnel - por1 | ROBU GmbH | 21121 | |
| polycarbonate ultracentrifuagtion vials (30 mL) for T-865 | Beranek Laborgeräte (Laborgeraete-beranek.de) | 314348 | |
| Ultracentrifuge Discovery 90SE | Sorvall | n.a. | |
| rotor T 865 | ThermoFisher Scientific (thermofisher.com) | 51411 | |
| Neubauer Cell Counter Chamber (improved) | Carl Roth Laborbedarf (Carlroth.com) | T729.1 | |
| Zeiss Mikroskop Primostar (7) | Optik-Pro (optik-pro.de) | 51428 | |
| optical glass cuvettes (6040-OG) | Hellma Analytics (hellma-analytics.com) | "6040-10-10" | |
| V-630 UV-VIS Spectrophotometer (incl. software) | Jasco (jasco.de) | V-630 | |
| n-Dodecyl-β-D-Maltopyranoside | ANATRACE (anatrace.com) | D310LA | |
| Ultra-Clear tubes 17 ml for AH629 | Beranek Laborgeräte (Laborgeraete-beranek.de) | 344061 | |
| rotor AH629-17-mL | ThermoFisher Scientific (thermofisher.com) | 54285 | |
| Membrane concentrator_Centriprep 30 kDa cutoff | Millipore (merckmillipore.com) | 4307 | |
| Biometra Minigel-Twin | Analytik Jena AG (analytik-jena.de) | 846-010-100 | |
| Silver Stain Plus Kit | Bio-Rad (bio-rad.com) | 1610449 | |
| libre office spread sheet | The document foundation | https://de.libreoffice.org/download/libreoffice-still/ | |
| special glass cuvettes for fluorescence (101-0S) | Hellma Analytics (hellma-analytics.com) | 101-10-20 | |
| Spectrofluorometer FP-6500 (incl. Software) | Jasco (jasco.de) | FP-6500 | |
| SDS-loading buffer Roti-Load | ROTH (carlroth.com) | K929.1 | |
| n-octyl β-D-glucopyranoside | ANATRACE (anatrace.com) | O311 | |
| Monogalactosyl Diaclyglycerol (MGDG) | Larodan AB (larodan.com) | 59-1300 | make stock solution in chloroform |
| Digalactosyl Diacylglycerol (DGDG) | Larodan AB (larodan.com) | 59-1310 | make stock solution in chloroform |
| Sulphoquinovosyl Diacylglycerol (SQDG) | Larodan AB (larodan.com) | 59-1230 | make stock solution in chloroform |
| L-alpha-Phosphatidylglycerol (PG) | Larodan AB (larodan.com) | 37-0150 | make stock solution in chloroform |
| L-α-Phosphatidylcholine | Sigma-Aldrich (sigmaaldrich.com) | P3782 SIGMA | make stock solution in chloroform |
| sonicator bath S-50TH | Sonicor (getmedonline.com | SONICOR-S-50TH | |
| mini-Extruder | Avanti Polar Lipids (Avanti.com) | 610000 | |
| Nuleopore polycarbonate membrane | Avanti Polar Lipids (Avanti.com) | 610005 | |
| dialysis membrane Visking 14 kDa cutoff | ROTH (carlroth.com) | 0653.1 | boil in destilled water before use |
| Biobeads SM2 Adsorbent | Biorad (Bio-rad.com) | 152-3920 | |
| sucrose epichlorhydrin copolymer - Ficoll 400 | Sigma-Aldrich (sigmaaldrich.com) | F4375 | |
| Polycarbonate ultracentrifuagtion vials (2.7 mL) for TFT 80.4 | Beranek Laborgeräte (Laborgeraete-beranek.de) | 252150 | |
| rotor TFT 80.4 | Millipore (merckmillipore.com) | 54356 | |
| material listed in order of appearance | For specific safety instructions please refer to material safety sheets and repective manuals. Standard lab material and substances are not listed. |
Riferimenti
- Eberhard, S., Finazzi, G., Wollman, F. A. The Dynamics of Photosynthesis. Annual Review of Genetics. 42, 463-515 (2008).
- Li, Z. R., Wakao, S., Fischer, B. B., Niyogi, K. K. Sensing and Responding to Excess Light. Annual Review of Plant Biology. 60, 239-260 (2009).
- Niyogi, K. K., Truong, T. B. Evolution of flexible non-photochemical quenching mechanisms that regulate light harvesting in oxygenic photosynthesis. Current Opinion in Plant Biology. 16 (3), 307-314 (2013).
- Li, X. -. P., et al. A pigment-binding protein essential for regulation of photosynthetic light harvesting. Nature. 403 (6768), 391-395 (2000).
- Peers, G., et al. An ancient light-harvesting protein is critical for the regulation of algal photosynthesis. Nature. 462 (7272), 518-521 (2009).
- Correa-Galvis, V., et al. Photosystem II Subunit PsbS Is Involved in the Induction of LHCSR Protein-dependent Energy Dissipation in Chlamydomonas reinhardtii. The Journal of biological chemistry. 291 (33), 17478-17487 (2016).
- Bailleul, B., et al. An atypical member of the light-harvesting complex stress-related protein family modulates diatom responses to light. Proceedings of the National Academy of Sciences of the United States of America. 107 (42), 18214-18219 (2010).
- Taddei, L., et al. Multisignal control of expression of the LHCX protein family in the marine diatom Phaeodactylum tricornutum. Journal of experimental botany. 67 (13), 3939-3951 (2016).
- Lepetit, B., et al. The diatom Phaeodactylum tricornutum adjusts nonphotochemical fluorescence quenching capacity in response to dynamic light via fine-tuned Lhcx and xanthophyll cycle pigment synthesis. New Phytologist. 214 (1), 205-218 (2017).
- Büchel, C. Evolution and function of light harvesting proteins. Journal of Plant Physiology. 172, 62-75 (2015).
- Lavaud, J., Rousseau, B., van Gorkom, H. J., Etienne, A. -. L. Influence of the Diadinoxanthin Pool Size on Photoprotection in the Marine Planktonic Diatom Phaeodactylum tricornutum. Plant Physiology. 129 (3), 1398-1406 (2002).
- Ruban, A., et al. The super-excess energy dissipation in diatom algae: comparative analysis with higher plants. Photosynthesis Research. 82 (2), 165-175 (2004).
- Mann, D. G. The species concept in diatoms. Phycologia. 38 (6), 437-495 (1999).
- Papagiannakis, E., van Stokkum, I. H. M., Fey, H., Büchel, C., van Grondelle, R. Spectroscopic Characterization of the Excitation Energy Transfer in the Fucoxanthin-Chlorophyll Protein of Diatoms. Photosynthesis Research. 86 (1-2), 241-250 (2005).
- Premvardhan, L., Robert, B., Beer, A., Büchel, C. Pigment organization in fucoxanthin chlorophyll a/c2 proteins (FCP) based on resonance Raman spectroscgopy and sequence analysis. Biochimica et Biophysica Acta-Bioenergetics. 1797 (9), 1647-1656 (2010).
- Gildenhoff, N., Herz, J., Gundermann, K., Büchel, C., Wachtveitl, J. The excitation energy transfer in the trimeric fucoxanthin-chlorophyll protein from Cyclotella meneghiniana analyzed by polarized transient absorption spectroscopy. Chemical Physics. 373 (1), 104-109 (2010).
- Ramanan, C., et al. Exploring the mechanism(s) of energy dissipation in the light harvesting complex of the photosynthetic algae Cyclotella meneghiniana. Biochimica et Biophysica Acta-Bioenergetics. 1837 (9), 1507-1513 (2014).
- Haferkamp, S., Kirchhoff, H. Significance of molecular crowding in grana membranes of higher plants for light harvesting by photosystem II. Photosynthesis Research. 95 (2-3), 129-134 (2008).
- Wahadoszamen, M., et al. Stark fluorescence spectroscopy reveals two emitting sites in the dissipative state of FCP antennas. Biochimica et Biophysica Acta-Bioenergetics. 1837 (1), 193-200 (2014).
- Zhou, F., et al. Effect of monogalactosyldiacylglycerol on the interaction between photosystem II core complex and its antenna complexes in liposomes of thylakoid lipids. Photosynthesis Research. 99 (3), 185-193 (2009).
- Moya, I., Silvestri, M., Vallon, O., Cinque, G., Bassi, R. Time-resolved fluorescence analysis of the photosystem II antenna proteins in detergent micelles and liposomes. Biochemistry. 40 (42), 12552-12561 (2001).
- Natali, A., et al. Light-harvesting Complexes (LHCs) Cluster Spontaneously in Membrane Environment Leading to Shortening of Their Excited State Lifetimes. Journal of Biological Chemistry. 291 (32), 16730-16739 (2016).
- Gundermann, K., Büchel, C. Factors determining the fluorescence yield of fucoxanthin-chlorophyll complexes (FCP) involved in non-photochemical quenching in diatoms. Biochimica et Biophysica Acta-Bioenergetics. 1817 (7), 1044-1052 (2012).
- Ahmad, R. A., Dietzel, L. Relaxation of cellular K+ gradients by valinomycin induces diatoxanthin accumulation in Cyclotella meneghiniana cells and alters FCPa fluorescence yield in vitro. Physiologia Plantarum. , 171-180 (2017).
- Provasoli, L., McLaughlin, J. J. A., Droop, M. R. The development of artificial media for marine algae. Archiv für Mikrobiologie. 25 (4), 392-428 (1957).
- Jeffrey, S., Humphrey, G. New spectrophotometry equations for determining chlorophyll a, chlorophyll b, chlorophyll c-1 and chlorophyll c-2 in higher plants, algae and natural phytoplankton. Biochemie und Physiologie der Pflanzen. 167, 191-194 (1975).
- Beer, A., Gundermann, K., Beckmann, J., Büchel, C. Subunit Composition and Pigmentation of Fucoxanthin−Chlorophyll Proteins in Diatoms: Evidence for a Subunit Involved in Diadinoxanthin and Diatoxanthin Binding. Biochemistry. 45 (43), 13046-13053 (2006).
- Schägger, H., von Jagow, G. Tricine-sodium dodecyl sulfate-polyacrylamide gel electrophoresis for the separation of proteins in the range from 1 to 100 kDa. Analytical Biochemistry. 166 (2), 368-379 (1987).
- Büchel, C. Fucoxanthin-Chlorophyll Proteins in Diatoms: 18 and 19 kDa Subunits Assemble into Different Oligomeric States. Biochemistry. 42 (44), 13027-13034 (2003).
- Vieler, A., Wilhelm, C., Goss, R., Süß, R., Schiller, J. The lipid composition of the unicellular green alga Chlamydomonas reinhardtii and the diatom Cyclotella meneghiniana investigated by MALDI-TOF MS and TLC. Chemistry and Physics of Lipids. 150 (2), 143-155 (2007).
- Gundermann, K., Büchel, C. The fluorescence yield of the trimeric fucoxanthin-chlorophyll-protein FCPa in the diatom Cyclotella meneghiniana is dependent on the amount of bound diatoxanthin. Photosynthesis Research. 95 (2-3), 229-235 (2008).
- Miloslavina, Y., et al. Ultrafast fluorescence study on the location and mechanism of non-photochemical quenching in diatoms. Biochimica et Biophysica Acta-Bioenergetics. 1787 (10), 1189-1197 (2009).
- Grouneva, I., Jakob, T., Wilhelm, C., Goss, R. The regulation of xanthophyll cycle activity and of non-photochemical fluorescence quenching by two alternative electron flows in the diatoms Phaeodactylum tricornutum and Cyclotella meneghiniana. Biochimica et Biophysica Acta-Bioenergetics. 1787 (7), 929-938 (2009).
- Chukhutsina, V. U., Büchel, C., van Amerongen, H. Disentangling two non-photochemical quenching processes in Cyclotella meneghiniana by spectrally-resolved picosecond fluorescence at 77 K. Biochimica et Biophysica Acta-Bioenergetics. 1837 (6), 899-907 (2014).
- Ghazaryan, A., Akhtar, P., Garab, G., Lambrev, P. H., Büchel, C. Involvement of the Lhcx protein Fcp6 of the diatom Cyclotella meneghiniana in the macro-organisation and structural flexibility of thylakoid membranes. Biochimica Et Biophysica Acta-Bioenergetics. 1857 (9), 1373-1379 (2016).
- Darley, W. M. Biochemical composition. The biology of diatoms. 13, 198-223 (1977).
- Milsman, M. H. W., Schwendner, R. A., Weder, H. G. Preparation of large single bilayer liposomes by a fast and controlled dialysis. Biochimica Et Biophysica Acta. 512 (1), 147-155 (1978).
- Zumbuehl, O., Weder, H. G. Liposomes of controllable size in the range of 40 to 180 nm by defined dialysis of lipid-detergent-mixed micelles. Biochimica Et Biophysica Acta. 640 (1), 252-262 (1981).
- Verchere, A., Broutin, I., Picard, M. Photo-induced proton gradients for the in vitro investigation of bacterial efflux pumps. Scientific Reports. 2 (306), (2012).
- Chevallet, M., Luche, S., Rabilloud, T. Silver staining of proteins in polyacrylamide gels. Nature Protocols. 1 (4), 1852-1858 (2006).
Ristampe e Autorizzazioni
Richiedi autorizzazione per utilizzare il testo o le figure di questo articolo JoVE
Richiedi AutorizzazioneThis article has been published
Video Coming Soon
Personale delle biblioteche
Copyright © 2025 MyJoVE Corporation. Tutti i diritti riservati