
Method Article
Producción eficiente y escalable de variantes completas de huntingtina humana en células de mamíferos utilizando un sistema de expresión transitoria
En este artículo
Resumen
Proporcionamos protocolos escalables que cubren el diseño de constructo, la transfección transitoria y la expresión y purificación de variantes de proteína huntingtina humana de longitud completa en células HEK293.
Resumen
La huntingtina de longitud completa (FL HTT) es una proteína grande (aa 1-3,144), expresada ubicuamente, que contiene poliglutamina (polyQ) con una masa de aproximadamente 350 kDa. Si bien la función celular de FL HTT no se comprende completamente, una expansión mutante del tracto polyQ por encima de ~ 36 repeticiones se asocia con la enfermedad de Huntington (HD), y la longitud de polyQ se correlaciona aproximadamente con la edad de inicio. Para comprender mejor el efecto de la estructura en la función del HTT mutante (mHTT), se requieren grandes cantidades de la proteína. La producción de submiligramos de FL HTT en células de mamíferos se logró utilizando la expresión de líneas celulares estables inducibles por doxiciclina. Sin embargo, la producción de proteínas a partir de líneas celulares estables tiene limitaciones que pueden superarse con métodos de transfección transitoria.
Este documento presenta un método robusto para la producción en cantidades de bajos miligramos de FL HTT y sus variantes a partir de plásmidos optimizados para codones por transfección transitoria utilizando polietilenimina (PEI). El método es escalable (>10 mg) y produce consistentemente 1-2 mg/L de cultivo celular de FL HTT altamente purificado. De acuerdo con informes anteriores, se encontró que el estado de la solución purificada de FL HTT era altamente dinámico; La proteína tiene una propensión a formar dímeros y oligómeros de alto orden. Una clave para ralentizar la formación de oligómeros es trabajar rápidamente para aislar las fracciones monoméricas de las fracciones diméricas y oligoméricas de alto orden durante la cromatografía de exclusión de tamaño.
Se utilizó cromatografía de exclusión de tamaño con dispersión de luz multiángulo (SEC-MALS) para analizar el dímero y el contenido oligomérico de orden superior de HTT purificado. No se observó correlación entre la longitud de FL HTT polyQ (Q23, Q48 y Q73) y el contenido de oligómeros. La construcción exon1-deleted (aa 91-3,144) mostró una propensión a oligomerización comparable a FL HTT (aa 1-3,144). Aquí se describen los métodos de producción, purificación y caracterización por índice de refracción (RI) SEC/MALS, electroforesis en gel de dodecilsulfato-poliacrilamida de sodio (SDS-PAGE), western blot, PAGE nativo y Blue Native PAGE.
Introducción
La enfermedad de Huntington (EH) es una enfermedad neurodegenerativa rara caracterizada principalmente por movimientos motores inestables e involuntarios, así como alteraciones cognitivas y psiquiátricas, como cambios de personalidad y apatía 1,2. La EH se asocia con una expansión del tracto de repetición CAG localizado en el exón 1 del gen huntingtina (HTT) a más de 35 repeticiones, con un mayor número de repeticiones CAG que se correlacionan con un inicio más temprano de la enfermedad 3,4. El producto traslacional de HTT, la proteína huntingtina (HTT), está implicado en la viabilidad neuronal y el desarrollo cerebral 5,6,7,8,9.
HTT es una proteína de andamiaje que participa en una amplia gama de procesos celulares, transporte de vesículas, división celular, ciliogénesis y autofagia10,11. Sin embargo, la patogénesis molecular de la EH no está del todo clara, y falta la identificación de interactores proteicos clave que medien el impacto patológico de la mHTT expandida por poliQ. Algunas investigaciones sugieren una ganancia de la función tóxica de mHTT impulsada por la propensión a la oligomerización de la proteína HTT expandida, ya que se han identificado agregados de HTT en neuronas y glía en pacientes con EH y modelos animales de la enfermedad 12,13,14,15,16,17 . Para impulsar la investigación de la función y estructura de las variantes FL HTT y mHTT y suministrar a los investigadores estándares de proteínas de alta calidad para el desarrollo de ensayos, se necesita un suministro robusto y escalable de proteína recombinante homogénea.
Debido a su tamaño (aa 1-3,144, numeración basada en la longitud de polyQ Q23), inestabilidad proteolítica y propensión a agregarse, FL HTT ha demostrado ser difícil de expresar y aislar como proteína soluble. Anteriormente, la región del exón 1 (aa 2-90) de HTT se ha expresado y purificado a gran escala utilizando varias etiquetas que pueden aumentar la solubilidad de la proteína en Escherichia coli18,19,20. FL HTT se expresó y purificó por primera vez en un sistema de expresión de células de insectos utilizando baculovirus 21,22, y se informaron estructuras de microscopía electrónica (EM) de 30 Å de baja resolución de FL Q23-HTT y Q78-HTT químicamente reticuladas23. La investigación de la estructura de HTT avanzó aún más cuando la producción de FL Q17, Q46 y Q128-HTT con modificaciones postraduccionales nativas (PTM) se logró en células humanas utilizando líneas celulares estables o sistemas de expresión de adenovirus24. Estos estudios sugieren que aunque el HTT purificado existe principalmente en estado monomérico, también tiende a formar oligómeros y agregados de alto orden.
La ultracentrifugación analítica de FL Q128-HTT, con una región poliQ altamente expandida, proporcionó más fracciones oligoméricas y agregadas que la proteína con la región poliQ no expandida24. Utilizando una línea celular estable, se ha adaptado con éxito una estrategia para estabilizar FL HTT mediante la coexpresión con el socio de interacción HAP40. Una estructura crio-EM del complejo FL HTT y HAP40 ha sido resuelta a una resolución promedio de 4 Å utilizando el complejo proteico purificado (PDB:6EZ8)25. Esta estrategia de coexpresión ha sido adaptada con éxito a un sistema de baculovirus, y una serie de variantes de HTT de alta calidad con diferentes longitudes de poliQ han sido expresadas y purificadas a partir de células de insectos26. Desde entonces, más estructuras crio-EM del complejo de HTT con longitudes variables de polyQ y estructuras HAP40 y de mayor resolución fueron resueltas y depositadas en la Base de Datos de Proteínas27,28 (PDB: 7DXK, 7DXH, 6X9O).
Optimizamos un método de transfección y expresión en células HEK293, utilizando polietilenimina (PEI), para la expresión transitoria rápida de FL HTT. Como prueba de principio, las variantes de FL HTT que contienen 23 glutaminas (FL Q23-HTT) se purificaron y caracterizaron por primera vez utilizando una modificación de un método de purificación descrito anteriormente24. Este método de transfección transitoria es conveniente, altamente eficiente y escalable; puede producir HTT purificada con rendimientos de 1-2 mg/L, comparable al método de línea celular estable reportado24. Debido a que la proteína se produce en una línea celular humana, es más probable que la HTT producida tenga PTM humanos nativos cuando se somete a un análisis proteómico de espectrometría de masas 11,29,30,31. Se produjeron cantidades de miligramos de las variantes FL Q48-HTT, FL Q73-HTT y exon1-deleted (ΔExon1-HTT) de FL HTT, lo que demuestra que el método de expresión transitoria es especialmente útil para producir rápidamente variantes alternativas de HTT sin depender del esfuerzo requerido para establecer líneas celulares estables para la producción.
El siguiente protocolo ejemplifica el método estándar utilizado en el laboratorio de estos autores para el cultivo celular, la transfección, la purificación de proteínas y la caracterización de proteínas posteriores a la purificación para producir FL Q23-HTT a partir de un cultivo celular de 2 L. El protocolo puede ampliarse a cultivos más grandes o adaptarse para purificar otras variantes de HTT. Hasta 10 L de cultivos celulares de FL HTT y varias mutaciones de sitio o truncamiento de homólogos de HTT y HTT se han realizado con éxito en el laboratorio utilizando el mismo protocolo. Purified FL HTT contiene un alto porcentaje de monómeros junto con dímeros y oligómeros de orden superior. El mismo perfil agregado se observa entre las variantes producidas (Q23, Q48, Q73 y Exon1 eliminado). Como la agregación puede ocurrir cuando no se toma el cuidado adecuado, se realizó un estudio de estabilidad de formulación y congelación-descongelación para identificar las mejores condiciones para el manejo de proteínas. También se describen métodos, como Blue Native PAGE y SEC/MALS-RI, para analizar el contenido de oligómeros HTT como parte del proceso de control de calidad. Para beneficiar a la comunidad de investigación de la EH, los plásmidos y las proteínas HTT descritas en este estudio también se depositan en el Repositorio de la Comunidad de la EH en el Instituto Coriell (www.coriell.org/1/CHDI).
Protocolo
1. Diseño y producción de constructos para la expresión de mamíferos HTT etiquetados con FLAG
- Recuperar la secuencia completa de proteína HTT humana (P42858) del Centro Nacional de Información Biotecnológica (https://www.ncbi.nlm.nih.gov/).
NOTA: Los investigadores deben estar familiarizados con las organizaciones de dominio de HTT y mantener la estructura 3D central de HTT al diseñar construcciones para mutantes de HTT. - Solicitar un servicio de síntesis génica para realizar la optimización de codones para la expresión celular humana basada en la secuencia de P42858. Cambie el número polyQ de Q16 a la longitud Q deseada (Q23 fue elegido como la primera construcción aquí) y sintetice el gen HTT de longitud completa.
NOTA: La construcción Q23-HTT de longitud completa optimizada para codones sintetizados se entregó como un inserto en el plásmido pUC18 en este estudio. - Opcional: Agregue características para facilitar la clonación de diferentes longitudes Q y la purificación en las construcciones.
NOTA: Se agregaron un sitio de escisión del virus del grabado del tabaco (TEV) y una etiqueta de purificación FLAG (AAAENLYFQGDYKDDDDK) al extremo C-terminal de las construcciones. Se diseñaron dos sitios HindIII en los constructos para abarcar la región polyQ (la secuencia de proteínas traducida no se cambia mediante la introducción de sitios HindIII). Esto permite al investigador cambiar la longitud Q de HTT mediante la digestión y ligadura de enzimas de restricción sin resintetizar el gen HTT completo.
2. Clone las construcciones HTT sintetizadas en pcDNA3.1.
- Digerir 5 μg de pUC18-Q23-HTT y 5 μg de pcDNA3.1 usando 2 μL de NheI y PmeI cada uno a 37 °C durante 2 h.
- Ejecute un gel de agarosa al 0,5% p/v y purifique el fragmento Q23-HTT y el vector pcDNA3.1 digerido utilizando un kit de extracción en gel de agarosa. Cuantificar las concentraciones de ADN purificado por OD280 utilizando un espectrómetro UV que puede medir microlitros de muestras.
NOTA: Normalmente se observa OD260/280 que oscila entre 1,8 y 2,0. El FL HTT sintetizado se suministra como un inserto con NheI y PmeI en ambos extremos en un plásmido pUC18. Use otras enzimas de restricción si la HTT se sintetiza de manera diferente. - Utilice 10 ng de vector pcDNA3.1 digerido en la reacción. Ligate los ADN purificados a una relación molar 1:1 (HTT:pcDNA3.1) en una reacción de 10 μL a temperatura ambiente durante 5 min usando ADN ligasa T4.
- Transformar el producto ligado en células competentes de E. coli (ver la Tabla de materiales) utilizando el protocolo especificado por el fabricante de la ligasa.
- Recoger 6 colonias individuales y hacer cultivos nocturnos en 4-6 mL de LB suplementados con 100 μg/mL de carbenicilina a 37 °C.
- Asigne 1 ml de cada cultivo nocturno. Añadir glicerol al 25% v/v y guardar el stock de glicerol a -80 °C. Purifique el cultivo restante durante la noche utilizando un mini kit de preparación de acuerdo con los pasos especificados en el manual del usuario.
- Secuencie todos los plásmidos utilizando cebadores de secuenciación que abarcan toda la región de transcripción del plásmido. Elija un stock de glicerol que tenga la secuencia correcta como stock de glicerol maestro y descarte el resto.
- Opcional: Solicite un servicio de síntesis de genes para sintetizar ADN con las diferentes longitudes Q (Q48, Q73 y Exon1) que abarcan los dos sitios HindIII en el plásmido pcDNA3.1-Q23-HTT. Digiera pcDNA3.1-Q23-HTT y los ADN recién sintetizados usando HindIII, y religarlos con ligasa T4 como en los pasos 2.2-2.7 para hacer FL HTT con diferentes longitudes de poliQ en el plásmido pcDNA3.1.
NOTA: Las construcciones de plásmidos utilizadas en este estudio también están disponibles directamente en el Repositorio Comunitario de HD en el Instituto Coriell (www.coriell.org/1/CHDI); véase la Tabla de materiales.
3. ADN plásmido libre de endotoxinas GIGA prep para transfección a gran escala
- Rayar las reservas bacterianas de glicerol de pcDNA3.1-Q23-HTT-TEV-FLAG en una placa de agar LB con carbenicilina (100 μg/mL). Incubar la placa a 37 °C durante 16-24 h hasta que aparezcan colonias individuales.
- Recoger una sola colonia, inocular un cultivo iniciador de 5 ml en un medio rico formulado para la amplificación de plásmidos con carbenicilina (100 μg/ml) y crecer a 37 °C durante 8 h.
- Elija un kit de purificación de plásmidos GIGA libre de endotoxinas. Siga los pasos descritos en el manual del kit GIGA del plásmido para purificar el plásmido pcDNA3.1-Q23-HTT-TEV-FLAG.
- Mida los niveles de endotoxinas plásmidas utilizando un kit de cuantificación de endotoxinas basado en lisado de amebocitos limulus (LAL). Siga el procedimiento especificado en el manual del fabricante.
NOTA: Una purificación de plásmidos de alta calidad y bajo nivel de endotoxinas es esencial para obtener una buena eficiencia de transfección. Usando este protocolo, se pueden obtener 20-40 mg de plásmido (forma superenrollada >80%) por L de cultivo bacteriano a concentraciones de plásmidos > 4 mg/ml. Un plásmido debidamente purificado debe tener un nivel de endotoxinas < 30 UE/mg. Normalmente se observa OD260/280 que oscila entre 1,8 y 2,0.
4. Transfección a gran escala de 2 L de células HEK293 mediante polietilenoimina (PEI)
- Agregue 1 g de PEI 25K a 1 L de agua libre de endotoxinas con agitación. Ajuste el pH a 2.0 usando HCl de 100 mM y revuelva hasta que todo el PEI 25K se disuelva. Ajustar el pH a 7,0 utilizando una solución de NaOH de 100 mM y filtrar a través de un filtro de 0,2 μm. Alícuota y conservar a -20 °C durante un máximo de un año.
NOTA: Las alícuotas de PEI pueden conservarse a 4 °C durante un máximo de dos semanas, pero nunca deben volver a congelarse después de la descongelación. - Propagar células HEK293 en el medio de crecimiento (ver la Tabla de materiales) suplementadas con penicilina-estreptomicina (concentración final a 5 U/ml para penicilina y 5 μg/ml para estreptomicina) en una incubadora agitadora humidificada a 37 °C, 90 rpm, 5% deCO2 durante 18-24 h. Diluir las células a 2 L a una densidad de ~1,2 × 106 células/ml utilizando el medio de cultivo en matraces Erlenmeyer de 5 L un día antes de la transfección.
- Continuar creciendo las células a 37 °C, 90 rpm, 5%CO2 durante 18-24 h. Mida los parámetros de la celda utilizando un contador automático de celdas capaz de medir la densidad y viabilidad de las celdas siguiendo el manual del usuario.
NOTA: La densidad celular debe duplicarse y la viabilidad debe ser del >95%. La densidad celular antes de la transfección debe ser de aproximadamente 2,0 × 10 6-2,4 × 106 células/ml. Diluir las células a la densidad deseada antes de la transfección cuando sea necesario. - Calcular las cantidades de plásmido y PEI requeridas para la transfección; usar 1 mg de plásmido y 3 mg de PEI para la transfección de cada litro de cultivo celular. Asigne 2 mg de plásmido y 6 mg de PEI necesarios para una transfección de 2 L.
- Diluir el plásmido y la PEI individualmente en un volumen de solución salina tamponada con fosfato igual a 1/20del volumen total de cultivo celular (100 ml cada uno para una transfección de 2 L) e incubar a temperatura ambiente durante 5 min. Mezclar el plásmido diluido y la PEI girando suavemente e incubar la mezcla a temperatura ambiente durante 30 min.
NOTA: La mezcla aparecerá ligeramente turbia después de la incubación. - Agregue la mezcla al cultivo celular y agite suavemente para mezclarlos.
- Cultivar las células a 37 °C, 5%CO2, 90 rpm durante 24 h.
- Añadir una solución de butirato sódico 2 M a una concentración final de 2 mM. Añadir al cultivo un agente antiaglutinante 1:1000 (v/v) y un antiespumante 1:1000 (v/v).
- Mover el matraz a una incubadora agitadora humidificada a 32 °C, 90 rpm, 5%CO2, y continuar creciendo durante 48 h.
- Mida los parámetros de la celda, incluida la densidad y la viabilidad de la celda, utilizando el contador automático de celdas siguiendo el manual del usuario.
- Transfiera 2,0 × 10 6 células (Vol = 2,0 × 106/densidad celular) en un tubo de microcentrífuga. Granular las células a 2.000 × g durante 1 minuto en una centrífuga para Western blot en la sección 5.
- Cosechar las células por centrifugación a 2.000 × g durante 30 min y almacenar el pellet de células a -80 °C antes de la purificación.
5. SDS-PAGE y western blot de lisado celular HEK293 para estimar el nivel de expresión de HTT
- Tomar una alícuota de 2,0 × 106 células previamente congeladas (paso 4.11) de la transfección a gran escala del cultivo celular HEK293. Agregue 250 μL de solución salina tamponada con Tris (TBS) suplementada con 50 μg / ml de digitonina, 5 mM EDTA y 1x cóctel de inhibidores de proteasa, y vuelva a suspender el gránulo celular aspirando varias veces con una pipeta.
- Gire los tubos suavemente durante 30 minutos a 4 °C utilizando un minirotador para lisar las células. Granular el material insoluble centrifugando a 17.000 × g durante 5 min.
- Agregue 1/3del volumen de tampón de carga de dodecilsulfato de litio (LDS) reductor 4x al sobrenadante y caliente a 70 °C durante 10 min.
- Cargue 5-20 μL de lisado celular en un gel PAGE prefabricado de acetato de Tris al 3-8%. Usando el tampón de funcionamiento SDS 1x Tris-acetato compatible con gel, ejecute el gel en un modo de voltaje constante a 150 V durante 60 min.
NOTA: Se utilizó Tris-acetato SDS-PAGE para el análisis FL HTT porque genera una resolución más alta que otros tipos de SDS-PAGE para las proteínas con peso molecular superior a 300 kDa. Las proteínas utilizadas en este estudio también están disponibles directamente en el Repositorio Comunitario de la EH en el Instituto Coriell (www.coriell.org/1/CHDI); véase la Tabla de materiales. - Para realizar Western Blotting, ensamble un sándwich de transferencia con un papel de transferencia grueso equilibrado con tampón de transferencia, una membrana de fluoruro de polivinilideno activado por metanol (PVDF) y un gel SDS-PAGE. Transfiera las proteínas a la membrana de PVDF utilizando un secante occidental semiseco de acuerdo con el manual del usuario del fabricante.
NOTA: Normalmente, 20-30 min a 135 mA es suficiente para una membrana de 10 cm x 10 cm. - Desmonte el sándwich de transferencia y bloquee la membrana en TBST (20 mM Tris pH 7.4, 150 mM NaCl y 0.1% v/v Tween-20) suplementado con 5% p/v de leche descremada.
- Incubar la membrana en un balancín durante 1 h a temperatura ambiente con 15 ml de anticuerpo primario (dilución 1:2.500 para el anticuerpo monoclonal anticuerpo anti-FLAG y 1:2.000 para todos los demás anticuerpos primarios).
NOTA: Los anticuerpos primarios utilizados en este estudio son anti-FLAG M2, MAB5492, MAB5490, MAB2166, MAB3E10, MAB4E10, MAB2168, MAB8A4 (consulte la Tabla de materiales). - Lave la membrana 3 x 5 min usando 30-50 ml de TBST.
- Incubar la membrana en un balancín con un anticuerpo secundario IgG de cabra conjugado con colorante fluorescente a 1:15.000, a temperatura ambiente, en 15 ml de TBST que contiene 5% p/v de leche en polvo.
- Visualice las bandas de Western blot en un generador de imágenes fluorescente utilizando la longitud de onda específica del anticuerpo secundario. Quantitar la señal de banda utilizando el software que acompaña al generador de imágenes según el manual del usuario.
NOTA: La Western Blot cuantitativa se puede realizar utilizando HTT purificada como estándar. Un rango estándar lineal de HTT es específico del instrumento y se estableció en este laboratorio de 25 ng a 250 ng de HTT por carril utilizando un anticuerpo anti-FLAG. La mancha occidental de HTT debe estar libre de degradación; normalmente se observa un nivel total de expresión de HTT de 2-4 pg/célula. Consulte un protocolo32 publicado anteriormente para obtener detalles sobre cómo realizar un western blot cuantitativo.
6. Purificación rápida de cromatografía líquida de proteínas (FPLC) de HTT utilizando columna anti-FLAG y SEC
- Purificación anti-FLAG
- Estimar la cantidad de resina FLAG necesaria para la purificación (típicamente, 12 mL de resina de afinidad anti-FLAG M2 para la purificación de 2-4 L de cultivo celular transfectado). Empaque 12-25 ml de resina anti-FLAG en una columna vacía (consulte la Tabla de materiales) utilizando FPLC a un caudal de 4 ml / min utilizando el tampón A (Tabla 1). Ajuste la altura del émbolo, para que no haya espacio entre el extremo del émbolo y el lecho de resina.
- Utilizando una proporción de 10 ml de tampón de lisis por 1 g de pellet celular, descongele y suspenda el pellet celular en tampón de lisis frío (Tabla 1).
- Pase la suspensión celular una vez a través de un homogeneizador de alto cizallamiento a 10,000 psi. Aclarar el lisado por centrifugación a 20.000 × g durante 1 h en una centrífuga equipada con un rotor de ángulo fijo compatible.
- Programe el FPLC (consulte la Tabla de materiales para el software utilizado en el estudio) y ejecute las siguientes secuencias.
- Cargue el lisado clarificado a través de la bomba de muestra.
- Lavar con volúmenes de 4 columnas (CV) del tampón A (Tabla 1).
- Lavar con 4 CVs de Tampón B (Tabla 1).
- Lavar con 8 CVs de Tampón C (Tabla 1).
- Lavar con 3 CVs de Tampón D (Tabla 1).
- Lavar con 3 CVs de Elution Buffer (Tabla 1).
- Analice 10 μL de las fracciones pico utilizando SDS-PAGE. Recolectar y combinar las fracciones pico con la pureza deseada. Ahorre ~50 μL de los eluidos combinados para el análisis SDS-PAGE.
NOTA: Normalmente, aparecerá un solo pico, y todas las fracciones eluidas en el pico contienen ~ 90% de HTT puro. - Regenere una columna anti-FLAG usando 5 CVs de Buffer de Regeneración (Tabla 1) y reequilibre la columna usando 5 CVs de Buffer A.
NOTA: La resina Anti-FLAG se puede reutilizar hasta cinco veces o hasta que el rendimiento relativo/litro descienda al 50% de la primera purificación.
- Purificación de la columna de exclusión de tamaño (SEC) mediante una columna SEC
- Preequilibrar una columna SEC que permite la separación de proteínas con peso molecular (MW) > 500 kDa (ver la Tabla de Materiales para la columna utilizada) utilizando 2 × CV de Tampón SEC (Tabla 1).
- Cargue directamente el eluido anti-FLAG (del paso 6.1.5) a través de un superloop de 50 ml. Ejecute 1.2 × CV de búfer SEC por inyección. Ejecute la separación SEC durante la noche a 4 °C.
NOTA: Se puede cargar un máximo de 5 ml o 15 ml de muestra de proteína en las columnas SEC seleccionadas en este estudio. Programe el FPLC para que se puedan realizar múltiples inyecciones automáticamente. Los scripts de métodos de ejemplo también se incluyen como archivo complementario 1 y archivo complementario 2. - Compare el perfil de elución con el perfil de elución HTT estándar para distinguir los picos de monómero, dímero y oligomérico de orden superior. Agrupe las fracciones monoméricas de HTT en función del perfil de elución de la columna SEC. Si lo desea, agrupe las fracciones HTT oligoméricas y diméricas de orden superior por separado.
- Concentrar la proteína HTT agrupada utilizando un concentrador centrífugo de 100 kDa a 4 °C. Calcule las concentraciones de proteínas dividiendo sus valores de OD280 por los respectivos coeficientes de extinción (los coeficientes de extinción teóricos de Q23-HTT, Q48-HTT, Q73-HTT y ΔExon1-HTT son 0.776, 0.769, 0.762 y 0.798 (mg/mL)-1 cm-1, respectivamente, para el cálculo). Mantener la concentración de HTT ≤ 1,0 mg/ml.
NOTA: Es esencial monitorear el proceso de concentración ya que la sobreconcentración dará como resultado la agregación. - Alícuota de la proteína HTT purificada en tubos de microcentrífuga crioseguros en un volumen < 100 μL. Congelar rápidamente las alícuotas con nitrógeno líquido y almacenarlas a -80 °C.
7. Analítica HPLC SEC-MALS-dRI para analizar la polidispersidad HTT
- Realice todos los análisis SEC-MALS a 4 °C en un sistema de cromatografía líquida (HPLC) de alta resolución junto con un detector UV, un detector de dispersión de luz multiángulo y un detector de índice de refracción diferencial (dRI).
- Antes de conectar la columna UHPLC al sistema, purgue la bomba y los detectores con agua filtrada (0,1 μm) de grado HPLC.
- Conecte la columna UHPLC (consulte la Tabla de materiales para la columna utilizada) al sistema. Equilibre la columna con agua filtrada (0,1 μm) y luego el búfer SEC-MALS (Tabla 1) hasta que todas las señales del detector alcancen la línea de base.
- Inyecte 2 μL de 6 mg/ml de albúmina sérica bovina (BSA) a un caudal de 0,3 ml/min durante 15 min por inyección e inspeccione la calidad de los datos. Realice la normalización, la alineación de picos y la corrección de ampliación de banda en función del perfil BSA y cree una plantilla para las siguientes ejecuciones de muestra HTT.
- Descongele rápidamente un vial de la muestra FL Q23-HTT en un baño de agua a temperatura ambiente utilizando un flotador. Filtre el HTT a través de un filtro de espín de 0,1 μm. Inyectar 2-4 μL de la muestra HTT y ejecutar durante 15 min a 4 °C a un caudal de 0,3 ml/min.
- Analice los datos cromatográficos y de dispersión de la luz utilizando el software que lo acompaña (consulte la Tabla de materiales). Utilice el detector dRI como detector de concentración y utilice 0,185 como incremento del índice de refracción (dn/dc) para HTT. Generar un diagrama Zimm para determinar la masa molecular promediada en peso para cada pico33,34.
NOTA: El incremento del índice de refracción de HTT se calcula como 0.185 utilizando el software SEDFIT35 del programa y la secuencia de aminoácidos primarios de HTT como entrada.
NOTA: El monómero HTT MW es determinado por SEC-MALS a ~370 kDa ± 30 kDa. El HTT purificado típicamente tiene un contenido de monómero entre 60 y 75% (en este laboratorio). El bajo contenido de monómeros puede indicar que se debe tener más cuidado en el manejo para evitar la agregación.
8. Blue Native PAGE para analizar la polidispersidad HTT
- Prepare 1 L de tampón de ánodo mezclando 50 ml de búfer de ejecución Blue Native PAGE 20x (consulte la tabla de materiales) con 950 ml deH2O. Prepare 2 L de tampón catódico azul oscuro mezclando 100 ml de búfer de ejecución Blue Native PAGE 20x y 100 ml de aditivo de cátodo PAGE nativo azul (20x) con 1.800 ml de H2O. Enfríe los tampones a 4 °C antes de usarlos.
- Descongele rápidamente un vial de muestra FL Q23-HTT en un baño de agua a temperatura ambiente utilizando un flotador. Mantenga la proteína descongelada en hielo antes de usarla.
- Mezcle 5 μg de FL Q23-HTT (~1 mg/ml), 1 μL de aditivo G250 al 0,5%, 2,5 μL de tampón de muestra Blue Native PAGE 4x y agua para llevar el volumen final a 10 μL.
- Cargue la muestra mixta FL Q23-HTT en un gel prefabricado Bis-Tris al 3-12%. Cargue 7,5 μL del patrón de proteína no teñida en el mismo gel que el patrón.
- Llene la parte delantera del tanque con el amortiguador de cátodo azul oscuro y la parte posterior del tanque con el búfer de ánodo.
NOTA: Rellene los búferes después de cargar la muestra para permitir una fácil visualización al cargar las muestras. - Deje correr el gel a 150 V durante 120 minutos en una habitación fría.
- Destain el gel con Solución Detinente (Tabla 1) hasta que se observen bandas; Transfiera el gel al agua. Visualice y documente el gel en una estación de imágenes.
NOTA: Blue Native PAGE fue diseñado originalmente para analizar proteínas de membrana. Se adaptó en este laboratorio como un método alternativo para estimar el contenido monomérico de HTT. Se une a las regiones hidrófobas de HTT y evita que forme agregados en condiciones de tampón que carecen de detergente. La PAGE nativa tradicional sin usar Coomassie blue G250 hace que HTT forme oligómeros y agregados solubles, probablemente debido a las muchas bolsas hidrofóbicas existentes en HTT.
9. SDS PAGE seguido de Coomassie o tinción de plata para analizar la pureza de HTT
- Agregue 4x tampón de muestra LDS y 10x reactivo reductor a FL Q23-HTT purificado para que la concentración final del tampón de carga y el reactivo reductor sea 1x.
- Calentar la muestra en un bloque calefactor seco a 70 °C durante 10 min.
- Cargue un máximo de 1 μg de proteína por pocillo en un gel de acetato de Tris al 3-8% y ejecute a 150 V durante 1 h utilizando el tampón de funcionamiento SDS de acetato de Tris.
NOTA: Las proteínas utilizadas en este estudio también están disponibles directamente en el Repositorio Comunitario de la EH en el Instituto Coriell (www.coriell.org/1/CHDI); véase la Tabla de materiales. - Mancha de Coomassie
- Lavar el gel conH2Odurante 5 min.
- Manchar el gel en la solución de tinción de Coomassie (Tabla 1) balanceando el gel en 30 ml de solución de tinción durante 15 min.
- De-manchar meciendo el gel en 50 mL deH2Odurante 5 min. Repita dos veces. Visualice y documente el gel teñido de Coomassie en una estación de imágenes.
- Tinción de plata usando un kit comercial de tinción de plata.
- Después de SDS-PAGE, fije el gel con la solución fijadora (Tabla 1) durante 1 h hasta pasar la noche a temperatura ambiente.
- Realice la mancha, lave y revele de acuerdo con las instrucciones del kit.
- Detenga el paso de desarrollo inmediatamente una vez que las bandas alcancen la intensidad deseada.
- Documente el gel en un sistema de documentación de gel equipado con una fuente de luz visible.
NOTA: HTT purificado al >95% puede ser detectado por Coomassie y tinción de plata con este protocolo. Consulte un protocolo32 publicado anteriormente para obtener detalles sobre cómo realizar análisis cuantitativos de proteínas.
Resultados
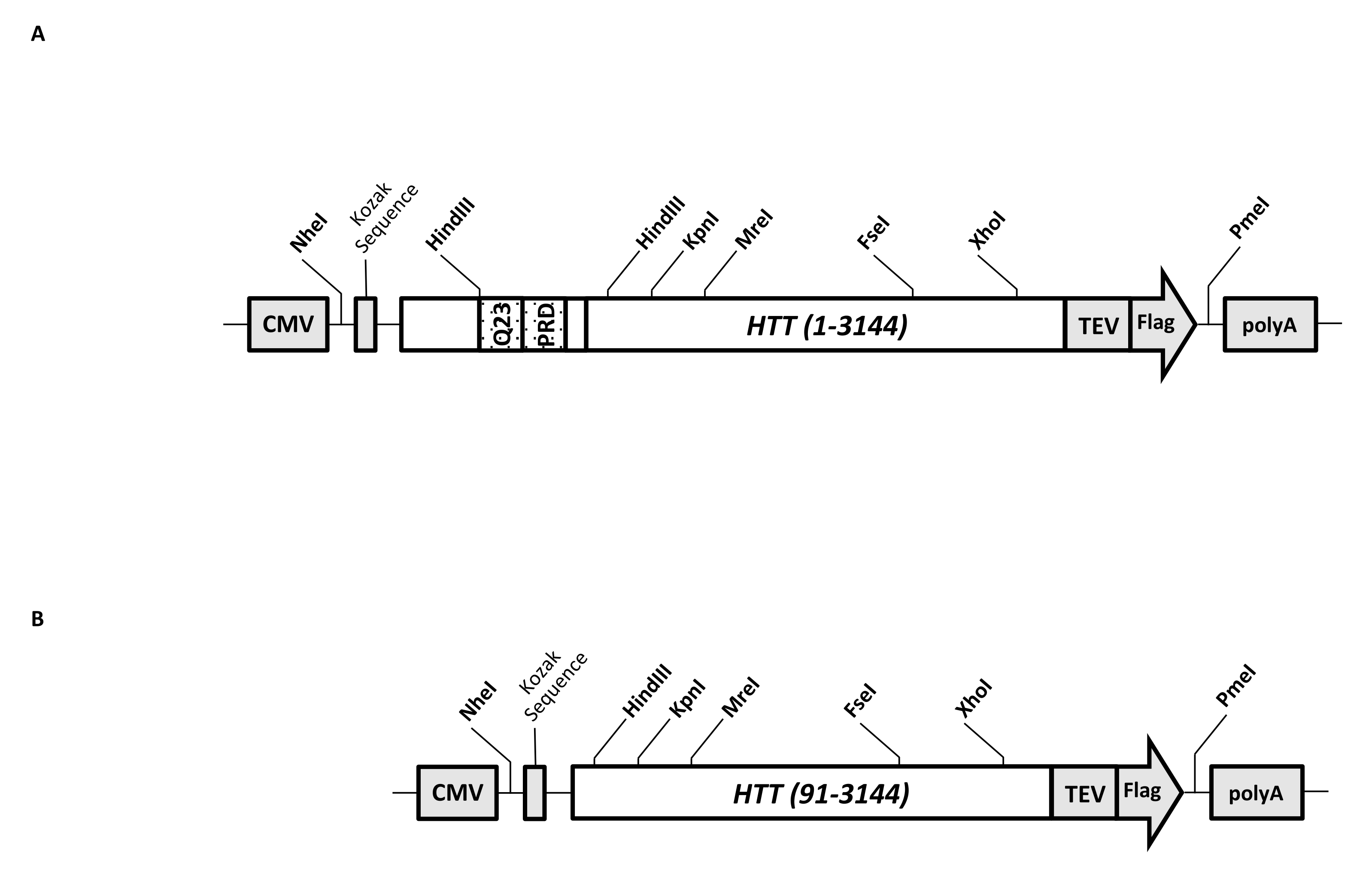
Un vector de expresión transitoria (pcDNA3.1-Q23-HTT-TEV-FLAG, Figura 1A) está diseñado para una producción rápida en células de mamíferos de FL Q23-HTT (aa 1-3,144, basado en la numeración Q23). Esta construcción tiene las características diseñadas para generar rápidamente varias construcciones de mutación HTT mediante clonación de casetes, facilitar la purificación de la proteína HTT a alta calidad y homogeneidad con pasos cromatográficos mínimos, y tener la opción de producir HTT FL sin etiquetar. La lista de características incluye 1. Los sitios de digestión de restricción HindIII, que rodean la repetición CAG en el exón 1 de HTT, se pueden usar para generar mutantes FL HTT con un estiramiento poliQ de varias longitudes mediante digestión y ligadura de enzimas de restricción; 2. el extremo C-terminal de FL HTT se marca con un epítopo FLAG con un sitio de reconocimiento de proteasa TEV para la purificación de afinidad de un solo paso de FL HTT con alta pureza y generación opcional de proteína FL HTT libre de etiquetas utilizando escisión de proteasa TEV; 3. Secuencia FL HTT optimizada para codones de células humanas para la expresión de alto nivel en células HEK293. El vector pcDNA 3.1 (+) se utiliza como columna vertebral del constructo para aprovechar la alta actividad de activación transcripcional del promotor del CMV en líneas celulares de mamíferos.
Usando pcDNA3.1-Q23-HTT-TEV-FLAG como plantilla inicial, las construcciones Q48 y Q73 FL HTT se produjeron sintetizando fragmentos de ADN con la longitud Q adecuada que abarcan dos sitios de enzimas de restricción HindIII e intercambiando la misma región en la plantilla. El mutante ΔExon1 de FL HTT (aa 91-3,144) (Figura 1B) se produjo utilizando cebadores dirigidos a residuos eliminados que abarcan la región del exón 1 en la plantilla. Las células HEK293 transfectadas con pcDNA3.1-Q23-HTT-TEV-FLAG utilizando PEI se cultivaron en matraces agitadores de 5 L por debajo del 5% deCO2. Una purificación típica a gran escala utiliza un pellet de celda de 2-10 L que contiene 6.0 × 10 9-3.0 × 1010 células. Antes de proceder a la purificación, el nivel de expresión de HTT de cada transfección se estimó mediante western blot cuantitativo utilizando HTT recombinante purificado marcado con FLAG como estándar y anticuerpo anti-FLAG como primer anticuerpo. Para la purificación se utilizaron pellets con un nivel estimado de expresión de HTT a ≥2 pg HTT/célula.
La purificación de FL HTT consiste en un proceso de columna de 2 pasos, primero con purificación de afinidad anti-FLAG y luego con SEC en una columna de filtración de gel con un rango de separación adecuado para HTT (Figura 2A; ver Tabla de materiales para ejemplos). Después de ambos pasos, HTT se obtuvo con una pureza de muestra del >95%, según lo determinado por SDS-PAGE con azul de Coomassie y >65% de contenido de monómero basado en SEC-MALS analítico. Debido a que tanto el tiempo de purificación prolongado como la temperatura tienen un impacto negativo en el contenido final de monómero HTT, FPLC se utilizó en ambos pasos de purificación para minimizar el manejo y obtener una calidad de muestra consistente. El principal contaminante durante la purificación anti-FLAG fue la chaperona Hsp70 determinada por espectrometría de masas (Figura 2B, carril 2). Esto es consistente con el hallazgo de que Hsp70 se co-purifica con FL HTT expresado de manera estable en líneas celulares humanas24, lo que sugiere que Hsp70 puede ser un estabilizador común para FL HTT in vivo.
La contaminación por Hsp70 se puede eliminar mediante un lavado extensivo con cloruro de magnesio y ATP durante el paso de purificación de afinidad anti-FLAG (Figura 2B, carril 1). Tras la eliminación de Hsp70, FL HTT es propenso a formar oligómeros de orden superior24 y debe mantenerse en una concentración ≤ 1 mg / ml. El paso de concentración antes de la SEC a menudo puede resultar en una agregación significativa. Por lo tanto, la mejor práctica es cargar directamente las fracciones máximas de la purificación anti-FLAG en la columna de exclusión de tamaño sin concentrarse. Después de la SEC, la muestra se concentró a ≤1 mg/ml para la recuperación máxima de FL HTT monomérica. La cantidad de HTT recuperada de cada paso de purificación se estimó mediante azul de Coomassie o western blot cuantitativo utilizando FL HTT purificado como estándar de cuantificación (Tabla 2). El rendimiento típico de las proteínas FL HTT purificadas producidas por el método descrito es de aproximadamente 1 mg / L de cultivo celular, pero puede caer muy por debajo de eso (Tabla 3) debido a la variabilidad de lote a lote, o si la resina de purificación anti-FLAG se reutiliza varias veces.
La sobreexpresión de FL HTT puede resultar en la fragmentación de la proteína22. FL Q23-HTT producido por el método descrito aquí resuelto como una sola banda con el MW correcto de 350 kDa por SDS PAGE, teñido por Coomassie G250 o por tinción de plata (Figura 2C). Por western blotting, FL Q23-HTT reaccionó con anticuerpos levantados contra epítopos en el N-terminal, C-terminal y varios dominios intermedios, sin que se observaran bandas adicionales relacionadas con fragmentos, lo que indica que la proteína se aisló sin truncamientos detectables significativos (Figura 3A). Las variantes de longitud de FL HTT polyQ Q23, Q48 y Q73 reaccionaron como se esperaba en Western blot, mostrando una señal progresivamente más fuerte para mAb MW1 dirigido por polyQ que se correlaciona con el aumento de la longitud Q: Q23-HTT < Q48-HTT < Q73-HTT (Figura 3B). No se observó señal para ΔExon1-HTT (aa 91-3,144) cuando se sondeó con los anticuerpos MW1 y MAB549, que se dirigen al exón 1 terminal N (Figura 3B).
SEC-MALS se empleó para analizar el estado de agregación y la masa molecular de la proteína HTT purificada. Las muestras se analizaron mediante análisis SEC monitoreados por detectores UV, MALS y dRI. La masa molar absoluta obtenida de SEC-MALS no depende de la forma de las moléculas33,34; por lo tanto, SEC-MALS proporciona una estimación imparcial de MW para fracciones monoméricas y oligoméricas cuando están bien separadas. Entre las columnas de HPLC probadas, la columna SEC (ver la Tabla de materiales) mostró una resolución suficiente entre el monómero HTT y el dímero de tal manera que se pudieran distinguir masas molares (Figura 4). La concentración de proteína se determinó mediante la detección de dRI. Los incrementos del índice de refracción (dn/dc) de FL HTT son 0,1853 ml/g calculados por el software SEDFIT35. Se observaron patrones analíticos similares de elución SEC para ΔExon1 HTT (91-3,144), FL Q23, Q48 y Q73 HTT (1-3,144), cada uno de los cuales consiste en un pico de monómero mayor con picos diméricos y oligoméricos menores (Tabla 4). El MW calculado para la forma monomérica es mayor que el MW teórico. Esto es probablemente causado por especies superpuestas de picos oligoméricos de orden superior y errores resultantes de señales dRI débiles, ya que las proteínas HTT se mantienen en baja concentración para evitar la formación de oligómeros de orden superior. Al integrar los picos UV de varios lotes de variantes purificadas de FL HTT, no se observó una correlación clara entre la longitud del poliQ y el perfil agregado (Tabla 4).
Además de la SEC analítica, se realizó PAGE nativa tradicional para determinar si se puede utilizar como un método complementario para caracterizar el estado oligomérico FL HTT. Los oligómeros de orden superior se resolvieron a través de geles de acetato de Tris al 3-8% utilizando tampón nativo sin detergente. El FL HTT purificado de SEC mostró múltiples bandas correspondientes a los estados de oligomerización (Figura 5A). La banda más baja se ubicó entre el marcador nativo 480 kDa y 720 kDa, similar a los resultados anteriores reportados para FL HTT purificado a partir de células de insectos22. Sin embargo, el monómero HTT no fue la banda más abundante cuando se utilizó PAGE nativo tradicional, y los resultados no se correlacionan con el perfil agregado determinado por SEC-MALS analítico. Varios parches hidrófobos presentes en FL HTT36,37,38, especialmente la interfaz hidrofóbica entre HAP40 y FL HTT25, es probable que contribuyan a la formación de oligómeros de orden superior durante la migración dentro del gel. Esto se debe a que se sabe que las regiones hidrofóbicas interactúan entre sí en ausencia de interacciones proteína-proteína estabilizadoras o detergentes. De acuerdo con las propiedades hidrofóbicas de HTT, FL HTT forma cantidades crecientes de fracciones oligoméricas de orden superior en ausencia de CHAPS durante el paso de purificación SEC.
Blue Native PAGE, que es ampliamente utilizado para investigar proteínas de membrana y grandes complejos de proteínas que contienen parches hidrófobos39, se comparó con PAGE nativo tradicional. El HTT purificado mostró tres bandas principales en Blue Native PAGE con MW estimados de 643, 927 y 1070 kDa (Figura 5B) que probablemente representan las especies monomérica, dimérica y trimérica de HTT, respectivamente. La banda monomérica siguió siendo la banda más abundante en la PAGE nativa azul, correspondiendo bien al perfil analítico SEC de las mismas muestras. La sobreestimación de MW del monómero HTT por Blue Native PAGE puede ser el resultado de la estructura esférica hueca única o regiones hidrófobas de HTT que causan una migración más lenta en relación con los marcadores de peso molecular correspondientes 11,23,25. En general, FL Q23-HTT, FL Q48-HTT, FL Q73-HTT y ΔExon1-HTT tienen perfiles PAGE nativos azules similares con solo ligeras diferencias en la migración de la banda de proteínas debido a sus diferencias de peso molecular.
Como control adicional de la calidad de las proteínas purificadas, la etiqueta C-terminal FLAG se puede eliminar de FL HTT mediante tratamiento con proteasa TEV. Después de la escisión proteolítica, las muestras se analizaron mediante Western blot utilizando cuatro anticuerpos para confirmar la eliminación de la etiqueta FLAG y detectar la degradación de HTT. La inmunorreactividad a anti-FLAG M2 y tres anticuerpos específicos de huntingtina con epítopos al extremo N, dominios intermedios y C-terminal de HTT mostró una eliminación exitosa de la etiqueta FLAG y ningún producto de degradación específico de HTT (Figura suplementaria S1).

Figura 1: Constructo para la expresión HTT de longitud completa. (A) El HTT Q23 de longitud completa se optimizó para codones y se clonó en plásmido pcDNA3.1 (+). El extremo 3' de HTT se marcó con epítopo Flag y sitio de escisión de proteasa TEV para producir proteína HTT libre de etiquetas. El estiramiento de poliglutamina y el dominio rico en prolina se diseñaron con sitios de endonucleasa de restricción HindIII flanqueados para insertar repeticiones CAG adicionales utilizando clonación de casetes, es decir , Q48 y Q73, para producir variantes HTT con diferentes longitudes de poliQ. (B) La construcción de ΔExon1 se realizó mutagénesis por PCR utilizando pcDNA3.1-Q23-HTT como plantilla. Los residuos 91-3.144 de HTT permanecieron en el constructo ΔExon1 para su expresión. Abreviaturas: HTT = huntingtina; CMV = citomegalovirus; Q23 = estiramiento de poliglutamina; PRD = dominio rico en prolina; TEV = sitio de escisión del virus del grabado del tabaco. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}

Figura 2: Purificación a gran escala de HTT. (A) Perfil SEC de Q23-HTT de longitud completa purificado anti-Flag en una columna FPLC. Se etiquetan oligómeros de alto orden, dímero y picos de monómeros de Q23-HTT. Las fracciones que contenían monómero se recogieron como la muestra final de HTT. (B) SDS-PAGE de Q23-HTT purificado con paso de lavado de ATP / magnesio (carril 1) o sin lavado de ATP / magnesio da como resultado la coelución de Hsp70 (carril 2). (C) Variantes finales purificadas de HTT de longitud completa en SDS-PAGE teñidas con azul Coomassie G-250 o tinción plateada. Abreviaturas: FL = longitud completa; HTT = huntingtina; SEC = cromatografía de exclusión de tamaño; FPLC = cromatografía líquida de proteínas rápidas; O = oligómero; D = dímero; M = monómero; SDS-PAGE = electroforesis en gel de poliacrilamida dodecilsulfato de sodio; Hsp70 = proteína de choque térmico 70. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}

Figura 3: Análisis de Western blot de variantes de HTT purificadas . (A) El FL Q23-HTT purificado se ejecutó en SDS-PAGE y se transfirió a la membrana de PVDF. Los anticuerpos primarios y los epítopos que interactúan son Lane 1, α-FLAG M2, etiqueta FLAG; Carril 2, MAB5492, HTT aa. 1-82; Carril 3, MAB5490, HTT aa 115-129; Carril 4, MAB2166, HTT aa 181-810; Carril 5, MAB3E10, HTT aa 1,171-1,177; Carril 6, MAB4E10, HTT aa 1,844-2,131; Carril 7, MAB2168, HTT aa 2,146-2,541; Carril 8, MAB8A4, HTT aa 2,703-2,911. (B) Se ejecutó 1 μg de variantes purificadas de FL HTT en SDS-PAGE y se transfirieron a PVDF (izquierda), y se ejecutó un gel SDS duplicado y se tiñó con Coomassie Blue (derecha). Los anticuerpos primarios y los epítopos que interactúan son la fila 1, MW1, repeticiones PolyQ expandidas; Fila 2, MAB2166, HTT aa 181-810; Fila 3, MAB5492, HTT aa 1-82. Abreviaturas: FLL Q23-HTT = proteína huntingtina de longitud completa que contiene 23 residuos de glutamina; SDS-PAGE = electroforesis en gel de poliacrilamida dodecilsulfato de sodio; WB = Western blot; M = marcador; PVDF = fluoruro de polivinilideno. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}

Figura 4: Análisis SEC-MALS de HTT completo. El Q23-HTT purificado de longitud completa se eluyó en una columna UPLC. Se indican las posiciones máximas de monómero, dímero y oligómero previstos. Los pesos moleculares se calcularon para los picos de monómero, dímero y trímero y se enumeraron en la Tabla 5. Se observan perfiles de elución similares para Q48, Q73 y ΔExon1 HTT, con contenidos variables de monómero, dímero y oligómero en cada purificación. Abreviaturas: SEC-MALS = Cromatografía de exclusión de tamaño con dispersión de luz multiángulo; UV = ultravioleta; LS = dispersión de la luz; MW = peso molecular; Q23-HTT = proteína huntingtina que contiene 23 residuos de glutamina; M = monómero; D = dímero; O = oligómero. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}

Figura 5: Caracterización de HTT purificado utilizando gel clear Native PAGE o Blue Native PAGE. El marcador nativo y el Q23-HTT monomérico aparente de SEC se resolvieron en geles de Tris-acetato al 3-8% en un sistema PAGE no desnaturalizante (A) y un sistema Blue Native PAGE (B). Abreviaturas: FL = longitud completa; Q23-HTT = proteína huntingtina que contiene 23 residuos de glutamina; PAGE = electroforesis en gel de poliacrilamida; M = marcador. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}
| Paso | Nombre | Composición | ||
| 6.1.1 | Búfer A | 50 mM Tris, 500 mM NaCl, 5% v/v glicerol, 5 mM EDTA, 0,01% v/v Tween-20, pH 8,0. | ||
| 6.1.2 | Tampón de lisis | 50 mM Tris, 500 mM NaCl, 5% v/v glicerol, 5 mM EDTA y 1x cóctel inhibidor de proteasa | ||
| 6.1.4.2 | Búfer A | 50 mM Tris, 500 mM NaCl, 5% v/v glicerol, 5 mM EDTA, 0,01% v/v Tween-20, pH 8,0. | ||
| 6.1.4.3 | Búfer B | 50 mM Tris; 500 mM KCl; 5 mM MgCl2; 5% v/v glicerol; 0.01% v/v Tween-20, pH 8.0 | ||
| 6.1.4.4 | Búfer C | 20 mM Tris; 200 mM KCl; 5 mM MgCl2; 5 mM ATP; 0,01% v/v Tween-20; 5% v/v glicerol, pH 8.0 | ||
| 6.1.4.5 | Búfer D | 50 mM Tris; 500 mM NaCl; 5% v/v glicerol; 5 mM EDTA; 0,5% p/v CHAPS, pH 8,0 | ||
| 6.1.4.6 | Tampón de elución | 50 mM Tris; 500 mM NaCl; 5% v/v glicerol; 0,5% p/v CHAPS; 0,2 mg/ml de péptido DYKDDDDK, pH 8,0 | ||
| 6.1.6 | Búfer de regeneración | 0,1 M glicina HCl, pH 3,5; 0.01% v/v Tween-20 | ||
| 6.2.1 | Búfer SEC | 50 mM Tris, 500 mM NaCl, 5% v/v glicerol, 0,5% p/v CHAPS, 1 mM TCEP | ||
| 7.3 | Búfer SEC-MALS | 50 mM HEPES, pH 7.2, 500 mM NaCl, 5% v/v glicerol, 0.5% p/v CHAPS | ||
| 8.7 | Solución decolorante | 40% v/v metanol y 7% v/v ácido acético | ||
| 9.4.2 | Solución de tinción Coomassie | 0.01% p/v Coomassie G250, 50% v/v/ metanol, 10% v/v ácido acético | ||
| 9.5.1 | Solución de reparación | 50% v/v metanol, 10% v/v ácido acético, 50 μL de formaldehído/100 ml de solución | ||
Tabla 1: Composición de tampones y soluciones
| Pasos | Concentración de HTT (mg/ml) | Volumen total (ml) | Contenido de HTT (mg) | Rendimiento HTT por celda (pg/celda) | % Rendimiento |
| Sobrenadante | 0.1792 | 220 | 39.4 | 4.4 | 100 |
| Anti-Bandera | 1.524 | 8.6 | 13.1 | 1.47 | 33.4 |
| SEC | 0.91 | 3.9 | 3.54 | 0.4 | 9.1 |
Tabla 2: Rendimiento de HTT de un pellet HEK293 de 2 L transfectado con pcDNA3.1-Q23-HTT-TEV-Flag. Abreviaturas: FL Q23-HTT = proteína huntingtina de longitud completa que contiene 23 residuos de glutamina; TEV = sitio de escisión del virus del grabado del tabaco; SEC = cromatografía de exclusión de tamaño.
| Ejemplo HTT | Rendimiento HTT (mg/L) | Pureza media (%) | ||
| BCA | A280 | |||
| 1 | FL DEx1-HTT (N=3) | 0.67-1.30 | 0.69-1.18 | 99.3 |
| 2 | FL Q23-HTT (N=3) | 0.25-0.92 | 0.28-0.98 | 96.9 |
| 3 | FL Q48-HTT (N=3) | 0.28-1.15 | 0.38-1.16 | 97.4 |
| 4 | FL Q73-HTT (N=3) | 0.58-1.05 | 0.57-0.97 | 98.8 |
Tabla 3: Resumen del rendimiento proteico de cuatro purificaciones de variantes FL HTT y su pureza final. Abreviatura: FLL HTT = proteína huntingtina de longitud completa.
| Ejemplo HTT | Un | D | M |
| FL Q23-HTT | 4.2-6.9% | 18.7-29.3% | 66.5-76.0% |
| FL Q48-HTT | 4.0-9.4% | 10.6-17.8% | 73.6-85.4% |
| FL Q73-HTT | 2.0-14.0% | 16.9-24.6% | 65.1-81.1% |
Tabla 4: Resumen del contenido representativo de agregados, dímeros y monómeros de las variantes de FL HTT de la purificación. Abreviaturas: FL HTT = proteína huntingtina de longitud completa; A = agregado; D = dímero; M = monómero; SEC = cromatografía de exclusión de tamaño.
Figura suplementaria S1: Análisis de Western blot después de la digestión de la proteasa TEV. Los FL Q23-HTT y FL Q48-HTT purificados se ejecutaron en SDS-PAGE, se transfirieron a membranas de PVDF y se analizaron mediante Western blot después del digest TEV. Los anticuerpos primarios utilizados fueron anti-Flag M2 (etiqueta Flag), MAB5492 (HTT aa 1-82), MAB3E10 (HTT aa 997-1,276) y MAB2168 (HTT aa 2,146-2,541). Carril 1, Estándar de Proteína; Carril 2, Q23-HTT-TEV-Flag; Carril 3, Q48-HTT-TEV-Flag; Carril 4, Q23-HTT-TEV-Flag tratado con proteasa TEV a 1:5, durante la noche a 4 °C; Carril 5, Q48-HTT-TEV-Flag tratado con proteasa TEV a 1:5, durante la noche a 4 °C. Abreviaturas: FL HTT = proteína huntingtina de longitud completa; SDS-PAGE = electroforesis en gel de poliacrilamida dodecilsulfato de sodio; TEV = virus del grabado del tabaco; PVDF = fluoruro de polivinilideno. Haga clic aquí para descargar este archivo.
Figura suplementaria S2: Análisis SEC-MALS de variantes de FL HTT sometidas a ciclos de congelación-descongelación. Los Q23-HTT (A) y Q48-HTT (B) purificados se congelaron a -80 °C y se descongelaron a temperatura ambiente hasta 6 veces. Q23-HTT y Q48-HTT después del primer ciclo de congelación-descongelación y sexto ciclo de congelación-descongelación fueron analizados por SEC-MALS. Se observó una ligera disminución en la fracción de monómeros y un aumento en las fracciones de dímeros y oligómeros de orden alto por dispersión de luz después de ciclos repetidos de congelación-descongelación. Se indican las posiciones máximas de monómero, dímero y oligómero de orden alto predichos. Abreviaturas: FL HTT = proteína huntingtina de longitud completa; O = oligómero; D = dímero; M = monómero; SEC-MALS = Cromatografía de exclusión de tamaño con dispersión de luz multiángulo. Haga clic aquí para descargar este archivo.
Figura suplementaria S3: PÁGINA DE SDS de las variantes FL HTT sometidas a ciclos de congelación-descongelación. Los Q23-HTT purificados (carriles 2-7) y Q48-HTT (carriles 9-14) se congelaron a -80 °C y se descongelaron a temperatura ambiente hasta 6 veces. Las alícuotas de Q23-HTT y Q48-HTT se almacenaron después de cada ciclo de congelación-descongelación y luego se analizaron mediante SDS PAGE. No se observó ningún aumento de los productos agregados o de degradación; Las muestras se consideraron estables y >95% puras por densitometría de banda. Abreviaturas: FL HTT = proteína huntingtina de longitud completa; SDS-PAGE = electroforesis en gel de poliacrilamida dodecilsulfato de sodio. Haga clic aquí para descargar este archivo.
Archivo complementario 1: script HTT anti-FLAG FPLC de 15 ml. Abreviaturas = FPLC = cromatografía líquida de proteínas rápidas; HTT = proteína huntingtina. Haga clic aquí para descargar este archivo.
Archivo complementario 2: script FPLC SEC_MALS HTT. Abreviaturas: SEC-MALS = Cromatografía de exclusión de tamaño con dispersión de luz multiángulo; FPLC = cromatografía líquida de proteínas rápidas; HTT = proteína huntingtina. Haga clic aquí para descargar este archivo.
Discusión
Describimos aquí un método transitorio de transfección, expresión y purificación para generar múltiples construcciones de proteína FL HTT con pureza y homogeneidad adecuadas para su uso como estándares para el desarrollo de inmunoensayo y ensayo de EM, controles para análisis de Western blot y para estudios de estructura-función. Este método de expresión transitoria es escalable y versátil y permite al usuario generar cantidades bajas de miligramos de variantes de FL HTT de manera más eficiente que el uso de líneas celulares estables o métodos basados en virus descritos anteriormente21,22,23,24. Rutinariamente, se pueden generar 2-5 mg de FL HTT altamente purificado a partir de una producción de proteína a escala de 2 L en menos de una semana utilizando el método de expresión transitoria una vez que se construye el plásmido, con un rendimiento típico de 1-2.5 mg de FL HTT por litro de cultivo celular.
El método de expresión transitoria descrito aquí supera muchos obstáculos en la expresión de líneas celulares estables, como el largo tiempo necesario para establecer líneas celulares y las dificultades para almacenar y mantener líneas celulares estables. PEI también es relativamente barato en comparación con otros reactivos de transfección en el mercado, lo que hace que la transfección a gran escala sea económicamente viable. También hay limitaciones en el protocolo: la eficiencia de la transfección depende en gran medida de la calidad de los plásmidos, el crecimiento celular óptimo y qué tan bien se almacena y prepara la PEI. Los operadores deben tener especial cuidado y realizar controles de calidad en esos pasos críticos para evitar una caída drástica en los rendimientos de proteínas. La resina anti-FLAG utilizada en el protocolo también es relativamente costosa y muestra una captura reducida de FL HTT después de varias purificaciones y regeneraciones. Algunos investigadores pueden encontrar más práctico cambiar a una etiqueta diferente para permitir una regeneración más robusta de la resina de afinidad.
Se probaron varias líneas celulares y condiciones de expresión para optimizar los niveles de expresión de FL HTT. Las células HEK293 fueron elegidas para la expresión de FL HTT debido a la alta expresión de proteínas y la facilidad de manejo en un formato de cultivo en suspensión, lo que hace que el método sea adecuado para la expresión a gran escala en agitadores o biorreactores. Se puede lograr un nivel más alto de expresión de proteína FL HTT a temperaturas de cultivo más bajas, como 32 ° C, en lugar de usar la temperatura habitual de 37 ° C. Es posible que la temperatura más baja pueda ralentizar la síntesis de proteínas y promover el plegamiento correcto de FL HTT40. Sin embargo, este fenómeno no es específico de FL HTT o de las líneas celulares probadas. La temperatura reducida posterior a la transfección se ha utilizado ampliamente en la expresión de proteínas farmacéuticas en células CHO. Aunque el mecanismo no se comprende completamente, se cree que las bajas temperaturas detienen el ciclo celular en la fase G1 y desvían la energía celular a la producción de proteínas41.
La HTT de longitud completa purificada a partir de células de mamíferos co-eluye con la chaperona Hsp7024, y los pasos de lavado de Mg-ATP pueden eliminar la proteína Hsp70. Curiosamente, el Hsp70 co-eluyente no se observa en FL HTT purificado de un sistema de expresión celular de insectos21,22,23. Esto puede reflejar una diferencia en los PTM de FL HTT o respuestas de proteínas de choque térmico a la sobreexpresión de FL HTT en células de mamíferos e insectos. Una vez que la proteína recombinante ha sido despojada de Hsp70, se requieren detergentes no iónicos como CHAPS o DDM para estabilizar la forma monomérica de FL HTT.
Los estados de oligomerización de las variantes FL HTT se analizaron utilizando Blue Native PAGE y SEC-MALS. Una pequeña fracción de HTT dimérico y oligomérico de orden superior estaba presente cuando fue analizado por Blue Native PAGE o SEC-MALS. Cabe destacar que los oligómeros de orden superior formados por FL HTT no parecen correlacionarse con la longitud del poliQ, e incluso el mutante de deleción Exon1 muestra una relación oligómero-dímero-monómero similar. Las variaciones reales en el contenido de oligómeros entre estas construcciones probablemente se deban a pequeñas diferencias en la producción y manejo de cada lote. En contraste con los agregados y fibrillas formados por HTT Exon140,41, los oligómeros de orden superior de FL HTT permanecieron solubles y pudieron ser analizados por SEC y Native PAGE.
El FL HTT monomérico purificado es solo relativamente estable. El almacenamiento prolongado a 4 °C, las incubaciones cortas a temperatura ambiente o concentraciones > 1 mg/ml convertirán la FL HTT monomérica en formas diméricas y oligoméricas de orden superior aunque no se observe precipitación visible en esas condiciones. La FL HTT monomérica purificada mantenida a ≤1 mg/mL se mantuvo relativamente estable a -80 °C en tampón de almacenamiento (50 mM Tris, pH 8,0, 500 mM NaCl, 5% v/v glicerol, 0,5% p/v CHAPS y 5 mM DTT) como se describió anteriormente24. Hasta 6 ciclos de congelación-descongelación de FL HTT preparados y almacenados de esta manera no causaron precipitación visible de la proteína, aunque SEC-MALS observó un ligero cambio a un estado oligomérico más alto (Figura Suplementaria S2). Las muestras también fueron analizadas por SDS PAGE después de repetidos ciclos de congelación-descongelación. No se observaron precipitados visibles; no se observaron agregados ni productos de degradación adicionales en SDS-PAGE (Figura Suplementaria S3). La estabilidad a largo plazo de FL HTT purificado todavía está bajo investigación. En ausencia de datos concluyentes a largo plazo, recomendamos almacenar FL HTT purificado a -80 °C durante no más de 6 meses.
Las variantes de proteína FL HTT recombinante de alta calidad y los métodos para producirlas tienen una gran demanda por parte de la comunidad de investigación de la EH. Estas proteínas se utilizan como inmunoensayo y estándares analíticos de EM, en estudios estructurales y para el desarrollo de nuevos ensayos específicos de FL HTT. Los métodos de expresión transitoria a gran escala descritos aquí han producido consistentemente cantidades de miligramos de variantes de FL HTT con una pureza del >95%, proporcionando herramientas esenciales para los estudios de HTT. La producción de decenas de miligramos de variantes poliQ FL HTT altamente purificadas y otros mutantes en apoyo de la investigación de la EH se ha convertido en rutina.
Divulgaciones
Los autores declaran que no tienen conflictos de intereses con el contenido de este artículo.
Agradecimientos
Agradecemos al Departamento de Ciencias Farmacéuticas de la Universidad Estatal de Nueva York en Buffalo por realizar el análisis de EM de HTT. Este trabajo fue un esfuerzo de colaboración con la Fundación CHDI. Agradecemos específicamente a Elizabeth M. Doherty; Ignacio Muñoz-Sanjuan; Douglas Macdonald, Fundación CHDI; y Rory Curtis, Curia, por su inestimable aporte durante la preparación de este manuscrito. También agradecemos a Michele Luche, Mithra Mahmoudi y Stephanie Fox por su apoyo a este esfuerzo de investigación.
Materiales
| Name | Company | Catalog Number | Comments |
| 100 kDa concentrator-Amicon | Millipore | UFC910096 | Protocol Section Number-6.2.4 |
| 20x blue native PAGE running buffer | Invitrogen | BN2001 | Protocol Section Number-8.1 |
| 20x TBS | Thermo Fisher | PI28358 | Protocol Section Number-5.1 |
| 4x blue native PAGE sample buffer | Invitrogen | BN2003 | Protocol Section Number-8.3 |
| 4x LDS loading buffer | Invitrogen | NP0007 | Protocol Section Number-5.3 |
| 5 L Erlenmeyer flasks | Corning | 431685 | Protocol Section Number-4.2 |
| Agarose gel extraction kit | Qiagen | 28704 | Protocol Section Number-2.2 |
| Anti-clumping agent | Thermo Fisher | 0010057AE | Protocol Section Number-4.8 |
| anti-FLAG M2 affinity gel | Sigma | A2220 | Protocol Section Number-6.1.1 |
| anti-FLAG M2 | Sigma | F3165 | Protocol Section Number-5.7 |
| Anti foam-Excell anti foam | Sigma | 59920C-1B | Protocol Section Number-4.8 |
| ATP | Sigma | A6419 | Protocol Section Number-6.1.4.4 |
| BEH 450 SEC | Waters | 186006851 | 2.5 µm x 4.6 mm x 150 mm Protocol Section Number-7.3 |
| blue native PAGE 5% G-250 sample additive | Invitrogen | BN2004 | Protocol Section Number-8.3 |
| carbenicillin | Thermo Fisher | 10177012 | Protocol Section Number-2.5 |
| centrifuge - Sorvall Lynx 6000 | Thermo Fisher | 75006590 | Protocol Section Number-6.1.3 |
| Cell Counter - ViCELL | BECKMAN COULTER | Protocol Section Number-4.3 | |
| CHAPS | Anatrace | C316S | Protocol Section Number-6.1.4.6 |
| Competent E. coli cells-TOP10 | Invitrogen | C404010 | Protocol Section Number-2.4 |
| digitonin | Sigma | D141 | Protocol Section Number-5.1 |
| differential refractive index detector | Wyatt | Protocol Section Number-7.1 | |
| DYKDDDDK peptide | Genscript | Peptide synthesis service Protocol Section Number-6.1.4.6 | |
| EDTA | Sigma | EDS | Protocol Section Number-5.1 |
| EndoFree Plasmid Giga Kit | Qiagen | 12391 | Protocol Section Number-3.3 |
| Endotoxin free water | Cytiva | SH30529.03 | Protocol Section Number-4.1 |
| endotoxin quantification kit-CRL Endosafe Nexgen-PTS detection system | Charles River | PTS150K | Protocol Section Number-3.4 |
| fixed angle rotor A23-6x100 rotor | Thermo Fisher | 75003006 | Protocol Section Number-6.1.3 |
| FPLC software- Unicorn 6.2 | Cytiva | Protocol Section Number-6.1.4 | |
| Gene synthesis | Genscript | Gene synthesis service Protocol Section Number-1.2 | |
| Glycerol | Fisher Scientific Glycerol (Certified ACS) | G33-4 | Protocol Section Number-5.6 |
| Growth Medium-Expi293 expression medium | Thermo Fisher | A1435102 | Protocol Section Number-4.2 |
| HEK293 cells | Thermo Fisher | R79007 | Protocol Section Number-4 |
| high shear homogenizer-Microfluidizer | MicroFluidics | LM10 | Protocol Section Number-6.1.3 |
| HPLC - 1260 infinity II Bio-Insert HPLC | Agilent | Protocol Section Number-7.1 | |
| Image Studio | LiCor | Image analysis software Protocol Section Number-5.1 | |
| MAB2166 | Sigma | MAB2166 | Protocol Section Number-5.7 |
| MAB2168 | EMD | MAB2168 | Protocol Section Number-5.7 |
| MAB3E10 | Santa Cruz | SC-47757 | Protocol Section Number-5.7 |
| MAB4E10 | Santa Cruz | SC-7757 | Protocol Section Number-5.7 |
| MAB5490 | Sigma | MAB5490 | Protocol Section Number-5.7 |
| MAB5492 | Sigma | MAB5492 | Protocol Section Number-5.7 |
| MAB8A4 | Santa Cruz | SC-47759 | Protocol Section Number-5.7 |
| multi-angle light scattering detector | Wyatt | Protocol Section Number-7.1 | |
| NativeMark Unstained Protein Standard | Invitrogen | LC0725 | Protocol Section Number-8.4 |
| NaCl | Sigma | S9888 | Protocol Section Number-5.6 |
| NheI | New England Biolab | R0131S | Hi-Fi version available Protocol Section Number-2.2 |
| NuPAGE 3–8% Tris acetate gels | Invitrogen | EA0375PK2 | Protocol Section Number-5.4 |
| NuPAGE Tris-Acetate SDS Running buffer | Invitrogen | LA0041 | Protocol Section Number-5.4 |
| PEI 25K | Polysciences | 23966-1 | Protocol Section Number-4.1 |
| Penicillin-Streptomycin | Thermo Fisher | 15070063 | Protocol Section Number-4.2 |
| Phosphate Buffered Saline (PBS) | Cytiva | SH30256.02 | Protocol Section Number-4.5 |
| plasmid miniprep kit | Qiagen | 27104 | Protocol Section Number-2.6 |
| PmeI | New England Biolab | R0560S | Protocol Section Number-2.2 |
| precast Bis-tris gel- 3-12% NativePAGE Novex Bis-Tris Gel | Invitrogen | BN1003BOX | Protocol Section Number-8.4 |
| protease inhibitor cocktail | GoldBio | GB-331-1 | Protocol Section Number-5.1 |
| SEC-MALS analysis software - Astra 7 | Wyatt Technology | Protocol Section Number-7.6 | |
| secondary antibody -IRdye 800 CW goat anti-mouse IgG | LiCor | 926-32210 | Protocol Section Number-5.9 |
| Superose 6 pg XK 16/70 | Cytiva | 90100042 | Protocol Section Number-6.2 |
| Tris base | Fisher | BP152 | Protocol Section Number-5.6 |
| Tween-20 | Thermo Fisher | AAJ20605AP | Protocol Section Number-6.1.1 |
| UV spectrometer - Nanodrop 8000 | Thermo Fisher | ND-8000-GL | Protocol Section Number-2.2 |
| XK26/100 | Cytiva | 28988951 | Protocol Section Number-6.1.1 |
Referencias
- Walker, F. O. Huntington's disease. Lancet. 369 (9557), 218-228 (2007).
- McColgan, P., Tabrizi, S. J. Huntington's disease: a clinical review. European Journal of Neurology. 25 (1), 24-34 (2018).
- Duyao, M., et al. Trinucleotide repeat length instability and age of onset in Huntington's disease. Nature Genetics. 4 (4), 387-392 (1993).
- MacDonald, M. E., et al. A novel gene containing a trinucleotide repeat that is expanded and unstable on Huntington's disease chromosomes. Cell. 72 (6), 971-983 (1993).
- Nasir, J., et al. Targeted disruption of the Huntington's disease gene results in embryonic lethality and behavioral and morphological changes in heterozygotes. Cell. 81 (5), 811-823 (1995).
- Dragatsis, I., Levine, M. S., Zeitlin, S. Inactivation of Hdh in the brain and testis results in progressive neurodegeneration and sterility in mice. Nature Genetics. 26 (3), 300-306 (2000).
- Anne, S. L., Saudou, F., Humbert, S. Phosphorylation of huntingtin by cyclin-dependent kinase 5 is induced by DNA damage and regulates wild-type and mutant huntingtin toxicity in neurons. Journal of Neuroscience. 27 (27), 7318-7328 (2007).
- Dietrich, P., Johnson, I. M., Alli, S., Dragatsis, I. Elimination of huntingtin in the adult mouse leads to progressive behavioral deficits, bilateral thalamic calcification, and altered brain iron homeostasis. PLoS Genetics. 13 (7), 1006846(2017).
- Dragatsis, I., et al. Effect of early embryonic deletion of huntingtin from pyramidal neurons on the development and long-term survival of neurons in cerebral cortex and striatum. Neurobiology of Disease. 111, 102-117 (2018).
- Benn, C. L., et al. Huntingtin modulates transcription, occupies gene promoters in vivo, and binds directly to DNA in a polyglutamine-dependent manner. Journal of Neuroscience. 28 (42), 10720-10733 (2008).
- Saudou, F., Humbert, S. The biology of huntingtin. Neuron. 89 (5), 910-926 (2016).
- Davies, S. W., et al. Formation of neuronal intranuclear inclusions underlies the neurological dysfunction in mice transgenic for the HD mutation. Cell. 90 (3), 537-548 (1997).
- DiFiglia, M., et al. Aggregation of huntingtin in neuronal intranuclear inclusions and dystrophic neurites in brain. Science. 277 (5334), 1990-1993 (1997).
- Gutekunst, C. A., et al. Nuclear and neuropil aggregates in Huntington's disease: Relationship to neuropathology. Journal of Neuroscience. 19 (7), 2522-2534 (1999).
- Hodgson, J. G., et al. A YAC mouse model for Huntington's disease with full-length mutant huntingtin, cytoplasmic toxicity, and selective striatal neurodegeneration. Neuron. 23 (1), 181-192 (1999).
- Hoffner, G., Djian, P. Polyglutamine aggregation in Huntington disease: does structure determine toxicity. Molecular Neurobiology. 52 (3), 1297-1314 (2015).
- Waldvogel, H. J., Kim, E. H., Tippett, L. J., Vonsattel, J. P. G., Faull, R. L. M. The neuropathology of Huntington's disease. Current Topics in Behavioral Neurosciences. 22, 33-80 (2014).
- Kim, M. Beta conformation of polyglutamine track revealed by a crystal structure of huntingtin N-terminal region with insertion of three histidine residues. Prion. 7 (3), 221-228 (2013).
- Hoop, C. L., et al. Huntingtin exon 1 fibrils feature an interdigitated β-hairpin-based polyglutamine core. Proceedings of the National Academy of Sciences of the United States of America. 113 (6), 1546-1551 (2016).
- Vieweg, S., Ansaloni, A., Wang, Z. M., Warner, J. B., Lashuel, H. A. An intein-based strategy for the production of tag-free huntingtin exon 1 proteins enables new insights into the polyglutamine dependence of Httex1 aggregation and fibril formation. Journal of Biological Chemistry. 291 (23), 12074-12086 (2016).
- Seong, I. S., et al. Huntingtin facilitates polycomb repressive complex 2. Human Molecular Genetics. 19 (4), 573-583 (2009).
- Li, W., Serpell, L. C., Carter, W. J., Rubinsztein, D. C., Huntington, J. A. Expression and characterization of full-length human huntingtin, an elongated HEAT repeat protein. Journal of Biological Chemistry. 281 (23), 15916-15922 (2006).
- Vijayvargia, R., et al. Huntingtin's spherical solenoid structure enables polyglutamine tract-dependent modulation of its structure and function. eLife. 5, 11184(2016).
- Huang, B., et al. Scalable production in human cells and biochemical characterization of full-length normal and mutant huntingtin. PLoS ONE. 10 (3), 0121055(2015).
- Guo, Q., et al. The cryo-electron microscopy structure of huntingtin. Nature. 555 (7694), 117-120 (2018).
- Harding, R. J., et al. Design and characterization of mutant and wildtype huntingtin proteins produced from a toolkit of scalable eukaryotic expression systems. Journal of Biological Chemistry. 294 (17), 6986-7001 (2019).
- Harding, R. J., et al. HAP40 orchestrates huntingtin structure for 1 differential interaction with polyglutamine 2 expanded exon 1. bioRxiv. , (2021).
- Huang, B., et al. Pathological polyQ expansion does not alter the conformation of the Huntingtin-HAP40 complex. Structure. 29 (8), 804-809 (2021).
- Colin, E., et al. Huntingtin phosphorylation acts as a molecular switch for anterograde/retrograde transport in neurons. EMBO Journal. 27 (15), 2124-2134 (2008).
- Thompson, L. M., et al. IKK phosphorylates Huntingtin and targets it for degradation by the proteasome and lysosome. Journal of Cell Biology. 187 (7), 1083-1099 (2009).
- Ratovitski, T., et al. Post-translational modifications (PTMs), identified on endogenous Huntingtin, cluster within proteolytic domains between HEAT repeats. Journal of Proteome Research. 16 (8), 2692-2708 (2017).
- Taylor, S. C., Berkelman, T., Yadav, G., Hammond, M. A defined methodology for reliable quantification of western blot data. Molecular Biotechnology. 55 (3), 217-226 (2013).
- Tarazona, M. P., Saiz, E. Combination of SEC/MALS experimental procedures and theoretical analysis for studying the solution properties of macromolecules. Journal of Biochemical and Biophysical Methods. 56 (1-3), 95-116 (2003).
- Folta-Stogniew, E. Oligomeric states of proteins determined by size-exclusion chromatography coupled with light scattering, absorbance, and refractive index detectors. Methods in Molecular Biology. 328, Clifton, N.J. 97-112 (2006).
- McMeekin, T. L., Wilensky, M., Groves, M. L. Refractive indices of proteins in relation to amino acid composition and specific volume. Biochemical and Biophysical Research Communications. 7 (2), 151-156 (1962).
- Atwal, R. S., et al. Huntingtin has a membrane association signal that can modulate huntingtin aggregation, nuclear entry and toxicity. Human Molecular Genetics. 16 (21), 2600-2615 (2007).
- Kegel-Gleason, K. B. Huntingtin interactions with membrane phospholipids: Strategic targets for therapeutic intervention. Journal of Huntington's Disease. 2 (3), 239-250 (2013).
- Michalek, M., Salnikov, E. S., Werten, S., Bechinger, B. Membrane interactions of the amphipathic amino terminus of huntingtin. Biochemistry. 52 (5), 847-858 (2013).
- Wittig, I., Braun, H. P., Schägger, H. Blue native PAGE. Nature Protocols. 1 (1), 418-428 (2006).
- Nissley, D. A., O'Brien, E. P. Altered co-translational processing plays a role in huntington's pathogenesis-A hypothesis. Frontiers in Molecular Neuroscience. 9, 54(2016).
- Kumar, N., Gammell, P., Clynes, M. Proliferation control strategies to improve productivity and survival during CHO based production culture: A summary of recent methods employed and the effects of proliferation control in product secreting CHO cell lines. Cytotechnology. 53 (1-3), 33-46 (2007).
Reimpresiones y Permisos
Solicitar permiso para reutilizar el texto o las figuras de este JoVE artículos
Solicitar permisoExplorar más artículos
This article has been published
Video Coming Soon
ACERCA DE JoVE
Copyright © 2025 MyJoVE Corporation. Todos los derechos reservados