
Method Article
Ensayos in vitro estandarizados para visualizar y cuantificar las interacciones entre neutrófilos humanos y biopelículas de Staphylococcus aureus
En este artículo
Resumen
El presente protocolo describe el estudio de las interacciones neutrófilos-biofilm. Las biopelículas de Staphylococcus aureus se establecen in vitro y se incuban con neutrófilos humanos derivados de sangre periférica. La respuesta de explosión oxidativa de los neutrófilos se cuantifica, y la localización de neutrófilos dentro de la biopelícula se determina mediante microscopía.
Resumen
Los neutrófilos son la primera línea de defensa desplegada por el sistema inmune durante la infección microbiana. In vivo, los neutrófilos son reclutados al sitio de la infección donde utilizan procesos como la fagocitosis, la producción de especies reactivas de oxígeno y nitrógeno (ROS, RNS, respectivamente), NETosis (trampa extracelular de neutrófilos) y la desgranulación para matar microbios y resolver la infección. Las interacciones entre neutrófilos y microbios planctónicos han sido ampliamente estudiadas. Ha habido intereses emergentes en el estudio de las infecciones causadas por biopelículas en los últimos años. Las biopelículas exhiben propiedades, incluida la tolerancia a la muerte por neutrófilos, distintas de sus contrapartes cultivadas planctónicamente. Con el establecimiento exitoso de modelos de biopelículas in vitro e in vivo , ahora se pueden investigar las interacciones entre estas comunidades microbianas con diferentes células inmunes. Aquí, las técnicas que utilizan una combinación de modelos tradicionales de biopelícula y ensayos de actividad de neutrófilos bien establecidos se adaptan específicamente para estudiar las interacciones de neutrófilos y biopelículas. La microscopía de fluorescencia de campo amplio se utiliza para monitorear la localización de neutrófilos en biopelículas. Estas biopelículas se cultivan en condiciones estáticas, seguidas de la adición de neutrófilos derivados de la sangre periférica humana. Las muestras se tiñen con tintes apropiados antes de la visualización bajo el microscopio. Además, la producción de ROS, que es una de las muchas respuestas de neutrófilos contra patógenos, se cuantifica en presencia de una biopelícula. La adición de células inmunes a este sistema establecido ampliará la comprensión de las interacciones huésped-patógeno al tiempo que garantiza el uso de condiciones estandarizadas y optimizadas para medir estos procesos con precisión.
Introducción
Una biopelícula es una comunidad de microbios asociados a la superficie o agregados no unidos encerrados en una sustancia polimérica extracelular (EPS)1,2. Estas comunidades protegen los microorganismos encerrados de los factores estresantes ambientales, incluida la tolerancia a los agentes antimicrobianos y al sistema inmunológico3. Varias especies microbianas patógenas forman biofilms que han sido asociados con infecciones crónicas4. El desarrollo de biopelículas es un proceso intrincado que implica la fijación a superficies, la producción de EPS, la proliferación celular, la estructuración de biopelículas y el desprendimiento celular5. Una vez que las células se dispersan para formar una biopelícula, permanecen planctónicas o se translocan a un nuevo sustrato y reinician el desarrollo de la biopelícula6.
Staphylococcus aureus, un patógeno oportunista, sigue un esquema general de desarrollo de biopelículas, incluyendo adhesión, proliferación, maduración y dispersión7. El proceso de unión en biopelículas de S. aureus está dictado por interacciones hidrofóbicas, ácidos teicoicos y componentes de superficie microbiana que reconocen moléculas de matriz adhesiva (MSCRAMMs)8,9. A medida que comienza la proliferación de S. aureus, se produce EPS, que consiste principalmente en polisacáridos, proteínas, ADN extracelular y ácidos teicoicos5. A medida que se producen componentes EPS, también se producen varias exoenzimas y moléculas pequeñas, lo que contribuye a la estructura 3-dimensional de la biopelícula y ayuda en el desprendimiento5. S. aureus aprovecha este estilo de vida altamente coordinado para establecer diversas infecciones crónicas, incluyendo infecciones debidas a la permanencia de dispositivos médicos10.
El S. aureus resistente a la meticilina (SARM) es una de las principales causas de infecciones relacionadas con dispositivos médicos permanentes, como catéteres venosos y urinarios centrales, articulaciones protésicas, marcapasos, válvulas cardíacas mecánicas y dispositivos intrauterinos11. Durante tales infecciones, los neutrófilos son las primeras células inmunes del huésped reclutadas en el sitio de la infección para combatir los patógenos a través de múltiples estrategias12. Estos incluyen fagocitosis, desgranulación, producción reactiva de especies de oxígeno y nitrógeno (ROS/RNS) o liberación de trampas extracelulares de neutrófilos (TNE) para eliminar patógenos13.
La generación de ROS tras la fagocitosis de microbios es una de las respuestas antimicrobianas clave exhibidas por los neutrófilos14. La fagocitosis aumenta si los microbios están recubiertos de opsoninas, particularmente inmunoglobulinas y componentes del complemento que se encuentran en el suero15. Los microbios opsonizados son reconocidos por los receptores de la superficie celular en los neutrófilos y engullidos, formando un compartimento llamado fagosoma15. Los neutrófilos generan y liberan ROS en el fagosoma a través de la NADPH-oxidasa16 asociada a la membrana. Este complejo enzimático multicomponente genera aniones superóxido mediante la transferencia de electrones al oxígeno molecular16. Además, los neutrófilos también generan RNS a través de la expresión de óxido nítrico sintasa inducible (iNOS)17. Estos radicales con alto superóxido y óxido nítrico dentro del fagosoma tienen amplias actividades antimicrobianas. Pueden interactuar con centros metálicos en enzimas y dañar ácidos nucleicos, proteínas y membranas celulares del patógeno 18,19,20,21. Numerosos microbios adoptan un estilo de vida de biopelícula y emplean diferentes estrategias para evadir la muerte por ROS22,23. Por lo tanto, los ensayos estandarizados que acoplan biopelículas con neutrófilos para cuantificar ROS son beneficiosos para obtener resultados consistentes.
Mientras que los ensayos, como la cuantificación de la producción de ROS de neutrófilos, proporcionan información sobre las respuestas de los neutrófilos a las biopelículas, la capacidad de visualizar las interacciones de los neutrófilos dentro de una biopelícula también puede servir como una herramienta poderosa. El uso de colorantes fluorescentes para microscopía a menudo requiere optimización para obtener imágenes de alta calidad que se pueden utilizar para el análisis de imágenes de microscopía. La flexibilidad para optimizar algunas condiciones es limitada, ya que los neutrófilos pueden sufrir la muerte celular después del aislamiento. Además, las biopelículas generalmente se lavan para eliminar la población planctónica de la configuración experimental antes de la adición de neutrófilos. Durante el lavado, la variabilidad entre las biopelículas replicadas puede surgir debido a la pérdida de biomasa parcial si las biopelículas se adhieren libremente a la superficie.
En términos generales, los métodos actuales en el campo para analizar las interacciones entre neutrófilos y biopelículas incluyen principalmente microscopía, citometría de flujo y enumeración de unidades formadoras de colonias (UFC)24,25,26,27. La microscopía implica el uso de colorantes que tiñen directamente los neutrófilos y las biopelículas, o se dirigen a diversas respuestas de neutrófilos contra microbios como la formación de NET, la desgranulación y la muerte celular25,28. Un subconjunto de estas respuestas, como la muerte celular de neutrófilos y la desgranulación, también puede analizarse mediante citometría de flujo, pero requiere que los neutrófilos no se asocien preferentemente con grandes agregados de microbios en una biopelícula28,29. La citometría de flujo también puede cuantificar algunos parámetros del biofilm, como la viabilidad celular27. Estos procesos, sin embargo, requieren la interrupción de la biomasa de biofilm y no serían útiles para visualizar otras interacciones importantes como la distribución espacial de los neutrófilos y sus componentes dentro de un biofilm27,29,30.
El presente protocolo se centra en adaptar algunos de los métodos utilizados tradicionalmente para estudiar las interacciones neutrófilos-biofilm en biofilms que han sido optimizados para proporcionar una variabilidad mínima durante la manipulación. Por lo tanto, este protocolo proporciona métodos estandarizados para cultivar y cuantificar biopelículas, aislar neutrófilos humanos primarios de sangre periférica, cuantificar la producción de ROS y visualizar las interacciones biopelícula-neutrófilos a través de microscopía. Este protocolo se puede adaptar a diferentes sistemas para comprender las interacciones biopelícula-neutrófilos teniendo en cuenta la heterogeneidad entre los grupos de donantes.
Protocolo
Todos los procedimientos fueron aprobados por la Junta de Revisión Institucional de la Universidad Estatal de Ohio (IRB) (2014H0154). Se obtuvo el consentimiento informado por escrito de todos los donantes para recolectar sangre periférica para aislar neutrófilos humanos primarios. Staphylococcus aureus (USA300 LAC)31 se utilizó como organismo modelo para realizar los experimentos. Los experimentos se realizaron con el equipo de protección personal (EPP) adecuado debido a la posible exposición a un patógeno transmitido por la sangre.
1. Preparación del biofilm in vitro
- Obtener colonias aisladas de S. aureus a partir de un stock criopreservado 31 utilizando una técnica de placa rayada32,33 en una placa de agar rica en nutrientes, como el agar tríptico de soja (ver Tabla de materiales).
- Recubrir los pocillos individuales de una placa de 96 pocillos con 100 μL de poli-L-lisina (PLL) al 0,001% (v/v) diluida enH2Oestéril e incubar a temperatura ambiente durante 30 min. Asépticamente, aspire la solución de PLL utilizando una trampa de aspiración asistida por vacío. Deje que los pocillos se sequen durante la noche a temperatura ambiente.
NOTA: Todos los pasos de aspiración en el protocolo se realizan utilizando una trampa de aspiración asistida por vacío a menos que se indique lo contrario. - Preparar un cultivo nocturno inoculando una colonia de S. aureus en medio esencial mínimo alfa (MEMα) suplementado con glucosa al 2% e incubar a 37 °C, agitando a 200 rpm durante 16-18 h.
- Diluir el cultivo nocturno transfiriendo de 50 μL a 5 ml de MEMα fresco suplementado con glucosa al 2% e incubar a 37 °C, agitando a 200 rpm, hasta la fase logarítmica media, generalmente entre densidad óptica 600 (OD600nm) de 0.5-0.8. Utilice MEMα para normalizar el cultivo logarítmico medio a un OD600nm de 0,1.
- Transfiera 150 μL de cultivo normalizado a cada pocillo de la placa de 96 pocillos tratada con PLL. Incubar estáticamente durante 18-20 h en una cámara humidificada a 37 °C.
NOTA: Las biopelículas también se pueden cultivar en otros formatos, como diapositivas de μ canales (consulte la Tabla de materiales). - Aspirar el sobrenadante para eliminar las células planctónicas. Lave suavemente la biomasa restante con 150 μL de solución salina equilibrada (HBSS) de Hanks para eliminar las células no unidas. Agregue HBSS gota a gota para evitar interrumpir la biopelícula.
NOTA: Mientras aspira el sobrenadante y HBSS durante los lavados, deje suficiente líquido (sobrenadante o HBSS) en los pocillos que contienen biopelícula de modo que la biopelícula aún esté sumergida. Esto evita la interrupción de la estructura de la biopelícula cuando se agrega HBSS gota a gota para lavar la biopelícula. - Repita el paso 1.6 al menos dos veces más para eliminar todas las células planctónicas. En este punto, las biopelículas están listas para experimentos inmediatos aguas abajo.
NOTA: Si las biopelículas no se utilizan para experimentos con neutrófilos, HBSS se puede sustituir con solución salina tamponada con fosfato (PBS). Se prefiere HBSS sobre PBS ya que HBSS contiene componentes, incluida la glucosa, que proporcionan condiciones óptimas para la activación de neutrófilos34.
2. Cuantificación de biomasa de biofilm
- Prepare una reserva de solución de Crystal Violet (CV) al 0,1% (p/v) (consulte la Tabla de materiales) disolviendo en etanol al 20% (v/v) y al 80% (v/v) H2O. Asegúrese de que el CV esté completamente disuelto en etanol antes de agregarH2O. Filtre la solución.
- Añadir 150 μL de solución CV al 0,1% al biofilm lavado e incubar durante 20 min a temperatura ambiente. Utilice al menos tres pozos vacíos como controles de sólo medios.
- Aspirar la solución CV al 0,1% de los biofilms y lavar los biofilms teñidos con 200 μL de 1x PBS. Repita este proceso para un total de tres lavados para eliminar cualquier exceso de CV de los pocillos.
- Añadir 150 μL de ácido acético glacial al 33% (v/v) diluido conH2O. Incubar a temperatura ambiente en un balancín a 50 rpm durante 30 min para permitir que el CV unido a la biomasa se disuelva completamente.
PRECAUCIÓN: Realice este paso en una campana de flujo laminar con el EPP adecuado, ya que el ácido acético glacial es un producto químico corrosivo. - Mientras tanto, configure el lector de microplacas (consulte la Tabla de materiales) para leer los valores de tinción CV. Después del tratamiento con ácido acético glacial, lea la placa a una longitud de onda de 595 nm.
NOTA: La longitud de onda utilizada para medir el OD de CV puede variar de 500-600 nm35.
3. Aislamiento de neutrófilos
NOTA: Los neutrófilos fueron aislados siguiendo un método previamente publicado con cambios menores36. Este protocolo de aislamiento combina primero la centrifugación por gradiente de densidad, seguida de la sedimentación con dextrano al 3%. Esta sección solo cubre el protocolo general de aislamiento de neutrófilos, centrándose en los cambios realizados en el protocolo publicado. Además, el protocolo descrito a continuación es uno de los muchos métodos que pueden aislar neutrófilos y pueden sustituirse según sea necesario. Otros métodos para aislar neutrófilos incluyen el uso de medios de separación celular o separación celular de anticuerpos magnéticos37.
- Extraer sangre de un donante adulto por venopunción, según el protocolo descrito en el IRB institucional. Antes de la extracción de sangre, asegúrese de que la jeringa tenga suficiente heparina sin conservantes, de modo que la concentración final de heparina sea de 20 U/ml.
- Diluir la sangre heparinizada con 3/4 del volumen de NaCl libre de endotoxinas al 0,9% (ver Tabla de Materiales) enH2Oa temperatura ambiente.
- Por cada 20 ml de la muestra de sangre diluida, alícuota 14 ml de un medio de gradiente de densidad disponible comercialmente (ver Tabla de materiales) en un tubo cónico fresco de 50 ml. Coloque cuidadosamente la muestra de sangre diluida sobre el medio de gradiente de densidad.
- Centrifugar la muestra de sangre en capas a 400 x g durante 40 min a temperatura ambiente. Asegúrese de que la centrífuga tenga una rotura lenta para evitar alterar la capa una vez que se complete la centrifugación.
NOTA: La muestra de sangre tendrá cinco capas que contienen una mezcla de solución salina y plasma, una capa de células mononucleares, medio de gradiente de densidad, neutrófilos y eritrocitos. - Usando una pipeta serológica, aspirar todas las capas por encima de los neutrófilos y el pellet de eritrocitos, seguido de una suave resuspensión del pellet en NaCl libre de endotoxinas frías al 0,9% enH2O. Para cada pellet generado a partir de una muestra de sangre de 20 ml, resuspenda el pellet a 20 ml de volumen total. Agregue un volumen de 1:1 de dextrano al 3% (consulte la Tabla de materiales). Incubar el tubo en posición vertical durante 18-20 minutos sobre hielo.
NOTA: Asegúrese de que el dextrano al 3% esté hecho con NaCl al 0,9% libre de endotoxinas enH2O. - Extraer 20 ml de la capa superior que contiene neutrófilos y algunos eritrocitos en un nuevo tubo cónico de 50 ml y centrifugarlo a 355 x g durante 10 min a 4 °C. Vierta el sobrenadante dejando atrás una bolita roja.
- Resuspender suavemente el pellet en 10 ml deH2Ofrío y estéril durante 30 s para lisar los eritrocitos restantes. Agregue inmediatamente 10 ml de solución salina fría al 0,9% libre de endotoxinas a la mezcla para restaurar la tonicidad. Centrifugar la solución a 233 x g durante 3 min a 4 °C.
- Vierta el sobrenadante y vuelva a suspender el pellet que contiene 95% -97% de neutrófilos en 1 ml de HBSS frío por 20 ml de la muestra de sangre.
- Transfiera 10 μL de los neutrófilos resuspendidos en 90 μL de colorante de exclusión azul de tripano al 0,4% y cuente las células utilizando un hemocitómetro (ver Tabla de materiales).
NOTA: Las células no viables se tiñen de azul ya que el colorante de exclusión azul tripano es impermeable en las células viables. Este protocolo proporciona >99% de viabilidad celular37,38. - Agregue HBSS adicional de tal manera que la concentración final de neutrófilos sea de 4 x 106 células/ml.
NOTA: Para casos con <99% de viabilidad celular, aún se puede lograr la concentración final de 4 x 106 células/ml; sin embargo, el volumen total de solución que contiene 4 x 106 células/ml obtenido disminuirá. La concentración final de neutrófilos se puede ajustar de acuerdo con las necesidades experimentales del usuario. Los neutrófilos se resuspendieron a una concentración final de 4 x 106 células/ml para todos los experimentos descritos a continuación. Para tener en cuenta la variabilidad de donante a donante, se recomienda encarecidamente que todos los experimentos relacionados con neutrófilos se realicen con al menos tres donantes diferentes.
4. Medición de ROS producidas por neutrófilos
- Añadir 100 μL de suero humano normal al 20% (diluido en HBSS) gota a gota al biofilm lavado (paso 1.6) e incubar a 37 °C en condiciones estáticas durante 30 min para opsonizar el biofilm.
- Aspirar la solución sérica al 20% y lavar los biofilms gota a gota con 150 μL de HBSS una vez. Aspirar el HBSS, dejando atrás pozos con biofilms opsonizados.
NOTA: Para la interpretación del experimento, se recomienda un mínimo de cuatro grupos: (A) Neutrófilos + Biopelícula, (B) Neutrófilos + PMA (control positivo, ver Tabla de materiales), (C) Neutrófilos solamente, y (D) Biopelícula solamente. - Agregue luminol (ver Tabla de materiales) a los neutrófilos resuspendidos en HBSS a una concentración de 4 x 106 células/ml tal que la concentración final de luminol sea de 50 μM. Esta solución está lista para usar para los grupos (A) y (C). Añadir 4 x 105 neutrófilos mezclados con luminol a los pocillos con biofilms opsonizados.
- En un tubo separado, prepare una solución de luminol de 50 μM en HBSS sin neutrófilos y agréguela al biofilm que contiene el pocillo (grupo D).
- Alícuota 350 μL de neutrófilos mezclados con luminol y añadir forbol 12-miristato 13-acetato (PMA) a una concentración final de 500 ng/ml a la mezcla. Para el grupo (B), añadir 4 x 105 neutrófilos de esta mezcla en pocillos sin biopelícula. Esto sirve como un control positivo.
NOTA: La concentración de PMA indicada en este paso es relativamente alta para garantizar una respuesta robusta a la ráfaga, ya que los neutrófilos estimulados por PMA son un control positivo. PMA se puede utilizar a una concentración más baja para activar los neutrófilos, dependiendo del experimento. - Centrifugar la placa a 270 x g durante 30 s a 4 °C.
- Asegúrese de que el lector de placas esté ajustado a 37 °C junto con el ajuste de luminiscencia y lectura cinética durante 60 minutos con intervalos de 3 minutos. Coloque la placa en el lector de placas para medir la producción de ROS por neutrófilos durante 60 min.
NOTA: Para este ensayo, las biopelículas se cultivaron en placas blancas utilizadas para ensayos de luminiscencia. El PMA es un agonista conocido para la respuesta de explosión oxidativa39. Cuando realice estudios con PMA, asegúrese de que PMA se agregue en el paso final mientras la solución que contiene neutrófilos esté fría, ya que PMA inicia inmediatamente la respuesta de ráfaga.
5. Imágenes de las interacciones biopelícula-neutrófilos
- Configure una biopelícula siguiendo los pasos 1.2-1.6. Para facilitar las imágenes de biopelícula, emplee una cepa fluorescente de S. aureus, como USA300 que expresa la proteína fluorescente verde (GFP)40,41, para aumentar la facilidad de las imágenes de microscopía.
NOTA: Se utilizó un portaobjetos de 6 μ canales (consulte la Tabla de materiales) en lugar de una placa de 96 pocillos para demostrar el modelo de biopelícula in vitro (paso 1). - Incubar 4 x 106 células/ml de neutrófilos con 100 μM de colorante azul CMAC (7-amino-4-clorometilcumarina) (BCD, ver Tabla de materiales) durante 30 min en un balancín a 37 °C y 5% deCO2. Asegúrese de que las muestras se incuben en la oscuridad y limite la exposición a la luz durante los pasos restantes.
- Para lavar el exceso de BCD, centrifugar los neutrófilos a 270 x g durante 5 min y aspirar el sobrenadante. Resuspender los neutrófilos en HBSS fresco. En este punto, agregue etidio homodímero-1 (consulte la Tabla de materiales) a los neutrófilos teñidos con BCD a una concentración final de 4 μM para controlar la muerte bacteriana y de neutrófilos.
- Agregue 150 μL de neutrófilos a la biopelícula de S. aureus que se ha cultivado en μ portaobjetos, de modo que la proporción de neutrófilos a bacterias sea de 1:30 (neutrófilos: bacterias). Incubar los μ portaobjetos en una cámara humidificada durante 30 min. El número de células bacterianas se basa en los recuentos celulares obtenidos al colocar una biopelícula de 18 h.
- Imagen de la interacción neutrófilo-biopelícula utilizando canales fluorescentes correspondientes a las longitudes de onda de excitación y emisión de los colorantes/proteínas fluorescentes.
NOTA: Para el presente estudio, BCD es 353/466 nm, ethidium homodimer-1 es 528/617 nm y GFP es 395/509 nm. Limite la exposición de la muestra al láser o a la luz para evitar el fotoblanqueo de las muestras. - Analice las imágenes utilizando software de análisis de imágenes de microscopía o programas como FIJI / ImageJ, COMSTAT2, BiofilmQ y BAIT, entre muchos más42,43,44,45.
NOTA: Cuando se trabaja con manchas, es importante tener en cuenta la especificidad de los tintes en uso. Algunas tinciones funcionan en células procariotas y eucariotas, mientras que otras funcionan solo en una. Si los neutrófilos y las biopelículas se tiñen por separado con tintes que pueden teñir ambos tipos de células, asegúrese de lavar cualquier tinte restante antes de combinar neutrófilos y biopelículas para evitar la tinción cruzada.
Resultados
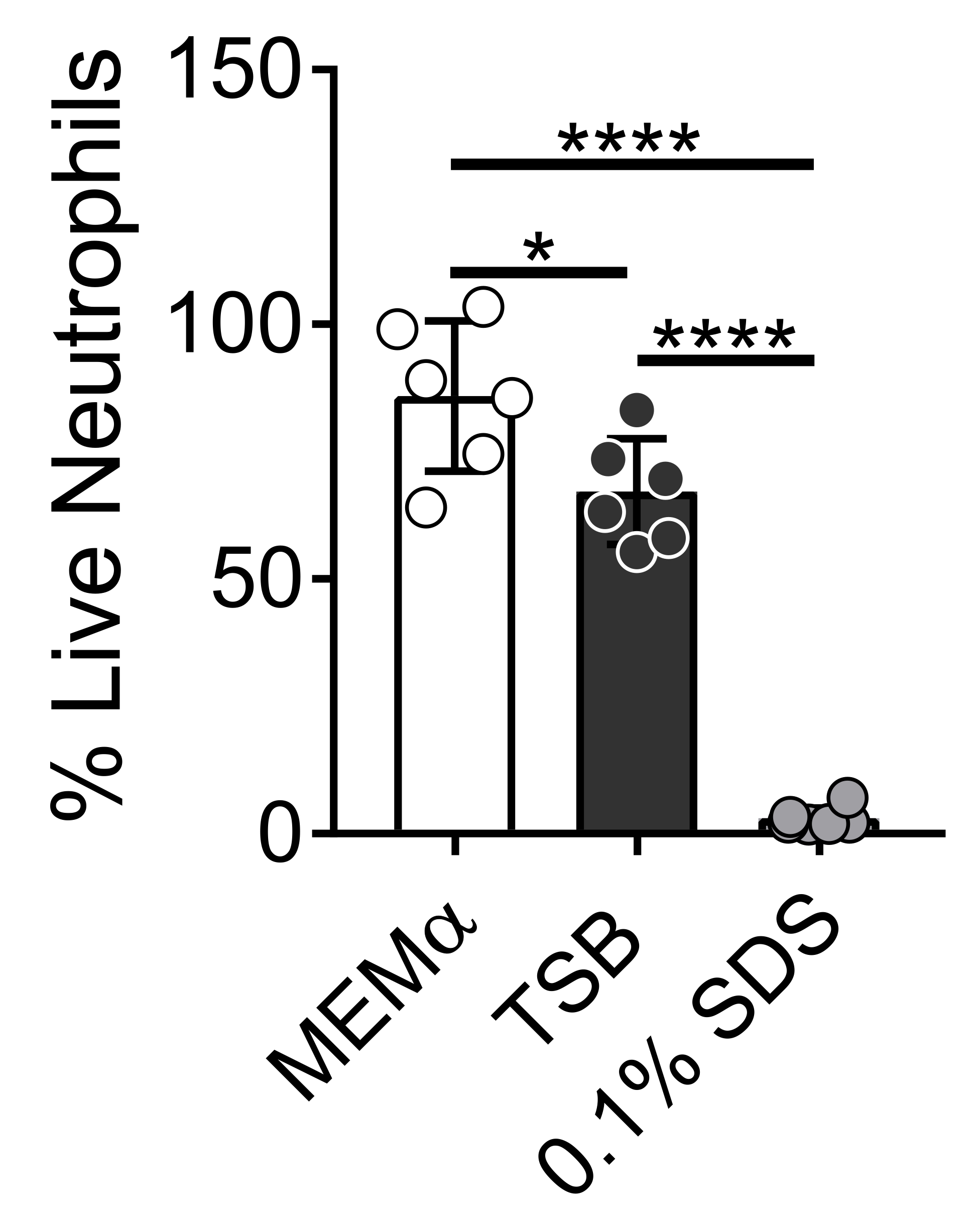
Los medios utilizados para cultivar biopelículas bacterianas influyen en la supervivencia de los neutrófilos. Se probaron diferentes medios para reducir el efecto de los medios solos sobre la viabilidad de los neutrófilos para estudiar las interacciones neutrófilos-biopelícula (Figura 1). Los medios de crecimiento bacteriano como el caldo de soja tríptico minimizan la viabilidad de los neutrófilos, de modo que ~ 60% de los neutrófilos están vivos después de un período de incubación de 30 minutos a 37 ° C con 5% deCO2. Los medios de cultivo de células de mamíferos, como MEMα, no afectan la viabilidad de los neutrófilos y apoyan el crecimiento de biopelículas de S. aureus. De hecho, los medios mínimos promueven el crecimiento robusto de biopelículas en otras bacterias46,47.
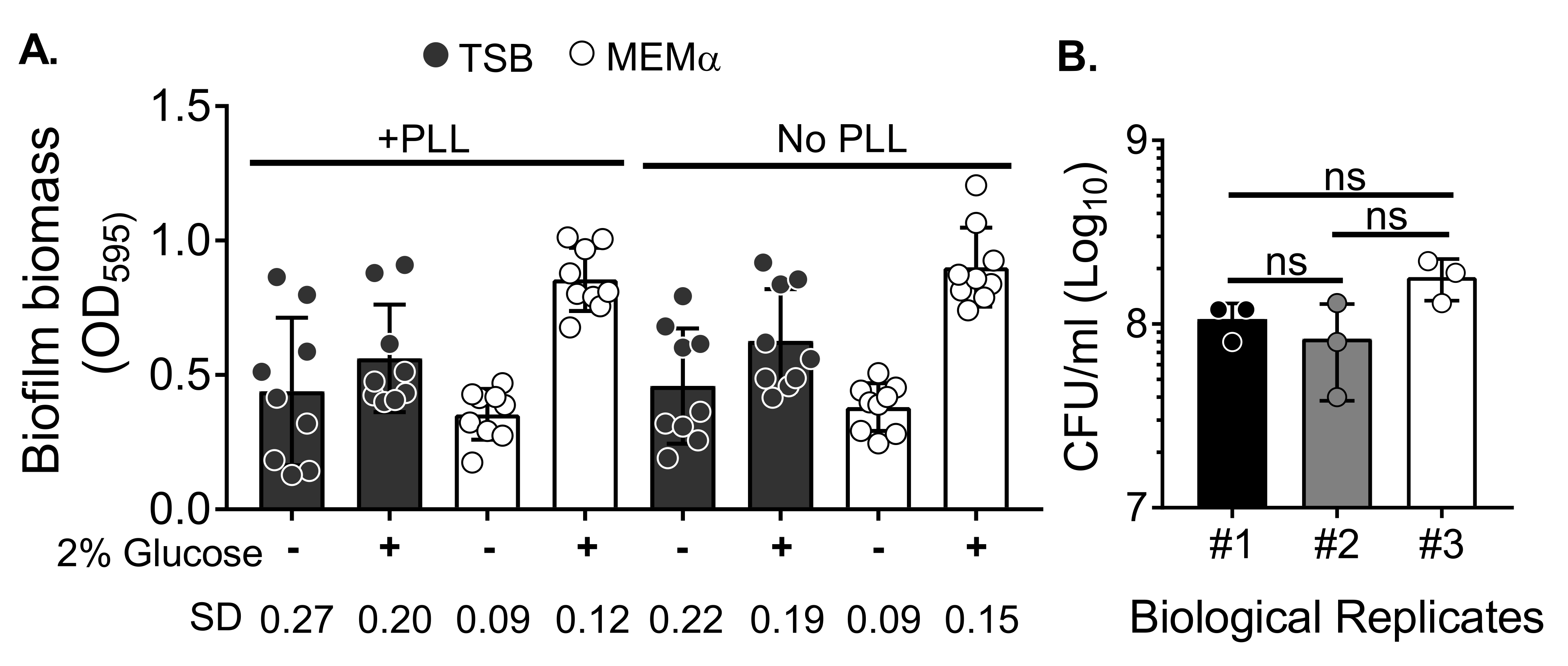
Para evaluar el efecto de los medios sobre el crecimiento del biofilm y la variabilidad en la cuantificación de la biomasa del biofilm después del lavado de la biomasa para eliminar las células planctónicas, se cultivó un biofilm de S. aureus de 18 h en una placa de 96 pocillos, con pocillos tratados o no tratados con poli-L-lisina. Se utilizó un medio rico en nutrientes (caldo de soja tríptico [TSB]) y un medio mínimo (MEMα) tal cual o suplementado con glucosa al 2%. La biomasa de biofilm teñida con CV reveló que la biopelícula de S. aureus cultivada en MEMα suplementada con 2% de glucosa produjo la biopelícula más robusta entre todos los medios probados (Figura 2A). Además, las biopelículas cultivadas en pocillos pretratados con PLL que contenían MEMα + 2% de glucosa mostraron menos variabilidad que las biopelículas en pocillos no tratados con PLL que contenían MEMα + 2% de glucosa. Estas biopelículas mostraron menos variabilidad en la cuantificación a través del ensayo CV35 y la UFC / ml cuando se colocaron después de manipular con precisión las biopelículas para la cuantificación de biomasa. Estas biopelículas contenían, en promedio, 1 x 108 UFC/ml, como se demuestra al colocar las biopelículas en 3 días separados (Figura 2B). Este número es útil para determinar el número de neutrófilos que se agregarán a las biopelículas para los ensayos de funcionalidad de neutrófilos.
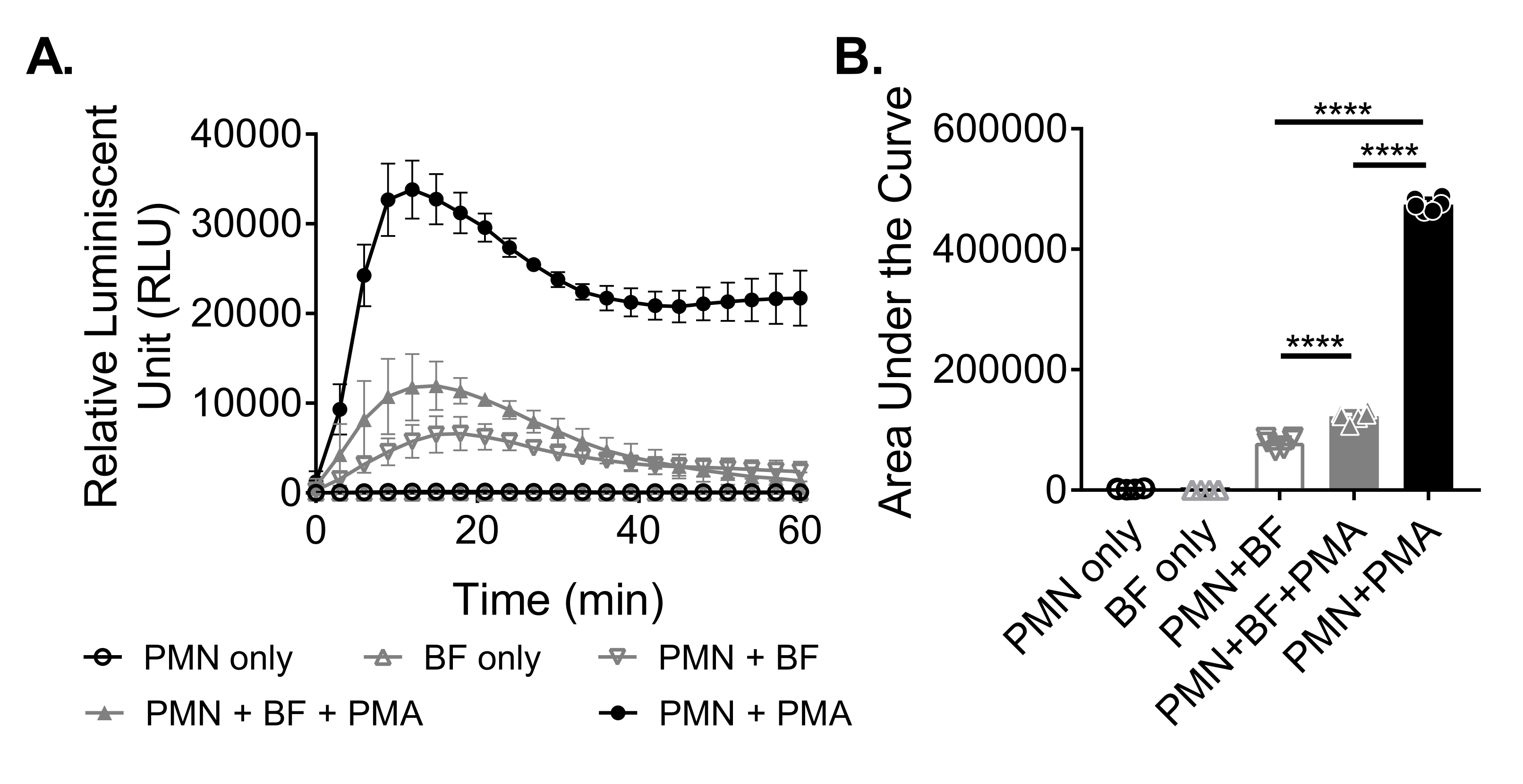
Para medir la producción de ROS por neutrófilos en respuesta a biopelículas, las biopelículas de S. aureus se cultivaron estáticamente durante 18-20 h en una placa de 96 pocillos. Luego se opsonizaron biopelículas y se agregaron neutrófilos. La producción de ROS se midió durante 60 min (Figura 3A). El área bajo la curva se calcula a partir de la curva cinética para cuantificar la producción total de ROS por los neutrófilos. Los neutrófilos tratados con un agonista, como PMA, utilizado como control, muestran un aumento de la producción de ROS. En ausencia de biopelículas, los neutrófilos tratados con PMA mostraron una producción robusta de ROS. En presencia de biopelícula de S. aureus , la producción global de ROS por neutrófilos tratados con PMA disminuyó. En ausencia de PMA, los neutrófilos dependen únicamente de su interacción con la biopelícula, lo que reduce aún más la cantidad de ROS producida (Figura 3B).
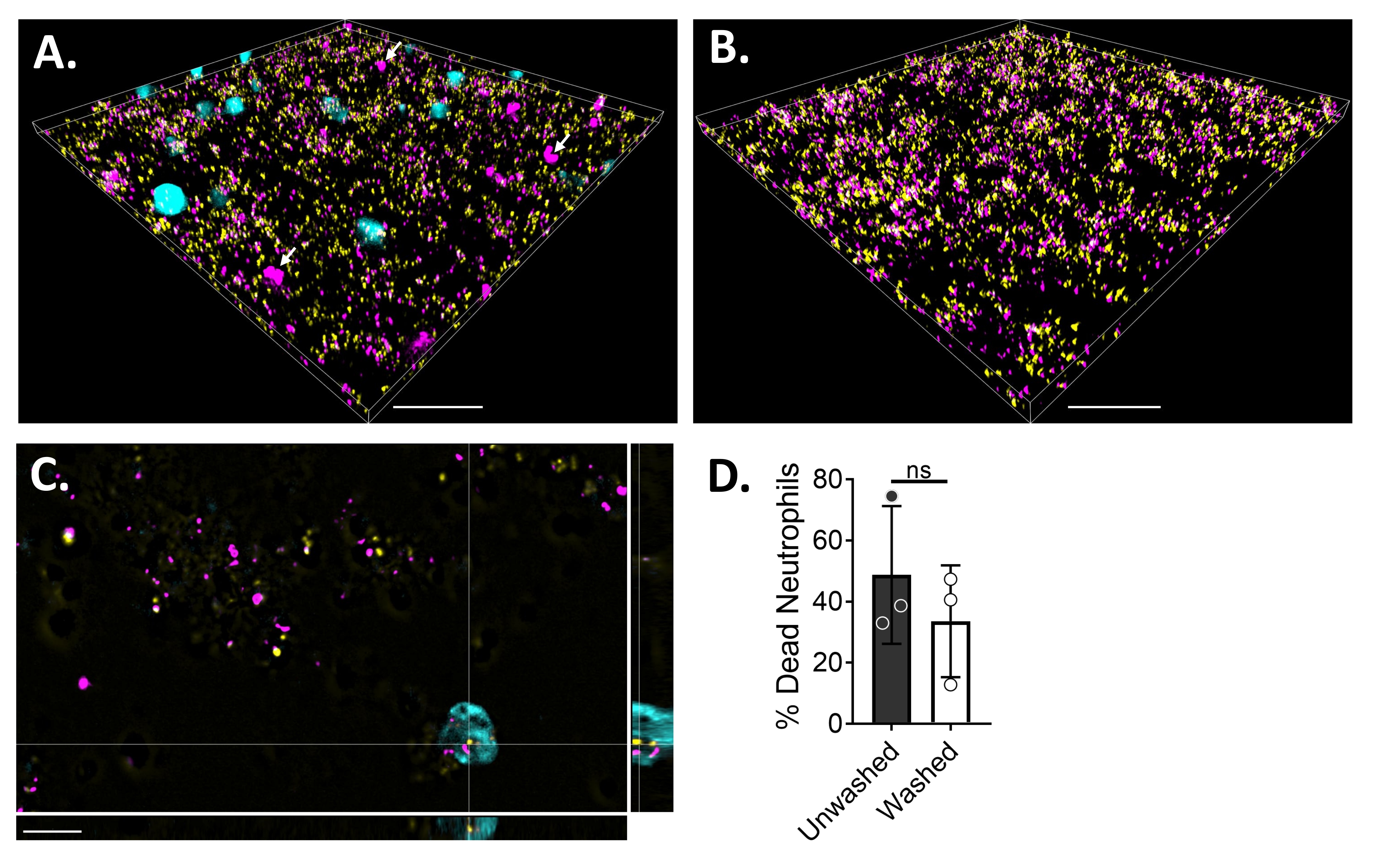
Para visualizar las interacciones neutrófilos-biopelícula utilizando microscopía de fluorescencia, se utilizó una cepa de S. aureus que expresa GFP, colorante CMAC azul y homodímero de etidio-1, que tiñe el citoplasma de células vivas y el ADN de células muertas, respectivamente. La biopelícula de S. aureus se cultivó durante 18 h en un portaobjetos de 6 μ canales. Se agregaron neutrófilos marcados con colorante CMAC azul junto con homodímero-1 de etidio a las biopelículas lavadas y se incubaron durante 30 minutos a 37 ° C con 5% deCO2 antes de la obtención de imágenes. La microscopía fluorescente de campo amplio reveló que muchos neutrófilos estaban localizados en la superficie de las biopelículas de S. aureus, mientras que unos pocos están dentro de la biopelícula (Figura 4A). La interacción entre las células de S. aureus dentro de los neutrófilos también fue evidente (Figura 4C). La mayoría de las células de S. aureus que interactúan con los neutrófilos (cian) estaban muertas (magenta), mientras que unas pocas permanecieron vivas (amarillas) según lo determinado por la tinción de muertos vivos (Figura 4C). A modo de comparación, las biopelículas de S. aureus que expresan GFP se tiñeron con homodímero-1 de etidio, lo que reveló una fracción de la población muerta de S. aureus dentro de la biopelícula (Figura 4B). Los neutrófilos no viables que fueron positivos para el homodímero-1 de etidio se cuantificaron utilizando un software de análisis (ver Tabla de materiales) después de la incubación con biopelículas de S. aureus. Aproximadamente el 48% de los neutrófilos ya estaban muertos dentro de los 30 minutos de la incubación con biopelícula de S. aureus. Durante la optimización del protocolo de microscopía, también se evaluó el efecto del lavado del biofilm y los neutrófilos después de 30 min de incubación para eliminar los neutrófilos no adheridos, revelando alrededor del 33% de neutrófilos muertos todavía unidos al biofilm (Figura 4D).

Figura 1: El ensayo LIVE-DEAD compara la supervivencia de neutrófilos entre los medios de crecimiento bacterianos y de mamíferos. Los neutrófilos se aislaron e incubaron en HBSS, MEMα, TSB o SDS al 0,1% durante 30 min. La tinción LIVE-DEAD se realizó utilizando Calcein AM (vivo) y ethidium homodimer-1 (muerto). Se determinó el porcentaje de neutrófilos vivos, donde los neutrófilos incubados con HBSS se trataron como neutrófilos 100% vivos. Los resultados representan un promedio de dos experimentos independientes realizados por triplicado, con neutrófilos obtenidos de dos donantes diferentes. Los datos se presentan como media ± DE (*p < 0,05, ****p < 0,0001. ANOVA unidireccional). Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}

Figura 2: Cuantificación de biomasa de biofilm en diferentes condiciones y recuento de viabilidad bacteriana de biofilms cultivados en las condiciones optimizadas. (A) S. aureus se sembró en una placa de 96 pocillos recubierta o no con poli-L-lisina (PLL). Los biofilms se cultivaron en TSB, MEMα o cualquiera de los medios suplementados con glucosa al 2% en condiciones estáticas durante 18 h. Se realizó tinción de violeta cristal (CV) para teñir la biomasa de biopelícula. La tinción CV eluyida se diluyó a 1:10 y se leyó en un lector de microplacas. Los resultados representan un promedio de tres experimentos independientes realizados por triplicado. Los datos se presentan como media ± DE. La SD para cada grupo se muestra en la parte inferior para demostrar la variabilidad de las diferentes condiciones de crecimiento de la biopelícula. (B) Los recuentos bacterianos de UFC se obtuvieron a partir de biopelículas cultivadas en un medio optimizado (MEMα + 2% de glucosa). Las biopelículas estáticas de 18 h se sometieron al mismo número de lavados seguidos de una sonicación de 10 minutos para aflojar la biomasa de biopelícula y pasaron a través de una aguja de 22G para interrumpir los agregados antes del recubrimiento. Los resultados representan tres réplicas realizadas por triplicado. Los datos se presentan como media ± DE (ns = no significativo. ANOVA unidireccional). Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}

Figura 3: Cuantificación de la producción de ROS por neutrófilos mediante ensayo de quimioluminiscencia. (A) Los neutrófilos (PMN) se incubaron con biopelículas de S. aureus (BF) lavadas con HBSS en presencia (triángulo gris cerrado) o ausencia (triángulo invertido gris abierto) de PMA para medir la producción de ROS por los neutrófilos. Luminol se utilizó para detectar ROS cada 3 minutos durante 60 minutos en un lector de microplacas. Mientras que los neutrófilos tratados con PMA en ausencia de una biopelícula (círculo negro cerrado) sirvieron como control positivo, los grupos de neutrófilos solo (círculo negro abierto) y solo biopelícula (triángulo gris abierto) sirvieron como controles negativos. Los datos representan un promedio de dos experimentos independientes realizados por triplicado con neutrófilos obtenidos de dos donantes diferentes. Los datos se presentan como media ± DE. (B) El área bajo la curva de (A) se calculó para cuantificar el ROS total generado por los neutrófilos. Los datos se representan como media ± DE. (***p < 0,0001. ANOVA unidireccional). Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}

Figura 4: Visualización de la interacción entre el biofilm de S. aureus y los neutrófilos mediante microscopía de fluorescencia de campo amplio. Los neutrófilos marcados con colorante CMAC azul (cian) se suplementaron con homodímero-1 de etidio (magenta; muerto) antes de incubar con una biopelícula de S. aureus de 18 h (amarillo). Las interacciones biopelícula-neutrófilos se obtuvieron imágenes mediante microscopía fluorescente de campo amplio y las imágenes se procesaron utilizando un software de análisis de imágenes. Los experimentos se realizaron con tres donantes diferentes. Las imágenes representativas se presentan como (A) vista 3D de la biopelícula de S. aureus con neutrófilos vivos (cian) y muertos (magenta; algunos indicados con flechas blancas), (B) vista 3D de una biopelícula de S. aureus en ausencia de neutrófilos con S. aureus vivo que expresa GFP (amarillo) o S. aureus muerto teñido con homodímero-1 de etidio (magenta), (C) una vista ortogonal de S. aureus e interacción de neutrófilos como se muestra en los planos xy, yz y xz, y (D) cuantificación de la viabilidad de neutrófilos en presencia de biopelícula de S. aureus después de 30 minutos, ya sea inmediatamente (sin lavar) o después de tres rondas de lavados con HBSS para eliminar los neutrófilos no adheridos (lavados). La muerte celular de neutrófilos se presenta como media ± SD (prueba t de Student). La barra de escala indica 50 μm en (A) y (B) y 10 μm en (C). Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}
Discusión
Ha habido numerosos esfuerzos para cultivar biofilms robustos y reproducibles de S. aureus para experimentos posteriores in vitro48,49,50. Se describe un protocolo estandarizado que aprovecha la naturaleza catiónica de PLL, así como la suplementación de los medios con glucosa para el crecimiento de biopelículas robustas in vitro de S. aureus. La adición de PLL permite una mejor unión de la célula bacteriana cargada negativamente a las superficies recubiertas de PLL cargadas positivamente. Es importante tener en cuenta que la PLL a una concentración de 10 μg/ml tiene actividad antimicrobiana contra Pseudomonas aeruginosa, Escherichia coli y S. aureus cuando se incuba durante 24 h51. La misma concentración se utiliza para recubrir superficies; sin embargo, el exceso de PLL se aspira, lo que hace que la concentración de PLL sea inferior a 10 μg / ml cuando se siembra para el crecimiento de biopelículas.
Es importante señalar que la PLL ha funcionado solo en medios de crecimiento específicos como MEMα con glucosa al 2%, donde se observó que S. aureus produjo biofilms robustos con una variabilidad mínima (Figura 2A). La concentración de PLL que se utilizará junto con otros tipos de medios requeriría una mayor optimización, como el uso de una mayor concentración de PLL para recubrir los pozos. Además, estas condiciones se han optimizado para una biopelícula monoespecie de S. aureus. Si bien las biopelículas de heridas crónicas son a menudo polimicrobianas, la estandarización de ensayos para estudiar la biopelícula monoespecie y sus interacciones con neutrófilos y otras células inmunes es clave para comprender su contribución a la patogénesis52. Estos protocolos estandarizados se pueden optimizar aún más para mantener y estudiar biopelículas polimicrobianas y sus interacciones con los neutrófilos.
También se observó que los medios de cultivo bacterianos ricos, como la TSB, condujeron a una pérdida de viabilidad de neutrófilos (Figura 1). Por lo tanto, se optimizaron las condiciones de crecimiento de las biopelículas de S. aureus en MEMα, utilizadas para cultivos celulares de mamíferos. Para estudios con neutrófilos, este medio apoya la viabilidad de los neutrófilos y promueve el crecimiento de S. aureus. Si bien se observó que los medios afectan la viabilidad de los neutrófilos, también es importante considerar que los neutrófilos aislados de sangre humana periférica sufren apoptosis ex vivo con aproximadamente un 70% de neutrófilos apoptóticos a las 20 h53. Esto requiere un manejo adecuado, como almacenar los neutrófilos en hielo cuando se prepara para los experimentos, usar reactivos libres de endotoxinas y prevenir la activación de neutrófilos evitando el vórtice de muestras con neutrófilos.
La evaluación del estallido oxidativo en neutrófilos se realiza rutinariamente para determinar el efecto destructor de los neutrófilos sobre el patógeno14,54,55. Estos estudios se realizan con frecuencia con bacterias planctónicas donde se agregan neutrófilos, y la respuesta de explosión oxidativa se cuantifica utilizando quimioluminiscencia amplificada por luminol que detecta aniones superóxido producidos por neutrófilos. El presente protocolo se modifica reemplazando las bacterias planctónicas con biopelícula de S. aureus de 18 h cultivada estáticamente. Como tal, los neutrófilos se pueden agregar directamente a la biopelícula para evaluar su activación. Por otro lado, las bacterias en las biopelículas producen enzimas, como la catalasa y la superóxido dismutasa para desintoxicar ROS 23,56. Las biopelículas de Staphylococcus epidermidis producen una catalasa más alta que su contraparte planctónica bajo estrés57. La quimioluminiscencia total de los neutrófilos estimulados por PMA en una biopelícula de S. aureus es significativamente menor que la de los neutrófilos estimulados por PMA donde la biopelícula está ausente (Figura 2). Esto puede deberse a la actividad de estas enzimas desintoxicantes. Además, las biopelículas de S. aureus producen varias toxinas formadoras de poros llamadas leucocidinas que matan a los neutrófilos58. La respuesta reducida al estallido también es probable que se deba a la viabilidad reducida de los neutrófilos en presencia de biopelícula de S. aureus. Si bien este estudio utiliza luminol que detecta las ROS totales producidas tanto dentro como fuera de las células, otros reactivos, como CM-H 2 DCFDA (5-(y-6)-clorometil-2'7'-diclorodihidrofluoresceína diacetato) o isoluminol, deben considerarse si el objetivo del trabajo es estudiar específicamente la producción de ROS intracelular o extracelular14,53,54.
La capacidad de visualizar las interacciones neutrófilos-biopelícula a través de la microscopía puede ser informativa sobre el comportamiento de los neutrófilos y las biopelículas en presencia mutua. Los espectros de excitación y emisión de los colorantes fluorescentes y proteínas representan una instantánea de la interacción entre una biopelícula de S. aureus de 18 h y neutrófilos después de una incubación de 30 minutos. Para capturar eficazmente las señales de las células teñidas, es importante limitar la exposición de las muestras a fuentes de luz mientras se configuran las muestras para microscopía. Durante la obtención de imágenes, se evitó el fotoblanqueo rápido de las muestras al reducir la intensidad de la fuente de luz al ajustar todos los parámetros, como la altura de la pila Z y el tiempo de exposición para diferentes canales.
Estas prácticas simples permitieron obtener imágenes de microscopía adecuadas donde se observó que pocos neutrófilos se localizan dentro de la biopelícula (Figura 4A). Esto puede deberse a los espacios presentes dentro del biofilm ya que el biofilm de 18 h de S. aureus cultivado en MEMα con 2% de glucosa no cubre uniformemente la superficie (Figura 4B). Sin embargo, el uso de medios ricos en otros estudios ha demostrado un césped uniforme de crecimiento de biopelícula de S. aureus y leucocitos que penetran a través de la biopelícula30,58. Además, también se observa que hubo muerte celular de neutrófilos después de 30 min de incubación con biofilms de S. aureus debido a leucocidinas producidas por biofilm de S. aureus que lisan neutrófilos58 (Figura 4A, D). La adición de un paso de lavado para eliminar los neutrófilos no adheridos después de incubarlos con biofilm durante 30 minutos eliminó ~ 15% de los neutrófilos muertos del sistema en comparación con el grupo sin lavar, en el que la microscopía se realizó inmediatamente después de 30 minutos de incubación (Figura 4D). También se observaron neutrófilos interactuando con S. aureus (Figura 4C). Se requieren experimentos adicionales para evaluar si S. aureus es engullido por neutrófilos o unido a la superficie celular de los neutrófilos54. La obtención de imágenes de neutrófilos y biopelículas es el primer paso para evaluar varias funcionalidades de neutrófilos aguas abajo, como la fagocitosis y la NETosis54,59. El efecto de los neutrófilos sobre las biopelículas también se puede evaluar cuantificando la biomasa de la biopelícula, los cambios estructurales de la biopelícula y la viabilidad de la biopelícula, entre muchos otros, utilizando las herramientas de análisis de imágenes enumeradas en el paso 5.6. Por último, existe variabilidad de donante a donante en los neutrófilos; Por lo tanto, se recomienda que se utilicen al menos tres donantes diferentes para estudios con neutrófilos.
En general, los ensayos in vitro estandarizados se combinaron para evaluar las interacciones entre los neutrófilos y las biopelículas. Aunque estos ensayos utilizan S. aureus, los protocolos descritos pueden adaptarse fácilmente para estudiar otros patógenos. Si bien existen varios modelos in vivo para estudiar las interacciones huésped-patógeno, pueden ser costosos y laboriosos, especialmente si las condiciones no están optimizadas. Trabajar con ensayos in vitro estandarizados permite optimizar las condiciones experimentales y confirmar las observaciones antes de pasar a un sistema in vivo. Finalmente, se han utilizado varios modelos de infección animal para estudiar las interacciones biopelícula-neutrófilos in vivo. Sin embargo, es importante considerar las diferencias inmunológicas entre los modelos humanos y animales60,61,62,63. Esto requiere el uso de neutrófilos derivados de humanos para estudiar estas complejas interacciones huésped-patógeno.
Divulgaciones
Los autores no tienen nada que revelar.
Agradecimientos
Este trabajo fue financiado por el Instituto Nacional de Alergias y Enfermedades Infecciosas (R01AI077628) a DJW y un Premio de Desarrollo Profesional de la Asociación Americana del Corazón (19CDA34630005) a ESG. Agradecemos al Dr. Paul Stoodley por proporcionarnos la cepa USA 300 LAC GFP. Además, reconocemos los recursos del Centro de Microscopía e Imágenes del Campus (CMIF) y el Recurso Compartido de Microscopía (MSR) del Centro Integral del Cáncer de OSU (OSUCCC), la Universidad Estatal de Ohio. También agradecemos a Amelia Staats, Peter Burback y Lisa Coleman del laboratorio Stoodley por realizar extracciones de sangre.
Materiales
| Name | Company | Catalog Number | Comments |
| 0.9% sodium chloride irrigation, USP | Baxter | 2F7124 | Endotoxin-free; Used for isolation of neutrophils |
| 150 mL rapid-flow filter unit | Thermo Scientific | 565-0020 | |
| 200 proof ethanol | VWR | 89125-188 | |
| 3 mL syringe | BD | 309657 | Used for blood draw |
| 50 mL conical centrifuge tubes | Thermo Scientific | 339652 | |
| 60 mL syringe | BD | 309653 | Used for blood draw |
| Agar | Fisher Bioreagents | BP1423-2 | |
| Alcohol swab | BD | Used for blood draw | |
| Band-aids | Used for blood draw | ||
| BD Bacto Tryptic Soy Broth | BD | DF0370-07-5 | Combine with 1.5% agar to make Tryptic Soy Agar |
| Cell counter | Bal Saupply | 202C | |
| CellTracker blue CMCH | Invitrogen | C2111 | Blue CMAC Dye (BCD) |
| Clear bottom 96-well flat bottom polystyrene plates | Costar | 3370 | |
| Cotton gauze | Fisherbrand | 13-761-52 | Used for blood draw |
| Crystal violet | Acros Organic | 40583-0250 | |
| Culture tubes | Fisherbrand | 14-961-27 | Borosilicate Glass 13 x 100 mm |
| D-(+)-glucose | Sigma | G-8270 | |
| Dextran from Leuconostoc spp. | Sigma | 31392-250G | Used for isolation of neutrophils |
| Dulbecco's phosphate buffered saline (DPBS) 1x | Gibco | 14190-144 | |
| Ethidium homodimer-1 | Invitrogen | L3224 B | |
| Ficoll-Paque plus | Cytiva | 17144003 | Used for isolation of neutrophils (density gradient medium) |
| Hanks' balanced salt solution (HBSS) 1x | Corning cellgro | 21-022-CV | without calcium, magnesium, and phenol red |
| Hemacytometer | Bright Line | ||
| Heparin | Novaplus | NDC 63323-540-57 | 1000 USP units/mL, Used for blood draw |
| IMARIS 9.8 | Oxford Instruments | Microscopy image analysis software | |
| Luminol | Sigma | A8511-5G | |
| Minimal essential media (MEM) Alpha 1x | Gibco | 41061-029 | |
| Needle (23 G1) | BD | 305145 | Used for blood draw |
| Nikon Eclipse Ti2 | Nikon | ||
| NIS-Elements | Nikon | Quantification of dead neutrophils | |
| Normal human serum | Complement Technology | NHS | |
| Petri Dish (100 x 15 mm) | VWR | 25384-342 | |
| Phorbol 12-myristate 13-acetate | |||
| Poly-L-lysine solution | Sigma | P4707-50ML | |
| Sodium chloride | Fisher Bioreagents | BP358-10 | Used for neutrophil isolation |
| SoftMax Pro Software | Molecular Devices | Microplate reader software used for data acquisition | |
| SpectraMax i3x | Molecular Devices | Microplate reader | |
| Sterile water for irrigation, USP | Baxter | 2F7114 | Endotoxin-free; Used for neutrophil isolation |
| Surflo winged infusion set | Terumo | SC*19BLK | 19 G x 3/4", used for blood draw |
| Trypan blue stain (0.4%) | Gibco | 15250-061 | |
| Turnicate | Used for blood draw | ||
| UltraPure distilled water | Invitrogen | 10977015 | |
| White opaque 96-well plates | Falcon | 353296 | Tissue culture treated and flat bottom plate |
| μ-Slide VI 0.4 | Ibidi | 80601 | μ-channel slide |
Referencias
- Donlan, R. M. Biofilms: microbial life on surfaces. Emerging Infectious Diseases. 8 (9), 881-890 (2002).
- Alhede, M., et al. Phenotypes of non-attached Pseudomonas aeruginosa aggregates resemble surface attached biofilm. PLoS One. 6 (11), 27943(2011).
- Hall-Stoodley, L., Costerton, J. W., Stoodley, P. Bacterial biofilms: from the natural environment to infectious diseases. Nature Reviews: Microbiology. 2 (2), 95-108 (2004).
- Donlan, R. M., Costerton, J. W. Biofilms: survival mechanisms of clinically relevant microorganisms. Clinical Microbiology Reviews. 15 (2), 167-193 (2002).
- Schilcher, K., Horswill, A. R. Staphylococcal biofilm development: structure, regulation, and treatment strategies. Microbiology and Molecular Biology Reviews. 84 (3), 0002(2020).
- Kaplan, J. B. Biofilm dispersal: mechanisms, clinical implications, and potential therapeutic uses. Journal of Dental Research. 89 (3), 205-218 (2010).
- Otto, M. Staphylococcal Biofilms. Microbiology Spectrum. 6 (4), 10(2018).
- Moormeier, D. E., Bayles, K. W. Staphylococcus aureus biofilm: a complex developmental organism. Molecular Microbiology. 104 (3), 365-376 (2017).
- Gross, M., Cramton, S. E., Gotz, F., Peschel, A. Key role of teichoic acid net charge in Staphylococcus aureus colonization of artificial surfaces. Infection and Immunity. 69 (5), 3423-3426 (2001).
- Zheng, Y., He, L., Asiamah, T. K., Otto, M. Colonization of medical devices by staphylococci. Environmental Microbiology. 20 (9), 3141-3153 (2018).
- Donlan, R. M. Biofilms and device-associated infections. Emerging Infectious Diseases. 7 (2), 277-281 (2001).
- Kolaczkowska, E., Kubes, P. Neutrophil recruitment and function in health and inflammation. Nature Reviews Immunology. 13 (3), 159-175 (2013).
- Amulic, B., Cazalet, C., Hayes, G. L., Metzler, K. D., Zychlinsky, A. Neutrophil function: from mechanisms to disease. Annual Review of Immunology. 30, 459-489 (2012).
- Chen, Y., Junger, W. G. Measurement of oxidative burst in neutrophils. Methods in Molecular Biology. 844, 115-124 (2012).
- van Kessel, K. P., Bestebroer, J., van Strijp, J. A. Neutrophil-mediated phagocytosis of Staphylococcus aureus. Frontiers in Immunology. 5, 467(2014).
- Nguyen, G. T., Green, E. R., Mecsas, J. Neutrophils to the ROScue: Mechanisms of NADPH oxidase activation and bacterial resistance. Frontiers in Cellular and Infection Microbiology. 7, 373(2017).
- Saini, R., Singh, S. Inducible nitric oxide synthase: An asset to neutrophils. Journal of Leukocyte Biology. 105 (1), 49-61 (2019).
- Imlay, J. A. The molecular mechanisms and physiological consequences of oxidative stress: lessons from a model bacterium. Nature Reviews Microbiology. 11 (7), 443-454 (2013).
- Segal, A. W. The function of the NADPH oxidase of phagocytes and its relationship to other NOXs in plants, invertebrates, and mammals. The International Journal of Biochemistry and Cell Biology. 40 (4), 604-618 (2008).
- Fang, F. C. Antimicrobial reactive oxygen and nitrogen species: concepts and controversies. Nature Reviews Microbiology. 2 (10), 820-832 (2004).
- Bogdan, C. Nitric oxide and the immune response. Nature Immunology. 2 (10), 907-916 (2001).
- Chua, S. L., et al. Reactive oxygen species drive evolution of pro-biofilm variants in pathogens by modulating cyclic-di-GMP levels. Open Biology. 6 (11), 160162(2016).
- El Haj, C., Lichtenberg, M., Nielsen, K. L., Bjarnsholt, T., Jensen, P. O. Catalase protects biofilm of Staphylococcus aureus against daptomycin activity. Antibiotics. 10 (5), 511(2021).
- Ghimire, N., et al. Direct microscopic observation of human neutrophil-Staphylococcus aureus interaction in vitro suggests a potential mechanism for initiation of biofilm infection on an implanted medical device. Infection and Immunity. 87 (12), 00745(2019).
- Bhattacharya, M., et al. Leukocidins and the nuclease nuc prevent neutrophil-mediated killing of Staphylococcus aureus biofilms. Infection and Immunity. 88 (10), 00372(2020).
- Bogachev, M. I., et al. Fast and simple tool for the quantification of biofilm-embedded cells sub-populations from fluorescent microscopic images. PLoS One. 13 (5), 0193267(2018).
- Kerstens, M., et al. A flow cytometric approach to quantify biofilms. Folia Microbiologica. 60 (4), 335-342 (2015).
- Meyle, E., et al. Destruction of bacterial biofilms by polymorphonuclear neutrophils: relative contribution of phagocytosis, DNA release, and degranulation. The International Journal of Artificial Organs. 33 (9), 608-620 (2010).
- Oveisi, M., et al. Novel assay to characterize neutrophil responses to oral biofilms. Infection and Immunity. 87 (2), 00790(2019).
- Leid, J. G., Shirtliff, M. E., Costerton, J. W., Stoodley, P. Human leukocytes adhere to, penetrate, and respond to Staphylococcus aureus biofilms. Infection and Immunity. 70 (11), 6339-6345 (2002).
- Fey, P. D., et al. A genetic resource for rapid and comprehensive phenotype screening of nonessential Staphylococcus aureus genes. mBio. 4 (1), 00537(2013).
- Cody, W. L., et al. Skim milk enhances the preservation of thawed -80 degrees C bacterial stocks. Journal of Microbiological Methods. 75 (1), 135-138 (2008).
- Sanders, E. R. Aseptic laboratory techniques: plating methods. Journal of Visualized Experiments. 63, 3064(2012).
- Freitas, M., Porto, G., Lima, J. L., Fernandes, E. Optimization of experimental settings for the analysis of human neutrophils oxidative burst in vitro. Talanta. 78 (4-5), 1476-1483 (2009).
- Merritt, J. H., Kadouri, D. E., O'Toole, G. A. Growing and analyzing static biofilms. Current Protocols in Microbiology. , Chapter 1 Unit 1B 1 (2005).
- Nauseef, W. M. Isolation of human neutrophils from venous blood. Methods in Molecular Biology. 412, 15-20 (2007).
- Zhou, L., et al. Impact of human granulocyte and monocyte isolation procedures on functional studies. Clinical and Vaccine Immunology. 19 (7), 1065-1074 (2012).
- Quach, A., Ferrante, A. The application of dextran sedimentation as an initial step in neutrophil purification promotes their stimulation, due to the presence of monocytes. Journal of Immunology Research. 2017, 1254792(2017).
- Karlsson, A., Nixon, J. B., McPhail, L. C. Phorbol myristate acetate induces neutrophil NADPH-oxidase activity by two separate signal transduction pathways: dependent or independent of phosphatidylinositol 3-kinase. Journal of Leukocyte Biology. 67 (3), 396-404 (2000).
- Staats, A., et al. Rapid aggregation of Staphylococcus aureus in synovial fluid is influenced by synovial fluid concentration, viscosity, and fluid dynamics, with evidence of polymer bridging. mBio. , 0023622(2022).
- Chiu, I. M., et al. Bacteria activate sensory neurons that modulate pain and inflammation. Nature. 501 (7465), 52-57 (2013).
- Hartig, S. M. Basic image analysis and manipulation in ImageJ. Current Protocols in Molecular Biology. 102 (1), 14-15 (2013).
- Heydorn, A., et al. Quantification of biofilm structures by the novel computer program COMSTAT. Microbiology. 146 (10), 2395-2407 (2000).
- Hartmann, R., et al. Quantitative image analysis of microbial communities with BiofilmQ. Nature Microbiology. 6 (2), 151-156 (2021).
- Luo, T. L., et al. Introducing BAIT (Biofilm Architecture Inference Tool): a software program to evaluate the architecture of oral multi-species biofilms. Microbiology. 165 (5), 527-537 (2019).
- Naves, P., et al. Measurement of biofilm formation by clinical isolates of Escherichia coli is method-dependent. Journal of Applied Microbiology. 105 (2), 585-590 (2008).
- Eze, E. C., El Zowalaty, M. E. Combined effects of low incubation temperature, minimal growth medium, and low hydrodynamics optimize Acinetobacter baumannii biofilm formation. Infection and Drug Resistance. 12, 3523-3536 (2019).
- Harris, L. G., Tosatti, S., Wieland, M., Textor, M., Richards, R. G. Staphylococcus aureus adhesion to titanium oxide surfaces coated with non-functionalized and peptide-functionalized poly(L-lysine)-grafted-poly(ethylene glycol) copolymers. Biomaterials. 25 (18), 4135-4148 (2004).
- Miao, J., et al. Biofilm formation of Staphylococcus aureus under food heat processing conditions: first report on cml production within biofilm. Scientific Reports. 9 (1), 1312(2019).
- Lade, H., et al. Biofilm formation by Staphylococcus aureus clinical isolates is differentially affected by glucose and sodium chloride supplemented culture media. Journal of Clinical Medicine. 8 (11), 1853(2019).
- Guzel Kaya, G., et al. Antibacterial activity of linezolid against gram-negative bacteria: utilization of epsilon-Poly-l-Lysine capped silica xerogel as an activating carrier. Pharmaceutics. 12 (11), 1126(2020).
- Clinton, A., Carter, T. Chronic wound biofilms: pathogenesis and potential therapies. Laboratory Medicine. 46 (4), 277-284 (2015).
- Scheel-Toellner, D., et al. Reactive oxygen species limit neutrophil life span by activating death receptor signaling. Blood. 104 (8), 2557-2564 (2004).
- Pestrak, M. J., et al. Pseudomonas aeruginosa rugose small-colony variants evade host clearance, are hyper-inflammatory, and persist in multiple host environments. PLoS Pathogens. 14 (2), 1006842(2018).
- Guerra, F. E., et al. Staphylococcus aureus SaeR/S-regulated factors reduce human neutrophil reactive oxygen species production. Journal of Leukocyte Biology. 100 (5), 1005-1010 (2016).
- Suo, Y., Huang, Y., Liu, Y., Shi, C., Shi, X. The expression of superoxide dismutase (SOD) and a putative ABC transporter permease is inversely correlated during biofilm formation in Listeria monocytogenes 4b G. PLoS One. 7 (10), 48467(2012).
- Olwal, C. O., Ang'ienda, P. O., Ochiel, D. O. Alternative sigma factor B (sigma(B)) and catalase enzyme contribute to Staphylococcus epidermidis biofilm's tolerance against physico-chemical disinfection. Scientific Reports. 9 (1), 5355(2019).
- Bhattacharya, M., et al. Staphylococcus aureus biofilms release leukocidins to elicit extracellular trap formation and evade neutrophil-mediated killing. Proceedings of the National Academy of Sciences of the United States of America. 115 (28), 7416-7421 (2018).
- Masuda, S., et al. NETosis markers: Quest for specific, objective, and quantitative markers. Clinica Chimica Acta. 459, 89-93 (2016).
- Dworsky, E. M., et al. Novel in vivo mouse model of implant related spine infection. Journal of Orthopaedic Research. 35 (1), 193-199 (2017).
- Pletzer, D., Mansour, S. C., Wuerth, K., Rahanjam, N., Hancock, R. E. New mouse model for chronic infections by gram-negative bacteria enabling the study of anti-infective efficacy and host-microbe interactions. mBio. 8 (1), 00140(2017).
- Davis, M. M. A prescription for human immunology. Immunity. 29 (6), 835-838 (2008).
- Mestas, J., Hughes, C. C. Of mice and not men: differences between mouse and human immunology. The Journal of Immunology. 172 (5), 2731-2738 (2004).
Reimpresiones y Permisos
Solicitar permiso para reutilizar el texto o las figuras de este JoVE artículos
Solicitar permisoThis article has been published
Video Coming Soon
ACERCA DE JoVE
Copyright © 2025 MyJoVE Corporation. Todos los derechos reservados