
Se requiere una suscripción a JoVE para ver este contenido. Inicie sesión o comience su prueba gratuita.
Method Article
Un método de bajo costo para medir la productividad primaria in situ de las comunidades de perifitón de las aguas lénticas
En este artículo
Resumen
Aquí se presenta un método / instalación rentable y transportable para medir la productividad primaria de las esteras microbianas en condiciones reales de temperatura ambiental y luz in situ . La configuración experimental se basa en materiales ampliamente disponibles y se puede utilizar en diversas condiciones, al tiempo que ofrece las ventajas de los modelos basados en laboratorio.
Resumen
La medición de la productividad primaria in situ del perifitón durante el gradiente de la temporada de crecimiento puede dilucidar el efecto cuantitativo de los impulsores ambientales (principalmente la concentración de fósforo y la intensidad de la luz) y la composición de las especies en la productividad primaria. La productividad primaria es impulsada principalmente por la intensidad de la luz, la temperatura, la disponibilidad de nutrientes y la distribución de las especies iónicas del sistema carbonatado en las profundidades respectivas de la zona eufótica. Es un sistema complejo que es muy difícil de simular en el laboratorio. Esta barcaza flotante barata, transportable y fácil de construir permite medir la productividad primaria de forma precisa y directa en las condiciones naturales reales. La metodología se basa en medir la productividad primaria en tiempo real utilizando sensores de oxígeno no invasivos integrados en frascos de vidrio herméticamente sellados, lo que permite el monitoreo del flujo de oxígeno en línea y proporciona nuevos conocimientos sobre las actividades metabólicas. Las mediciones estacionales detalladas in situ de la productividad primaria bruta de las esteras microbianas (u otros organismos bentónicos) pueden mejorar el conocimiento actual de los procesos que controlan la dinámica de la productividad primaria en las aguas lénticas.
Introducción
La productividad primaria es la única entrada de carbono autóctono en los sistemas acuáticos que forman toda la red alimentaria del sistema1. Por lo tanto, la estimación precisa de la productividad primaria es un paso esencial hacia la comprensión del funcionamiento de los ecosistemas acuáticos. Las zonas litorales son áreas de alta productividad primaria y biodiversidad. Además del fitoplancton, se supone que el perifitón (en lo sucesivo denominados esteras microbianas) y las macroalgas contribuyen significativamente a la productividad primaria en las zonas litorales2. Debido a su estilo de vida sésil y su significativa heterogeneidad espacial, la cuantificación de la productividad primaria no es trivial.
La productividad primaria es impulsada principalmente por la intensidad de la luz, la temperatura, la disponibilidad de nutrientes y la distribución de las especies iónicas del sistema carbonatado en las respectivas profundidades de las zonas eufóticas 3,4. La profundidad influye notablemente en la distribución espacial de las esteras microbianas. Las comunidades microbianas deben hacer frente a los efectos adversos de la alta irradiación y las pronunciadas variaciones estacionales de temperatura en profundidades poco profundas y con menor intensidad de luz a mayores profundidades. Además del gradiente de profundidad, las interacciones tróficas dinámicas generan patrones espaciales múltiples y complejos a diferentes escalas5. Este complejo sistema es complicado de simular en el laboratorio. La forma más precisa de inferir la actividad metabólica de los productores primarios individuales a partir de zonas litorales es establecer experimentos in situ.
La metodología introducida en este trabajo se basa en el método tradicional de cámara 2,6,7, junto con una barcaza flotante de bajo costo transportable y fácil de construir. Esto permite la medición de la productividad primaria a diferentes profundidades bajo el espectro de luz natural, la temperatura y la diferente distribución de las especies iónicas del sistema carbonato con la profundidad. El método se basa en el principio de oxígeno de botella claro versus oscuro, que se empleó por primera vez para medir la fotosíntesisdel fitoplancton 6 y todavía se usa comúnmente 6,7. Compara la tasa de cambio en el oxígeno en las botellas mantenidas en la luz (que incluye los efectos de la productividad primaria y la respiración) con las que se mantienen en la oscuridad (solo respiración)8. El método utiliza la evolución del oxígeno (fotosíntesis) como un proxy para la productividad primaria. Las variables medidas son la productividad neta del ecosistema (NEP, como un cambio en la concentración de O2 a lo largo del tiempo en condiciones de luz) y la respiración del ecosistema (RE, como un cambio en la concentración deO2 a lo largo del tiempo en la oscuridad). La productividad bruta del ecosistema (GEP) es el cálculo de la diferencia entre los dos (Tabla 1). El término "ecosistema" se utiliza aquí para denotar que el perifitón está compuesto de organismos autótrofos y heterótrofos. La mejora más significativa de este método de cámara tradicional es el uso de sensores ópticos de oxígeno no invasivos y la optimización de este método principalmente planctónico para medir la productividad primaria perifítica.
La técnica se describe en el ejemplo de medición de esteras microbianas en la zona litoral de lagos post-minería recién emergidos en la República Checa: Milada, Most y Medar. La actividad metabólica de las esteras microbianas se determina mediante la medición directa in situ de los flujos deO2 realizada directamente a profundidades específicas, donde las comunidades estudiadas ocurren naturalmente. La actividad heterótrofa y fototrófica se mide en botellas de vidrio cerradas equipadas con sensores ópticos de oxígeno no invasivos. Estos sensores detectan la presión parcial de oxígeno utilizando la fluorescencia de tintes sensibles a la luz. Las botellas con esteras microbianas se suspenden y se incuban en un dispositivo flotante a las profundidades adecuadas. La concentración de oxígeno dentro de las botellas se midió continuamente durante el período de luz del día desde el pequeño bote.
Los buzos recolectan muestras de esteras microbianas intactas y las colocan en botellas de incubación herméticas al gas a profundidades designadas. Cada botella está equipada con un microsensor óptico de oxígeno no invasivo, que monitorea la productividad/consumo deO2 a lo largo del tiempo. Todas las mediciones se realizan en cinco pares de réplica de oscuridad / luz en cada profundidad. La temperatura y las intensidades de radiación fotosintéticamente activa (PHAR) se miden a profundidades respectivas a lo largo de la incubación. Después de 6 h de incubación in situ (horas de luz), las esteras microbianas se cosechan de las botellas y se secan. Los flujos deO2 se normalizan a biomasa microbiana. Como control, los flujos se corrigen para detectar cambios en la concentración de O2 en botellas separadas de gas hermético claro y oscuro (controles en blanco) que contienen agua de lago sin biomasa microbiana. A continuación se presentan instrucciones detalladas para construir la barcaza flotante y realizar todo el experimento paso a paso. Este artículo también presenta resultados representativos de las mediciones de esteras microbianas a dos profundidades (1 m y 2 m), con cinco réplicas a cada profundidad. La temperatura real y la intensidad de la luz se midieron durante todo el experimento utilizando registradores de datos.
Protocolo
NOTA: Antes del muestreo, determine el grado de réplicas en función de las necesidades generales del proyecto, el diseño estadístico o la cantidad esperada de variabilidad de la muestra. Se sugieren cinco pares replicados de botellas de incubación claras y oscuras para un análisis estadístico preciso y para tener en cuenta la posible pérdida o rotura de la muestra. La barcaza experimental flotante descrita está diseñada para transportar cinco réplicas más un par de controles en blanco; ver Figura 1 para un dibujo técnico de la barcaza experimental.

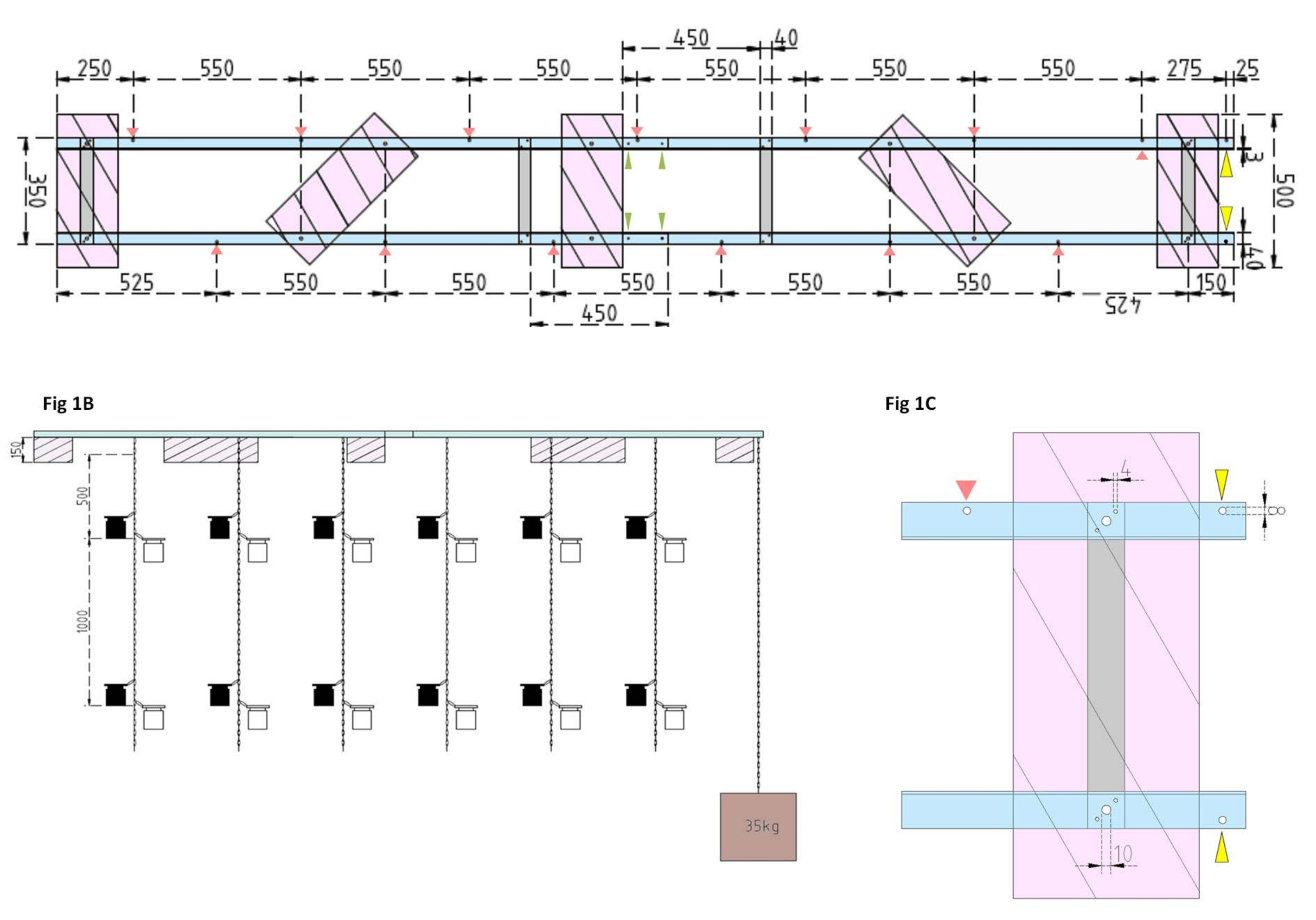
Figura 1: Dibujos técnicos de la barcaza experimental y el flotador lateral. (A) Vista superior: el marco de la barcaza consta de cuatro piezas de perfil L de ángulo de aluminio (azul) que están unidas entre sí por cuatro barras planas de aluminio (gris). Los flotadores XPS (rosa) se montan en el marco en dos puntos, cada uno en las piezas de aluminio paralelas. Las cadenas para botellas de incubación se unen al marco en ambos lados mediante ganchos de presión en orificios preperforados (flechas rojas) con 550 mm de separación entre ellos. Las cadenas se proporcionaron con ganchos a presión a distancias de 1 m y 2 m para la fijación de la botella de incubación (elija la posición de los ganchos de presión de acuerdo con la profundidad experimental). El ancla de hormigón está asegurada a la proa de la barcaza, donde un voladizo de 25 mm permite que dos agujeros preperforados (puntas de flecha amarillas) sirvan como punto de fijación para la cadena del ancla y el buque de investigación. El marco se monta o desmonta fácilmente a través de las uniones paralelas entre las cuatro piezas angulares de aluminio (puntas de flecha verdes). (B) La vista lateral muestra las cadenas suspendidas con botellas de incubación colgantes y anclaje de hormigón (cuadrado marrón). (C) El flotador lateral XPS: Las piezas L de ángulo de aluminio paralelo (azul) están unidas por barras planas verticales de aluminio (gris). Debajo de la sección de la barra transversal, el flotador XPS (rosa) está montado con los tamaños de orificio necesarios indicados (4 mm). Las cadenas suspendidas se fijan con ganchos de presión en agujeros de 8 mm (punta de flecha roja). En la proa de la barcaza, se perforan dos agujeros de 8 mm en el aluminio sobresaliente, uno para asegurar el ancla a la barcaza (punta de flecha amarilla) y otro para amarrar el buque de investigación a la barcaza (azul). Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}
1. Construcción de la barcaza experimental
NOTA: La barcaza flotante consta de dos secciones iguales montadas juntas, lo que permite un fácil montaje / desmontaje. Todas las piezas usadas se pueden comprar en cualquier mercado de pasatiempos o tienda de venta de materiales de construcción.
- Primero, ensamble el marco de la barcaza uniendo cuatro piezas de perfil en L de ángulo de aluminio (40 mm x 40 mm x 3 mm; longitud de 2,000 mm) juntas usando cuatro barras planas de aluminio (40 mm x 3 mm x 350 mm), 16 tornillos (4 mm x 15 mm con tuercas hexagonales) y 32 arandelas (4 mm x 10 mm).
NOTA: La distancia y la posición de las barras planas se muestran en el dibujo técnico de la Figura 1A. La fijación detallada de los flotadores a las barras planas laterales se muestra en la Figura 1B. - Para unir las dos secciones iguales del marco, utilice cuatro tornillos de 4 mm x 15 mm con tuercas de ala y ocho arandelas de 4 mm x 10 mm para atornillar los perfiles L angulares de aluminio en los extremos (Figura 1A, flechas verdes).
- Utilice cinco piezas de material de poliestireno extruido (XPS) (500 mm x 200 mm x 150 mm), diez tornillos de 10 mm x 170 mm con tuercas hexagonales y veinte arandelas de 10 mm x 50 mm para preparar cinco flotadores de poliestireno extruido (500 mm x 200 mm x 150 mm cada uno). Fije los flotadores al marco en cinco puntos que se muestran en el dibujo técnico (Figura 1A).
- Perfore agujeros en el marco (vea las flechas rojas en la Figura 1A que marcan las posiciones y distancias de los orificios para las cadenas). Fije las cadenas de acero de 12 m (diámetro de alambre de 3 mm, eslabón interior de 5,5 mm x 26 mm) para las botellas de incubación a los orificios del marco utilizando ganchos de carabina de acero (50 mm x 5 mm). Proporcione a cada cadena pares de ganchos de presión (50 mm x 5 mm) para instalar las botellas de incubación a la profundidad deseada de acuerdo con el diseño experimental. En este caso, estaban sentados a 1 m y 2 m de profundidad.
- Para el anclaje, llene el cubo de 15 L con concreto. Inserte un perno ocular en el concreto y deje que se seque sin ser molestado. Sujete la cadena de acero de 5 m al gancho. Asegure el ancla al orificio preperforado en la proa de la barcaza (marcado con flechas amarillas en la Figura 1A, B).
NOTA: Los dibujos técnicos con la descripción del ensamblaje se muestran en la Figura 1A-C. La Figura 2 muestra la foto de la barcaza experimental ensamblada. La figura 3 muestra la fijación de las botellas de incubación a la cadena.

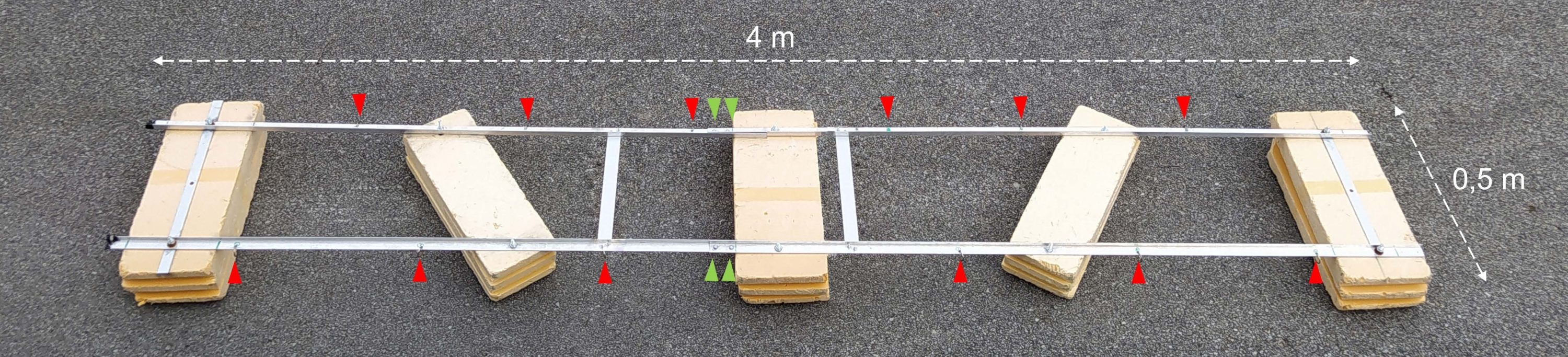
Figura 2: Barcaza experimental ensamblada. Fotografía de la barcaza experimental montada. Las puntas de flecha rojas muestran los orificios para la fijación de cadenas con botellas de incubación. Las puntas de flecha verdes apuntan a donde se unen las dos mitades del flotador. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}

Figura 3: Botellas de incubación. Foto de dos pares de botellas de incubación oscuras y claras colgando a una profundidad de 1 m. Un par de botellas contiene la muestra de esteras microbianas intactas que aún crecen en la piedra (punta de flecha roja). El segundo es la botella en blanco con el agua del lago desde la profundidad respectiva. Una punta de flecha amarilla apunta al punto del sensor de oxígeno unido a la pared interna de la botella de incubación. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}
2. Instalación en campo
- Se sugiere el uso de un kayak inflable para la colocación de barcazas y la realización de experimentos, ya que es fácilmente transportable.
- Seleccione un lugar con una profundidad ideal para anclar el flotador. Elija la profundidad de modo que las botellas de incubación inferiores estén al menos 2 m por encima del fondo para evitar perturbar el sedimento en la columna de agua alrededor de las botellas de incubación.
- Coloque la barcaza ensamblada detrás de la popa del barco. Baje con cuidado el ancla a lo largo del costado del barco y alinéela para que cuelgue ligeramente por debajo de la superficie del agua para que el flotador pueda ser fácilmente remolcado junto con el ancla hasta el lugar con la profundidad requerida.
- Desata el ancla del barco, bájala hasta el fondo y asegura la barcaza a la cadena del ancla.
- Asegure las cadenas para fijar las botellas de incubación a la barcaza.
3. Preparación del biberón de incubación
- Utilice las botellas transparentes de cuello ancho de 0,5 L con sellos herméticos al gas.
NOTA: Es posible ajustar el tamaño de las botellas, pero recuerde aumentar la flotación de la barcaza con más láminas de poliestireno también. La barcaza descrita aquí puede transportar de forma fiable 24 botellas de vidrio de 0,5 L. - Conecte los puntos del sensor óptico de oxígeno a la pared interna de cada botella.
- Agregue una capa opaca a las botellas de tratamiento oscuras envolviéndolas con cinta aislante negra.
- Corte un pequeño agujero en el lugar con el sensor óptico. Para evitar que la luz entre en la botella, haga que el orificio sea ligeramente más pequeño que el diámetro del sensor.
NOTA: Cualquier capa opaca que impida que la luz entre en la botella también funcionará. La ventaja de la cinta eléctrica negra es que resiste la abrasión y no se despega en el agua.
4. Recogida y manipulación de muestras
NOTA: Los buzos llevan a cabo la recolección manual de muestras en aguas más profundas. En aguas poco profundas, se puede hacer snorkeling o vadeando.
- Coloque las botellas de incubación en la caja portátil.
- Bucea con la caja a la profundidad respectiva. Evite perturbar el sedimento en el agua circundante.
- Llene las botellas de incubación con las muestras cuidadosamente. Trate de perturbar la biomasa de la muestra lo menos posible, por ejemplo, utilizando pinzas largas. Si las esteras microbianas crecen en una superficie sólida, como una piedra pequeña, transfiera cuidadosamente toda la piedra con biomasa intacta a la botella.
NOTA: Evite recolectar piedras grandes cuando las esteras de muestreo crezcan en piedras; las botellas de vidrio pueden romperse durante una manipulación adicional. - Llene un par de botellas claras/oscuras con agua limpia de las profundidades respectivas para que sirvan como controles en blanco.
NOTA: Las botellas sin la muestra de perifitón sirven como control para determinar la producción/consumo de oxígeno de los organismos del agua ambiental. Asegura que la productividad primaria neta o bruta calculada del perifitón sea imparcial. - Asegúrese de que el agua en todas las botellas de incubación esté limpia y no contenga sedimentos molestos.
- Cierre las botellas y llévelas al barco anclado a la barcaza flotante.
5. Medición de la productividad primaria
NOTA: La persona sentada en el bote toma la caja del buzo y realiza los siguientes pasos.
- Fije los dos primeros pares de botellas de incubación a los ganchos de presión de la primera cadena.
- Mida la concentración inicial de oxígeno en cada botella usando el medidor de oxígeno de fibra óptica. Conecte el cable óptico del medidor al sensor de oxígeno montado dentro de la botella e inmediatamente (en unos segundos) lea la concentración deO2 sin contacto (a través de la pared de la botella). Registre el valor medido.
NOTA: El tiempo de manipulación es corto; Desde llevar las botellas de los buzos hasta el ajuste inicial hasta la profundidad respectiva, solo toma unos minutos. - Inmediatamente después, baje con cuidado la cadena con las botellas adheridas al agua. Asegúrese de que las botellas de incubación se colocan a la misma profundidad que la biomasa que se colocó en ellas fue muestreada.
- Haga otra medición desde el barco después de 1 h (ver NOTA a continuación). Tire cuidadosamente de cada cadena con las botellas en el bote, lea el valor de oxígeno conectando el cable óptico al sensor y baje las muestras al agua nuevamente.
NOTA: Ajuste el tiempo entre mediciones individuales durante la incubación de acuerdo con la intensidad de la productividad/consumo deO2 de las muestras para evitar la sobresaturación de botellas. - Repita este procedimiento al menos cuatro o cinco veces con todos los pares de botellas.
NOTA: La configuración completa del experimento en el campo se muestra en la Figura 4.

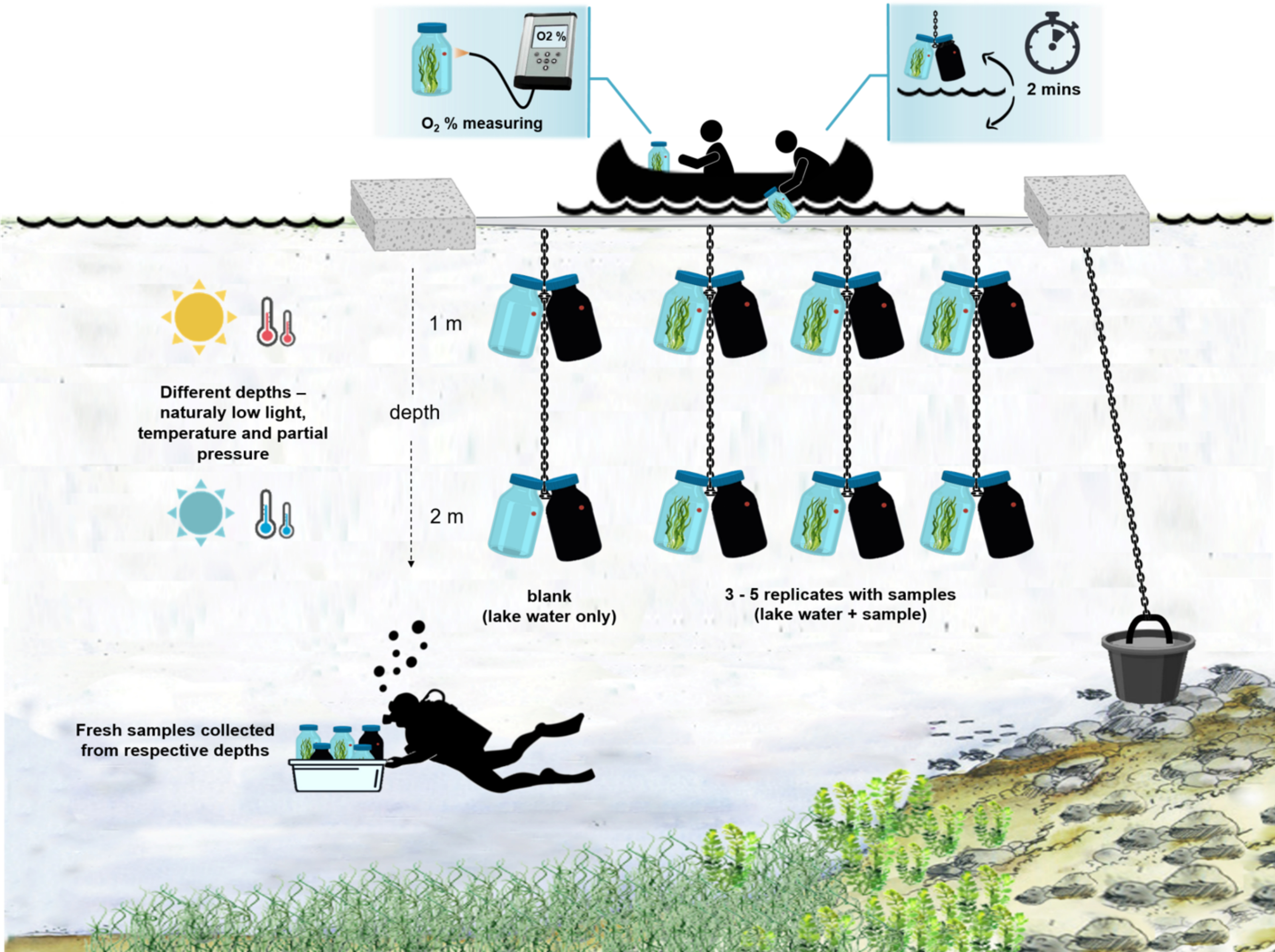
Figura 4: Esquema de la configuración experimental en el campo. Ilustración de la barcaza experimental anclada en la superficie del lago. Las botellas de incubación (0,5 L) con biomasa microbiana se cuelgan a dos profundidades diferentes (1 m y 2 m). Los buzos recolectaron muestras de esteras microbianas directamente en las botellas de incubación a las profundidades apropiadas. La concentración de oxígeno en botellas individuales se mide desde el barco. Las botellas se sacan del agua. El valor de concentración de oxígeno se mide en pocos segundos conectando un cable óptico al sensor de oxígeno. Las botellas se bajan cuidadosamente de nuevo en el agua. Todo el procedimiento de medición de dos pares de botellas de incubación desde dos profundidades toma ~ 2 min. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}
6. Análisis de muestras
- Después del final de las mediciones, tome las muestras directamente de las botellas y transfiera la biomasa de la estera microbiana a los pequeños matraces de plástico. Si las esteras crecen sobre sustratos sólidos (por ejemplo, piedras), frótelas con un cepillo de dientes o un cuchillo pequeño.
- En el laboratorio, cada réplica de filtro a través de filtros de fibra de vidrio prepesados para determinar el peso seco9.
7. Análisis de datos
- Durante el período de incubación, mida la concentración de oxígeno en las botellas claras y oscuras y compárela con la concentración de oxígeno en la columna de agua cuando se llenen las botellas.
NOTA: El cambio en el oxígeno en la botella de luz a lo largo del tiempo es el resultado combinado de la productividad bruta del ecosistema (GEP) y la respiración de todos los organismos en la botella (autótrofos y heterótrofos del agua ambiental y la comunidad perifítica). La disminución de oxígeno en la botella oscura mide las pérdidas respiratorias tanto de autótrofos como de heterótrofos. El cambio en la concentración de oxígeno en el control (es decir, botellas sin el perifitón) es solo el producto de organismos heterótrofos o autótrofos en el agua ambiente. La productividad y la respiración del perifitón se estimaron restando la productividad del agua ambiente y la respiración medidas en botellas de incubación en blanco. - Anote la concentración deO2 en porcentaje de saturación de oxígeno (es decir, calibre los sensores de oxígeno al 0% y al 100% de saturación de oxígeno). Antes de estimar la productividad primaria, convierta los datos sin procesar (
 ) a alguna unidad razonable.
) a alguna unidad razonable.
NOTA: En este estudio, los datos se convierten a mmol deO2 por gramo de materia orgánica (OM) de la masa de perifitón en la base de peso seco, según Benson y Krause10.- Calcula la conversión usando la ecuación (1):
 (1)
(1)
Donde Cp es la concentración de O2 en agua (mg (O 2) L-1) cuando está completamente saturada por O2, V Bottle es el volumen de la botella en mL, VStones es el volumen ocupado por la piedra en mL, es el peso de la masa de perifitón en g y32 es el peso molar de O2. - Calcula Cp usando la ecuación (2):
 (2)
(2)
Donde Cs es la concentración estándar deO2, P es la presión atmosférica en la superficie del lago, Pw es la presión parcial del vapor de agua en la superficie del lago y ω es la densidad del agua. - Calcule Cs, ω, P y Pw a partir de ecuaciones empíricas previamente definidas (3-6) cuando se conocen la elevación del lago (h en km) y la temperatura del agua en la superficie del lago (t en °C):
 (3)
(3) (4)
(4) (5)
(5) (6)
(6)
NOTA: A partir de las ecuaciones (1-6), es evidente que la concentración deO2 calculada es más precisa para profundidades poco profundas. A medida que aumenta la profundidad, la concentración calculada se vuelve más sesgada en términos de concentración absoluta. Es óptimo cuando se conoce la tasa de cambio de concentración de oxígeno a lo largo de la profundidad para cada lago, de modo que la concentración absoluta deO2 se puede corregir si es necesario. Una vez que se calcula la concentración deO2 , su cambio en el tiempo se puede utilizar para calcular dos flujos diferentes deO2 en dos condiciones diferentes. En condiciones de luz, la productividad neta del ecosistema (NEP) es directamente proporcional al cambio en la concentración deO2 a lo largo del tiempo (ver más abajo). El término "ecosistema" se utiliza aquí para denotar que el perifitón está compuesto de organismos autótrofos y heterótrofos. En la oscuridad, el cambio en la concentración deO2 a lo largo del tiempo es proporcional a la suma de las pérdidas respiratorias de organismos autótrofos y heterótrofos, definiendo así la respiración del ecosistema (RE). La diferencia entre NEP y RE define la productividad bruta de los ecosistemas (GEP). Si las pérdidas respiratorias de la parte heterótrofa de la comunidad son insignificantes, la GEP se convierte en igual a la productividad bruta del ecosistema.
- Calcula la conversión usando la ecuación (1):
- Determine la tasa de cambio de la concentración deO2 a lo largo del tiempo mediante regresión polinómica de tercer grado, como se muestra en la ecuación (7).
 (7)
(7)
Donde A 0 es la concentración deO2 en el tiempo cero y A 1-A 2 son coeficientes de regresión polinómica.
NOTA: La función polinómica se utiliza porque puede servir como una aproximación de cualquier ecuación diferencial. Por lo tanto, no es necesario conocer la relación funcional precisa entre la concentración deO2 y el tiempo. Por lo tanto, cualquier suposición asociada con la relación funcional (por ejemplo, linealidad) no necesita ser controlada. Por definición, el segundo término de regresión polinómica, A 1 define la tasa de cambio de la concentración deO2 en el tiempo cero (es decir, la tasa instantánea), que es independiente de A0 y, por lo tanto, la concentración absoluta de O2 en el tiempo cero. Por esa razón, la estimación del flujo de O2 no se ve afectada por el sesgo en los cálculos de concentración absoluta deO2 causado por el cambio de presión a través del gradiente de profundidad. A 1 tiene unidadesO2 (mmol g(OM)-1) por tiempo (1 h en este estudio).- Calcule A1, calculado por separado para botellas con y sin exposición a la luz y que contienen biomasa microbiana (es decir, V Stones > 0) y para botellas de control con y sin exposición a la luz y que contienen agua libre (es decir, VStones = 0).
NOTA: Asignando estas diferentes notaciones de coeficientes de regresión , , , y , respectivamente,
de coeficientes de regresión , , , y , respectivamente, 

 la ecuación (8) para el cálculo de productividad GEP se puede escribir como:
la ecuación (8) para el cálculo de productividad GEP se puede escribir como: (8.1)
(8.1) (8.2)
(8.2) (8.3)
(8.3)
El término define la productividad neta del ecosistema (Figura 5A; es decir, productividad neta de oxígeno), y el término
 representa la suma de la respiración autótrofa y heterótrofa (Figura 5B; RE, es decir, suponiendo que ambas respiraciones son similares en condiciones de oscuridad y luz).
representa la suma de la respiración autótrofa y heterótrofa (Figura 5B; RE, es decir, suponiendo que ambas respiraciones son similares en condiciones de oscuridad y luz). - Reste R de NEP para obtener GEP (Figura 5C).
NOTA: La ecuación (8) asume implícitamente que y son ambos positivos, y ambos son negativos. Si es positivo, verifique cuidadosamente los datos brutos para valores atípicos. pueden ser teóricamente negativos porque las pérdidas respiratorias causadas por la actividad heterótrofa pueden ser mayores que la actividad fotosintética.
- Calcule A1, calculado por separado para botellas con y sin exposición a la luz y que contienen biomasa microbiana (es decir, V Stones > 0) y para botellas de control con y sin exposición a la luz y que contienen agua libre (es decir, VStones = 0).
Resultados

Figura 5: Productividad neta y bruta del ecosistema de esteras microbianas durante el día. (A) Productividad del ecosistema de la red de botellas ligeras: datos del curso del tiempo de la productividad neta de oxígeno de las esteras microbianas de las botellas de luz. El cambio de concentración de oxígeno en las botellas de incubación se midió...
Discusión
La metodología descrita en este trabajo se basa en el principio de la técnica de oxígeno de botella clara y oscura en combinación con la técnica no invasiva de medir la concentración deO2 utilizando sensores ópticos de oxígeno. Este sistema permite la medición paralela de diferentes configuraciones de incubación, ya que la fibra óptica para medirO2 se puede mover rápidamente de una botella a otra. Las comunidades bentónicas de diversas profundidades pueden diferir en composición taxon?...
Divulgaciones
Los autores confirman que no tienen conflictos de intereses que revelar.
Agradecimientos
Este estudio fue apoyado por la Fundación Checa de Ciencias (GACR 19-05791S), RVO 67985939, y por el CAS dentro del programa de la Estrategia AV 21, Ahorro y recuperación de tierras. Muchas gracias a Ondřej Sihelský por tomar las fotos en el campo, sin él, la filmación habría sido un infierno. El proyecto no sería posible sin una estrecha cooperación con las empresas, Palivový Kombinát Ústí s.p. y Sokolovská Uhelná, que proporcionaron acceso a las localidades estudiadas.
Materiales
| Name | Company | Catalog Number | Comments |
| Aluminum angle L profile 40 x 40 mm x 3 mm, length 2,000 mm | |||
| Aluminum flat bar 40 x 3 x 350 mm | |||
| Bucket 15 L with concrete infill | |||
| Carabine hook with screw lock 50 x 5 mm | |||
| electric tape black | |||
| Extruded polystyrene (XPS) material 500 x 200 x 150 mm | |||
| Fibox 3 LCD trace | PreSens Precision Sensing GmbH | stand-alone fiber optic oxygen meter | |
| Hondex PS-7 Portable Depth Sounder | Hondex - Honda Electronics | to measures distances through water - to bottom depth measurement; https://www.honda-el.net/industry/ps-7e | |
| KORKEN - glass tight-seal jar 0.5 L | IKEA | incubation bottles; https://www.ikea.com/cz/en/p/korken-jar-with-lid-clear-glass-70213545/ | |
| metal hook | |||
| Oxygen Sensor Spot SP-PSt3-NAU-D5 | PreSens Precision Sensing GmbH | non-invasive optical oxygen sensor for measurements under Real Conditions | |
| SCOUT infantable canoe | GUMOTEX | https://www.gumotexboats.com/en/scout-standard#0000-044667-021-13/11C | |
| Screw 10 x 170 mm with hexagonal nuts | |||
| Screw 4 x 15 mm with hexagonal nuts | |||
| Screw 4 x 15 mm with wing nuts | |||
| Snap hooks 50 x 5 mm | |||
| Steel Carabine hook 50 x 5 mm | |||
| Steel chain with wire diameter 3 mm, inside link 5.5 x 26 mm | |||
| Steel chain, 5 m | |||
| toothbrush | |||
| tweezer | |||
| Washer 10 x 50 mm | |||
| Washer 4 x 10 mm | |||
| Washer 4 x 10 mm |
Referencias
- Blachart, J. L., et al. Potential consequences of climate change for primary production and fish production in large marine ecosystems. Philosophical Transactions of the Royal Society B: Biological Sciences. 367 (1605), 2979-2989 (2012).
- Howarth, R. W., Michaels, A. F., Sala, O. E., Jackson, R. B., Mooney, H. A., Howarth, R. W. The Measurement of primary production in aquatic ecosystems. Methods in Ecosystem Science. , 72-85 (2000).
- Vadenbecouer, Y. E. G., Peterson, M. J., Vander, Z., Kalff, J. Benthic algal production across lake size gradients: Interactions among morphometry, nutrients, and light. Ecology. 89 (9), 2542-2552 (2008).
- Reimer, A., Landmann, G., Kempe, S. Lake Van, eastern Anatolia, hydrochemistry and history. Aquatic Geochemistry. 15 (1), 195-222 (2009).
- Cantonati, M., Lowe, R. L. Lake benthic algae: toward an understanding of their ecology. Freshwater Sciences. 33 (2), 475-486 (2014).
- Gaarder, T., Gran, H. H. Investigation of the production of plankton in the Oslo Fjord. Rapports et Proces-verbaux des Réunions. Conseil International pour l'Éxploration de la Mer. 42, 1-48 (1927).
- Hall, R. O., Thomas, S., Gaiser, E. E., Fahey, T. J., Knapp, A. K. Measuring Freshwater Primary Productivity and Respiration. Principles and Standards for Measuring Primary Productivity. , (2007).
- Howart, R., Michaels, A. Chapter 6 The Measurement of Primary Production in Aquatic Ecosystems. Springer Science and Business Media LLC. , (2000).
- Kopáček, J., Hejzlar, J. Semi-micro determination of total phosphorus in soils, sediments, and organic materials: a simplified perchloric acid digestion procedure. Communications in Soil Science and Plant Analysis. 26 (11-12), 1935-1946 (1995).
- Benson, B. B., Krause, D. The concentration and isotopic fractionation of oxygen dissolved in freshwater and seawater in equilibrium with the atmosphere1. Limnology and Oceanography. 29 (3), 620-632 (1984).
- Dodds, W. K., Biggs, B. J., Lowe, R. L. Photosynthesis-irradiance patterns in benthic microalgae: variations as a function of assemblage thickness and community structure. Journal of Phycology. 35 (1), 42-53 (1999).
- Bott, T. L., et al. An evaluation of techniques for measuring periphyton metabolism in chambers. Canadian Journal of Fisheries and Aquatic Sciences. 54 (3), 715-725 (1997).
- Blankenship, R. E. Structural and functional dynamics of photosynthetic antenna complexes. Proceedings of the National Academy of Sciences. 112 (45), 13751-13752 (2015).
- Hawes, I., Schwartz, A. -. M. Photosynthesis in an extreme shade environment, benthic microbial mats from Lake Hoare, a permanently ice-covered Antarctic lake. Journal of Phycology. 35 (3), 448-459 (1999).
- Aristegui, J., et al. Planktonic primary production and microbial respiration measured by 14C assimilation and dissolved oxygen changes in coastal waters of the Antarctic peninsula during austral summer: Implications for carbon flux studies. Marine Ecology-Progress Series. 132, 191-201 (1996).
- Steemann-Nielsen, C. The use of radioactive carbon (14C) for measuring organic production in the sea. Journal of Experimental Marine Biology and Ecology. 18 (2), 117-140 (1952).
- Sanz-Martín, M., et al. Relationship between carbon-and oxygen-based primary productivity in the Arctic Ocean, svalbard archipelago. Frontiers in Marine Science. 6, 468 (2019).
- Nielsen, E. S. Measurement of the production of organic matter in the sea by means of carbon-14. Nature. 167 (4252), 684-685 (1951).
- Jönsson, B. A 14C-incubation technique for measuring microphytobenthic primary productivity in intact sediment cores. Limnology and Oceanography. 36 (7), 1485-1492 (1991).
- Bender, M. L., et al. A comparison of four methods for determining planktonic community production. Limnology and Oceanography. 32 (5), 1085-1098 (1987).
- Šimek, K., et al. Spatio-temporal patterns of bacterioplankton productivity and community composition related to phytoplankton composition and protistan bacterivory in a dam reservoir. Aquatic Microbial Ecology. 51 (3), 249-262 (2008).
Reimpresiones y Permisos
Solicitar permiso para reutilizar el texto o las figuras de este JoVE artículos
Solicitar permisoThis article has been published
Video Coming Soon
ACERCA DE JoVE
Copyright © 2025 MyJoVE Corporation. Todos los derechos reservados