
Method Article
Microscopía de superresolución del complejo sinaptonémico dentro de la línea germinal Caenorhabditis elegans
En este artículo
Resumen
La microscopía de superresolución puede proporcionar una visión detallada de la organización de los componentes dentro del complejo sinaptonémico en la meiosis. Aquí, demostramos un protocolo para resolver proteínas individuales del complejo sinaptonémico Caenorhabditis elegans .
Resumen
Durante la meiosis, los cromosomas homólogos deben reconocerse y adherirse entre sí para permitir su correcta segregación. Uno de los eventos clave que asegura la interacción de cromosomas homólogos es el ensamblaje del complejo sinaptonémico (SC) en la profase meiótica I. A pesar de que hay poca homología de secuencia entre los componentes proteicos dentro del SC entre diferentes especies, la estructura general del SC ha sido altamente conservada durante la evolución. En las micrografías electrónicas, el SC aparece como una estructura tripartita, similar a una escalera, compuesta de elementos o ejes laterales, filamentos transversales y un elemento central.
Sin embargo, la identificación precisa de la localización de componentes individuales dentro del complejo por microscopía electrónica para determinar la estructura molecular del SC sigue siendo un desafío. Por el contrario, la microscopía de fluorescencia permite la identificación de componentes proteicos individuales dentro del complejo. Sin embargo, dado que el SC tiene solo ~ 100 nm de ancho, su subestructura no puede resolverse mediante microscopía de fluorescencia convencional limitada por difracción. Por lo tanto, la determinación de la arquitectura molecular del SC requiere técnicas de microscopía óptica de súper resolución, como la microscopía de iluminación estructurada (SIM), la microscopía de agotamiento por emisión estimulada (STED) o la microscopía de localización de molécula única (SMLM).
Para mantener la estructura y las interacciones de los componentes individuales dentro del SC, es importante observar el complejo en un entorno cercano a su entorno nativo en las células germinales. Por lo tanto, demostramos un protocolo de inmunohistoquímica e imagen que permite el estudio de la subestructura del SC en tejido germinal intacto, extruido de Caenorhabditis elegans con microscopía SMLM y STED. La fijación directa del tejido al cubreobjetos reduce el movimiento de las muestras durante la obtención de imágenes y minimiza las aberraciones en la muestra para lograr la alta resolución necesaria para visualizar la subestructura del SC en su contexto biológico.
Introducción
Reducir el número de cromosomas a la mitad durante la meiosis es clave para generar una progenie saludable en los organismos que se reproducen sexualmente. Para lograr esta reducción en el número de cromosomas, los cromosomas homólogos deben emparejarse y segregarse durante la meiosis I. Para garantizar la segregación precisa de los cromosomas homólogos, las células germinales se someten a una profase I extendida, durante la cual los cromosomas homólogos se emparejan, hacen sinapsis y se recombinan para generar vínculos físicos entre homólogos1. El SC se ha convertido en la estructura central clave para regular la correcta progresión a través de la profasemeiótica 2.
El SC es un complejo cuya estructura general se conserva evolutivamente, aunque hay poca homología entre sus componentes proteicos. El SC se identificó por primera vez en micrografías electrónicas como una estructura tripartita, similar a una escalera, que consta de dos elementos laterales o ejes, una región central formada por filamentos transversales y un elemento central 3,4. Determinar la organización de los componentes individuales dentro del complejo es clave para avanzar en nuestra comprensión del papel del SC durante la profase meiótica.
El organismo modelo C. elegans es ideal para estudiar la estructura y función del SC ya que sus líneas germinales contienen un gran número de núcleos meióticos con SCs completamente ensamblados5. Estudios genéticos y bioquímicos han revelado que los ejes cromosómicos están formados por tres complejos distintos de cohesina6,7 y cuatro proteínas de dominio HORMA llamadas HTP-1/2/3 y HIM-3 7,8,9,10,11 en C. elegans. En la región central del SC, se han identificado seis proteínas que contienen dominios de bobina enrollada hasta la fecha 12,13,14,15,16,17. Para salvar la distancia entre los dos ejes, SYP-1, -5 y -6 dimerizan de manera cara a cara (Figura 1), mientras que tres proteínas adicionales estabilizan su interacción en el elemento central16,17,18,19.
Obtener información detallada sobre la organización de estas proteínas es esencial para comprender las muchas funciones del SC durante la meiosis. Dado que el ancho de la región central del SC es solo ~ 100 nm, su subestructura no puede resolverse mediante microscopía de fluorescencia limitada por difracción. Sin embargo, la visualización de componentes dentro de una estructura de este tamaño es fácilmente alcanzable por microscopía de súper resolución. De hecho, la microscopía de iluminación estructurada (SIM), la microscopía de expansión 20, la microscopía de agotamiento de emisión estimulada (STED) 21 y la microscopía de localización de molécula única (SMLM)22,23 han surgido como herramientas esenciales para estudiar la arquitectura molecular del SC en especies 16,24,25,26,27,28,29, 30.
Para superar el límite de resolución, la microscopía STED se basa en superponer el punto limitado por difracción de la luz de emisión con un haz en forma de rosquilla del láser STED, que teóricamente constriñe la función de dispersión puntual hasta dimensiones moleculares31,32. Sin embargo, la resolución que es prácticamente alcanzable por STED dentro de muestras biológicas permanece en el rango de unas pocas decenas de nanómetros en xy33.
Se puede obtener una resolución aún mayor en muestras biológicas con técnicas SMLM. SMLM aprovecha las propiedades de parpadeo de fluoróforos específicos para resolver objetos a nivel de subdifracción separando fluoróforos espacialmente superpuestos en el tiempo. Luego, la muestra se visualiza repetidamente para capturar diferentes subconjuntos de fluoróforos. La posición de los fluoróforos dentro de la muestra se determina ajustando la función de dispersión puntual (PSF) a las señales obtenidas en todas las imágenes, que pueden resolver estructuras de hasta 15 nm23,34.
En conjunto, las imágenes localizadas codifican las posiciones de todos los fluoróforos. La resolución de SMLM está determinada por la densidad de etiquetado y las características de parpadeo del fluoróforo. De acuerdo con el criterio de Nyquist-Shannon, es imposible resolver de manera confiable objetos que son menos del doble de la distancia promedio de etiqueta a etiqueta. Por lo tanto, se necesita una alta densidad de etiquetado para imágenes de alta resolución. Para el SC en C. elegans, se puede lograr una alta densidad de etiquetado mediante el uso de etiquetas de epítopos unidas a sitios específicos de proteínas endógenas utilizando la edición del genoma. Las etiquetas de epítopos pueden teñirse a una alta densidad utilizando anticuerpos monoclonales específicos con altas afinidades19,30. Al mismo tiempo, el ciclo de fluoróforos individuales debe ser lo suficientemente corto como para garantizar que los fluoróforos espacialmente superpuestos no se capturen al mismo tiempo35.
Debido a estos dos requisitos, resolver la estructura de grandes complejos macromoleculares como el SC requiere obtener imágenes de un número suficientemente grande de imágenes y, por lo tanto, puede llevar varias horas. La trampa de los largos tiempos de obtención de imágenes es que las muestras tienden a desplazarse debido al movimiento de la etapa o a pequeñas corrientes dentro del búfer de muestra; Incluso pequeños movimientos del orden de 10 nm son perjudiciales a la resolución de nm y deben corregirse. Sin embargo, los métodos de corrección de deriva comúnmente utilizados no son lo suficientemente robustos como para superponer con precisión imágenes de dos canales fotografiados secuencialmente36. Esto es problemático porque las preguntas biológicas a menudo requieren la detección precisa y la localización de múltiples objetivos dentro de la misma muestra. Para evitar estos problemas, se han desarrollado métodos como la imagen ratiométrica. La imagen ratiométrica permite la obtención simultánea de múltiples fluoróforos con espectros de excitación y emisión superpuestos, con una asignación posterior de cada señal detectada a su respectivo fluoróforo basado en la relación de intensidades en canales espectralmente distintos37,38.
Además, el estudio de la organización de complejos macromoleculares como el SC requiere información tridimensional (3D). Para lograr una superresolución en tres dimensiones (3D-SMLM), se incorpora una lente cilíndrica en la trayectoria óptica de la luz emitida que distorsiona la forma del PSF de un fluoróforo dependiendo de su distancia del plano focal. Por lo tanto, la posición precisa de un fluoróforo en el plano z se puede extrapolar analizando la forma de su señal de emisión35,39. La combinación de estos avances en SMLM permite obtener imágenes de la organización 3D de complejos macromoleculares, incluido el SC.
Protocolo
1. Preparación de soluciones y cubreobjetos
NOTA: Consulte la Tabla de materiales para obtener detalles relacionados con todos los materiales y reactivos y la Tabla 1 para la composición de las soluciones utilizadas en este protocolo.
- Cubreobjetos recubiertos de poli-L-lisina
- Preparar poli-L-lisina al 0,01% (p/v) (ver Tabla 1).
- Lave un cubreobjetos de precisión (24 mm de diámetro; 0,17 ± 0,005 mm, nº 1,5) en etanol durante 10-30 min. Enjuague el cubreobjetos conddH2Opara eliminar el etanol y deje que el cubreobjetos se seque a temperatura ambiente.
- Limpie con plasma el cubreobjetos con un limpiador de plasma.
NOTA: La limpieza por plasma aumenta la hidrofilicidad del cubreobjetos y facilita los siguientes pasos. Si no se dispone de un limpiador de plasma, se puede omitir este paso, aunque esto puede requerir ajustar el volumen y / o la concentración de la solución de poli-L-lisina. Esta modificación no ha sido probada. - Coloque una gota (120 μL) de poli-L-lisina al 0,01% (p/v) en el cubreobjetos. Incubar durante 10 min a temperatura ambiente.
- Después de la incubación, enjuague el cubreobjetos enddH2Oy séquelo a temperatura ambiente. Conservar a 4 °C hasta 1 mes.
- F(ab')2 fragmentos conjugados con colorantes orgánicos fluorescentes
- Agregue el siguiente orden a un tubo de PCR: 10 μL de 0.6-0.7 mg / ml de fragmento F(ab')2 en PBS, 1 μL de 0.1 M NaHCO 3 (pH8.3 ) y 1 μL de fluoróforo reactivo de éster de succinimidil (NHS) de 1 mM en DMSO (relación molar de F(ab')2: colorante es ~ 1:17). Mezclar bien pipeteando hacia arriba y hacia abajo.
- Incubar durante 1 h a temperatura ambiente.
- Separe el fragmento F(ab')2 del colorante reactivo libre restante utilizando una columna de desalinización (7K MWCO) siguiendo las especificaciones del fabricante. Utilice 1x PBS para el equilibrio de la columna y la elución del fragmento F(ab')2 etiquetado.
- Conservar el fragmento F(ab')2 etiquetado a 4 °C durante un máximo de 3 meses.
NOTA: No se han probado tiempos de almacenamiento superiores a 3 meses.
2. Disección y fijación
NOTA: Los procedimientos de disección y fijación se modifican a partir de los procedimientos recomendados anteriormente16,40 para obtener muestras óptimas para microscopía de superresolución.
- Disección
- Elija gusanos C. elegans de la misma edad (cultivados a 20 ° C para este estudio) en una gota de 30 μL de EBTT (1x tampón de huevo41 con 0.2% de detergente no iónico, Tabla 1) en un cubreobjetos (22 mm x 22 mm, No. 1). Coloque el cubreobjetos en un portaobjetos de vidrio para facilitar la manipulación. Lavar con 30 μL de EBTT pipeteando hacia arriba y hacia abajo varias veces. Retire 30 μL de la solución para dejar una gota de 30 μL en el cubreobjetos.
NOTA: Se deben agregar pequeñas cantidades de detergente no iónico a todas las soluciones en las que se pipetean los gusanos para evitar que los gusanos se peguen a las puntas de plástico. - Use una hoja de bisturí para cortar las cabezas y/o las colas de los gusanos para extruir la gónada (Figura 2A).
- Elija gusanos C. elegans de la misma edad (cultivados a 20 ° C para este estudio) en una gota de 30 μL de EBTT (1x tampón de huevo41 con 0.2% de detergente no iónico, Tabla 1) en un cubreobjetos (22 mm x 22 mm, No. 1). Coloque el cubreobjetos en un portaobjetos de vidrio para facilitar la manipulación. Lavar con 30 μL de EBTT pipeteando hacia arriba y hacia abajo varias veces. Retire 30 μL de la solución para dejar una gota de 30 μL en el cubreobjetos.
- Fijación
- Pipeta 30 μL de solución fijadora (Tabla 1) en la gota de los gusanos disecados y pipeta arriba y abajo para mezclar.
NOTA: Subir y bajar varias veces puede ayudar a liberar más gónadas. - Arregle durante exactamente 1 minuto después de agregar la solución fijadora.
- Detenga la fijación transfiriendo los gusanos a un tubo de PCR lleno de TBST (Tabla 1). Transfiera los gusanos en el menor volumen posible (~ 15 μL).
- Descienda el tubo de PCR en una mini centrífuga de sobremesa (2.000 × g, 10 s). Retire el sobrenadante y lave 2 veces con 200 μL de TBST cada una.
- Lavar con 200 μL de PBST (Tabla 1) durante 5-10 min. Repita los pasos 2.1.1 a 2.2.5 para un máximo de cuatro muestras mientras mantiene las muestras disecadas en hielo.
NOTA: Si procesa más de cuatro muestras, continúe con los pasos 2.2.6 a 2.2.7 después de cada cuatro muestras para garantizar que la fijación de la muestra permanezca constante en todas las muestras. - Girar las muestras diseccionadas en una mini centrífuga de sobremesa (2.000 × g, 10 s), retirar el PBST y añadir 50-100 μL de metanol frío (-20 °C).
PRECAUCIÓN: El metanol es tóxico. Use equipo de protección y evite la inhalación. - Mezclar pipeteando hacia arriba y hacia abajo y dejar las muestras en metanol durante 30-60 s. Lavar las muestras 2 veces en 200 μL de PBST.
NOTA: Si se procesan más de cuatro muestras, proceda a la disección de las muestras restantes (pasos 2.1.1 a 2.2.7). - Lavar las muestras una tercera vez con 200 μL de PBST.
- Pipeta 30 μL de solución fijadora (Tabla 1) en la gota de los gusanos disecados y pipeta arriba y abajo para mezclar.
3. Incubaciones de anticuerpos
- Bloqueante
- Bloquear las muestras en 1x solución de bloqueo (Tabla 1) durante 45-60 min a temperatura ambiente.
NOTA: El tiempo de incubación puede variar de 30 minutos a temperatura ambiente a varios días a 4 °C (la prueba se realizó hasta 3 días).
- Bloquear las muestras en 1x solución de bloqueo (Tabla 1) durante 45-60 min a temperatura ambiente.
- Solución primaria de anticuerpos
- Diluir anticuerpos anti-HTP-3 (pollo42) y anti-HA (ratón) (o los anticuerpos de elección) a las soluciones de trabajo (1:250 para SMLM y 1:1.000 para muestras de microscopía STED) en 1x solución de bloqueo.
NOTA: Los anticuerpos utilizados para marcar muestras de SMLM están más concentrados que para las muestras de ETS, ya que se recomienda una mayor densidad de etiquetado para la microscopía SMLM. - Haga girar las muestras en una mini centrífuga de sobremesa (2.000 × g, 10 s), retire el tampón de bloqueo y agregue 30-50 μL de la solución de anticuerpos primarios. Incubar durante la noche a 4 °C (preferiblemente) o durante 1-2 h a temperatura ambiente.
- Después de la incubación, lavar 3 x 5-15 min con PBST.
- Diluir anticuerpos anti-HTP-3 (pollo42) y anti-HA (ratón) (o los anticuerpos de elección) a las soluciones de trabajo (1:250 para SMLM y 1:1.000 para muestras de microscopía STED) en 1x solución de bloqueo.
- Solución de trabajo de fragmentos F(ab')2 conjugados con colorante fluorescente
- Diluir los fragmentos F(ab')2 marcados (paso 1.2.4) en las soluciones de trabajo (1:100 para SMLM y 1:1.000 para muestras de microscopía STED) en 1x solución de bloqueo.
NOTA: Para ambas técnicas de superresolución, se utilizaron pares de fluoróforos previamente informados, a saber, AlexaFluor647 / CF680 para SMLM y AlexaFluor594 / Abberior STAR635P para STED. AlexaFluor647 y STAR645P se utilizaron para etiquetar fragmentos anti-ratón (Fab')2 para apuntar al extremo C de SYP-5, y fragmentos anti-pollo (Fab')2 marcados con CF680/AlexaFluor594 para apuntar a HTP-3. - Gire las muestras en una mini centrífuga de sobremesa (2,000 × g, 10 s), retire el PBST y agregue 30-50 μL de solución de anticuerpos secundarios. Incubar durante 30 min a 2 h a temperatura ambiente (preferible) o durante la noche a 4 °C. Lavar 3 x 5-15 min con PBST.
- Diluir los fragmentos F(ab')2 marcados (paso 1.2.4) en las soluciones de trabajo (1:100 para SMLM y 1:1.000 para muestras de microscopía STED) en 1x solución de bloqueo.
4. Montaje de muestras en un cubreobjetos
- Postfijación
NOTA: Procese muestras individualmente a través de los pasos 4.1.1-4.2.1.- Gire las muestras manchadas y retire el sobrenadante. Añadir 50 μL de PBST0,2 y transferir los gusanos teñidos a un cubreobjetos No. 1 de 22 mm x 22 mm.
NOTA: Use PBST 0.2 fresco con0.2 % de detergente no iónico (Tabla 1) para este paso para evitar que los gusanos se adhieran al cubreobjetos. - Pipeta 5.7-6.3 μL de solución posfijadora sobre un cubreobjetos de poli-L-lisina.
NOTA: Los cubreobjetos de poli-L-lisina almacenados a 4 °C deben llevarse primero a temperatura ambiente. - Pipetea los gusanos disecados en el mismo volumen (5.7-6.3 μL) y transfiérelos a la gota del fijador en el cubreobjetos de poli-L-lisina (Figura 2B).
NOTA: En este y el siguiente paso, es muy importante retener el tejido disecado en el centro del cubrecubierto con poli-L-lisina. Esto es especialmente importante si se montan las muestras en un soporte personalizado para adaptarse al microscopio SMLM personalizado utilizado aquí (consulte el paso 5.1, Figura 2B). - Cubra la muestra con un cubreobjetos pequeño (12 mm de diámetro, figura 2B). Elimine el exceso de líquido con un pequeño trozo de papel de filtro (Figura 2B). Fijar durante 3-5 minutos en una cámara oscura.
- Gire las muestras manchadas y retire el sobrenadante. Añadir 50 μL de PBST0,2 y transferir los gusanos teñidos a un cubreobjetos No. 1 de 22 mm x 22 mm.
- "Craqueo por congelación"
- Congele las muestras colocándolas sobre un bloque de aluminio en hielo seco (Figura 2B).
NOTA: El bloque de aluminio debe enfriarse bien en el hielo seco antes de colocar las muestras sobre él. Proceder a la postfijación de las muestras restantes (pasos 4.1.1 a 4.2.1). La muestra debe estar en hielo seco durante al menos 20 minutos o hasta 1 h antes del siguiente paso (4.2.2). - Retire el cubreobjetos más pequeño con una navaja de afeitar (Figura 2B).
NOTA: Para STED, continúe con el paso 5.2.1. Para SMLM, continúe con el paso 4.2.3. - Sumerja el cubreobjetos en un tubo cónico de 50 ml que contenga PBS helado (preferido) o metanol a -20 °C durante aproximadamente 10 s.
NOTA: La temperatura es un factor muy importante para este paso. Por lo tanto, use PBS que esté recién descongelado o mantenido en un baño de hielo / etanol. - Coloque el cubreobjetos en un pocillo de una placa de seis pocillos llena de tampón PBST. Retire el PBST de los pozos y agregue PBS fresco. Deje las muestras en PBS durante 5 minutos.
NOTA: Pipetea el PBS en el lado del pozo para evitar dañar y separar las muestras. - Lavar con PBS fresco y dejar las muestras a 4 °C hasta que se obtengan imágenes.
NOTA: Las muestras son estables hasta por 2 semanas, pero los mejores resultados se logran si las muestras se visualizan dentro de los 2 días. - Antes de obtener imágenes, evalúe la calidad del montaje de la muestra bajo un microscopio estéreo.
NOTA: Las líneas germinales montadas con éxito se unen de manera estable sin movimiento discernible en relación con el cubreobjetos. Las líneas germinales mal adheridas aletearán en la solución tampón.
- Congele las muestras colocándolas sobre un bloque de aluminio en hielo seco (Figura 2B).
5. Imágenes
- Microscopía de localización de molécula única
NOTA: Las imágenes se adquirieron en el Centro de Imágenes EMBL utilizando un microscopio de localización de molécula única hecho a medida que se construyó alrededor de un cuerpo personalizado, como se informó anteriormente.38,43, con las características únicas especificadas en el Tabla de materiales; Consulte https://www.embl.org/about/info/imaging-centre- Adquisición de calibración de perlas 3D
- Preparar un cubreobjetos de precisión (24 mm de diámetro; 0,17 ± 0,005 mm, Nº 1,5) con perlas fluorescentes adherentes de 100 nm como se describió anteriormente38,44.
- Coloque la muestra de calibración del paso 5.1.1.1 en un portamuestras.
- Agregue una gota de aceite de inmersión en el objetivo de aceite limpio de 100x/1.5 y monte la muestra de calibración en el microscopio.
- En MicroManager 2 45,46, especifique15-20 posiciones en la muestra de calibración.
- Dentro de la ventana47 del plugin EMU, configure la adquisición de una imagen z-stack para cada una de las posiciones del paso 5.1.1.5.
NOTA: Aquí, una lente cilíndrica compuesta proporciona el astigmatismo requerido para las imágenes 3D, y se adquirieron 201 cortes z para cada posición que abarca el rango entre -1 μm y 1 μm, con un incremento de 10 nm. Se utilizó una iluminación láser de 2 kW/cm2 640 nm durante 25 ms para cada z-slice. - Adquiera las imágenes de la pila z de las perlas fluorescentes de 100 nm a través de una ruta óptica idéntica que se utilizará para adquirir imágenes de muestra en el paso 5.1.11.
- Utilizando la plataforma de análisis de microscopía de superresolución (SMAP48), genere un modelo cspline de la función experimental de dispersión puntual (PSF) que se utilizará para ajustar los datos 3D-SMLM en el paso 5.1.13.
- Prepare el portamuestras. Para el soporte personalizado utilizado aquí que utiliza un anillo magnético para crear la cámara de imágenes (Figura 2B), envuelva el anillo magnético con parafilm.
NOTA: Alternativamente, se puede usar un portaobjetos de microscopio con una cavidad de depresión cóncava para montar muestras para microscopios con portaportaobjetos. - Prepare 1 ml de tampón de imágenes44 (Tabla 1).
- Tome un cubreobjetos del paso 4.2.6 y colóquelo en el soporte hecho a medida. Fije el cubreobjetos en el soporte con el anillo magnético envuelto en parafilm (paso 5.1.2).
- Pipetear suavemente el tampón de imagen (paso 5.1.3) en la cámara creada por el anillo magnético en la parte superior de la muestra (Figura 2B). Selle la cámara con un trozo de parafilm.
- Para montar la muestra, agregue una gota de aceite de inmersión en el objetivo de aceite limpio 100x/1.5. Sin introducir aire en el aceite de inmersión, coloque suavemente el portamuestras con la muestra montada (paso 5.1.5) sobre la platina del microscopio.
NOTA: Antes de colocar la muestra en el microscopio, limpie la parte inferior del cubreobjetos con pañuelo de papel y etanol al 70%. - Usando la ventana47 del plugin EMU dentro de MicroManager 245,46, mueva la etapa piezoeléctrica hasta que se detecte la señal del láser de bloqueo de enfoque en el fotodiodo del cuadrante (QPD).
NOTA: Para mantener un enfoque fijo a lo largo del tiempo de imagen, el bloqueo del enfoque se logra mediante la reflexión interna total de un láser acoplado a fibra de infrarrojo cercano desde el cubreobjetos y la posterior detección sensible a la altura en un fotodiodo cuadrante (QPD). La señal QPD proporcionó un control de bucle cerrado de la montura piezoeléctrica de la lente del objetivo. - Adquiera una imagen de plano focal posterior con un láser de excitación de 640 nm a baja potencia (es decir, 1-5%) para confirmar la ausencia de burbujas de aire en el aceite de inmersión.
NOTA: Extraiga la muestra del escenario si se detecta una burbuja de aire. Limpie la parte inferior del cubreobjetos y el objetivo y repita los pasos 5.1.6-5.1.8. De lo contrario, proceda a bloquear el foco dentro del software EMU47. - Localice el tejido de la gónada utilizando la iluminación de campo claro. Usando una iluminación de baja intensidad de 640 nm, concéntrese en la sección del tejido que contiene muchos estiramientos SC.
NOTA: No se centre en estructuras que estén a más de 2 μm del cubreobjetos. No utilice una potencia láser mayor para localizar la muestra, ya que esto puede convertir algunos fluoróforos en un estado de parpadeo prematuro. Aquí, se utilizó 1 kW / cm2 en modo ascendente con un pulso ajustado a 1.000. - Proceda a exponer la muestra con iluminación de 640 nm a alta irradiancia (27 kW/cm2) durante ~30 s hasta que se logre una velocidad de parpadeo adecuada (Video complementario 1).
- Adquiera 200.000 fotogramas con un tiempo de exposición de 20 ms utilizando la herramienta de adquisición multidimensional de MicroManager 245,46.
- Mientras tanto, configure la activación UV utilizando la opción de activación del complemento EMU38,47 para mantener la velocidad de parpadeo deseada.
NOTA: Utilice el láser UV a una irradiancia de 3 kW/cm2 en modo ascendente con una longitud máxima de pulso establecida en 10.000. - Realice la reconstrucción y el postprocesamiento de imágenes SMLM.
NOTA: Para reconstruir imágenes a partir de datos SMLM sin procesar, consulte los métodos publicados. Los datos aquí presentados fueron procesados utilizando el software SMAP48,49. La reconstrucción de imágenes superresueltas, la asignación de canales, la corrección de derivas y el filtrado de localizaciones con poca precisión de localización y un filtro de máxima verosimilitud se realizaron en el software SMAP48.
- Adquisición de calibración de perlas 3D
- Microscopía de agotamiento por emisión estimulada
NOTA: Las imágenes se adquirieron en el sistema microscópico STED integrado equipado con un láser de luz blanca, un láser STED pulsado de 775 nm y el módulo de envejecimiento FALCON Fluorescence Lifetime IM(Tabla de materiales) en el EMBL Imaging Centre (https://www.embl.org/about/info/imaging-centre).- Coloque una gota de 20 μL de medio de montaje (Tabla de materiales) en un portaobjetos de microscopio. Tome un cubreobjetos del paso 4.2.2 y coloque la muestra suavemente sobre el portaobjetos orientado hacia el medio de montaje (figura 2B).
NOTA: Evite introducir bolsas de aire dentro del medio de montaje. - Deje que el medio de montaje se cure durante la noche.
NOTA: Imagen de las muestras al día siguiente o manténgalas a 4 °C hasta que se obtengan imágenes. - Para montar la muestra, añadir una gota de aceite de inmersión en el cubreobjetos de la muestra del paso 5.2.2. Coloque suavemente la muestra en la etapa del microscopio utilizando un objetivo de aceite 100x/1.40.
- Concéntrese en la muestra y localice el tejido de la línea germinal utilizando la iluminación de campo brillante.
- Utilizando el software del microscopio, especifique la región de interés para la que se adquirirá la imagen TauSTED.
- Seleccione los láseres de excitación y su potencia adecuada utilizada para excitar los fluoróforos utilizados en la muestra.
NOTA: Aquí, el láser de 580 nm al 4% de potencia se utilizó para obtener imágenes de fragmentos de anticuerpos secundarios F(ab')2 conjugados con AlexaFluor 594, y 635 nm al 3% de potencia para obtener imágenes de fragmentos F(ab')2 conjugados con STAR 635P. - Usando el software del microscopio, seleccione una potencia láser de agotamiento STED adecuada y configure la detección de imágenes.
NOTA: Aquí, la potencia del láser de agotamiento STED de 775 nm se estableció en 40%. El detector se utilizó en modo de conteo con un valor de ganancia de 10 para la detección de fotones, con una velocidad de escaneo de 100 Hz y con un tamaño de píxel de 17 nm. La acumulación de cuatro líneas se utilizó para la adquisición de TauSTED.
- Coloque una gota de 20 μL de medio de montaje (Tabla de materiales) en un portaobjetos de microscopio. Tome un cubreobjetos del paso 4.2.2 y coloque la muestra suavemente sobre el portaobjetos orientado hacia el medio de montaje (figura 2B).
Resultados
Para obtener imágenes del SC dentro del tejido de la línea germinal de C. elegans por SMLM, hemos empleado 3D-SMLM ratiométrico de 2 colores para localizar HTP-3, un componente de los ejes cromosómicos, y el extremo C del filamento transversal SYP-5 marcado endógenamente con una etiqueta de hemaglutinina (HA). La localización de ambas proteínas dentro del SC de C. elegans fue previamente determinada por otros estudios16,30.
Para minimizar la dispersión de la luz y las aberraciones ópticas inherentes a las muestras biológicas gruesas, obtuvimos imágenes de la sección z más inferior de los núcleos meióticos que contienen los SC (Figura 3, líneas amarillas). Para cada imagen adquirida, la posición de la etapa piezoeléctrica del plano de imagen se marcó en relación con la posición de la etapa piezoeléctrica cuando el objetivo se centró en el cubreobjetos. Esto permitió el cálculo de la distancia piezoeléctrica desde el cubreobjetos. Las muestras montadas con éxito se fijan de manera estable cerca del cubreobjetos y conservan la forma de la gónada (es decir, el tejido no se aplasta entre los dos cubreobjetos durante el paso posterior a la fijación). La calidad del montaje de la muestra puede evaluarse fácilmente bajo un microscopio estereoscópico, ya que las gónadas bien unidas no muestran ningún movimiento en la solución (paso 4.2.6). Sin embargo, debido a la estocasticidad del proceso de montaje, el tejido de la gónada no necesariamente se colocará completamente plano sobre el cubreobjetos. Por lo tanto, el plano inferior de los núcleos que contienen SC se puede encontrar a distancias variables en relación con el cubreobjetos dentro de la misma gónada.
Para ilustrar cómo cambia la resolución dependiendo de la unión del tejido al cubreobjetos, adquirimos imágenes a diferentes distancias piezoeléctricas al cubreobjetos. Para evaluar la calidad de una imagen individual, se calcularon las curvas de correlación de anillo de Fourier (FRC)50,51 y la resolución se determinó utilizando el complemento FRCResolution dentro del software SMAP48. En la Figura 4 se muestran dos núcleos representativos extraídos de dos imágenes 3D-SMLM separadas tomadas a diferentes distancias del cubreobjetos. En los SC situados cerca del cubreobjetos, los ejes cromosómicos y el extremo C de SYP-5::HA están bien resueltos en las tres dimensiones (Figura 4A, 0,8 μm del cubreobjetos). Para resolver dos estructuras separadas por una distancia dada, la resolución FRC alcanzada generalmente debe ser menor que la mitad de esta distancia en la resolución axial.
Para separar lateralmente las mismas estructuras, es necesario alcanzar valores de resolución FRC aún más pequeños. De hecho, en muestras que se encuentran muy cerca del cubreobjetos, la resolución FRC es de 38 nm para el canal AlexaFluor 647 y 34 nm para el canal CF680, y por lo tanto muy por debajo de la distancia esperada de 84 nm entre los C-termini de SYP-516. Por lo tanto, esta resolución resuelve fácilmente la organización del SC no sólo en vistas frontales sino también laterales (Figura 4B i,ii). Por el contrario, la resolución se deteriora en los SC ubicados a una distancia de 5 μm del cubreobjetos debido a la dispersión de la luz y las aberraciones esféricas (Figura 4B). Las resoluciones FRC a esta distancia caen a 47 nm (AlexaFluor 647) y 41 nm (CF680), que no pueden resolver completamente los extremos C de SYP-5. Dado que las aberraciones ópticas perjudican la resolución lateral más severamente que la resolución axial, las bandas HTP-3 y SYP-5 ya no se resuelven claramente en la sección transversal de la vista lateral en muestras ubicadas a una distancia de 5 μm del cubreobjetos (Figura 4B ii). La comparación de la resolución FRC de las imágenes adquiridas a diferentes distancias piezoeléctricas del cubreobjetos reveló que el tejido fotografiado no debería estar a más de 2 μm del cubreobjetos (Figura 5). Este resultado resalta la importancia de la ejecución correcta de la etapa posterior a la fijación, durante la cual el tejido debe ser reticulado con éxito al recubrimiento de poli-L-lisina del cubreobjetos.
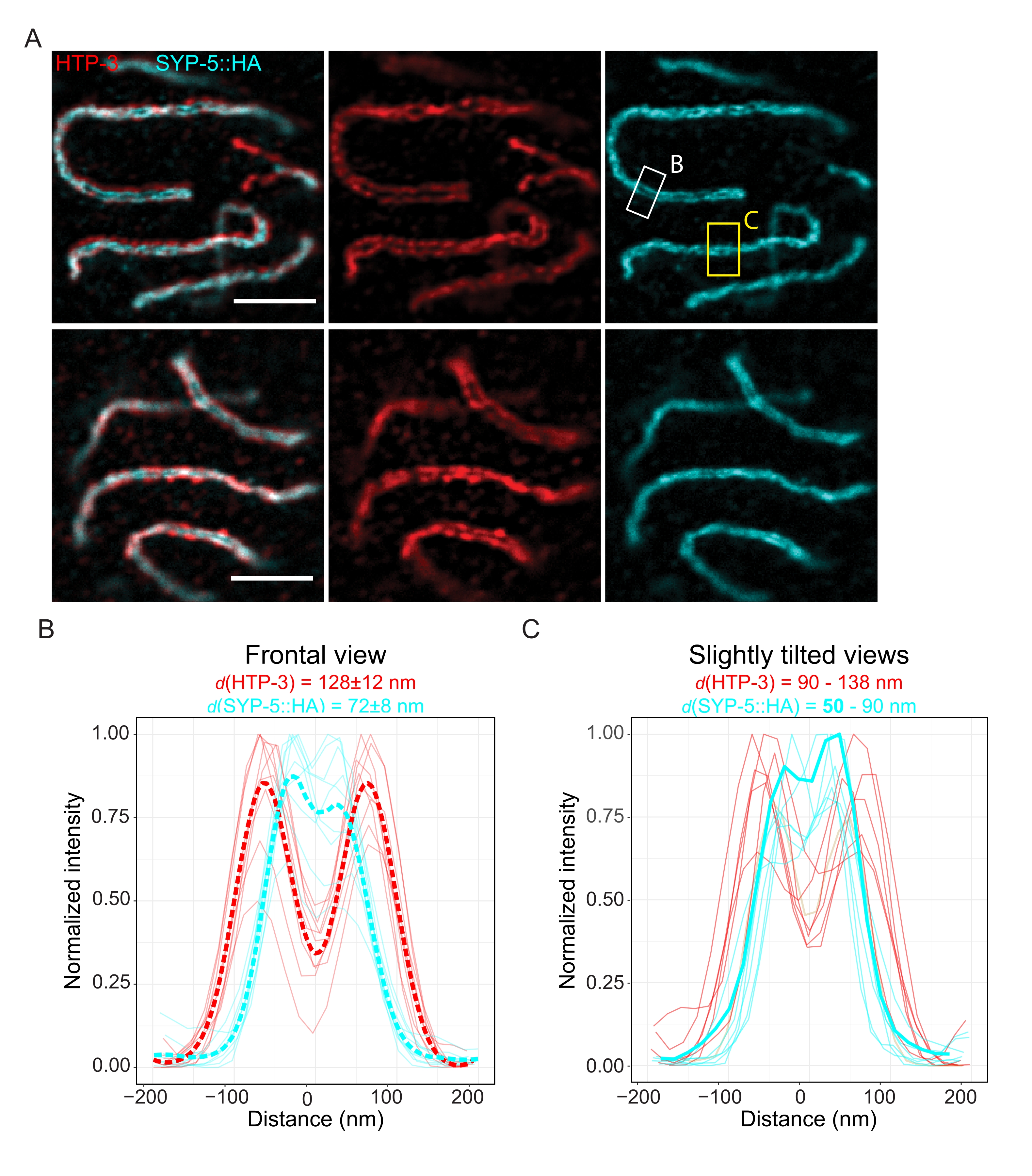
Para demostrar la resolución alcanzable con otra técnica de súper resolución, también tomamos imágenes de SC en tejido germinal intacto fijo con microscopía TauSTED. La Figura 6A muestra imágenes TauSTED con la resolución más alta y más baja alcanzada dentro de este estudio según lo estimado a partir de perfiles de línea del SC en vista frontal (Figura 6B). En ambos núcleos, podríamos resolver las dos bandas de localización de HTP-3 en los ejes cromosómicos y los C-terminales de SYP-5 en la región central, demostrando que la resolución alcanzable en TauSTED utilizando este protocolo optimizado es inferior a 84 nm. En condiciones óptimas (Figura 6A, arriba), pudimos resolver los extremos C en vistas ligeramente inclinadas del SC que estaban separadas por solo 50 nm (Figura 6A, rectángulo amarillo y 6C).

Figura 1: Esquema de la organización del complejo sinaptonémico en Caenorhabditis elegans. La caricatura muestra una estructura simplificada del SC en C. elegans uniendo dos cromosomas homólogos (gris). La estructura se muestra en vistas frontales, laterales y transversales. Los ejes cromosómicos se muestran como barras rojas, mientras que los filamentos transversales se muestran en cian. Las proteínas de filamentos transversales (SYP-1, 5, 6 en C. elegans) están orientadas de manera cabeza a cabeza (gráficos de bola cian) en la región central para salvar la distancia entre los dos ejes. Se indican las distancias esperadas entre los ejes y los extremos C de los filamentos transversales. Abreviatura: SC = complejo sinaptonémico. Haga clic aquí para ver una versión más grande de esta figura.
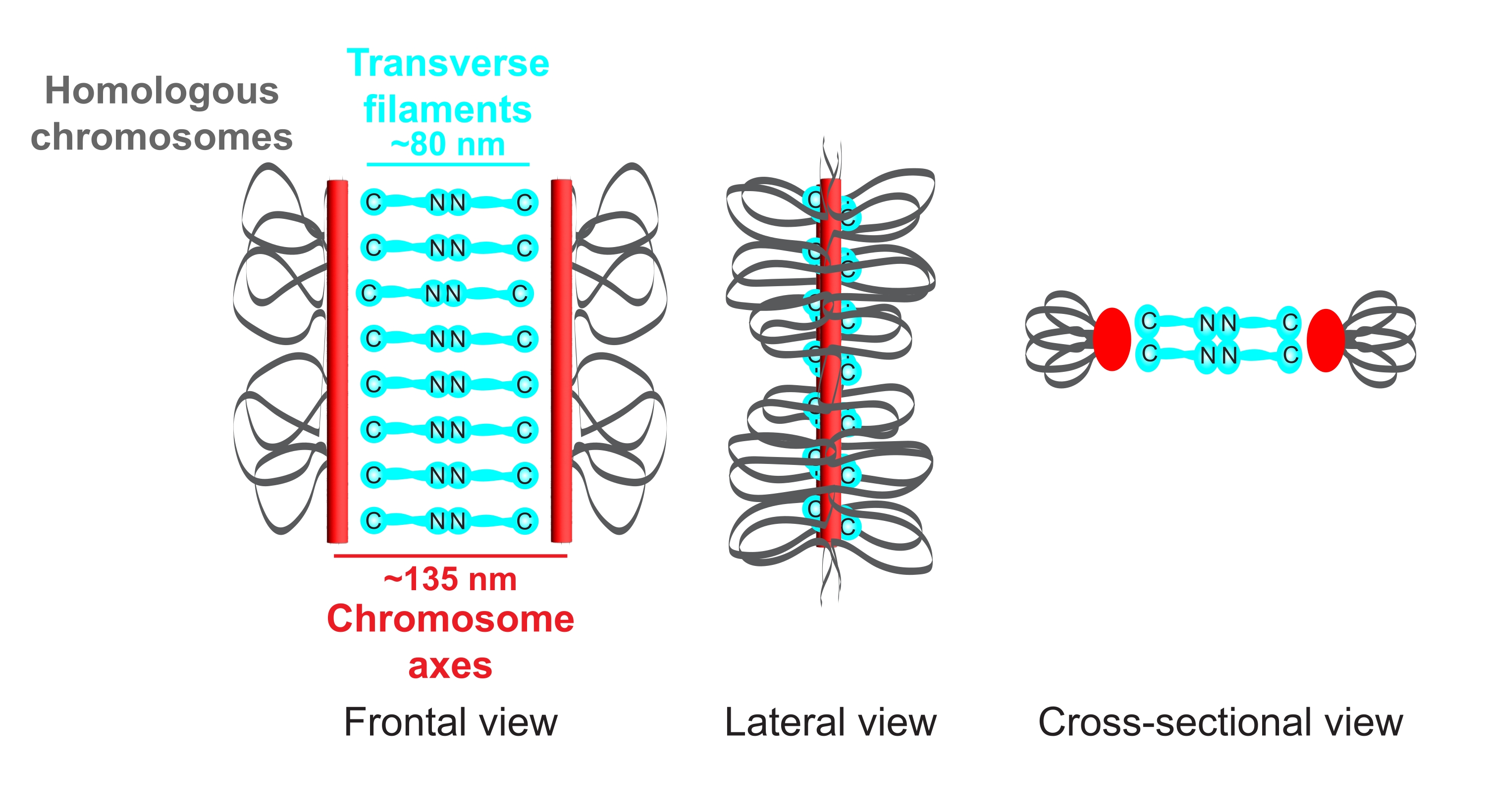{kind=link}

Figura 2: Ilustración de la preparación de la muestra utilizada en el estudio. (A) Los adultos jóvenes de C. elegans se diseccionan en la cabeza o la cola (líneas verdes y discontinuas) y se procesan como se describe en el protocolo. (B) Los pasos individuales del método se indican con gráficos que están conectados con flechas grises. Abreviaturas: STED = agotamiento de emisiones estimuladas; SMLM = microscopía de localización de molécula única; PBS = solución salina tamponada con fosfato. Haga clic aquí para ver una versión más grande de esta figura.
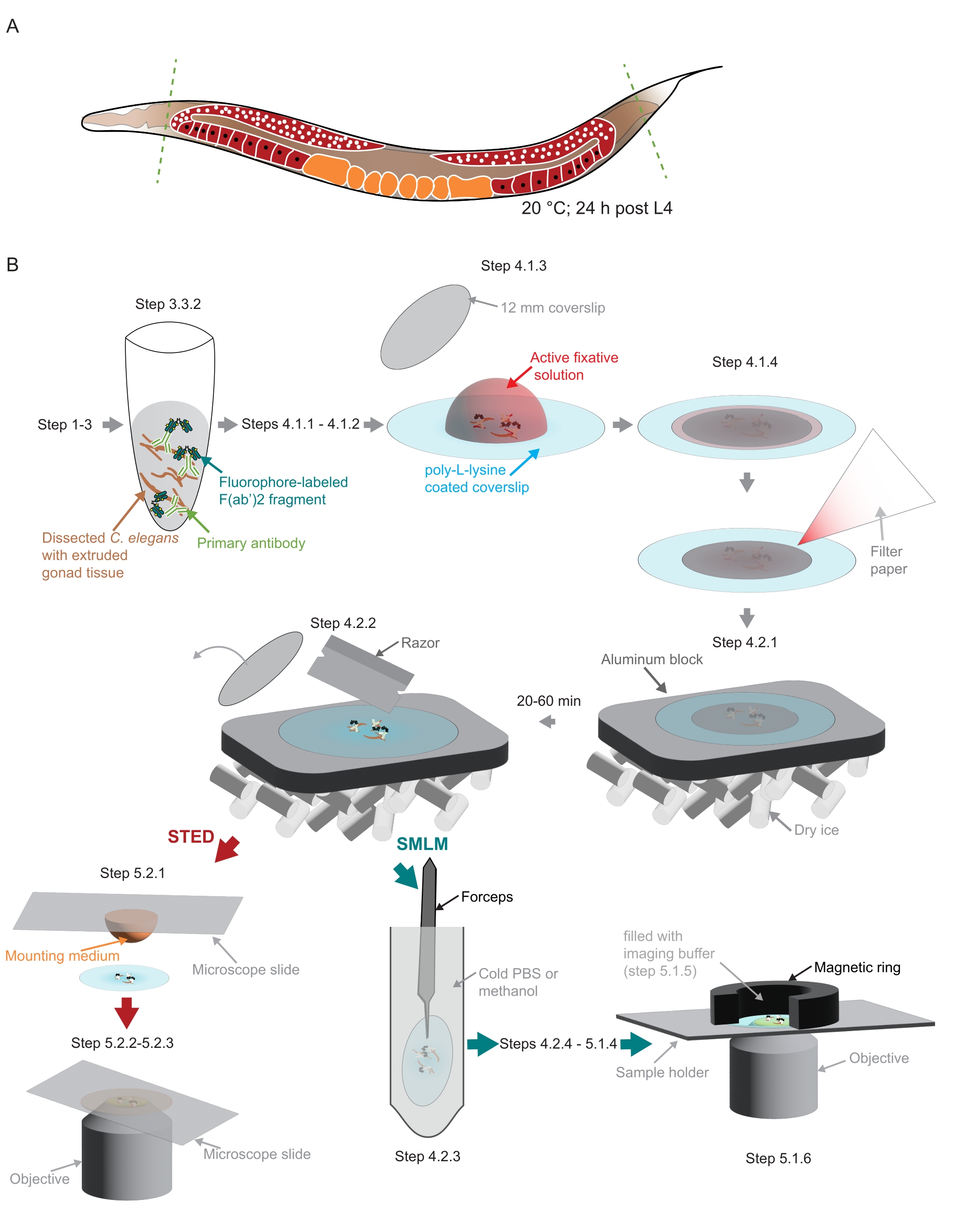{kind=link}

Figura 3: Ubicación de la sección de tejido que se puede observar mediante microscopía de localización de molécula única. MIP de una imagen confocal de disco giratorio de una gónada completa de C. elegans. El tejido se tiñó para HTP-3 y el extremo C de SYP-5 (SYP-5::HA), y la señal combinada se muestra en gris. Las imágenes confocales individuales se unieron utilizando el plugin Grid/Collection stitching Fiji52 para crear una imagen de toda la gónada. El recuadro muestra una vista xy del plano z inferior que contiene los SC. La localización de este plano se muestra en vistas ortogonales de la sección de tejido indicada por un rectángulo en la imagen MIP de la gónada (líneas amarillas). Barras de escala = 10 μm. Abreviaturas: MIP = proyección de intensidad máxima; SCs = complejos sinaptonemales. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}

Figura 4: Microscopía de localización de molécula única de HTP-3 y los C-terminales del SYP-5. (A,B) Izquierda: Imágenes SMLM que muestran núcleos de paquiteno teñidos para HTP-3 (rojo) y el extremo C de SYP-5 (SYP-5::HA, cian) (barra de escala = 1 μm). Centro: Imágenes ampliadas de regiones de interés que se indican en A y B con las correspondientes vistas transversales que se muestran debajo de cada imagen (i, ii; barra de escala = 100 nm). Los tramos del SC dentro de las imágenes ampliadas se giran para orientar los ejes cromosómicos paralelos al eje y. Derecha: Representación gráfica de la localización de las proteínas de interés dentro del SC retratando la orientación del SC en las regiones ampliadas que se muestran en el centro de la figura. Abreviaturas: SMLM = microscopía de localización de molécula única; SC = complejo sinaptonémico. Los datos brutos para reconstruir imágenes SMLM están disponibles a través de la base de datos BioStudies60 (ID de acceso: S-BIAD504). Haga clic aquí para ver una versión más grande de esta figura.
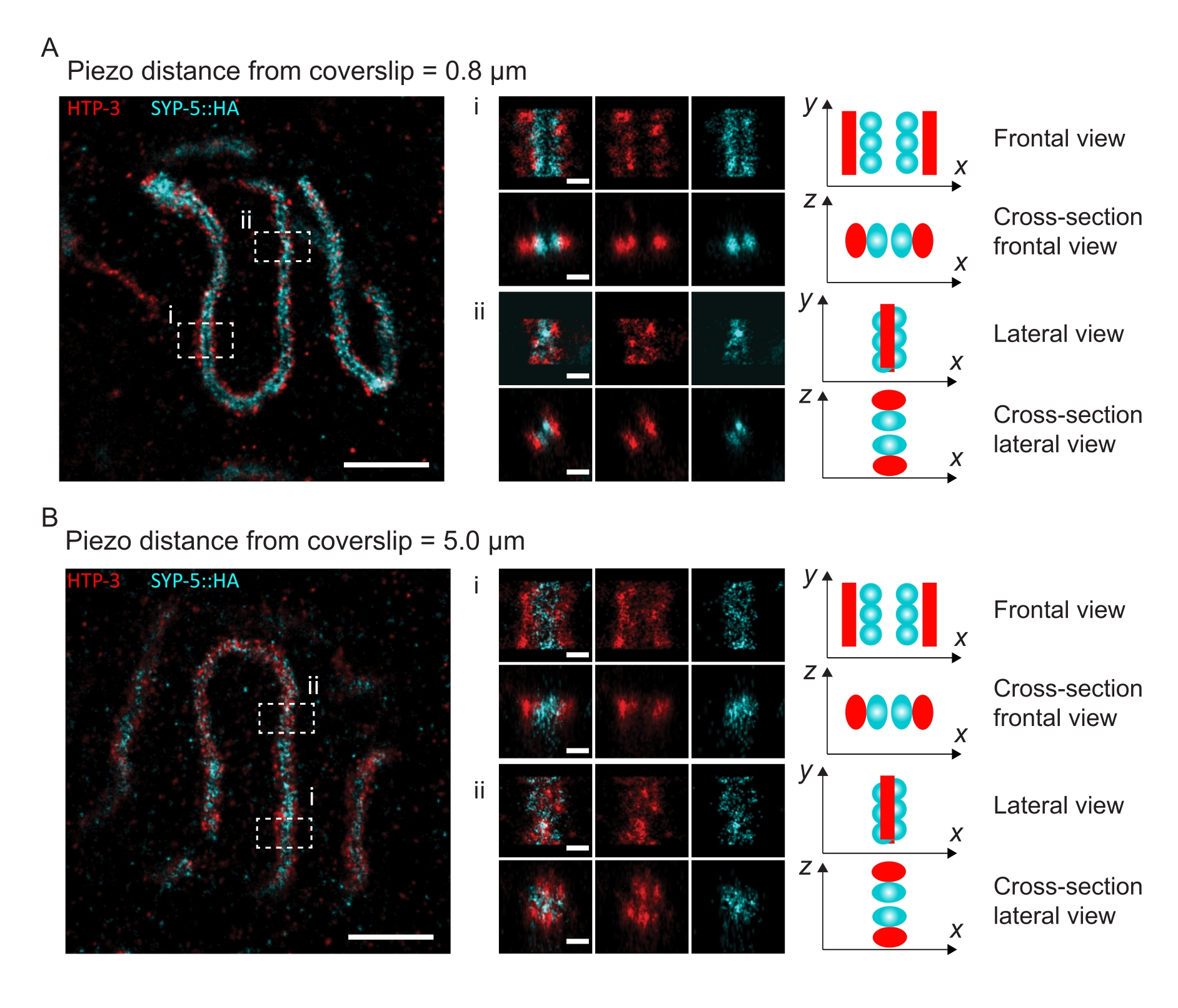{kind=link}

Figura 5: La resolución de correlación de anillo de Fourier de las imágenes de microscopía de localización de molécula única depende de la distancia del plano z fotografiado desde el plano del cubreobjetos. Las líneas de color muestran curvas FRC de imágenes adquiridas a diferentes distancias (como se muestra en la barra de color) desde el cubreobjetos. El umbral de 1/7 utilizado para determinar la resolución FRC se indica mediante una línea horizontal negra. Los recuadros muestran la dependencia de la resolución FRC en la distancia piezoeléctrica desde el cubreobjetos. El trazado se realizó mediante un script R escrito a medida (versión 4.1.2, Archivo Suplementario 1) en el que las curvas originales se suavizaron con funciones del paquete "ggplot2". Abreviaturas: FRC = correlación de anillo de Fourier; SMLM = microscopía de localización de molécula única; SC = complejo sinaptonémico. Los datos para las curvas FRC y los datos SMLM están disponibles a través de la base de datos BioStudies60 (ID de acceso: S-BIAD504). Haga clic aquí para ver una versión más grande de esta figura.
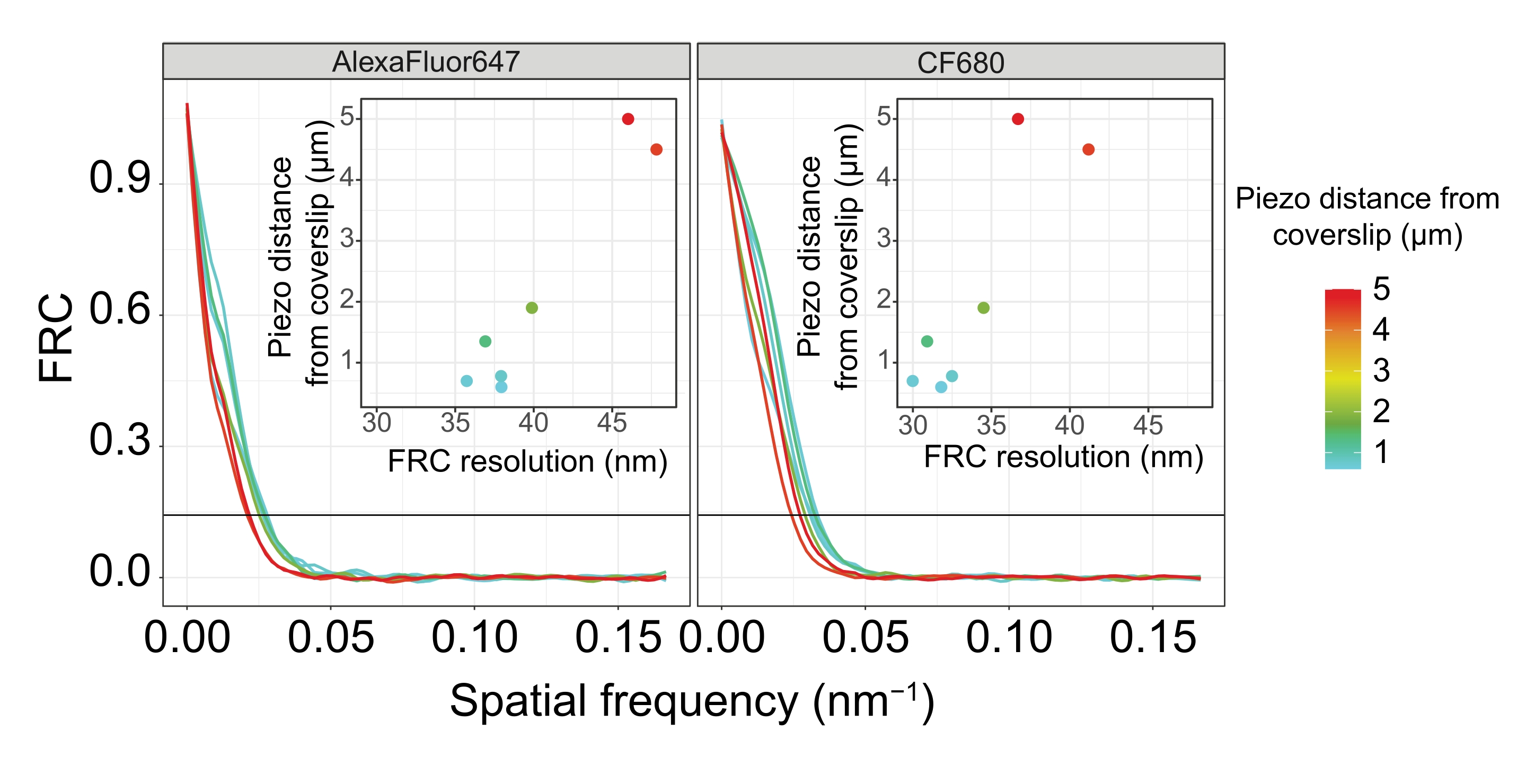{kind=link}

Figura 6: La microscopía de agotamiento de emisión estimulada mejorada por información basada en la vida útil de fluorescencia (TauSTED) resuelve dos bandas de localización tanto para HTP-3 como para el extremo C de SYP-5. (A) Dos imágenes representativas de TauSTED muestran núcleos de paquiteno teñidos para HTP-3 (rojo) y el extremo C de SYP-5 (SYP-5::HA, cian) con definición estructural más alta (arriba) e inferior (abajo) (barra de escala = 1 μm). Los rectángulos marcan regiones con los extremos C resueltos de SYP-5 en frontal (blanco) y una vista ligeramente inclinada (amarillo) del SC. (B,C) Distribución de la señal HTP-3 (rojo) y C-terminal de SYP-5 (cian) resuelta por TauSTED. Los perfiles de línea de las regiones de interés que contienen el SC en vistas frontales (B) o ligeramente inclinadas (C) se muestran como líneas completas con intensidad normalizada al valor máximo. Los perfiles de línea se generaron utilizando Fiji ImageJ. Las líneas discontinuas en B muestran los datos promediados para cada proteína. La línea cian gruesa en C corresponde al perfil de línea con la distancia resuelta más corta entre los C-terminales de SYP-5. Para determinar las distancias entre los anticuerpos dirigidos a proteínas específicas, los perfiles de línea (n = 9 (B), n = 7 (C)) se ajustaron con gaussianos dobles utilizando un script R escrito a medida (versión 4.1.2, Archivo Suplementario 1). La distancia media ± la desviación estándar (B) y el rango con el valor mínimo resaltado en negrita (C) se indican en la parte superior de cada gráfico, respectivamente. Abreviaturas: STED = microscopía de agotamiento por emisión estimulada; SC = complejo sinaptonémico. Las imágenes mostradas y los puntos de datos de los perfiles de líneas trazadas están disponibles a través de la base de datos BioStudies60 (ID de acceso: S-BIAD504). Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}
Tabla 1: Composición de tampones y soluciones utilizadas en este protocolo. Haga clic aquí para descargar esta tabla.
Video complementario 1: Adquisición de microscopía de localización de molécula única. Video que muestra fluoróforos parpadeando a una velocidad adecuada (se muestran 50 cuadros, barra de escala = 5 μm, 20 ms / cuadro). Haga clic aquí para descargar este video.
Archivo complementario 1: Script de análisis de datos. Haga clic aquí para descargar este archivo.
Discusión
La organización en forma de escalera del SC, que es esencial para la correcta recombinación y segregación de cromosomas homólogos, se observó por primera vez hace casi 70 años en microscopía electrónica 3,4. Si bien la organización general del SC se resuelve fácilmente en microscopía electrónica, la localización de componentes individuales dentro de este complejo requiere un enfoque más específico. Con su ancho de sólo ~100 nm, la subestructura del SC no puede ser resuelta por microscopía de fluorescencia convencional. Sin embargo, la microscopía de súper resolución se ha convertido en un importante impulsor de nuevos descubrimientos sobre la estructura y función del complejo sinaptonémico 16,19,24,25,26,27,28,29,30. Para facilitar esta investigación, hemos demostrado un procedimiento de montaje que permite estudiar la arquitectura del SC dentro del tejido de la gónada de C. elegans con microscopía SMLM y STED.
Un paso crítico para optimizar la resolución en las imágenes SMLM es la reticulación directa del tejido de la línea germinal a un cubrecubierto recubierto de poli-L-lisina (paso 4). La unión covalente del tejido al cubreobjetos es esencial para reducir los movimientos dentro de la muestra que darían lugar a grandes derivas y harían imposible la obtención de imágenes durante largos períodos de tiempo para SMLM. Además, incluso una unión subóptima que deja los núcleos que contienen SC a una distancia del cubreobjetos conduce a una caída significativa en la resolución alcanzable como resultado de aberraciones esféricas (Figura 4). Alternativamente a la unión covalente utilizada aquí, el tejido de la línea germinal teñido también se puede inmovilizar entre dos cubreobjetos sellados en una pequeña gota de tampón de imagen19,30. Sin embargo, este método de inmovilización reduce severamente el volumen de tampón de imagen en la muestra de 1 ml utilizado en el protocolo optimizado aquí a solo unos pocos μL, lo que resultará en una acidificación del tampón de imágenes y reducirá severamente el tiempo durante el cual se puede obtener una imagen de la muestra 38,53,54.
Los largos tiempos de adquisición para la microscopía SMLM y STED limitan el uso de estos métodos a la obtención de imágenes de muestras fijadas químicamente. Aquí, la fijación de paraformaldehído asegura que la estructura del SC se conserve durante la preparación de la muestra y la obtención de imágenes. Sin embargo, a pesar de las precauciones tomadas aquí para obtener imágenes del SC dentro del tejido intacto, la estructura resultante del SC después de la fijación no es necesariamente idéntica a la estructura en su estado nativo dentro de un organismo vivo. Además, dado que una sola imagen del SC fijo representa una única "instantánea" de la estructura biológica, este enfoque permanece ciego a la dinámica de la estructura nativa in vivo.
Sin embargo, la información sobre la dinámica y la variabilidad de las estructuras macromoleculares también se puede obtener adquiriendo no solo una sino muchas "instantáneas". Si bien este enfoque puede resolver cambios en la estructura del SC durante el paquiteno19, hay varios factores que limitan el número de imágenes que se pueden adquirir de una sola muestra preparada utilizando este protocolo. En primer lugar, las altas potencias láser utilizadas durante la adquisición de imágenes conducen a un blanqueamiento permanente de los fluoróforos e impiden la obtención de imágenes de regiones adyacentes de interés o múltiples planos z, lo que reduce significativamente el número de imágenes que se pueden adquirir de una sola muestra. En segundo lugar, la densidad de muestra/tejido en el cubreobjetos preparado por este método es baja, lo que limita significativamente el número de imágenes que se pueden adquirir de un solo cubreobjetos. La baja densidad de la muestra también prohíbe el uso de tuberías automatizadas de adquisición de imágenes que ayudaron a arrojar luz sobre otras cuestiones biológicas 34,55,56,57,58,59. Sin embargo, la densidad de la muestra puede ser aumentada ligeramente por un usuario experimentado.
El protocolo presentado aquí está optimizado para obtener una alta densidad de etiquetado que es necesaria para lograr una resolución óptima en SMLM35. Mientras que los protocolos anteriores unen covalentemente el tejido al cubreobjetos antes de la inmunotinción16, este nuevo protocolo vincula el tejido al cubreobjetos solo después de que las muestras se tiñeron en solución. Esta modificación permite que los anticuerpos utilizados para el inmunomarcaje accedan libremente al tejido desde todos los lados, mientras que la unión covalente del tejido al cubreobjetos puede restringir que los anticuerpos lleguen a los núcleos más cercanos al cubreobjetos, reduciendo así el grado de marcado. En conjunto, las modificaciones descritas aquí mejoran la resolución de 40-50 nm (resolución FRC)16 a 30-40 nm (este protocolo).
Es importante destacar que, si bien una alta densidad de etiquetado y una alta concentración de anticuerpos son esenciales para SMLM, encontramos que se obtienen mejores imágenes de microscopía STED utilizando concentraciones de anticuerpos más bajas (paso 3). Con una resolución de decenas de nanómetros, el tamaño de las moléculas utilizadas para etiquetar la proteína de interés se vuelve cada vez más importante. Por lo tanto, empleamos fragmentos F(ab')2 que son la mitad del tamaño de los anticuerpos de longitud completa. La mejora en el contraste local debido a una fuente de señal más pequeña, y por lo tanto la resolución obtenida por esta modificación en comparación con el uso de anticuerpos secundarios de longitud completa, permitió la resolución de los dos C-terminales de SYP-5 dentro de la región central por TauSTED, que no se resuelven mediante ETS convencionales utilizando anticuerpos de longitud completa (16 y datos no mostrados). Anticipamos que este protocolo optimizado para obtener imágenes de SC en líneas germinales intactas de C. eleganfacilitará la investigación de la relación estructura-función del SC durante la meiosis.
Divulgaciones
Los autores declaran no tener conflictos de intereses.
Agradecimientos
Nos gustaría agradecer a Jonas Ries y al laboratorio de Ries por compartir búferes de imágenes para imágenes SMLM. También agradecemos a Yumi Kim por la cepa C. elegans utilizada en este protocolo y a Abby F. Dernburg por el anticuerpo pollo-anti-HTP-3. Agradecemos a Marko Lampe y Stefan Terjung del Centro de Microscopía Óptica Avanzada del EMBL Heidelberg por su apoyo en el uso del microscopio confocal Olympus iXplore SPIN SR. Este trabajo fue apoyado por el Laboratorio Europeo de Biología Molecular y la Deutsche Forschungsgemeinschaft (DFG, Fundación Alemana de Investigación - 452616889, SK). Reconocemos el acceso y los servicios proporcionados por el Centro de Imágenes del Laboratorio Europeo de Biología Molecular (EMBL IC), generosamente apoyado por la Fundación Boehringer Ingelheim.
Materiales
| Name | Company | Catalog Number | Comments |
| 100x/1.5 oil objective | Olympus | UPLAPO100XOHR | UPLAPO100XOHR |
| 2-mercaptoethylamine (MEA) | Sigma-Aldrich | 30070-10G | Dissolved in MilliQ water to 5 M solution, pH 8.7 adjusted with HCl. Aliquoted to a single-use volume, frozen, and kept at -80 °C. |
| Additional 640 nm booster laser | Toptica | IBEAM-SMART-640-S-HP | |
| AlexaFluor 594, NHS ester | ThermoFischer Scientific | A37572 | Dissolved in DMSO to 1 mM solution, aliquoted to single use volume, frozen and kept at -80 °C |
| AlexaFluor 647, NHS ester | ThermoFischer Scientific | A37573 | Dissolved in DMSO to 1 mM solution, aliquoted to single use volume, frozen and kept at -80 °C |
| anti-HA | Thermo Fisher Scientific | 2-2.2.14 | Mouse monoclonal, 1:250 (SMLM), 1:1,000 (STED Microscopy) |
| anti-HTP-3 | a gift from Abby F. Dernburg | MacQueen et al., 2005 | Chicken polyclonal, 1:250 (SMLM), 1:1,000 (STED Microscopy) |
| Caenorhabditis elegans strain YKM349 | a gift from Yumi Kim | Hurlock et al., 2020 | syp-5(kim9[syp-5::HA]) I; meIs8[pie-1p::GFP::cosa-1, unc-119(+)] II |
| CF6680, NHS ester | Biotium | 92139 | Dissolved in DMSO to 1mM solution, aliquoted to single use volume, frozen and kept at -80 °C |
| Circular cover glass 12 mm No. 1 | Menzel-Gläser; VWR | 631-0713 | |
| Circular cover glass 24 mm No. 1.5 | Carl Roth | PK26.1 | |
| Cylindrical lenses | Thorlabs | LJ1516RM-A, LK1002RM-A | |
| Egg Buffer (10x) | Edgar 1995 | 250 mM HEPES, 1.18 M NaCl, 480 mM KCl, 20 mM EDTA, 5 mM EGTA, pH 7.4 | |
| Ethanol (absolute for analysis) | Merck | 64-17-5 | |
| F(ab’)2 fragment anti-chicken IgY | Jackson Immunoresearch | AB_2340347 | Donkey polyclonal, 1:100 (SMLM), 1:1,000 (STED Microscopy) |
| F(ab’)2 fragment anti-mouse IgG | Jackson Immunoresearch | AB_2340761 | Donkey polyclonal, 1:100 (SMLM), 1:1,000 (STED Microscopy) |
| Fisherbrand Microscope slides T/F Ground 0.8-1.0 mm thick | Fisher scientific | 7107 | |
| Gauge Worm Pick 30 diameter 0.254 mm - Iridium 10% | Kisker | 789265 | |
| Glucose oxidase/Catalase enzyme mix (GlOX/Cat ) | a gift from Jonas Ries | Hoess, Mund, Reitberger, & Ries, 2018 | 20x, 1916 U/mL glucose oxidase (Sigma G7141), 42350 U/mL catalase (Sigma C3155), 50 mM Tris-HCl pH 8.0, 51% glycerol, MilliQ water. Stored at -20 °C. |
| Imaging buffer base | a gift from Jonas Ries | Hoess, Mund, Reitberger, & Ries, 2018 | 50 mM Tris-HCl, pH 8.0, 10 mM NaCl, 10% D-Glucose. Aliquoted to a single-use volume (950 μL), frozen, and kept at -80 °C. |
| Invitrogen ProLong Glass Antifade Mountant | ThermoFischer Scientific | P36982 | |
| Leica Stellaris 8 STED FALCON | Leica | N/A | The microscope is equiped with the latest generation white light laser, a 775nm pulsed STED laser, the FALCON Fluorescence Lifetime IMaging module, HC PL APO CS2 100x/1.40 oil objective, and Leica HyD X detector. The system is capable of FLIM module enhanced Tau-STED which measures the specific fluorescence lifetime of a dye and is therefore capable of removing background signal based on differences in fluorescence lifetimes of the dyes, and dye conditions in the sample. Additionally, the resolution is increased by accounting for the variation of fluorescence lifetimes in different areas of the depletion donut. |
| Longwave channel emission filter | AHF Analysentechnik | F47-702 | 700/100 nm bandpass |
| Methanol (absolute for analysis) | Merck | 67-56-1 | |
| NaHCO3 | Sigma-Aldrich/Merck | S5761-500G | 100 mM NaHCO3, pH 8.3 |
| Near-infrared fiber-coupled laser | Toptica | IBEAM-SMART-PT-CD | Custom Design, 808 nm - 75mW |
| Objective lens piezo mount (PIFOC ) | Physik Instrumente | P-726.1.CD | 100 µm travel range |
| Orca Fusion BT sCMOS camera | Hamamatsu | C15440-20UP | |
| PCR tubes | Greiner Bio-One | 673283 | 0.2 mL |
| Phosphate Saline Buffer (PBS 10x) | N/A | 137 mM NaCl, 2.7 mM KCl, 10 mM Na2HPO4, 1.8 mM KH2PO4, pH 7.4 | |
| Pierce 16% formaldehyde (w/v), methanol free | ThermoFischer Scientific | 28906 | 16% formaldehyde is transferred from the original glass ampule into 1.5 mL tube and kept at room temperature. |
| PlasmaPrep2 plasma cleaner | GaLa Instrumente GmbH | N/A | |
| Poly-L-lysine hydrobromide | Sigma-Aldrich/Merck | P2636-25MG | 0.1% w/v solution was prepared in Milli-Q water, and stored in aliquots at -20 °C. |
| Primary dichroic (illumination reflecting) | AHF Analysentechnik | F73-866S | quad bandpass @ 405, 488, 561, 640 |
| Quadnotch filter | AHF Analysentechnik | F40-072 | 405/488/561/640 nm |
| Quadrant photodiode (QPD) | Laser Components | SD197-23-21-041, LC301DQD-PV | |
| Razor blades | Apollo Herkenrath Solinger | N/A | |
| Refractive beam shaper | AdlOptica | PiShaper 6_6_VIS | |
| Roche Blocking Reagent | Roche | 11096176001 | 10x solution was prepared according to recommendation. Frozen aliquots were stored at -20 °C. |
| Scalpel blade (Feather brand #11, No. 3) | Heinz Herenz Medizinalbedarf GmbH | 1110911 | |
| Scalpel removal box | Fisher scientific | 10002-50 | |
| Secondary dichroic (emission reflecting) | AHF Analysentechnik | F38-785S | 750 nm longpass |
| Shortpass filter | Semrock | BSP01-785R-25 | 750 nm |
| Shortwave channel emission filter | AHF Analysentechnik | F37-677 | 676/37 nm bandpass |
| Single molecule localization microscope | EMBL Imaging Centre | Diekmann et al., 2020 with modifications | The microscope provides widefield epi-illumination via a single-mode fiber-coupled laser engine, additional booster laser, and refractive beam shaper to provide a uniform illumination field (Stehr et al, 2019). Widefield images are captured on a sCMOS camera and appropriate relay optics for a system magnification of 61x and a pixel size of 106 nm. For ratiometric imaging of spectrally overlapping far-red dyes, an image splitter produces two spectrally distinct images on the camera (splitting dichroic: 665 nm long pass, shortwave channel emission filter: 676/37 nm bandpass, longwave emission filter: 700/100 nm bandpass. An additional 405/488/561/640 nm quadnotch filter and 750 nm shortpass filter are common to the two paths and provide additional laser blocking). A compound cylindrical lens provides the astigmatism required for 3D imaging. To maintain a fixed focus across acquisitions exceeding 2 hours in time (comprising 200 000 - 250 000 images), focus locking is achieved by total internal reflection of a near-infrared fiber-coupled laser from the coverslip and subsequent height sensitive detection on a quadrant photodiode (QPD). The QPD signal provided closed-loop control of the objective lens piezo mount. For access to this microscope, refer to https://www.embl.org/about/info/imaging-centre or contact ic-contact@embl.de |
| Single-mode fiber-coupled multi-laser engine | Toptica | iCHROME MLE-LFA-HP | Provides widefield epi-illumination of 100 mW at 405, 488, 561, 640 nm |
| Splitting dichroic | AHF Analysentechnik | F48-665SG | 665 nm long pass |
| Square cover glass 22 x 22 mm No.1 | Menzel-Gläser; VWR | 630-2882 | |
| STAR 635P, NHS ester | Abberior | ST635P-0002-1MG | Dissolved in DMSO to 1 mM solution, aliquoted to single use volume, frozen and kept at -80 °C |
| Stereo microscope Stemi 305 Stand K LAB | Zeiss | N/A | |
| Tetramisole hydrochloride | Sigma-Aldrich/Merck | T1512-2G | 1% (w/v) solution was prepared in Milli-Q water. Frozen aliquots were stored at -20 °C. Thawed aliquot was kept at 4 °C and used for several months. |
| TetraSpeck Microspheres | ThermoFischer Scientific | T7279 | 0.1 µm, fluorescent blue/green/orange/dark red |
| Tris Saline Buffer (TBS 10x) | N/A | 200 mM Tris-HCl, 1.5 M NaCl, pH 7.5 | |
| TWEEN 20 | Sigma-Aldrich/Merck | P9416-50ML | Kept at room temperature in original packaging. |
| WormStuff worm pick | Kisker | 789277 | |
| XY microscope stage | Smaract | N/A | Custom Design |
| Zeba Micro Spin Desalting Column | ThermoFischer Scientific | 89877 | 7K MWCO, 75 µL |
Referencias
- Zickler, D., Kleckner, N. Meiotic chromosomes: integrating structure and function. Annual Review of Genetics. 33, 603 (1999).
- Ur, S. N., Corbett, K. D. Architecture and dynamics of meiotic chromosomes. Annual Review of Genetics. 55, 497-526 (2021).
- Fawcett, D. W. The fine structure ot chromosomes in the meiotic prophase of vertebrate spermatocytes. The Journal of Biophysical and Biochemical Cytology. 2 (4), 403-406 (1956).
- Moses, M. J. Chromosomal structures in crayfish spermatocytes. The Journal of Biophysical and Biochemical Cytology. 2 (2), 215-218 (1956).
- Hillers, K. J., Jantsch, V., Martinez-Perez, E., Yanowitz, J. L. Meiosis. WormBook. , 433-434 (2017).
- Pasierbek, P., et al. A Caenorhabditis elegans cohesion protein with functions in meiotic chromosome pairing and disjunction. Genes & Development. 15 (11), 1349-1360 (2001).
- Severson, A. F., Ling, L., Van Zuylen, V., Meyer, B. J. The axial element protein HTP-3 promotes cohesin loading and meiotic axis assembly in C. elegans to implement the meiotic program of chromosome segregation. Genes & Development. 23 (15), 1763-1778 (2009).
- Zetka, M. C., Kawasaki, I., Strome, S., Müller, F. Synapsis and chiasma formation in Caenorhabditis elegans require HIM-3, a meiotic chromosome core component that functions in chromosome segregation. Genes & Development. 13 (17), 2258-2270 (1999).
- Martinez-Perez, E. HTP-1-dependent constraints coordinate homolog pairing and synapsis and promote chiasma formation during C. elegans meiosis. Genes & Development. 19 (22), 2727-2743 (2005).
- Couteau, F., Zetka, M. HTP-1 coordinates synaptonemal complex assembly with homolog alignment during meiosis in C. elegans. Genes & Development. 19 (22), 2744-2756 (2005).
- Goodyer, W., et al. HTP-3 Links DSB Formation with Homolog Pairing and Crossing Over during C. elegans Meiosis. Developmental Cell. 14 (2), 263-274 (2008).
- Colaiácovo, M. P., et al. Synaptonemal complex assembly in C. elegans is dispensable for loading strand-exchange proteins but critical for proper completion of recombination. Developmental Cell. 5 (3), 463-474 (2003).
- MacQueen, A. J., Colaiácovo, M. P., McDonald, K., Villeneuve, A. M. Synapsis-dependent and -independent mechanisms stabilize homolog pairing during meiotic prophase in C. elegans. Genes & Development. 16 (18), 2428-2442 (2002).
- Smolikov, S., et al. Synapsis-defective mutants reveal a correlation between chromosome conformation and the mode of double-strand break repair during Caenorhabditis elegans meiosis. Genetics. 176 (4), 2027-2033 (2007).
- Smolikov, S., Schild-Prüfert, K., Colaiácovo, M. P. A yeast two-hybrid screen for SYP-3 interactors identifies SYP-4, a component required for synaptonemal complex assembly and chiasma formation in Caenorhabditis elegans meiosis. PLoS Genetics. 5 (10), 1000669 (2009).
- Hurlock, M. E., et al. Identification of novel synaptonemal complex components in C. Elegants. The Journal of Cell Biology. 219 (5), (2020).
- Zhang, Z., et al. Multivalent weak interactions between assembly units drive synaptonemal complex formation. The Journal of Cell Biology. 219 (5), (2020).
- Schild-Prüfert, K., et al. Organization of the synaptonemal complex during meiosis in Caenorhabditis elegans. Genetics. 189 (2), 411-421 (2011).
- Köhler, S., Wojcik, M., Xu, K., Dernburg, A. F. The interaction of crossover formation and the dynamic architecture of the synaptonemal complex during meiosis. bioRxiv. , (2020).
- Chen, F., Tillberg, P. W., Boyden, E. S. Expansion microscopy. Science. 347 (6621), 543-548 (2015).
- Klar, T. A., Jakobs, S., Dyba, M., Egner, A., Hell, S. W. Fluorescence microscopy with diffraction resolution barrier broken by stimulated emission. Proceedings of the National Academy of Sciences. 97 (15), 8206-8210 (2000).
- Rust, M. J., Bates, M., Zhuang, X. Sub-diffraction-limit imaging by stochastic optical reconstruction microscopy (STORM). Nature Methods. 3 (10), 793-796 (2006).
- Betzig, E., et al. Imaging intracellular fluorescent proteins at nanometer resolution. Science. 313 (5793), 1642-1645 (2006).
- Schücker, K., Holm, T., Franke, C., Sauer, M., Benavente, R. Elucidation of synaptonemal complex organization by super-resolution imaging with isotropic resolution. Proceedings of the National Academy of Sciences. 112 (7), 2029-2033 (2015).
- Cahoon, C. K., et al. Superresolution expansion microscopy reveals the three-dimensional organization of the Drosophila synaptonemal complex. Proceedings of the National Academy of Sciences. 114 (33), 6857-6866 (2017).
- Zwettler, F. U., et al. Tracking down the molecular architecture of the synaptonemal complex by expansion microscopy. Nature Communications. 11 (1), 1-11 (2020).
- Yoon, S., Choi, E. H., Kim, J. W., Kim, K. P. Structured illumination microscopy imaging reveals localization of replication protein A between chromosome lateral elements during mammalian meiosis. Experimental & Molecular Medicine. 50 (8), 1-12 (2018).
- Prakash, K., et al. Superresolution imaging reveals structurally distinct periodic patterns of chromatin along pachytene chromosomes. Proceedings of the National Academy of Sciences. 112 (47), 14635-14640 (2015).
- Xu, H., et al. Molecular organization of mammalian meiotic chromosome axis revealed by expansion STORM microscopy. Proceedings of the National Academy of Sciences. 116 (37), 18423-18428 (2019).
- Köhler, S., Wojcik, M., Xu, K., Dernburg, A. F. Superresolution microscopy reveals the three-dimensional organization of meiotic chromosome axes in intact Caenorhabditis elegans tissue. Proceedings of the National Academy of Sciences. 114 (24), 4734-4743 (2017).
- Hell, S. W. Far-field optical nanoscopy. Science. 316 (5828), 1153-1158 (2007).
- Hein, B., Willig, K. I., Hell, S. W. Stimulated emission depletion (STED) nanoscopy of a fluorescent protein-labeled organelle inside a living cell. Proceedings of the National Academy of Sciences. 105 (38), 14271-14276 (2008).
- Jahr, W., Velicky, P., Danzl, J. G. Strategies to maximize performance in STimulated Emission Depletion (STED) nanoscopy of biological specimens. Methods. 174, 27-41 (2019).
- Thevathasan, J. V., et al. Nuclear pores as versatile reference standards for quantitative superresolution microscopy. Nature Methods. 16 (10), 1045-1053 (2019).
- Xu, K., Shim, S. -. H., Zhuang, X. Super-resolution imaging through stochastic switching and localization of single molecules: an overview. Far-Field Optical Nanoscopy. , 27-64 (2013).
- Wang, Y., et al. Localization events-based sample drift correction for localization microscopy with redundant cross-correlation algorithm. Optics Express. 22 (13), 15982 (2014).
- Winterflood, C. M., Platonova, E., Albrecht, D., Ewers, H. Dual-color 3D superresolution microscopy by combined spectral-demixing and biplane imaging. Biophysical Journal. 109 (1), 3-6 (2015).
- Diekmann, R., et al. Optimizing imaging speed and excitation intensity for single molecule localization microscopy. Nature Methods. 17 (9), 909 (2020).
- Huang, B., Wang, W., Bates, M., Zhuang, X. Three-dimensional super-resolution imaging by stochastic optical reconstruction microscopy. Science. 319 (5864), 810-813 (2008).
- Phillips, C. M., McDonald, K. L., Dernburg, A. F. Cytological analysis of meiosis in Caenorhabditis elegans. Methods in Molecular Biology. 558, 171-195 (2009).
- Edgar, L. G. Blastomere culture and analysis. Methods in Cell Biology. 48, 303-321 (1995).
- MacQueen, A. J., et al. Chromosome sites play dual roles to establish homologous synapsis during meiosis in C. elegans. Cell. 123 (6), 1037-1050 (2005).
- Stehr, F., Stein, J., Schueder, F., Schwille, P., Jungmann, R. Flat-top TIRF illumination boosts DNA-PAINT imaging and quantification. Nature Communications. 10 (1), 1-8 (2019).
- Hoess, P., Mund, M., Reitberger, M., Ries, J. Dual-color and 3D super-resolution microscopy of multi-protein assemblies. Methods in Molecular Biology. 1764, 237-251 (2018).
- Edelstein, A., Amodaj, N., Hoover, K., Vale, R., Stuurman, N. Computer Control of microscopes using µManager. Current Protocols in Molecular Biology. 92 (1), 14-20 (2010).
- Edelstein, A. D., et al. Advanced methods of microscope control using µManager software. Journal of Biological Methods. 1 (2), 10 (2014).
- Deschamps, J., Ries, J. EMU: reconfigurable graphical user interfaces for Micro-Manager. BMC Bioinformatics. 21 (1), 1-13 (2020).
- Ries, J. SMAP: a modular super-resolution microscopy analysis platform for SMLM data. Nature Methods. 17 (9), 870-872 (2020).
- Li, Y., et al. Global fitting for high-accuracy multi-channel single-molecule localization. Nature Communications. 13 (1), 1-11 (2022).
- Nieuwenhuizen, R. P. J., et al. Measuring image resolution in optical nanoscopy. Nature Methods. 10 (6), 557-562 (2013).
- Banterle, N., Bui, K. H., Lemke, E. A., Beck, M. Fourier ring correlation as a resolution criterion for super-resolution microscopy. Journal of Structural Biology. 183 (3), 363-367 (2013).
- Preibisch, S., Saalfeld, S., Tomancak, P. Globally optimal stitching of tiled 3D microscopic image acquisitions. Bioinformatics. 25 (11), 1463-1465 (2009).
- Shi, X., Lim, J., Ha, T. Acidification of the oxygen scavenging system in single-molecule fluorescence studies: in situ sensing with a ratiometric dual-emission probe. Analytical Chemistry. 82 (14), 6132-6138 (2010).
- Olivier, N., Keller, D., Rajan, V. S., Gönczy, P., Manley, S. Simple buffers for 3D STORM microscopy. Biomedical Optics Express. 4 (6), 885-899 (2013).
- Mund, M., et al. Superresolution microscopy reveals partial preassembly and subsequent bending of the clathrin coat during endocytosis. bioRxiv. , (2022).
- Mund, M., et al. Systematic nanoscale analysis of endocytosis links efficient vesicle formation to patterned actin nucleation. Cell. 174 (4), 884-896 (2018).
- Sabinina, V. J., et al. Three-dimensional superresolution fluorescence microscopy maps the variable molecular architecture of the nuclear pore complex. Molecular Biology of the Cell. 32 (17), 1523-1533 (2021).
- Cieslinski, K., et al. Nanoscale structural organization and stoichiometry of the budding yeast kinetochore. bioRxiv. , (2021).
- Sieben, C., Banterle, N., Douglass, K. M., Gönczy, P., Manley, S. Multicolor single-particle reconstruction of protein complexes. Nature Methods. 15 (10), 777-780 (2018).
- Sarkans, U., et al. The BioStudies database—one stop shop for all data supporting a life sciences study. Nucleic Acids Research. 46, (2018).
Reimpresiones y Permisos
Solicitar permiso para reutilizar el texto o las figuras de este JoVE artículos
Solicitar permisoThis article has been published
Video Coming Soon
ACERCA DE JoVE
Copyright © 2025 MyJoVE Corporation. Todos los derechos reservados