
Method Article
Microscopia de super-resolução do complexo sinaptonemal dentro da linha germinal de Caenorhabditis elegans
Neste Artigo
Resumo
A microscopia de super-resolução pode fornecer uma visão detalhada da organização dos componentes dentro do complexo sinaptonemal na meiose. Aqui, demonstramos um protocolo para resolver proteínas individuais do complexo sinaptonemal de Caenorhabditis elegans .
Resumo
Durante a meiose, os cromossomos homólogos devem reconhecer e aderir uns aos outros para permitir sua segregação correta. Um dos principais eventos que assegura a interação dos cromossomos homólogos é a montagem do complexo sinaptonemal (SC) na prófase meiótica I. Embora haja pouca homologia de sequência entre os componentes proteicos dentro do SC entre diferentes espécies, a estrutura geral do SC tem sido altamente conservada durante a evolução. Em micrografias eletrônicas, o SC aparece como uma estrutura tripartida, semelhante a uma escada, composta de elementos laterais ou eixos, filamentos transversais e um elemento central.
No entanto, identificar com precisão a localização de componentes individuais dentro do complexo por microscopia eletrônica para determinar a estrutura molecular do SC permanece desafiador. Por outro lado, a microscopia de fluorescência permite a identificação de componentes proteicos individuais dentro do complexo. No entanto, uma vez que o SC tem apenas ~ 100 nm de largura, sua subestrutura não pode ser resolvida por microscopia de fluorescência convencional limitada por difração. Assim, a determinação da arquitetura molecular do SC requer técnicas de microscopia de luz de super-resolução, como microscopia de iluminação estruturada (SIM), microscopia de depleção de emissão estimulada (STED) ou microscopia de localização de molécula única (SMLM).
Para manter a estrutura e as interações de componentes individuais dentro do SC, é importante observar o complexo em um ambiente que esteja próximo ao seu ambiente nativo nas células germinativas. Portanto, demonstramos um protocolo de imuno-histoquímica e imagem que permite o estudo da subestrutura do SC em tecido germinativo de Caenorhabditis elegans extrudado e intacto com SMLM e microscopia STED. A fixação direta do tecido à lâmina de cobertura reduz o movimento das amostras durante a imagem e minimiza as aberrações na amostra para alcançar a alta resolução necessária para visualizar a subestrutura do SC em seu contexto biológico.
Introdução
Reduzir o número de cromossomos pela metade durante a meiose é fundamental para gerar progênie saudável em organismos que se reproduzem sexualmente. Para alcançar essa redução no número de cromossomos, os cromossomos homólogos devem emparelhar e segregar durante a meiose I. Para garantir a segregação precisa dos cromossomos homólogos, as células germinativas passam por uma prófase I estendida, durante a qual os cromossomos homólogos emparelham, fazem sinapse e se recombinam para gerar ligações físicas entre os homólogos1. O SC emergiu como a estrutura central que é fundamental na regulação da progressão correta através da prófasemeiótica 2.
O SC é um complexo cuja estrutura geral é conservada evolutivamente, embora haja pouca homologia entre seus componentes proteicos. O SC foi identificado pela primeira vez em micrografias eletrônicas como uma estrutura tripartida, semelhante a uma escada, consistindo de dois elementos laterais ou eixos, uma região central formada por filamentos transversais e um elemento central 3,4. Determinar a organização de componentes individuais dentro do complexo é fundamental para avançar nossa compreensão do papel do SC durante a prófase meiótica.
O organismo modelo C. elegans é ideal para estudar a estrutura e a função do SC, uma vez que suas linhas germinativas contêm um grande número de núcleos meióticos com SCs totalmente montados5. Estudos genéticos e bioquímicos revelaram que os eixos cromossômicos são formados por três complexos distintos de coesina6,7 e quatro proteínas do domínio HORMA chamadas HTP-1/2/3 e HIM-3 7,8,9,10,11 em C. elegans. Na região central do SC, seis proteínas contendo domínios de bobina enrolada foram identificadas até o momento 12,13,14,15,16,17. Para preencher a distância entre os dois eixos, SYP-1, -5 e -6 dimerizam de forma cabeça a cabeça (Figura 1), enquanto três proteínas adicionais estabilizam sua interação no elemento central16,17,18,19.
Obter uma visão detalhada sobre a organização dessas proteínas é essencial para entender as muitas funções do SC durante a meiose. Como a largura da região central do SC é de apenas ~100 nm, sua subestrutura não pode ser resolvida por microscopia de fluorescência limitada por difração. No entanto, a visualização de componentes dentro de uma estrutura desse tamanho é facilmente alcançável por microscopia de super-resolução. De fato, a microscopia de iluminação estruturada (SIM), a microscopia de expansão 20, a microscopia de depleção por emissão estimulada (STED) 21 e a microscopia de localização de molécula única (SMLM)22,23 emergiram como ferramentas essenciais para estudar a arquitetura molecular da SC em todas as espécies 16,24,25,26,27,28,29, 30.
Para superar o limite de resolução, a microscopia STED baseia-se na sobreposição do ponto limitado por difração da luz de emissão com um feixe em forma de rosquinha do laser STED, que teoricamente contrai a função de propagação pontual até dimensões moleculares31,32. No entanto, a resolução que é praticamente alcançável pelo STED dentro de amostras biológicas permanece na faixa de algumas dezenas de nanômetros em xy33.
Uma resolução ainda maior em amostras biológicas pode ser obtida com técnicas de SMLM. O SMLM aproveita as propriedades piscantes de fluoróforos específicos para resolver objetos no nível de subdifração separando fluoróforos espacialmente sobrepostos no tempo. A amostra é então fotografada repetidamente para capturar diferentes subconjuntos de fluoróforos. A posição dos fluoróforos dentro da amostra é então determinada ajustando-se a função de dispersão pontual (PSF) aos sinais obtidos em todas as imagens, o que pode resolver estruturas até 15 nm23,34.
Tomadas em conjunto, as imagens localizadas codificam as posições de todos os fluoróforos. A resolução do SMLM é determinada pela densidade de marcação e pelas características de piscar do fluoróforo. De acordo com o critério de Nyquist-Shannon, é impossível resolver de forma confiável objetos que são menos do que o dobro da distância média de rótulo a rótulo. Assim, uma alta densidade de rotulagem é necessária para imagens de alta resolução. Para o SC em C. elegans, uma alta densidade de marcação pode ser alcançada usando tags de epítopos anexadas a locais específicos de proteínas endógenas usando edição de genoma. As marcas de epítopos podem então ser coradas em alta densidade utilizando anticorpos monoclonais específicos com altas afinidades19,30. Ao mesmo tempo, o ciclo de fluoróforos individuais deve ser curto o suficiente para garantir que os fluoróforos espacialmente sobrepostos não sejam capturados ao mesmo tempo35.
Devido a esses dois requisitos, a resolução da estrutura de grandes complexos macromoleculares, como o SC, requer imagens de um número suficientemente grande de imagens e, portanto, pode levar várias horas. A armadilha de longos tempos de imagem é que as amostras tendem a derivar devido ao movimento do estágio ou pequenas correntes dentro do buffer da amostra; mesmo pequenos movimentos na ordem de 10 nm são prejudiciais na resolução nm e devem ser corrigidos. No entanto, os métodos de correção de deriva comumente utilizados não são robustos o suficiente para sobrepor com precisão imagens de dois canais fotografados sequencialmente36. Isso é problemático porque as questões biológicas geralmente pedem detecção e localização precisas de vários alvos dentro da mesma amostra. Para contornar esses problemas, métodos como imagens ratiométricas foram desenvolvidos. A imagem ratiométrica permite a imagem simultânea de múltiplos fluoróforos com espectros de excitação e emissão sobrepostos, com uma atribuição subsequente de cada sinal detectado ao seu respectivo fluoróforo com base na razão de intensidades em canais espectralmente distintos37,38.
Além disso, estudar a organização de complexos macromoleculares, como o SC, exige informações tridimensionais (3D). Para alcançar a super-resolução em três dimensões (3D-SMLM), uma lente cilíndrica é incorporada no caminho óptico da luz emitida que distorce a forma do PSF de um fluoróforo, dependendo da sua distância do plano focal. Assim, a posição precisa de um fluoróforo no plano z pode ser extrapolada pela análise da forma de seu sinal de emissão35,39. A combinação desses avanços no SMLM permite a obtenção de imagens da organização 3D de complexos macromoleculares, incluindo o SC.
Protocolo
1. Preparação de soluções e folhas de cobertura
NOTA: Consulte a Tabela de Materiais para obter detalhes relacionados a todos os materiais e reagentes e a Tabela 1 para a composição das soluções usadas neste protocolo.
- Coberturas revestidas com poli-L-lisina
- Preparar 0,01% (p/v) de poli-L-lisina (ver Tabela 1).
- Lave uma tampa de precisão (24 mm de diâmetro; 0,17 ± 0,005 mm, nº 1,5) em etanol por 10-30 min. Lave a tampa com ddH2O para remover o etanol e deixe a tampa secar à temperatura ambiente.
- Limpe a tampa a plasma usando um limpador de plasma.
NOTA: A limpeza a plasma aumenta a hidrofilicidade da tampa e facilita as seguintes etapas. Se um limpador de plasma não estiver disponível, esta etapa pode ser ignorada, embora isso possa exigir o ajuste do volume e / ou concentração da solução de poli-L-lisina. Esta modificação não foi testada. - Coloque uma gota (120 μL) de poli-L-lisina a 0,01% (p/v) no deslizamento da tampa. Incubar por 10 min à temperatura ambiente.
- Após a incubação, enxaguar a folha de cobertura em ddH2O e secar à temperatura ambiente. Conservar a 4 °C até 1 mês.
- Fragmentos de F(ab')2 conjugados com corantes orgânicos fluorescentes
- Adicione a seguinte ordem a um tubo de PCR: 10 μL de 0,6-0,7 mg/mL de fragmento F(ab')2 em PBS, 1 μL de 0,1 M de NaHCO 3 (pH8,3 ) e 1 μL de 1 mM de fluoróforo reativo de éster succinimidil (NHS) em DMSO (razão molar de F(ab')2:corante é ~1:17). Misture bem pipetando para cima e para baixo.
- Incubar durante 1 h à temperatura ambiente.
- Separe o fragmento F(ab')2 do corante reativo livre restante usando uma coluna de dessalinização (7K MWCO) de acordo com as especificações do fabricante. Use 1x PBS para equilíbrio da coluna e eluição do fragmento rotulado F(ab')2.
- Conservar o fragmento de F(ab')2 rotulado a 4 °C durante um período máximo de 3 meses.
NOTA: Tempos de armazenamento superiores a 3 meses não foram testados.
2. Dissecção e fixação
NOTA: Os procedimentos de dissecção e fixação são modificados em relação aos procedimentos recomendados anteriormente16,40 para obter amostras ótimas para microscopia de super-resolução.
- Dissecção
- Colher vermes de C. elegans pareados por idade (cultivados a 20 °C para este estudo) em uma gota de 30 μL de EBTT (1x tampão de ovo41 com detergente não iônico a 0,2%, Tabela 1) em uma folha de cobertura (22 mm x 22 mm, No. 1). Coloque a tampa em uma lâmina de vidro para facilitar a manipulação. Lave com 30 μL de EBTT pipetando para cima e para baixo várias vezes. Remova 30 μL da solução para deixar uma gota de 30 μL na folha de cobertura.
NOTA: Pequenas quantidades de detergente não iônico devem ser adicionadas a todas as soluções em que os vermes são pipetados para evitar que os vermes grudem nas pontas de plástico. - Use uma lâmina de bisturi para cortar as cabeças e/ou as caudas dos vermes para extrudar a gônada (Figura 2A).
- Colher vermes de C. elegans pareados por idade (cultivados a 20 °C para este estudo) em uma gota de 30 μL de EBTT (1x tampão de ovo41 com detergente não iônico a 0,2%, Tabela 1) em uma folha de cobertura (22 mm x 22 mm, No. 1). Coloque a tampa em uma lâmina de vidro para facilitar a manipulação. Lave com 30 μL de EBTT pipetando para cima e para baixo várias vezes. Remova 30 μL da solução para deixar uma gota de 30 μL na folha de cobertura.
- Fixação
- Pipetar 30 μL de solução fixadora (Tabela 1) na gota dos vermes dissecados e pipetar para cima e para baixo para misturar.
NOTA: Agitar para cima e para baixo algumas vezes pode ajudar a liberar mais gônadas. - Corrija por exatamente 1 min depois de adicionar a solução fixadora.
- Pare a fixação transferindo os vermes para um tubo de PCR preenchido com TBST (Tabela 1). Transfira os vermes no menor volume possível (~15 μL).
- Gire o tubo de PCR em uma mini centrífuga de bancada (2.000 × g, 10 s). Retire o sobrenadante e lave 2x com 200 μL de TBST cada.
- Lave com 200 μL de PBST (Tabela 1) por 5-10 min. Repetir os passos 2.1.1 a 2.2.5 para até quatro amostras, mantendo as amostras dissecadas no gelo.
NOTA: Se estiver a processar mais de quatro amostras, prossiga com os passos 2.2.6 a 2.2.7 após cada quatro amostras para garantir que a fixação da amostra permanece consistente em todas as amostras. - Gire as amostras dissecadas em uma mini centrífuga de bancada (2.000 × g, 10 s), remova o PBST e adicione 50-100 μL de metanol frio (-20 °C).
CUIDADO: O metanol é tóxico. Use equipamentos de proteção e evite a inalação. - Misture pipetando para cima e para baixo e deixe as amostras em metanol por 30-60 s. Lave as amostras 2x em 200 μL de PBST.
NOTA: Se estiver a processar mais de quatro amostras, proceda à dissecação das restantes amostras (passos 2.1.1 a 2.2.7). - Lave as amostras uma terceira vez com 200 μL de PBST.
- Pipetar 30 μL de solução fixadora (Tabela 1) na gota dos vermes dissecados e pipetar para cima e para baixo para misturar.
3. Incubações de anticorpos
- Bloqueio
- Bloquear as amostras em Solução de Bloqueio 1x (Tabela 1) por 45-60 min à temperatura ambiente.
NOTA: O tempo de incubação pode variar de 30 min à temperatura ambiente a vários dias a 4 °C (o teste foi feito até 3 dias).
- Bloquear as amostras em Solução de Bloqueio 1x (Tabela 1) por 45-60 min à temperatura ambiente.
- Solução primária de anticorpos
- Diluir anticorpos anti-HTP-3 (frango42) e anti-HA (rato) (ou os anticorpos de eleição) para as soluções de trabalho (1:250 para SMLM e 1:1.000 para amostras de microscopia STED) em 1x Solução de Bloqueio.
NOTA: Os anticorpos utilizados para rotular amostras SMLM são mais concentrados do que para amostras STED, uma vez que uma maior densidade de marcação é recomendada para a microscopia SMLM. - Gire as amostras em uma mini centrífuga de bancada (2.000 × g, 10 s), remova o tampão de bloqueio e adicione 30-50 μL da solução primária de anticorpos. Incubar durante a noite a 4 °C (de preferência) ou durante 1-2 h à temperatura ambiente.
- Após a incubação, lave 3 x 5-15 min com PBST.
- Diluir anticorpos anti-HTP-3 (frango42) e anti-HA (rato) (ou os anticorpos de eleição) para as soluções de trabalho (1:250 para SMLM e 1:1.000 para amostras de microscopia STED) em 1x Solução de Bloqueio.
- Solução de trabalho de fragmentos de F(ab')2 conjugados a corante fluorescente
- Diluir os fragmentos de F(ab')2 rotulados (passo 1.2.4) para as soluções de trabalho (1:100 para SMLM e 1:1.000 para amostras de microscopia STED) em 1x Solução de Bloqueio.
NOTA: Para ambas as técnicas de super-resolução, foram utilizados pares de fluoróforos previamente relatados, nomeadamente AlexaFluor647/CF680 para SMLM e AlexaFluor594/Abberior STAR635P para STED. AlexaFluor647 e STAR645P foram usados para rotular fragmentos anti-rato (Fab')2 para atingir o terminal C de SYP-5 e fragmentos anti-frango (Fab')2 marcados com CF680/AlexaFluor594 para atingir o HTP-3. - Gire as amostras em uma mini centrífuga de bancada (2.000 × g, 10 s), remova o PBST e adicione 30-50 μL de solução de anticorpos secundários. Incubar durante 30 min a 2 h à temperatura ambiente (de preferência) ou durante a noite a 4 °C. Lave 3 x 5-15 min com PBST.
- Diluir os fragmentos de F(ab')2 rotulados (passo 1.2.4) para as soluções de trabalho (1:100 para SMLM e 1:1.000 para amostras de microscopia STED) em 1x Solução de Bloqueio.
4. Amostras de montagem em uma folha de cobertura
- Pós-fixação
NOTA: Processar amostras individualmente através das etapas 4.1.1-4.2.1.- Gire as amostras coradas e remova o sobrenadante. Adicionar 50 μL de PBST0,2 e transferir os vermes corados para uma folha de cobertura n.º 1 de 22 mm x 22 mm.
NOTA: Utilize PBST 0,2 fresco com detergente não iónico a0,2 % (Tabela 1) para este passo para evitar que os vermes adiram à folha de cobertura. - Pipetar 5,7-6,3 μL de solução pós-fixadora em uma tampa de poli-L-lisina.
NOTA: As folhas de cobertura de poli-L-lisina armazenadas a 4 °C devem ser primeiro levadas à temperatura ambiente. - Pipetar os vermes dissecados no mesmo volume (5,7-6,3 μL) e transferir para a gota do fixador no deslizamento da cobertura de poli-L-lisina (Figura 2B).
NOTA: Nesta e na etapa seguinte, é muito importante reter o tecido dissecado no centro da tampa revestida com poli-L-lisina. Isso é especialmente importante se a montagem das amostras em um suporte personalizado para se ajustar ao microscópio SMLM personalizado usado aqui (consulte a etapa 5.1, Figura 2B). - Cobrir a amostra com uma pequena folha de cobertura (12 mm de diâmetro, figura 2B). Remova o excesso de líquido usando um pequeno pedaço de papel de filtro (Figura 2B). Fixe por 3-5 min em uma câmara escura.
- Gire as amostras coradas e remova o sobrenadante. Adicionar 50 μL de PBST0,2 e transferir os vermes corados para uma folha de cobertura n.º 1 de 22 mm x 22 mm.
- "Congelamento-rachadura"
- Congelar as amostras colocando-as em um bloco de alumínio em gelo seco (Figura 2B).
NOTA: O bloco de alumínio deve ser bem resfriado no gelo seco antes de colocar as amostras sobre ele. Proceder à pós-fixação das restantes amostras (passos 4.1.1 a 4.2.1). A amostra deve estar em gelo seco durante, pelo menos, 20 minutos ou até 1 h antes da fase seguinte (4.2.2). - Remova a tampa menor usando uma navalha (Figura 2B).
NOTA: Para STED, avance para o passo 5.2.1. Para SMLM, continue com a etapa 4.2.3. - Mergulhe a tampa num tubo cónico de 50 ml contendo PBS gelado (de preferência) ou metanol de -20 °C durante aproximadamente 10 s.
NOTA: A temperatura é um fator muito importante para esta etapa. Portanto, use PBS que é recém-descongelado ou mantido em um banho de gelo / etanol. - Coloque a tampa em um poço de uma placa de seis poços preenchida com buffer PBST. Remova o PBST dos poços e adicione o PBS fresco. Deixe as amostras em PBS por 5 min.
NOTA: Pipetar o PBS para o lado do poço para evitar danificar e separar as amostras. - Lavar com PBS fresco e deixar as amostras a 4 °C até à obtenção de imaginos.
NOTA: As amostras são estáveis por até 2 semanas, mas os melhores resultados são alcançados se as amostras forem fotografadas dentro de 2 dias. - Antes da imagem, avalie a qualidade da montagem da amostra sob um microscópio estéreo.
NOTA: As linhas germinativas montadas com sucesso são fixadas de forma estável, sem movimento discernível em relação à tampa. Linhas germinativas mal anexadas baterão na solução tampão.
- Congelar as amostras colocando-as em um bloco de alumínio em gelo seco (Figura 2B).
5. Imagens
- Microscopia de localização de molécula única
NOTA: As imagens foram adquiridas no EMBL Imaging Centre usando um microscópio de localização de molécula única personalizado que foi construído em torno de um corpo personalizado, conforme relatado anteriormente38,43, com os recursos exclusivos especificados no Tabela de Materiais; consulte https://www.embl.org/about/info/imaging-centre- Adquirindo calibração de grânulos 3D
- Preparar uma tampa de precisão (24 mm de diâmetro; 0,17 ± 0,005 mm, nº 1,5) com esferas fluorescentes aderentes de 100 nm, conforme descrito anteriormente38,44.
- Colocar a amostra de calibração da etapa 5.1.1.1 num suporte de amostra.
- Adicione uma gota de óleo de imersão na objetiva de óleo limpa de 100x/1,5 e monte a amostra de calibração no microscópio.
- No MicroManager 2 45,46, especifique15-20 posições na amostra de calibração.
- Dentro da janela47 do plug-in da UEM, configure a aquisição de uma imagem z-stack para cada uma das posições a partir da etapa 5.1.1.5.
NOTA: Aqui, uma lente cilíndrica composta fornece o astigmatismo necessário para imagens 3D, e 201 z-slices foram adquiridas para cada posição abrangendo a faixa entre -1 μm a 1 μm, com um incremento de 10 nm. Uma iluminação a laserde 2 kW/cm 2 640 nm foi utilizada para 25 ms para cada z-slice. - Adquira as imagens z-stack dos grânulos fluorescentes de 100 nm através de um caminho óptico idêntico que será usado para adquirir imagens de amostra na etapa 5.1.11.
- Usando a plataforma de análise de microscopia de super-resolução (SMAP48), gere um modelo cspline da função experimental de propagação de ponto (PSF) que será usado para ajustar os dados 3D-SMLM na etapa 5.1.13.
- Prepare o suporte da amostra. Para o suporte personalizado usado aqui que usa um anel magnético para criar a câmara de imagem (Figura 2B), envolva o anel magnético com parafilme.
NOTA: Alternativamente, uma lâmina de microscópio com uma cavidade de depressão côncava pode ser usada para montar amostras para microscópios com suportes de lâmina. - Preparar 1 mL de tampão de imagem44 (Tabela 1).
- Pegue uma tampa da etapa 4.2.6 e coloque-a no suporte personalizado. Fixar a folha de cobertura no suporte com o anel magnético embrulhado em parafilme (passo 5.1.2).
- Pipetar suavemente o tampão de imagem (etapa 5.1.3) na câmara criada pelo anel magnético no topo da amostra (Figura 2B). Sele a câmara com um pedaço de parafilme.
- Para montar a amostra, adicione uma gota de óleo de imersão na objetiva de óleo limpa de 100x/1,5. Sem introduzir qualquer ar no óleo de imersão, coloque suavemente o suporte da amostra com a amostra montada (passo 5.1.5) na fase do microscópio.
NOTA: Antes de colocar a amostra no microscópio, limpe o fundo da tampa com tecido e etanol a 70%. - Usando a janela 47 do plug-in da UEM dentro do MicroManager 2 45,46, mova o estágio piezo até que o sinal do laser de bloqueio de foco seja detectado no fotodiodo do quadrante (QPD).
NOTA: Para manter um foco fixo durante todo o tempo de imagem, o bloqueio do foco é alcançado pela reflexão interna total de um laser acoplado à fibra de infravermelho próximo a partir do deslizamento da tampa e subsequente detecção sensível à altura em um fotodiodo de quadrante (QPD). O sinal QPD fornecia controle de malha fechada da montagem piezo da lente objetiva. - Adquira uma imagem do plano focal traseiro com um laser de excitação de 640 nm em baixa potência (ou seja, 1-5%) para confirmar a ausência de bolhas de ar no óleo de imersão.
Observação : remova a amostra do estágio se uma bolha de ar for detectada. Limpe a parte inferior da folha de cobertura e o objetivo e repita as etapas 5.1.6-5.1.8. Caso contrário, proceda ao bloqueio da concentração no âmbito do software da UEM47. - Localize o tecido da gônada usando a iluminação de campo brilhante. Usando uma iluminação de baixa intensidade de 640 nm, concentre-se na seção do tecido que contém muitos trechos SC.
NOTA: Não se concentre em estruturas que estejam a mais de 2 μm da folha de cobertura. Não use uma potência de laser maior para localizar a amostra, pois isso pode transformar alguns fluoróforos em um estado de piscar prematuramente. Aqui, 1 kW/cm2 foi usado no modo de subida com um pulso ajustado para 1.000. - Proceder à exposição da amostra com iluminação de 640 nm a uma irradiância elevada (27 kW/cm2) durante ~30 s até se atingir uma taxa de piscar adequada (Vídeo Suplementar 1).
- Adquira 200.000 quadros com um tempo de exposição de 20 ms usando a ferramenta de aquisição multidimensional no MicroManager 245,46.
- Enquanto isso, configure a ativação UV usando a opção de ativação do pluginEMU 38,47 para manter a taxa de piscar desejada.
NOTA: Use o laser UV a uma irradiância de 3 kW/cm2 no modo de subida com comprimento máximo de pulso definido para 10.000. - Realizar reconstrução e pós-processamento de imagens SMLM.
NOTA: Para reconstruir imagens a partir de dados SMLM brutos, consulte os métodos publicados. Os dados aqui apresentados foram processados utilizando-se o software SMAP 48,49. Reconstrução de imagem super-resolvida, atribuição de canais, correção de desvio e filtragem de localizações com baixa precisão de localização e filtro de máxima verossimilhança foram realizadas no software SMAP48.
- Adquirindo calibração de grânulos 3D
- Microscopia de depleção de emissão estimulada
NOTA: As imagens foram adquiridas no sistema microscópico STED integrado equipado com um laser de luz branca, um laser STED pulsado de 775 nm e o módulo de envelhecimento FALCON Fluorescence Lifetime IM(Table of Materials) no EMBL Imaging Centre (https://www.embl.org/about/info/imaging-centre).- Coloque uma gota de 20 μL de meio de montagem (Tabela de Materiais) em uma lâmina de microscópio. Pegue uma tampa da etapa 4.2.2 e coloque a amostra suavemente no slide voltado para o meio de montagem (Figura 2B).
NOTA: Evite introduzir bolsas de ar dentro do meio de montagem. - Deixe o meio de montagem curar durante a noite.
NOTA: Apresentar uma imagem das amostras no dia seguinte ou mantê-las a 4 °C até à obtenção de imaginos. - Para montar a amostra, adicionar uma gota de óleo de imersão na folha de cobertura da amostra a partir do passo 5.2.2. Coloque suavemente a amostra no estágio do microscópio usando uma objetiva de óleo de 100x/1,40.
- Concentre-se na amostra e localize o tecido da linha germinativa usando a iluminação de campo brilhante.
- Usando o software do microscópio, especifique a região de interesse para a qual a imagem TauSTED será adquirida.
- Selecione os lasers de excitação e sua potência apropriada usada para excitar os fluoróforos usados na amostra.
NOTA: Aqui, o laser de 580 nm a 4% de potência foi usado para visualizar fragmentos de anticorpos secundários F(ab')2 conjugados com AlexaFluor 594 e 635 nm a 3% de potência para visualizar fragmentos de F(ab')2 conjugados com STAR 635P. - Usando o software do microscópio, selecione uma potência de laser de depleção STED apropriada e configure a detecção de imagem.
NOTA: Aqui, a potência do laser de depleção STED de 775 nm foi ajustada para 40%. O detector foi utilizado no modo de contagem com um valor de ganho de 10 para detecção de fótons, com uma taxa de varredura de 100 Hz, e a um tamanho de pixel de 17 nm. O acúmulo de quatro linhas foi utilizado para a aquisição do TauSTED.
- Coloque uma gota de 20 μL de meio de montagem (Tabela de Materiais) em uma lâmina de microscópio. Pegue uma tampa da etapa 4.2.2 e coloque a amostra suavemente no slide voltado para o meio de montagem (Figura 2B).
Resultados
Para a imagem do SC dentro do tecido germinativo de C. elegans por SMLM, empregamos 3D-SMLM ratiometric de 2 cores para localizar o HTP-3, um componente dos eixos cromossômicos, e o C-terminal do filamento transversal SYP-5 endogenamente marcado com uma tag hemaglutinina (HA). A localização de ambas as proteínas dentro do SC de C. elegans foi previamente determinada por outros estudos16,30.
Para minimizar o espalhamento de luz e as aberrações ópticas inerentes a amostras biológicas espessas, foram fotografadas a seção z mais inferior dos núcleos meióticos que contêm os SCs (Figura 3, linhas amarelas). Para cada imagem adquirida, a posição do estágio piezo do plano de imagem foi marcada em relação à posição do estágio piezo quando a objetiva foi focalizada na lâmina de cobertura. Isso permitiu o cálculo da distância piezo da folha de cobertura. As amostras montadas com sucesso são fixadas de forma estável perto da tampa e mantêm a forma da gônada (ou seja, o tecido não é esmagado entre as duas folhas de cobertura durante a etapa de pós-fixação). A qualidade da montagem da amostra pode ser facilmente avaliada sob um microscópio estéreo, uma vez que as gônadas bem fixadas não mostram nenhum movimento na solução (etapa 4.2.6). No entanto, devido à estocasticidade do processo de montagem, o tecido da gônada não será necessariamente colocado completamente plano na tampa. Portanto, o plano inferior dos núcleos contendo SCs pode ser encontrado a distâncias variáveis em relação à cobertura dentro da mesma gônada.
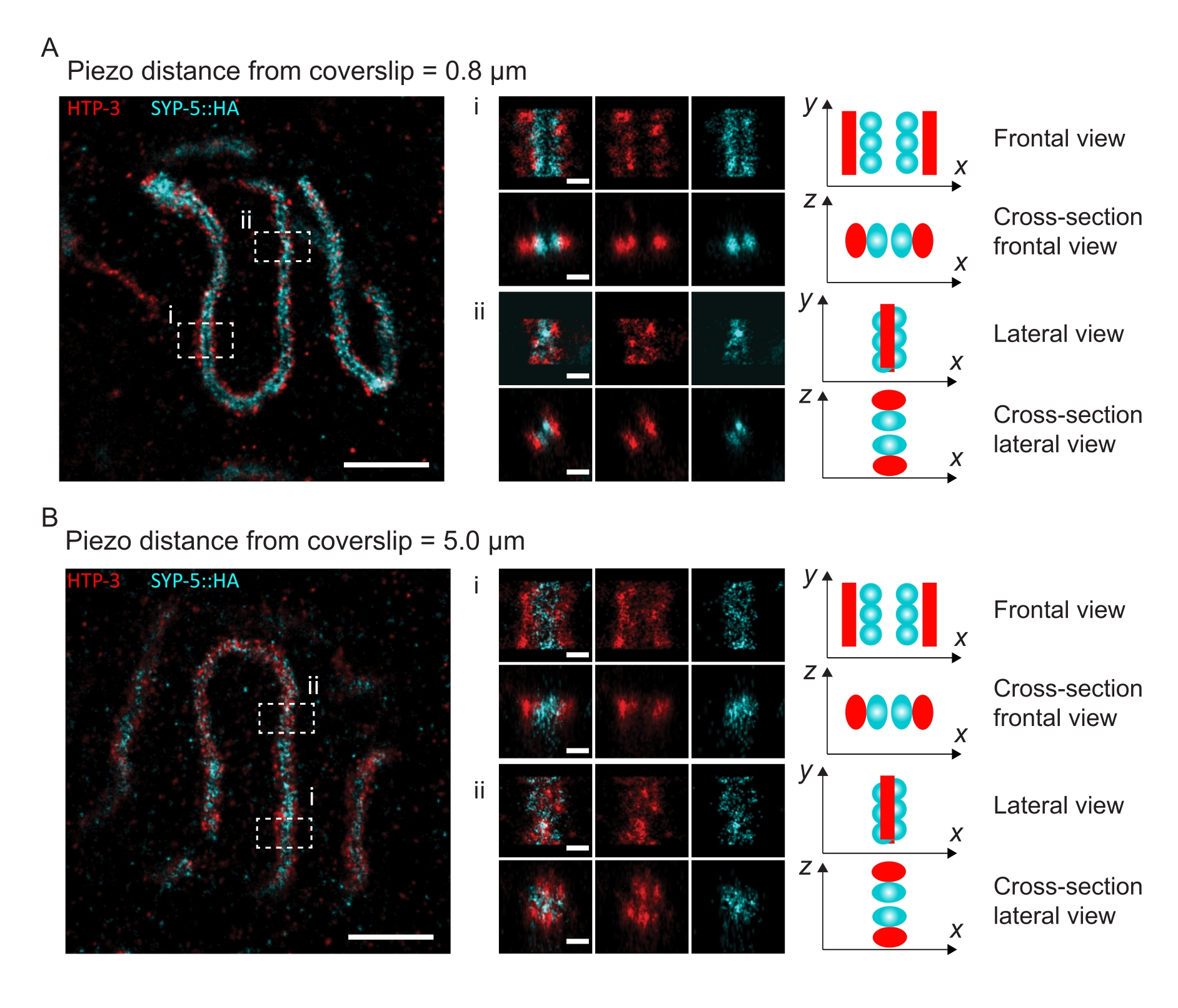
Para ilustrar como a resolução muda dependendo da fixação do tecido à folha de cobertura, adquirimos imagens em diferentes distâncias piezo para a folha de cobertura. Para avaliar a qualidade de uma imagem individual, foram calculadas curvas de correlação de anel de Fourier (FRC)50,51 e a resolução foi determinada usando o plugin FRCResolution dentro do software SMAP48. Dois núcleos representativos extraídos de duas imagens 3D-SMLM separadas tiradas a distâncias diferentes da folha de cobertura são exibidos na Figura 4. Em SCs localizados próximos ao deslizamento de cobertura, os eixos cromossômicos e o terminal C de SYP-5::HA são bem resolvidos em todas as três dimensões (Figura 4A, 0,8 μm do deslizamento de cobertura). Para resolver duas estruturas separadas por uma determinada distância, a resolução de FRC alcançada geralmente deve ser menor que a metade dessa distância na resolução axial.
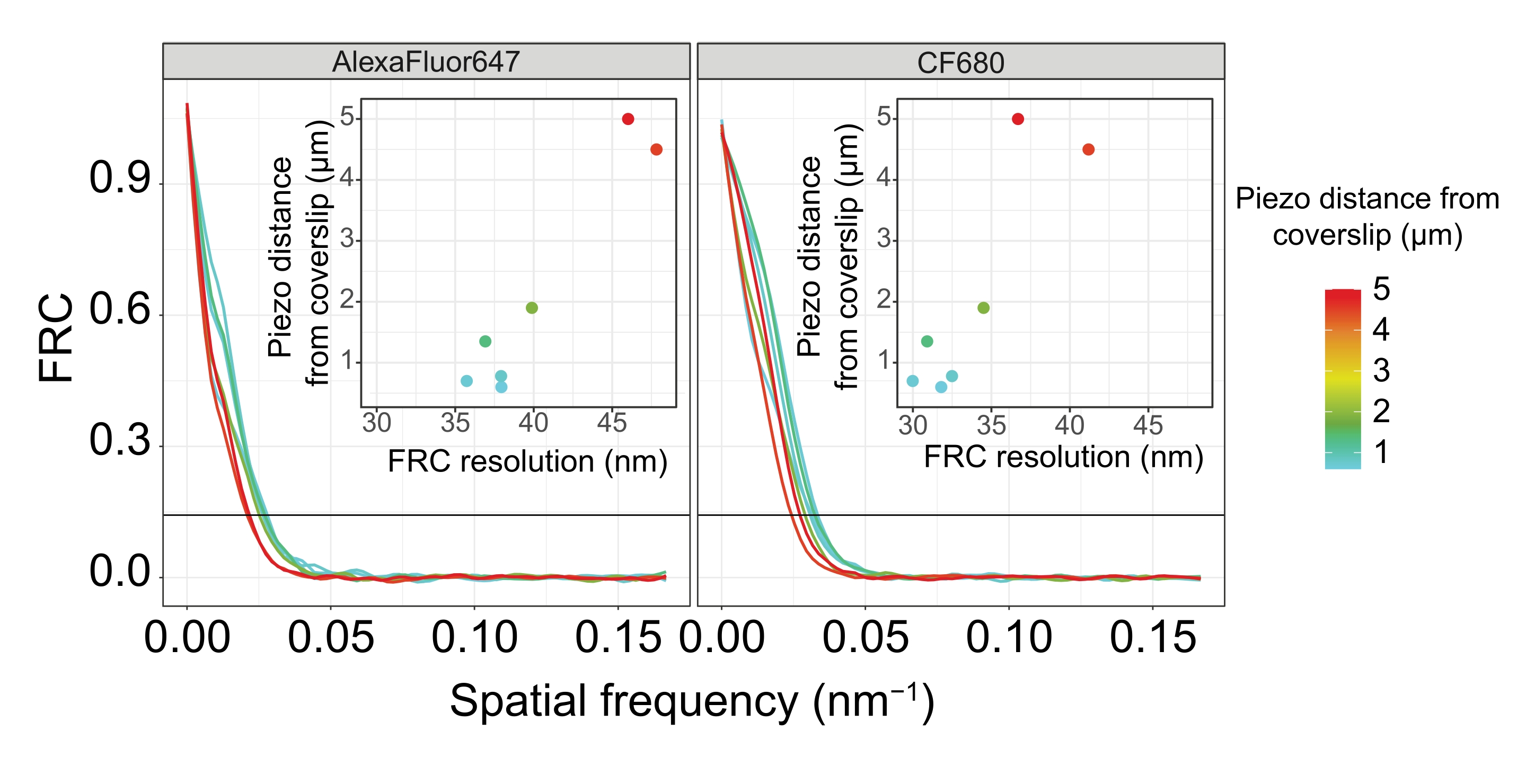
Para separar as mesmas estruturas lateralmente, valores de resolução de FRC ainda menores precisam ser alcançados. De fato, em amostras localizadas próximas ao deslizamento de cobertura, a resolução FRC é de 38 nm para o canal AlexaFluor 647 e 34 nm para o canal CF680 e, portanto, bem abaixo da distância esperada de 84 nm entre o C-termini do SYP-516. Esta resolução, portanto, resolve prontamente a organização do CS não apenas em vistas frontais, mas também em vistas laterais (Figura 4B i,ii). Por outro lado, a resolução se deteriora em SCs localizados a uma distância de 5 μm da cobertura devido ao espalhamento de luz e aberrações esféricas (Figura 4B). As resoluções FRC a esta distância caem para 47 nm (AlexaFluor 647) e 41 nm (CF680), o que não pode resolver totalmente o C-termini do SYP-5. Como as aberrações ópticas prejudicam a resolução lateral mais severamente do que a resolução axial, as bandas HTP-3 e SYP-5 não são mais claramente resolvidas na seção transversal da vista lateral em amostras localizadas a uma distância de 5 μm da cobertura (Figura 4B ii). A comparação da resolução FRC das imagens adquiridas a diferentes distâncias piezo da lâmina de cobertura revelou que o tecido fotografado não deve estar a mais de 2 μm da lâmina de cobertura (Figura 5). Este resultado destaca a importância da execução correta da etapa de pós-fixação, durante a qual o tecido deve ser reticulado com sucesso ao revestimento de poli-L-lisina da lâmina de cobertura.
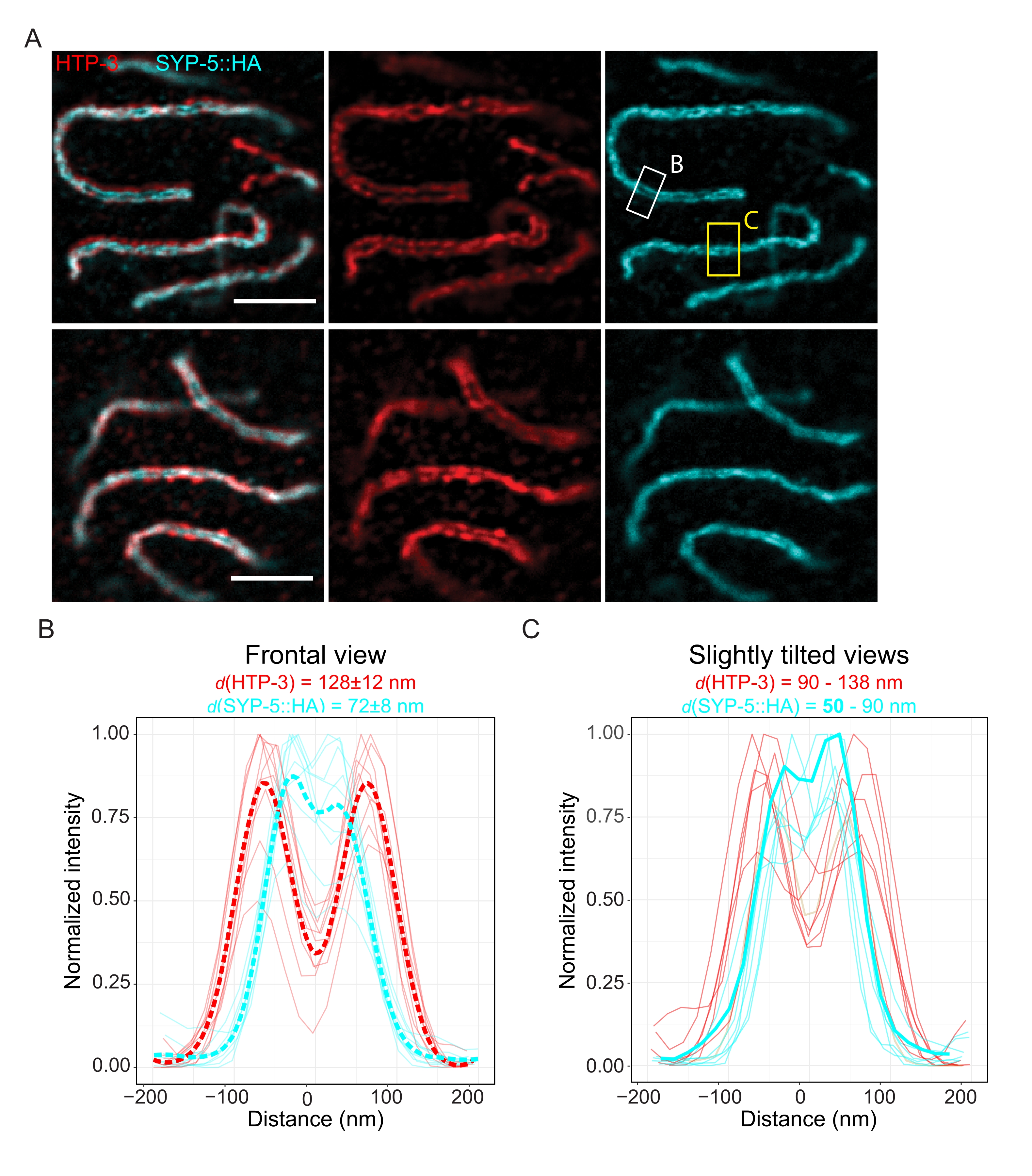
Para demonstrar a resolução alcançável com outra técnica de super-resolução, também fotografamos SCs em tecido germinativo intacto fixo com microscopia TauSTED. A Figura 6A mostra as imagens de TauSTED com a maior e menor resolução alcançadas neste estudo, estimadas a partir de perfis de linha do SC em vista frontal (Figura 6B). Em ambos os núcleos, foi possível resolver as duas bandas de localização do HTP-3 nos eixos cromossômicos e o C-termini do SYP-5 na região central, demonstrando que a resolução alcançável no TauSTED utilizando este protocolo otimizado é inferior a 84 nm. Em condições ótimas (Figura 6A, acima), foi possível resolver o C-termini em vistas levemente inclinadas do SC que estavam separadas por apenas 50 nm (Figura 6A, retângulo amarelo e 6C).

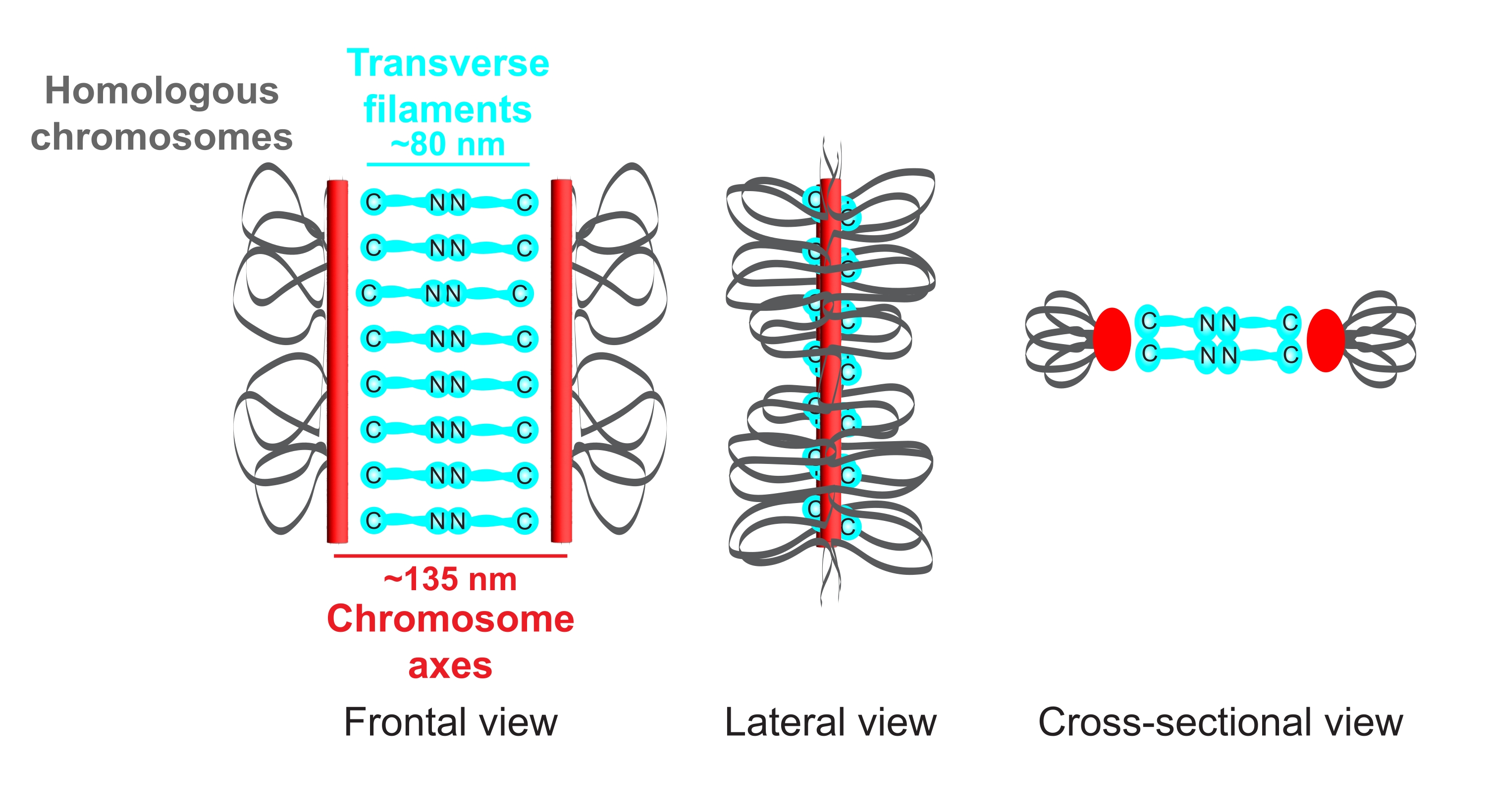
Figura 1: Esquema da organização do complexo sinaptonemal em Caenorhabditis elegans. O desenho animado mostra uma estrutura simplificada do SC em C. elegans fazendo a ponte entre dois cromossomos homólogos (cinza). A estrutura é mostrada nas vistas frontal, lateral e transversal. Os eixos cromossômicos são exibidos como barras vermelhas, enquanto os filamentos transversais são mostrados em ciano. As proteínas do filamento transversal (SYP-1, 5, 6 em C. elegans) são orientadas de forma cabeça a cabeça (gráficos de vara de bola ciano) na região central para preencher a distância entre os dois eixos. As distâncias esperadas entre os eixos e o C-termini dos filamentos transversais são indicadas. Abreviação: SC = complexo sinaptonemal. Por favor, clique aqui para ver uma versão maior desta figura.
{kind=link}

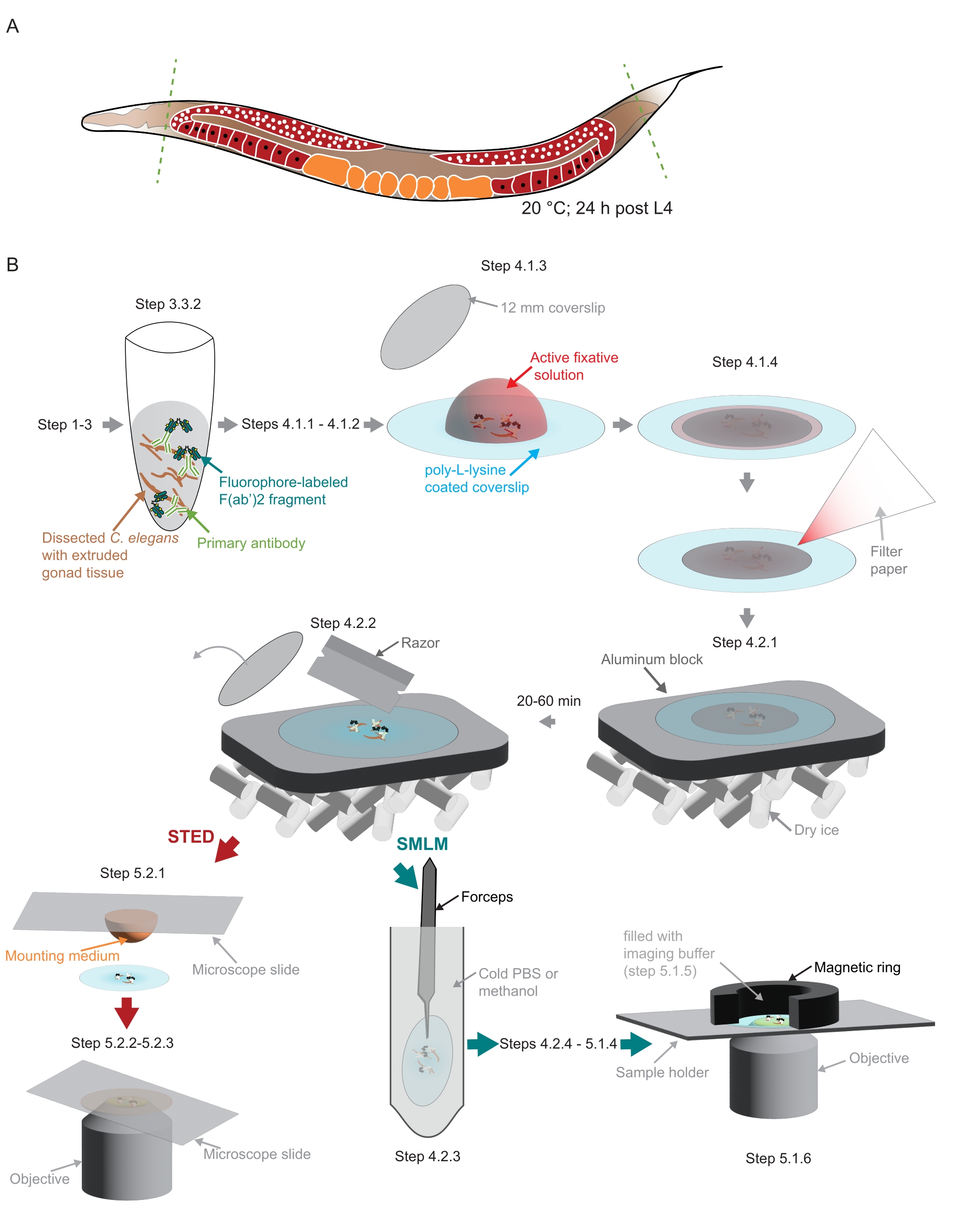
Figura 2: Ilustração do preparo da amostra utilizada no estudo . (A) Adultos jovens de C. elegans são dissecados na cabeça ou cauda (linhas verdes tracejadas) e processados conforme descrito no protocolo. (B) Etapas individuais do método são indicadas com gráficos que estão conectados com setas cinzas. Abreviaturas: STED = depleção de emissão estimulada; SMLM = microscopia de localização de molécula única; PBS = solução salina tamponada com fosfato. Por favor, clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 3: Localização da seção tecidual que pode ser observada por microscopia de localização de molécula única. MIP de uma imagem confocal de disco giratório de uma gônada inteira de montagem C. elegans. O tecido foi corado para HTP-3 e o terminal C de SYP-5 (SYP-5::HA), e o sinal combinado é mostrado em cinza. Imagens confocais individuais foram costuradas usando o plugin Grid/Collection stitching Fiji52 para criar uma imagem de toda a gônada. A inserção mostra uma exibição xy do plano z mais inferior que contém os SCs. A localização deste plano é mostrada em vistas ortogonais da seção tecidual indicada por um retângulo na imagem MIP da gônada (linhas amarelas). Barras de escala = 10 μm. Abreviaturas: PImáx = projeção de intensidade máxima; SCs = complexos sinaptonemais. Por favor, clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 4: Microscopia de localização de molécula única do HTP-3 e do C-termini do SYP-5. (A,B) Esquerda: Imagens SMLM mostrando núcleos de paquiteno corados para HTP-3 (vermelho) e o terminal C de SYP-5 (SYP-5::HA, ciano) (barra de escala = 1 μm). Centro: Imagens ampliadas de regiões de interesse indicadas em A e B com as visualizações transversais correspondentes exibidas abaixo de cada imagem (i, ii; barra de escala = 100 nm). Os trechos do SC dentro de imagens ampliadas são girados para orientar os eixos cromossômicos paralelos ao eixo y. Direita: Representação gráfica da localização das proteínas de interesse dentro do SC retratando a orientação do SC nas regiões ampliadas exibidas no centro da figura. Abreviaturas: SMLM = microscopia de localização de molécula única; SC = complexo sinaptonemal. Os dados brutos para reconstruir imagens SMLM estão disponíveis através da base de dados BioStudies60 (Accession ID: S-BIAD504). Por favor, clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 5: A resolução da correlação do anel de Fourier das imagens de microscopia de localização de molécula única depende da distância do plano z imageado do plano da folha de cobertura. As linhas coloridas mostram curvas FRC de imagens adquiridas a diferentes distâncias (conforme representado pela barra de cores) da folha de cobertura. O limite de 1/7 usado para determinar a resolução FRC é indicado por uma linha horizontal preta. As inserções mostram a dependência da resolução FRC da distância piezo da folha de cobertura. A plotagem foi realizada por um script R personalizado (versão 4.1.2, Arquivo Suplementar 1) no qual as curvas originais foram suavizadas com funções do pacote "ggplot2". Abreviaturas: FRC = correlação do anel de Fourier; SMLM = microscopia de localização de molécula única; SC = complexo sinaptonemal. Os dados relativos às curvas FRC e aos dados SMLM estão disponíveis na base de dados BioStudies60 (ID de Adesão: S-BIAD504). Por favor, clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 6: A microscopia de depleção de emissão estimulada aprimorada por informações baseadas no tempo de vida da fluorescência (TauSTED) resolve duas bandas de localização para o HTP-3 e o terminal C do SYP-5. (A) Duas imagens TauSTED representativas mostram núcleos de paquiteno corados para HTP-3 (vermelho) e o terminal C de SYP-5 (SYP-5::HA, ciano) com definição estrutural maior (superior) e inferior (inferior) (barra de escala = 1 μm). Os retângulos marcam regiões com o C-termini resolvido de SYP-5 em frontal (branco) e uma visão ligeiramente inclinada (amarelo) do SC. (B,C) Distribuição do HTP-3 (vermelho) e do C-terminal do sinal SYP-5 (ciano) resolvido por TauSTED. Os perfis de linha de regiões de interesse que contêm o SC em vistas frontal (B) ou ligeiramente inclinada (C) são mostrados como linhas completas com intensidade normalizada para o valor máximo. Os perfis de linha foram gerados usando o Fiji ImageJ. Linhas tracejadas em B mostram os dados médios para cada proteína. A linha ciano espessa em C corresponde ao perfil da linha com a menor distância resolvida entre o C-termini do SYP-5. Para determinar as distâncias entre os anticorpos direcionados a proteínas específicas, os perfis de linha (n = 9 (B), n = 7 (C)) foram equipados com gaussianos duplos usando um script R personalizado (versão 4.1.2, Arquivo Suplementar 1). A distância média ± desvio padrão (B) e o intervalo com valor mínimo destacado em negrito (C) são indicados no topo de cada gráfico, respectivamente. Abreviaturas: STED = microscopia de depleção de emissão estimulada; SC = complexo sinaptonemal. As imagens exibidas e os pontos de dados dos perfis de linhas plotadas estão disponíveis através do banco de dados BioStudies60 (ID de Adesão: S-BIAD504). Por favor, clique aqui para ver uma versão maior desta figura.
{kind=link}
Tabela 1: Composição dos buffers e soluções utilizados neste protocolo. Por favor, clique aqui para baixar esta Tabela.
Vídeo Suplementar 1: Aquisição de microscopia de localização de molécula única. Vídeo mostrando fluoróforos piscando a uma taxa apropriada (50 quadros são mostrados, barra de escala = 5 μm, 20 ms/quadro). Clique aqui para baixar este vídeo.
Arquivo Suplementar 1: Script de análise de dados. Clique aqui para baixar este arquivo.
Discussão
A organização em forma de escada do SC, essencial para a correta recombinação e segregação dos cromossomos homólogos, foi observada pela primeira vez há quase 70 anos na microscopia eletrônica 3,4. Embora a organização geral do SC seja prontamente resolvida na microscopia eletrônica, a localização de componentes individuais dentro desse complexo requer uma abordagem mais direcionada. Com sua largura de apenas ~100 nm, a subestrutura do SC não pode ser resolvida pela microscopia de fluorescência convencional. No entanto, a microscopia de super-resolução tornou-se um dos principais impulsionadores de novas descobertas sobre a estrutura e a função do complexo sinaptonemal 16,19,24,25,26,27,28,29,30. Para facilitar esta pesquisa, demonstramos um procedimento de montagem que permite estudar a arquitetura do SC dentro do tecido da gônada de C. elegans com SMLM e microscopia STED.
Um passo crítico para otimizar a resolução na imagem SMLM é a ligação direta do tecido germinativo a uma tampa revestida com poli-L-lisina (etapa 4). A fixação covalente do tecido à lâmina de cobertura é essencial para reduzir os movimentos dentro da amostra que resultariam em grandes derivas e impossibilitariam a imagem durante longos períodos de tempo para a LME. Além disso, mesmo uma fixação subótima que deixa os núcleos contendo SCs a uma distância da deslizamento de cobertura leva a uma queda significativa na resolução alcançável resultante de aberrações esféricas (Figura 4). Alternativamente ao acessório covalente aqui utilizado, o tecido corado da linha germinativa também pode ser imobilizado entre duas coberturas seladas em uma pequena gota de tampão de imagem19,30. No entanto, esse método de imobilização reduz severamente o volume de tampão de imagem na amostra de 1 mL utilizado no protocolo otimizado aqui para apenas alguns μL, o que resultará em uma acidificação do tampão de imagem e reduzirá severamente o tempo durante o qual a amostra pode ser fotografada 38,53,54.
Longos tempos de aquisição para microscopia SMLM e STED limitam o uso desses métodos à imagem de amostras quimicamente fixas. Aqui, a fixação de paraformaldeído garante que a estrutura do SC seja preservada durante a preparação da amostra e a imagem. No entanto, apesar das precauções tomadas aqui para a imagem do SC dentro do tecido intacto, a estrutura resultante do SC após a fixação não é necessariamente idêntica à estrutura em seu estado nativo dentro de um organismo vivo. Além disso, uma vez que uma única imagem do SC fixo representa um único "instantâneo" da estrutura biológica, essa abordagem permanece cega para a dinâmica da estrutura nativa in vivo.
No entanto, informações sobre a dinâmica e variabilidade das estruturas macromoleculares também podem ser obtidas através da aquisição não apenas de um único, mas muitos "instantâneos". Embora essa abordagem possa resolver mudanças na estrutura do SC durante o paquiteno19, existem vários fatores que limitam o número de imagens que podem ser adquiridas a partir de uma única amostra preparada usando esse protocolo. Primeiro, as altas potências do laser usadas durante a aquisição da imagem levam ao branqueamento permanente dos fluoróforos e impedem a imagem de regiões adjacentes de interesse ou múltiplos planos z, reduzindo significativamente o número de imagens que podem ser adquiridas de uma única amostra. Em segundo lugar, a densidade da amostra/tecido na folha de cobertura preparada por este método é baixa, o que limita significativamente o número de imagens que podem ser adquiridas a partir de uma única folha de cobertura. A baixa densidade amostral também proíbe o uso de pipelines automatizados de aquisição de imagens que ajudaram a esclarecer outras questões biológicas 34,55,56,57,58,59. No entanto, a densidade da amostra pode ser ligeiramente aumentada por um usuário experiente.
O protocolo aqui apresentado é otimizado para obter uma alta densidade de marcação que é necessária para alcançar a resolução ideal no SMLM35. Enquanto os protocolos anteriores anexam covalentemente o tecido à lâmina de cobertura antes da imunocoloração16, este novo protocolo reticula o tecido à lâmina de cobertura somente depois que as amostras foram coradas em solução. Essa modificação permite que os anticorpos usados para a imunomarcação acessem livremente o tecido de todos os lados, enquanto a fixação covalente do tecido à lâmina de cobertura pode restringir os anticorpos de atingir os núcleos mais próximos da lâmina de cobertura, reduzindo assim o grau de rotulagem. Juntas, as modificações descritas aqui melhoram a resolução de 40-50 nm (resolução FRC)16 para 30-40 nm (este protocolo).
É importante ressaltar que, embora uma alta densidade de marcação e uma alta concentração de anticorpos sejam essenciais para a SMLM, descobrimos que melhores imagens de microscopia STED são obtidas usando concentrações mais baixas de anticorpos (etapa 3). Com uma resolução de dezenas de nanômetros, o tamanho das moléculas usadas para rotular a proteína de interesse torna-se cada vez mais importante. Portanto, empregamos fragmentos de F(ab')2 que têm metade do tamanho dos anticorpos de comprimento total. A melhora no contraste local devido a uma menor fonte de sinal e, portanto, a resolução obtida por essa modificação em comparação com o uso de anticorpos secundários de comprimento total, permitiu a resolução dos dois C-termini de SYP-5 dentro da região central por TauSTED, que não são resolvidos por STED convencional usando anticorpos de comprimento total (16 e dados não mostrados). Prevemos que este protocolo otimizado para imagem de SCs em linhas germinativas intactas de C. eleganfacilitará a investigação da relação estrutura-função do SC durante a meiose.
Divulgações
Os autores declaram não haver conflitos de interesse.
Agradecimentos
Gostaríamos de agradecer a Jonas Ries e ao laboratório Ries por compartilhar buffers de imagem para imagens SMLM. Também agradecemos a Yumi Kim pela cepa de C. elegans usada neste protocolo e a Abby F. Dernburg pelo anticorpo frango-anti-HTP-3. Agradecemos a Marko Lampe e Stefan Terjung da Advanced Light Microscopy Facility da EMBL Heidelberg pelo seu apoio na utilização do microscópio confocal Olympus iXplore SPIN SR. Este trabalho foi apoiado pelo Laboratório Europeu de Biologia Molecular e pela Deutsche Forschungsgemeinschaft (DFG, Fundação Alemã de Investigação - 452616889, SK). Reconhecemos o acesso e os serviços prestados pelo Centro de Imagem do Laboratório Europeu de Biologia Molecular (EMBL IC), generosamente apoiado pela Fundação Boehringer Ingelheim.
Materiais
| Name | Company | Catalog Number | Comments |
| 100x/1.5 oil objective | Olympus | UPLAPO100XOHR | UPLAPO100XOHR |
| 2-mercaptoethylamine (MEA) | Sigma-Aldrich | 30070-10G | Dissolved in MilliQ water to 5 M solution, pH 8.7 adjusted with HCl. Aliquoted to a single-use volume, frozen, and kept at -80 °C. |
| Additional 640 nm booster laser | Toptica | IBEAM-SMART-640-S-HP | |
| AlexaFluor 594, NHS ester | ThermoFischer Scientific | A37572 | Dissolved in DMSO to 1 mM solution, aliquoted to single use volume, frozen and kept at -80 °C |
| AlexaFluor 647, NHS ester | ThermoFischer Scientific | A37573 | Dissolved in DMSO to 1 mM solution, aliquoted to single use volume, frozen and kept at -80 °C |
| anti-HA | Thermo Fisher Scientific | 2-2.2.14 | Mouse monoclonal, 1:250 (SMLM), 1:1,000 (STED Microscopy) |
| anti-HTP-3 | a gift from Abby F. Dernburg | MacQueen et al., 2005 | Chicken polyclonal, 1:250 (SMLM), 1:1,000 (STED Microscopy) |
| Caenorhabditis elegans strain YKM349 | a gift from Yumi Kim | Hurlock et al., 2020 | syp-5(kim9[syp-5::HA]) I; meIs8[pie-1p::GFP::cosa-1, unc-119(+)] II |
| CF6680, NHS ester | Biotium | 92139 | Dissolved in DMSO to 1mM solution, aliquoted to single use volume, frozen and kept at -80 °C |
| Circular cover glass 12 mm No. 1 | Menzel-Gläser; VWR | 631-0713 | |
| Circular cover glass 24 mm No. 1.5 | Carl Roth | PK26.1 | |
| Cylindrical lenses | Thorlabs | LJ1516RM-A, LK1002RM-A | |
| Egg Buffer (10x) | Edgar 1995 | 250 mM HEPES, 1.18 M NaCl, 480 mM KCl, 20 mM EDTA, 5 mM EGTA, pH 7.4 | |
| Ethanol (absolute for analysis) | Merck | 64-17-5 | |
| F(ab’)2 fragment anti-chicken IgY | Jackson Immunoresearch | AB_2340347 | Donkey polyclonal, 1:100 (SMLM), 1:1,000 (STED Microscopy) |
| F(ab’)2 fragment anti-mouse IgG | Jackson Immunoresearch | AB_2340761 | Donkey polyclonal, 1:100 (SMLM), 1:1,000 (STED Microscopy) |
| Fisherbrand Microscope slides T/F Ground 0.8-1.0 mm thick | Fisher scientific | 7107 | |
| Gauge Worm Pick 30 diameter 0.254 mm - Iridium 10% | Kisker | 789265 | |
| Glucose oxidase/Catalase enzyme mix (GlOX/Cat ) | a gift from Jonas Ries | Hoess, Mund, Reitberger, & Ries, 2018 | 20x, 1916 U/mL glucose oxidase (Sigma G7141), 42350 U/mL catalase (Sigma C3155), 50 mM Tris-HCl pH 8.0, 51% glycerol, MilliQ water. Stored at -20 °C. |
| Imaging buffer base | a gift from Jonas Ries | Hoess, Mund, Reitberger, & Ries, 2018 | 50 mM Tris-HCl, pH 8.0, 10 mM NaCl, 10% D-Glucose. Aliquoted to a single-use volume (950 μL), frozen, and kept at -80 °C. |
| Invitrogen ProLong Glass Antifade Mountant | ThermoFischer Scientific | P36982 | |
| Leica Stellaris 8 STED FALCON | Leica | N/A | The microscope is equiped with the latest generation white light laser, a 775nm pulsed STED laser, the FALCON Fluorescence Lifetime IMaging module, HC PL APO CS2 100x/1.40 oil objective, and Leica HyD X detector. The system is capable of FLIM module enhanced Tau-STED which measures the specific fluorescence lifetime of a dye and is therefore capable of removing background signal based on differences in fluorescence lifetimes of the dyes, and dye conditions in the sample. Additionally, the resolution is increased by accounting for the variation of fluorescence lifetimes in different areas of the depletion donut. |
| Longwave channel emission filter | AHF Analysentechnik | F47-702 | 700/100 nm bandpass |
| Methanol (absolute for analysis) | Merck | 67-56-1 | |
| NaHCO3 | Sigma-Aldrich/Merck | S5761-500G | 100 mM NaHCO3, pH 8.3 |
| Near-infrared fiber-coupled laser | Toptica | IBEAM-SMART-PT-CD | Custom Design, 808 nm - 75mW |
| Objective lens piezo mount (PIFOC ) | Physik Instrumente | P-726.1.CD | 100 µm travel range |
| Orca Fusion BT sCMOS camera | Hamamatsu | C15440-20UP | |
| PCR tubes | Greiner Bio-One | 673283 | 0.2 mL |
| Phosphate Saline Buffer (PBS 10x) | N/A | 137 mM NaCl, 2.7 mM KCl, 10 mM Na2HPO4, 1.8 mM KH2PO4, pH 7.4 | |
| Pierce 16% formaldehyde (w/v), methanol free | ThermoFischer Scientific | 28906 | 16% formaldehyde is transferred from the original glass ampule into 1.5 mL tube and kept at room temperature. |
| PlasmaPrep2 plasma cleaner | GaLa Instrumente GmbH | N/A | |
| Poly-L-lysine hydrobromide | Sigma-Aldrich/Merck | P2636-25MG | 0.1% w/v solution was prepared in Milli-Q water, and stored in aliquots at -20 °C. |
| Primary dichroic (illumination reflecting) | AHF Analysentechnik | F73-866S | quad bandpass @ 405, 488, 561, 640 |
| Quadnotch filter | AHF Analysentechnik | F40-072 | 405/488/561/640 nm |
| Quadrant photodiode (QPD) | Laser Components | SD197-23-21-041, LC301DQD-PV | |
| Razor blades | Apollo Herkenrath Solinger | N/A | |
| Refractive beam shaper | AdlOptica | PiShaper 6_6_VIS | |
| Roche Blocking Reagent | Roche | 11096176001 | 10x solution was prepared according to recommendation. Frozen aliquots were stored at -20 °C. |
| Scalpel blade (Feather brand #11, No. 3) | Heinz Herenz Medizinalbedarf GmbH | 1110911 | |
| Scalpel removal box | Fisher scientific | 10002-50 | |
| Secondary dichroic (emission reflecting) | AHF Analysentechnik | F38-785S | 750 nm longpass |
| Shortpass filter | Semrock | BSP01-785R-25 | 750 nm |
| Shortwave channel emission filter | AHF Analysentechnik | F37-677 | 676/37 nm bandpass |
| Single molecule localization microscope | EMBL Imaging Centre | Diekmann et al., 2020 with modifications | The microscope provides widefield epi-illumination via a single-mode fiber-coupled laser engine, additional booster laser, and refractive beam shaper to provide a uniform illumination field (Stehr et al, 2019). Widefield images are captured on a sCMOS camera and appropriate relay optics for a system magnification of 61x and a pixel size of 106 nm. For ratiometric imaging of spectrally overlapping far-red dyes, an image splitter produces two spectrally distinct images on the camera (splitting dichroic: 665 nm long pass, shortwave channel emission filter: 676/37 nm bandpass, longwave emission filter: 700/100 nm bandpass. An additional 405/488/561/640 nm quadnotch filter and 750 nm shortpass filter are common to the two paths and provide additional laser blocking). A compound cylindrical lens provides the astigmatism required for 3D imaging. To maintain a fixed focus across acquisitions exceeding 2 hours in time (comprising 200 000 - 250 000 images), focus locking is achieved by total internal reflection of a near-infrared fiber-coupled laser from the coverslip and subsequent height sensitive detection on a quadrant photodiode (QPD). The QPD signal provided closed-loop control of the objective lens piezo mount. For access to this microscope, refer to https://www.embl.org/about/info/imaging-centre or contact ic-contact@embl.de |
| Single-mode fiber-coupled multi-laser engine | Toptica | iCHROME MLE-LFA-HP | Provides widefield epi-illumination of 100 mW at 405, 488, 561, 640 nm |
| Splitting dichroic | AHF Analysentechnik | F48-665SG | 665 nm long pass |
| Square cover glass 22 x 22 mm No.1 | Menzel-Gläser; VWR | 630-2882 | |
| STAR 635P, NHS ester | Abberior | ST635P-0002-1MG | Dissolved in DMSO to 1 mM solution, aliquoted to single use volume, frozen and kept at -80 °C |
| Stereo microscope Stemi 305 Stand K LAB | Zeiss | N/A | |
| Tetramisole hydrochloride | Sigma-Aldrich/Merck | T1512-2G | 1% (w/v) solution was prepared in Milli-Q water. Frozen aliquots were stored at -20 °C. Thawed aliquot was kept at 4 °C and used for several months. |
| TetraSpeck Microspheres | ThermoFischer Scientific | T7279 | 0.1 µm, fluorescent blue/green/orange/dark red |
| Tris Saline Buffer (TBS 10x) | N/A | 200 mM Tris-HCl, 1.5 M NaCl, pH 7.5 | |
| TWEEN 20 | Sigma-Aldrich/Merck | P9416-50ML | Kept at room temperature in original packaging. |
| WormStuff worm pick | Kisker | 789277 | |
| XY microscope stage | Smaract | N/A | Custom Design |
| Zeba Micro Spin Desalting Column | ThermoFischer Scientific | 89877 | 7K MWCO, 75 µL |
Referências
- Zickler, D., Kleckner, N. Meiotic chromosomes: integrating structure and function. Annual Review of Genetics. 33, 603(1999).
- Ur, S. N., Corbett, K. D. Architecture and dynamics of meiotic chromosomes. Annual Review of Genetics. 55, 497-526 (2021).
- Fawcett, D. W. The fine structure ot chromosomes in the meiotic prophase of vertebrate spermatocytes. The Journal of Biophysical and Biochemical Cytology. 2 (4), 403-406 (1956).
- Moses, M. J. Chromosomal structures in crayfish spermatocytes. The Journal of Biophysical and Biochemical Cytology. 2 (2), 215-218 (1956).
- Hillers, K. J., Jantsch, V., Martinez-Perez, E., Yanowitz, J. L. Meiosis. WormBook. , 433-434 (2017).
- Pasierbek, P., et al. A Caenorhabditis elegans cohesion protein with functions in meiotic chromosome pairing and disjunction. Genes & Development. 15 (11), 1349-1360 (2001).
- Severson, A. F., Ling, L., Van Zuylen, V., Meyer, B. J. The axial element protein HTP-3 promotes cohesin loading and meiotic axis assembly in C. elegans to implement the meiotic program of chromosome segregation. Genes & Development. 23 (15), 1763-1778 (2009).
- Zetka, M. C., Kawasaki, I., Strome, S., Müller, F. Synapsis and chiasma formation in Caenorhabditis elegans require HIM-3, a meiotic chromosome core component that functions in chromosome segregation. Genes & Development. 13 (17), 2258-2270 (1999).
- Martinez-Perez, E. HTP-1-dependent constraints coordinate homolog pairing and synapsis and promote chiasma formation during C. elegans meiosis. Genes & Development. 19 (22), 2727-2743 (2005).
- Couteau, F., Zetka, M. HTP-1 coordinates synaptonemal complex assembly with homolog alignment during meiosis in C. elegans. Genes & Development. 19 (22), 2744-2756 (2005).
- Goodyer, W., et al. HTP-3 Links DSB Formation with Homolog Pairing and Crossing Over during C. elegans Meiosis. Developmental Cell. 14 (2), 263-274 (2008).
- Colaiácovo, M. P., et al. Synaptonemal complex assembly in C. elegans is dispensable for loading strand-exchange proteins but critical for proper completion of recombination. Developmental Cell. 5 (3), 463-474 (2003).
- MacQueen, A. J., Colaiácovo, M. P., McDonald, K., Villeneuve, A. M. Synapsis-dependent and -independent mechanisms stabilize homolog pairing during meiotic prophase in C. elegans. Genes & Development. 16 (18), 2428-2442 (2002).
- Smolikov, S., et al. Synapsis-defective mutants reveal a correlation between chromosome conformation and the mode of double-strand break repair during Caenorhabditis elegans meiosis. Genetics. 176 (4), 2027-2033 (2007).
- Smolikov, S., Schild-Prüfert, K., Colaiácovo, M. P. A yeast two-hybrid screen for SYP-3 interactors identifies SYP-4, a component required for synaptonemal complex assembly and chiasma formation in Caenorhabditis elegans meiosis. PLoS Genetics. 5 (10), 1000669(2009).
- Hurlock, M. E., et al. Identification of novel synaptonemal complex components in C. Elegants. The Journal of Cell Biology. 219 (5), (2020).
- Zhang, Z., et al. Multivalent weak interactions between assembly units drive synaptonemal complex formation. The Journal of Cell Biology. 219 (5), (2020).
- Schild-Prüfert, K., et al. Organization of the synaptonemal complex during meiosis in Caenorhabditis elegans. Genetics. 189 (2), 411-421 (2011).
- Köhler, S., Wojcik, M., Xu, K., Dernburg, A. F. The interaction of crossover formation and the dynamic architecture of the synaptonemal complex during meiosis. bioRxiv. , (2020).
- Chen, F., Tillberg, P. W., Boyden, E. S. Expansion microscopy. Science. 347 (6621), 543-548 (2015).
- Klar, T. A., Jakobs, S., Dyba, M., Egner, A., Hell, S. W. Fluorescence microscopy with diffraction resolution barrier broken by stimulated emission. Proceedings of the National Academy of Sciences. 97 (15), 8206-8210 (2000).
- Rust, M. J., Bates, M., Zhuang, X. Sub-diffraction-limit imaging by stochastic optical reconstruction microscopy (STORM). Nature Methods. 3 (10), 793-796 (2006).
- Betzig, E., et al. Imaging intracellular fluorescent proteins at nanometer resolution. Science. 313 (5793), 1642-1645 (2006).
- Schücker, K., Holm, T., Franke, C., Sauer, M., Benavente, R. Elucidation of synaptonemal complex organization by super-resolution imaging with isotropic resolution. Proceedings of the National Academy of Sciences. 112 (7), 2029-2033 (2015).
- Cahoon, C. K., et al. Superresolution expansion microscopy reveals the three-dimensional organization of the Drosophila synaptonemal complex. Proceedings of the National Academy of Sciences. 114 (33), 6857-6866 (2017).
- Zwettler, F. U., et al. Tracking down the molecular architecture of the synaptonemal complex by expansion microscopy. Nature Communications. 11 (1), 1-11 (2020).
- Yoon, S., Choi, E. H., Kim, J. W., Kim, K. P. Structured illumination microscopy imaging reveals localization of replication protein A between chromosome lateral elements during mammalian meiosis. Experimental & Molecular Medicine. 50 (8), 1-12 (2018).
- Prakash, K., et al. Superresolution imaging reveals structurally distinct periodic patterns of chromatin along pachytene chromosomes. Proceedings of the National Academy of Sciences. 112 (47), 14635-14640 (2015).
- Xu, H., et al. Molecular organization of mammalian meiotic chromosome axis revealed by expansion STORM microscopy. Proceedings of the National Academy of Sciences. 116 (37), 18423-18428 (2019).
- Köhler, S., Wojcik, M., Xu, K., Dernburg, A. F. Superresolution microscopy reveals the three-dimensional organization of meiotic chromosome axes in intact Caenorhabditis elegans tissue. Proceedings of the National Academy of Sciences. 114 (24), 4734-4743 (2017).
- Hell, S. W. Far-field optical nanoscopy. Science. 316 (5828), 1153-1158 (2007).
- Hein, B., Willig, K. I., Hell, S. W. Stimulated emission depletion (STED) nanoscopy of a fluorescent protein-labeled organelle inside a living cell. Proceedings of the National Academy of Sciences. 105 (38), 14271-14276 (2008).
- Jahr, W., Velicky, P., Danzl, J. G. Strategies to maximize performance in STimulated Emission Depletion (STED) nanoscopy of biological specimens. Methods. 174, 27-41 (2019).
- Thevathasan, J. V., et al. Nuclear pores as versatile reference standards for quantitative superresolution microscopy. Nature Methods. 16 (10), 1045-1053 (2019).
- Xu, K., Shim, S. -H., Zhuang, X. Super-resolution imaging through stochastic switching and localization of single molecules: an overview. Far-Field Optical Nanoscopy. , 27-64 (2013).
- Wang, Y., et al. Localization events-based sample drift correction for localization microscopy with redundant cross-correlation algorithm. Optics Express. 22 (13), 15982(2014).
- Winterflood, C. M., Platonova, E., Albrecht, D., Ewers, H. Dual-color 3D superresolution microscopy by combined spectral-demixing and biplane imaging. Biophysical Journal. 109 (1), 3-6 (2015).
- Diekmann, R., et al. Optimizing imaging speed and excitation intensity for single molecule localization microscopy. Nature Methods. 17 (9), 909(2020).
- Huang, B., Wang, W., Bates, M., Zhuang, X. Three-dimensional super-resolution imaging by stochastic optical reconstruction microscopy. Science. 319 (5864), 810-813 (2008).
- Phillips, C. M., McDonald, K. L., Dernburg, A. F. Cytological analysis of meiosis in Caenorhabditis elegans. Methods in Molecular Biology. 558, 171-195 (2009).
- Edgar, L. G. Blastomere culture and analysis. Methods in Cell Biology. 48, 303-321 (1995).
- MacQueen, A. J., et al. Chromosome sites play dual roles to establish homologous synapsis during meiosis in C. elegans. Cell. 123 (6), 1037-1050 (2005).
- Stehr, F., Stein, J., Schueder, F., Schwille, P., Jungmann, R. Flat-top TIRF illumination boosts DNA-PAINT imaging and quantification. Nature Communications. 10 (1), 1-8 (2019).
- Hoess, P., Mund, M., Reitberger, M., Ries, J. Dual-color and 3D super-resolution microscopy of multi-protein assemblies. Methods in Molecular Biology. 1764, 237-251 (2018).
- Edelstein, A., Amodaj, N., Hoover, K., Vale, R., Stuurman, N. Computer Control of microscopes using µManager. Current Protocols in Molecular Biology. 92 (1), 14-20 (2010).
- Edelstein, A. D., et al. Advanced methods of microscope control using µManager software. Journal of Biological Methods. 1 (2), 10(2014).
- Deschamps, J., Ries, J. EMU: reconfigurable graphical user interfaces for Micro-Manager. BMC Bioinformatics. 21 (1), 1-13 (2020).
- Ries, J. SMAP: a modular super-resolution microscopy analysis platform for SMLM data. Nature Methods. 17 (9), 870-872 (2020).
- Li, Y., et al. Global fitting for high-accuracy multi-channel single-molecule localization. Nature Communications. 13 (1), 1-11 (2022).
- Nieuwenhuizen, R. P. J., et al. Measuring image resolution in optical nanoscopy. Nature Methods. 10 (6), 557-562 (2013).
- Banterle, N., Bui, K. H., Lemke, E. A., Beck, M. Fourier ring correlation as a resolution criterion for super-resolution microscopy. Journal of Structural Biology. 183 (3), 363-367 (2013).
- Preibisch, S., Saalfeld, S., Tomancak, P. Globally optimal stitching of tiled 3D microscopic image acquisitions. Bioinformatics. 25 (11), 1463-1465 (2009).
- Shi, X., Lim, J., Ha, T. Acidification of the oxygen scavenging system in single-molecule fluorescence studies: in situ sensing with a ratiometric dual-emission probe. Analytical Chemistry. 82 (14), 6132-6138 (2010).
- Olivier, N., Keller, D., Rajan, V. S., Gönczy, P., Manley, S. Simple buffers for 3D STORM microscopy. Biomedical Optics Express. 4 (6), 885-899 (2013).
- Mund, M., et al. Superresolution microscopy reveals partial preassembly and subsequent bending of the clathrin coat during endocytosis. bioRxiv. , (2022).
- Mund, M., et al. Systematic nanoscale analysis of endocytosis links efficient vesicle formation to patterned actin nucleation. Cell. 174 (4), 884-896 (2018).
- Sabinina, V. J., et al. Three-dimensional superresolution fluorescence microscopy maps the variable molecular architecture of the nuclear pore complex. Molecular Biology of the Cell. 32 (17), 1523-1533 (2021).
- Cieslinski, K., et al. Nanoscale structural organization and stoichiometry of the budding yeast kinetochore. bioRxiv. , (2021).
- Sieben, C., Banterle, N., Douglass, K. M., Gönczy, P., Manley, S. Multicolor single-particle reconstruction of protein complexes. Nature Methods. 15 (10), 777-780 (2018).
- Sarkans, U., et al. The BioStudies database—one stop shop for all data supporting a life sciences study. Nucleic Acids Research. 46, (2018).
Reimpressões e Permissões
Solicitar permissão para reutilizar o texto ou figuras deste artigo JoVE
Solicitar PermissãoThis article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. Todos os direitos reservados